Introduction
Although improvement of systemic chemotherapy has
been demonstrated in gastrointestinal (GI) tract cancer including
gastric and colon cancer, the overall survival rate in patients
with advanced stage is still poor (1). Therefore, it would be desirable to
develop molecular target therapy more efficiently in GI-tract
cancer. For example, for the human epidermal growth factor receptor
(EGFR) related 2 (HER2)-overexpressing gastric cancer, Trastuzumab
in combination with chemotherapy vs. chemotherapy alone for
treatment of HER2-positive advanced gastric cancer (ToGA) trial
concluded that anti-HER2 monoclonal antibody (trastuzumab) plus
chemotherapy is a standard treatment option, in which trastuzumab
plus chemotherapy showed better survival in comparison to
chemotherapy alone (2).
It is generally accepted that trastuzumab can act on
gastric cancer cells through both anti-proliferative function
directly to cancer cells and antibody-dependent cellular
cytotoxicity (ADCC) activity via immune cells (3,4). It has
been reported that Trastuzumab-mediated ADCC can be influenced by
several factors including single nucleotide polymorphisms
(SNPs) in the Fc gamma receptor (FcγR) genes (5–7) or natural
killer (NK) cell dysfunction (8). In
fact, the SNPs can alter the FcγR binding affinity to the
therapeutic monoclonal antibodies (mAbs) and consequently resulted
in impairment of the ADCC activity. Of importance, a clinical trial
showed that therapeutic efficacy of trastuzumab against
HER2-positive breast cancer was significantly different between
patients with and without certain SNPs in the FcγR genes (9). Furthermore, the same observation was
also confirmed in colorectal cancer treated with anti-EGFR
antibody, cetuximab (10). These
results strongly suggest that enhancement of ADCC with some
modalities would be a promising approach to enhance the efficacy of
therapeutic mAbs.
It has been shown that removal of fucose from
antibody oligosaccharides attached to Asn297 of the
heavy chain (defucosylation) significantly enhanced FcγR binding
affinity between FcγR on NK cells and the mAbs, in comparison to
that of conventional antibody, leading to augmentation of ADCC
activity (11–15). Thus, the defucosylation technology
could be one of the most powerful approaches to enhance clinical
efficacy of therapeutic mAbs. There is, however, still limited
information describing the clinical usefulness of the defucosylated
therapeutic antibody on the ADCC, except for one report showing
that the use of the defucosylated antibodies may improve the
therapeutic effects of trastuzumab for breast cancer patients
(16). Thus, it is necessary to draw
solid conclusion for the effectiveness of the defucosylated
antibody in cancer patients or immunosuppressive state. In the
present study, using PBMCs from GI tract cancer patients and
healthy donors, we evaluated trastuzumab- and cetuximab-mediated
ADCC by comparing the defucosylated mAbs with conventional mAbs.
This is the first report using PBMCs from patients of GI tract
cancer. In addition, when ADCC-related molecules are modulated by
mitogen-activated protein kinase (MAPK) inhibitors, the
trastuzumab- and cetuximab-mediated ADCC were also evaluated.
Materials and methods
Preparation of human effector
cells
Twenty patients with histologically diagnosed GI
tract cancer, who were treated at Fukushima Medical University
Hospital (Fukushima, Japan) from February to August in 2016, were
enrolled. PBMCs were isolated from esophageal (n=4), gastric (n=9),
and colon cancer patients (n=7), and healthy individuals (n=10,
34.8±7.8 years old, Male: Female=9:1). PBMCs were separated by
lymphocyte separation solution (Lymphoprep™, Cosmo Bio Company)
with density gradient. None of the patients received radiotherapy,
chemotherapy, surgery, or other medical interventions before this
study. Patients' characteristics are shown in Table I. This study was approved by the
ethical committee of Fukushima Medical University (approval no.
2353), and informed consent for blood donations was obtained for
all individuals.
 | Table I.Characteristics of the patients
(n=20). |
Table I.
Characteristics of the patients
(n=20).
| Characteristic | Number of patients
(n) |
|---|
| Age, years (median,
range) | 54–80 (65) |
| Male:female | 17:3 |
| Location of
carcinoma |
|
|
Esophagus | 4 |
|
Stomach | 9 |
|
Colon | 7 |
| Clinical stage (TNM
classification) |
|
| 0 | 2 |
| 1 | 6 |
| 2 | 2 |
| 3 | 8 |
| 4 | 2 |
Cell lines
MKN-7 (HER-2 overexpressing gastric cancer cell
lines; cat. no. JCRB1025) and K562 (myelogenous leukemia cell
lines; cat. no. JCRB0019) were purchased from the Japanese
Collection of Research Bioresources (Osaka, Japan). MKN-28 (EGFR
overexpressing gastric cancer cell line) was obtained from the
American Type Culture Collection (Rockville, MD, USA). The MKN28
cell line has previously been reported to be a mixed gastric cancer
type, with MKN74 (an EGFR overexpressing cancer cell line)
contamination (17). However, this
contamination is not thought to have affected the results of the
present study as MKN28 and MKN74 share similar characteristics in
terms of EGFR overexpression, as described previously (18). All cell lines were maintained in
RPMI-1640 (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) with 10%
fetal bovine serum (Nichirei Biosciences, Inc., Tokyo, Japan) and
1% penicillin/streptomycin (Nichirei Biosciences, Inc.) at 37°C and
5% CO2.
Antibodies
Anti-HER-2 monoclonal antibody trastuzumab and
anti-human EGFR antibody cetuximab were used as clinical grade
products, and their defucosylated version were provided by Kyowa
Hakko Kirin Co., Ltd. (Tokyo, Japan), which were designed according
to the known amino acid sequences (19,20) and
produced with parent or α-1,6-fucosyltransferase knockout Chinese
hamster ovary cells (21).
Antibody-dependent cellular
cytotoxicity (ADCC) assay
Cytotoxicity was determined by the lactate
dehydrogenase (LDH) release assay using PBMCs as effector cells and
either MKN-7 cells or MKN-28 cells as target cells. Briefly, target
cells (5×103 per well) were distributed into 96-well
U-bottomed plates and pre-incubated with mAbs for 1.5 h at 37°C, 5%
CO2. Then, effector cells were added at indicated doses
and incubated for 7 h. Assays were performed in triplicate with or
without antibodies. The supernatant LDH activity was measured using
a nonradioactive cytotoxicity assay kit (Cytotoxicity Detection
kitPLUS; Roche Diagnostics, Basel, Switzerland) and was
measured at 490 nm excitation and 650 nm emission wavelengths using
spectrometer. Percentage cytotoxicity was calculated according to
the formula: Cytotoxicity (%)=100 × (Experimental
release-Spontaneous release)/(Maximum release-Spontaneous release).
The maximum release was prepared with target cells lysed with the
lysis solution. In the several pre-test run, the spontaneous
release of effector cells was approximately zero. Net ADCC was
calculated according to the formula: net ADCC (%)=ADCC activities
(%)-antibody-independent cellular cytotoxicity (AICC; %), where
AICC is the nonspecific cytotoxicity in the absence of
antibodies.
Cell treatment with MAPK signal
inhibitors
Tumor cells were cultured in a 6-well plate and
exposed to the MAPK signal inhibitor, PD0325901 (Selleck Chemicals,
Houston, TX, USA) as indicated in our previous report (22). Then, cytotoxicity assays were
performed after 48 h of incubation.
Statistical analysis
Date comparing differences between two groups
assessed using unpaired Student's t-test and two way ANOVA.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Optimal condition of defucosylated
therapeutic mAbs for ADCC activity
Cetuximab-mediated and trastuzumab-mediated ADCCs
were evaluated in various concentrations of conventional and
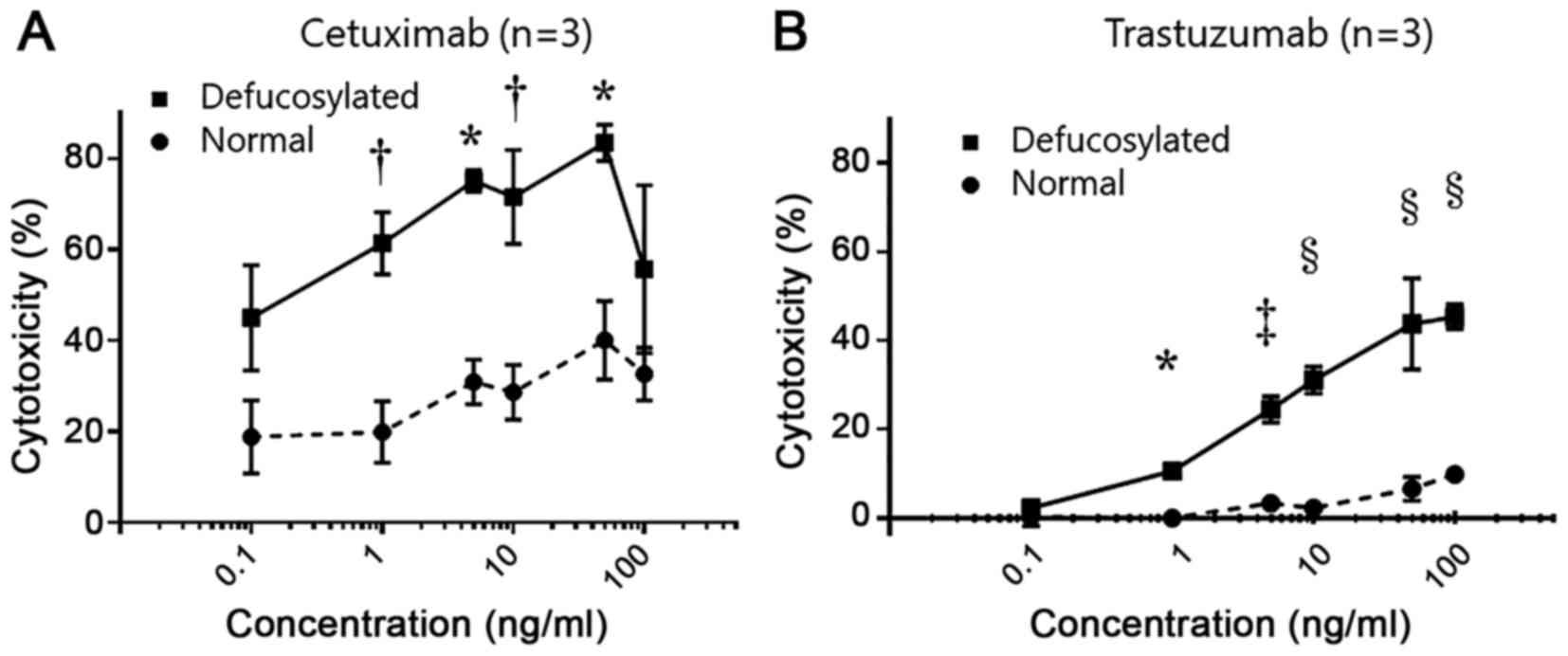
defucosylated mAbs using healthy donor's PBMCs (n=3, Fig. 1A and B). The EGFR-positive MKN28
gastric cancer cell line was used for cetuximab-mediated ADCC and
the HER2-positive MKN7 gastric cancer was used for
trastuzumab-mediated ADCCs, and the overexpression of EGFR or HER2
on tumor cells were repeatedly confirmed by flow cytometry (data
not shown).
As shown in Fig. 1A,
defucosylated cetuximab-mediated ADCCs at effector: Target ratio of
40:1 were significantly higher than conventional cetuximab-mediated
ADCCs in each concentration. We observed a dose-dependent increase
from 0.1 to 50 ng/ml and thereafter the ADCC leaded to a drop in
the present experimental condition, consistent with the previous
report (21). Therefore, 50 ng/ml of
defucosylated and conventional cetuximab were used for subsequent
experiments as optimal doses.
Also, the same tendency was observed in
trastuzumab-mediated ADCCs (Fig. 1B)
and 50 ng/ml of defucosylated and conventional trastuzumab were
used for subsequent experiments as optimal doses.
Augmented ADCC by defucosylated
cetuximab and trastuzumab
ADCC activities mediated by either conventional or
defucosylated mAbs using PBMCs from cancer patients (n=20) and
healthy volunteers (n=10) were evaluated. The patient's background
is shown in Table I. In order to
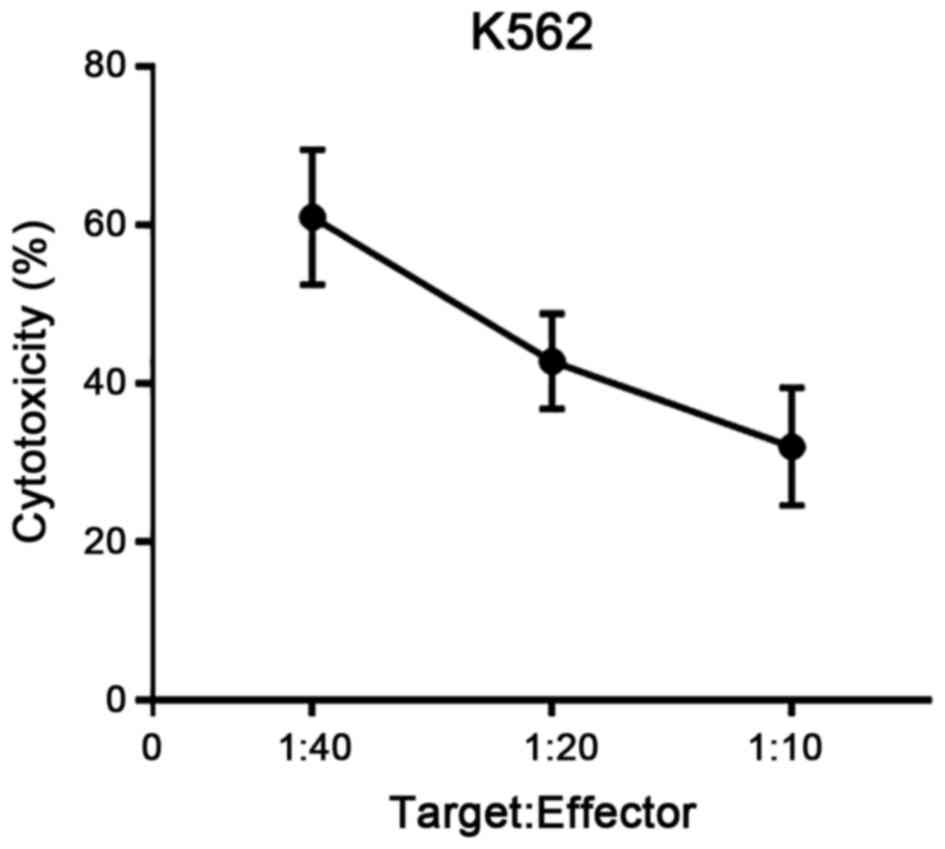
confirm the condition of PBMCs as effector cells, NK cell
activities targeted for K562 were also evaluated in parallel to the
ADCC assay and we confirmed condition of NK status in each
experiment (Fig. 2).
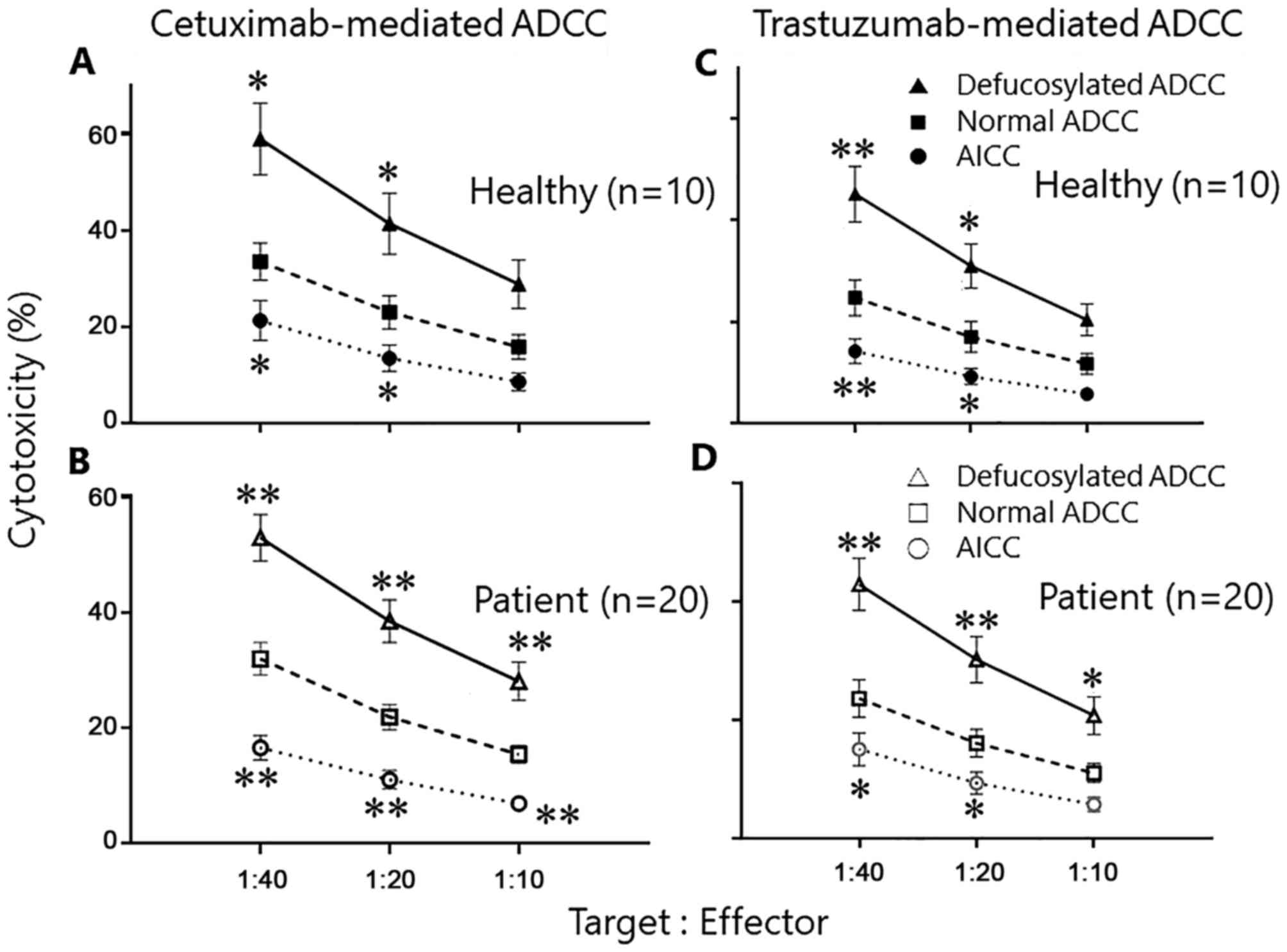
As shown in Fig. 3A,
the defucosylated cetuximab-mediated ADCCs were markedly enhanced
in comparison to conventional cetuximab-mediated ADCCs in healthy
donor's PBMCs. For example, the defucosylated and conventional
cetuximab-mediated ADCC at 40:1 ratio were 58.9±7.5 and 33.5±3.9%,
respectively. Similar observation was also confirmed using the
PBMCs from cancer patients (Fig. 3B),
in which the defucosylated and conventional trastuzumab-mediated
ADCC at 40:1 ratio were 52.9±4.0 and 32.0±2.8%, respectively.
Also, the enhancement by defucosylated mAbs was
confirmed in the trastuzumab-mediated ADCCs in both healthy donors
and cancer patients (Fig. 3C and
D).
Taken together, the defucosylated therapeutic mAbs
can enhance the ADCC activities in comparison to the conventional
mAbs using PBMCs from both healthy donors and cancer patients.
Defucosylated cetuximab- and
trastuzumab-mediated ADCC in advanced cancer cases
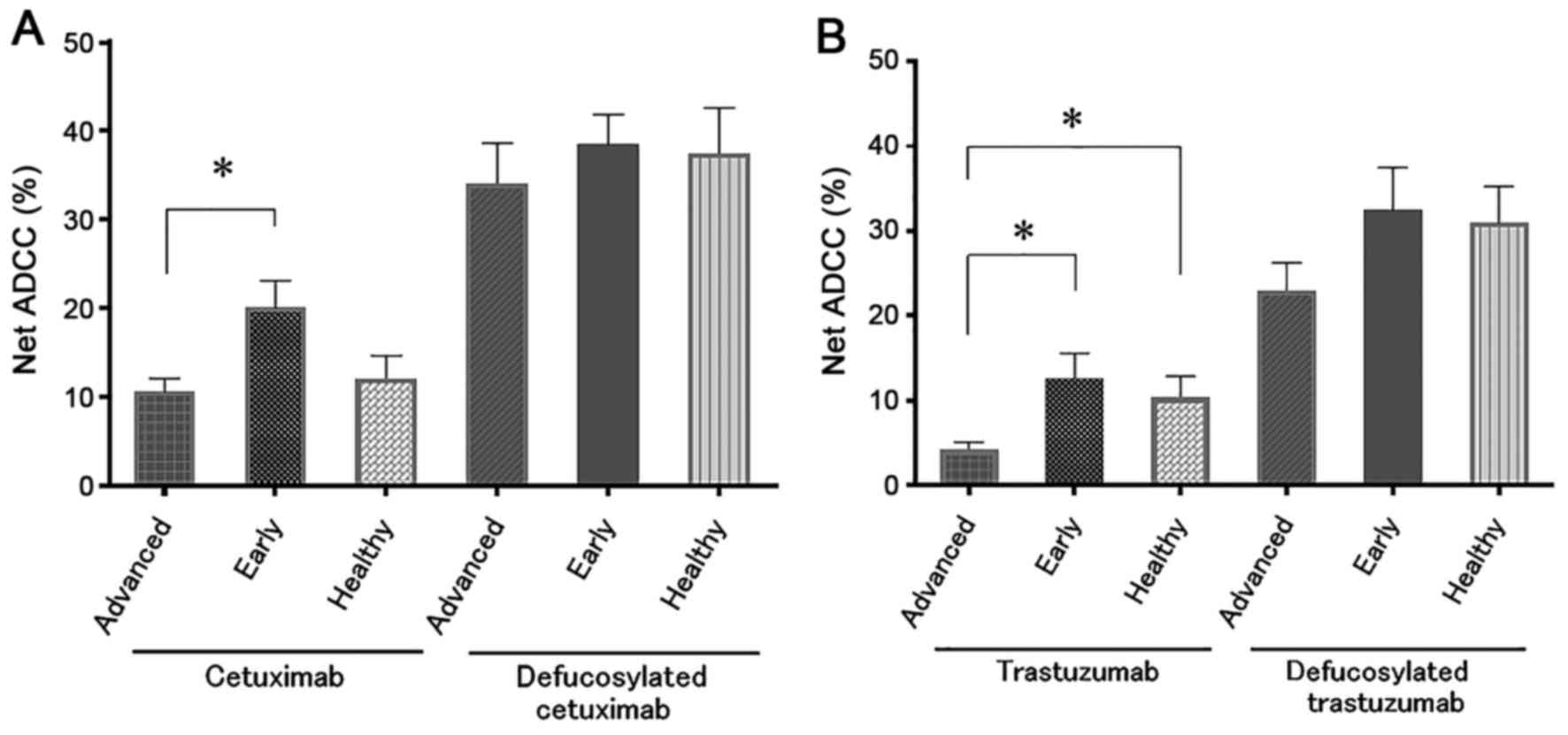
Based on the UICC-TNM classification, we classified
the cancer patients into advanced disease corresponded to stage III
and IV, or into early disease corresponded to stage 0, I, and II.
It has been already reported that the ADCC activities in advanced
cancer patients were impaired due to several mechanisms, including
NK cell dysfunction or immunosuppressive factors (8,23,24). As expected, we confirmed that the
cetuximab-mediated and trastuzumab-mediated ADCCs in advanced
disease were impaired in comparison to those in early disease or
healthy individuals (Fig. 4).
However, when the defucosylated mAbs were used instead of the
conventional mAbs, the ADCC activities in the advanced cases were
almost comparable to those in early disease or healthy individuals
(Fig. 4) and this observation was
confirmed in both defucosylated cetuximab and trastuzumab.
Thus, the defucosylated therapeutic mAbs can rescue
the impaired ADCC in advanced disease.
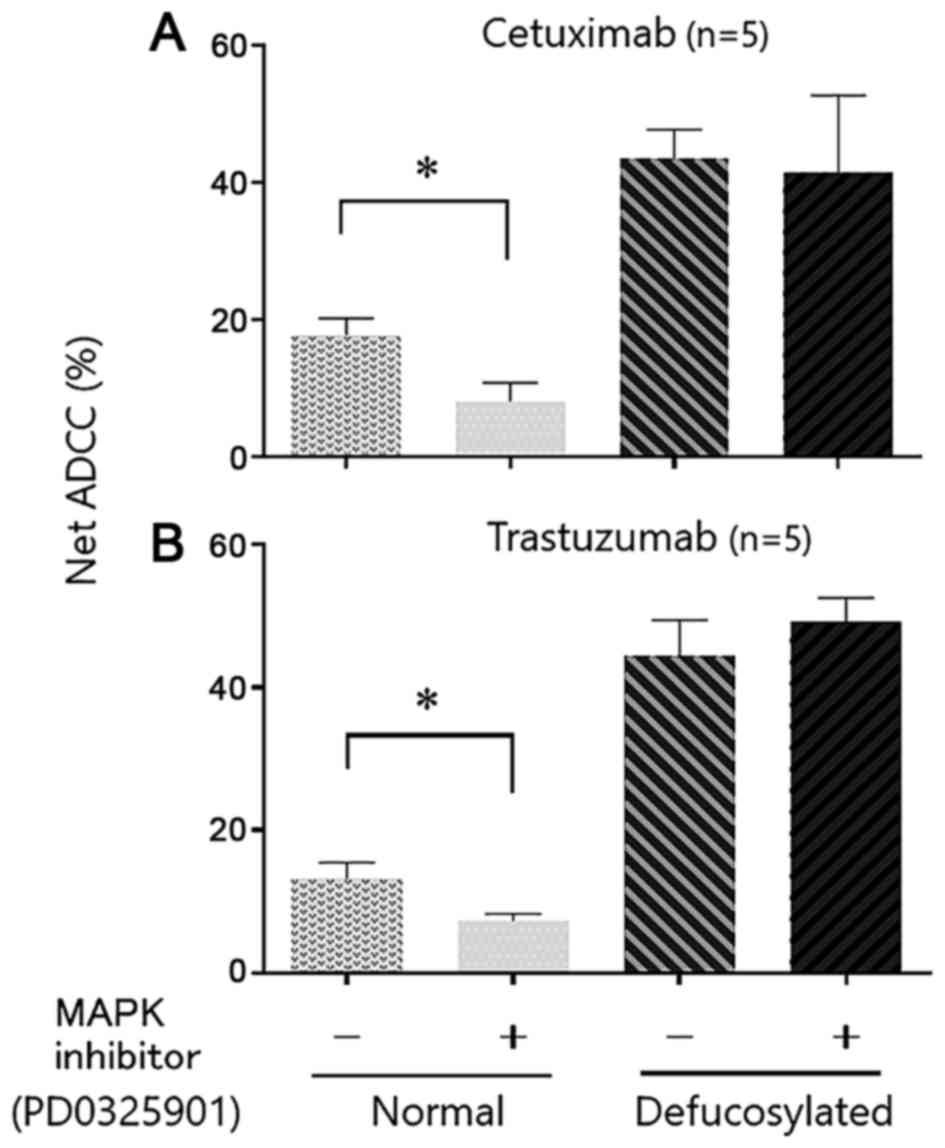
ADCC by defucosylated cetuximab and
trastuzumab when treated with MAPK inhibitors
In order to further investigate defucosylated
therapeutic mAbs-mediated ADCC, we modified the expression of
natural killer group 2 member D receptor (NKG2D) ligand and major
histocompatibility complex (MHC) class I on tumor cells by MAPK
inhibitors as indicated in our previous report (22). It is generally accepted that NK cells
can react with tumor cells through the balance of inhibitory and
stimulatory signals. The interaction between the killer
immunoglobulin-like receptor family on NK cells and MHC class I
molecules results in inhibitory signals, whereas activating signals
by NKG2D ligands expressed on targets induce stimulatory signals
leading to target cell killing (25–27). We
have shown that treatment of target tumor cells with MAPK
inhibitors can decrease ADCC activities through upregulation of MHC
class I and down-regulation of NKG2D ligands such as MICA/B
(22). As expected, conventional
cetuximab- and trastuzumab-mediated ADCC was significantly
decreased, when target tumor cells were pre-treated with the MAPK
inhibitor (Fig. 5). However, when the
defucosylated mAbs were used instead, ADCC activities did not alter
even if the target cells were pre-treated with the MAPK inhibitor.
In the current experiment condition, MAPK inhibitors do not have
any direct anti-proliferative effect on target cancer cells (data
not shown).
Taken together, defucosylated therapeutic mAbs can
efficiently enhance ADCC activities, even if the NKG2D ligand and
MHC class I expression on tumor cells, which are corresponding to
immune suppressive status, were modified.
Discussion
The present study provide an important finding
relevant to clinical cancer treatment with therapeutic mAbs. First,
we showed the augmentation of ADCC by defucosylated therapeutic
mAbs using PBMCs from healthy donors and cancer patients. Second,
although the ADCC activities were impaired in advanced disease, the
defucosylated mAbs can restore the impaired ADCC to the levels of
healthy individuals. Finally, the defucosylated therapeutic mAbs
can enhance ADCC activities even if the NKG2D ligand and MHC class
I expression on tumor cells were modified to induce
immunosuppressive environment.
There is accumulating evidence that ADCC is an
important antitumor mechanism when the therapeutic mAbs showed the
clinical benefit (28–32), and augmentation of ADCC will be able
to enhance the clinical efficacy of the therapeutic mAbs. For
example, modification of antibodies to increase binding to FcγR has
been pursued in order to augment ADCC (11,12). It
has finally been reported that removal of the α-1,6 fucose moiety
on the N-glycan at Asn297 of the heavy chain, which is
called defucosylation technology, significantly enhanced ADCC in
comparison to that of conventional antibody. Previously, there is
only one report describing the efficacy of defucosylated
trastuzumab using PBMCs from breast cancer patients (16). Herein, our present study is the first
report indicating usefulness of defucosylated cetuximab and
trastuzumab for ADCC using the PBMCs of GI-tract cancer patients.
Our observation and the previous report using clinical samples
clearly confirmed the defucosylation technology efficiently can
enhance the therapeutic mAbs-mediated ADCC.
In line with several previous reports (23,33), we
confirmed that conventional therapeutic mAbs-mediated ADCC in
advanced disease was impaired in comparison to those in early
disease or healthy donors. It is generally accepted that NK cells
in cancer-bearing hosts are impaired by many mechanisms, including
their reduced number, imbalances in their activating and inhibitory
receptor, impaired activation signaling cascade as well as
immunosuppressive cytokines (8,23,24). However, even in such a condition, the
current study clearly indicated that the defucosylated mAbs can
restore the impaired ADCC in advanced diseases.
MAPK inhibitors have been originally developed for
anti-cancer drugs based on their anti-proliferative action against
tumor cells (34,35). In addition, we and others previously
reported that the MAPK inhibitor can induce up-regulation of HLA
Class I and down-regulation of MICA/B expression on tumor cells,
leading to less NK sensitivity (26,27).
Therefore, in the present study, we tried to mimic
immunosuppressive status specific for NK-killing by modulating
ADCC-related molecules with MAPK inhibitors. As a result, we
confirmed that conventional cetuximab- and trastuzumab-mediated
ADCC was impaired when target tumor cells were pre-treated with the
MAPK inhibitor. Of importance, the defucosylated therapeutic mAbs
can enhance ADCC activities even if the NKG2D ligand and MHC class
I expression on tumor cells were modified by the MAPK inhibitor
into immunosuppressive status. Taking it into consideration, the
defucosylated therapeutic mAbs have a strong capability to enhance
ADCC activities.
Theoretically, it would be likely that the
defucosylated therapeutic mAbs has no ADCC activity against normal
tissues, since the therapeutic potential of the mAbs are dependent
on the level of target antigens expressed on the surface of target
cells. When we consider the clinical application of the
defucosylated therapeutic mAbs described in the present study,
further study to exclude potential toxicities of the defucosylated
mAbs to tissues expressing low levels of such target antigens will
be needed.
In conclusion, the defucosylated therapeutic mAbs
can restore the impaired ADCC activities in advanced stage of
cancer patients, leading to more effective anti-cancer
treatments.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bang YJ, Van Cutsem E, Feyereislova A,
Chung HC, Shen L, Sawaki A, Lordick F, Ohtsu A, Omuro Y, Satoh T,
et al: Trastuzumab in combination with chemotherapy versus
chemotherapy alone for treatment of HER2-positive advanced gastric
or gastro-oesophageal junction cancer (ToGA): A phase 3,
open-label, randomised controlled trial. Lancet. 376:687–697. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li K and Li J: Current molecular targeted
therapy in advanced gastric cancer: A comprehensive review of
therapeutic mechanism, clinical trials, and practical application.
Gastroenterol Res Pract. 2016:41056152016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arienti C, Zanoni M, Pignatta S, Del Rio
A, Carloni S, Tebaldi M, Tedaldi G and Tesei A: Preclinical
evidence of multiple mechanisms underlying trastuzumab resistance
in gastric cancer. Oncotarget. 7:18424–18439. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koene HR, Kleijer M, Algra J, Roos D, von
dem Borne AE and de Haas M: Fc gammaRIIIa-158V/F polymorphism
influences the binding of IgG by natural killer cell Fc gammaRIIIa,
independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood.
90:1109–1114. 1997.PubMed/NCBI
|
|
6
|
Shields RL, Namenuk AK, Hong K, Meng YG,
Rae J, Briggs J, Xie D, Lai J, Stadlen A, Li B, et al: High
resolution mapping of the binding site on human IgG1 for Fc gamma
RI Fc gamma RII Fc gamma RIII, and FcRn and design of IgG1 variants
with improved binding to the Fc gamma R. J Biol Chem.
276:6591–6604. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Warmerdam PA, van de Winkel JG, Vlug A,
Westerdaal NA and Capel PJ: A single amino acid in the second
Ig-like domain of the human Fc gamma receptor II is critical for
human IgG2 binding. J Immunol. 147:1338–1343. 1991.PubMed/NCBI
|
|
8
|
Watanabe M, Kono K, Kawaguchi Y, Mizukami
Y, Mimura K, Maruyama T, Izawa S and Fujii H: NK cell dysfunction
with down-regulated CD16 and up-regulated CD56 molecules in
patients with esophageal squamous cell carcinoma. Dis Esophagus.
23:675–681. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Musolino A, Naldi N, Bortesi B, Pezzuolo
D, Capelletti M, Missale G, Laccabue D, Zerbini A, Camisa R,
Bisagni G, et al: Immunoglobulin G fragment C receptor
polymorphisms and clinical efficacy of trastuzumab-based therapy in
patients with HER-2/neu-positive metastatic breast cancer. J Clin
Oncol. 26:1789–1796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wormald S, Milla L and O'Connor L:
Association of candidate single nucleotide polymorphisms with
somatic mutation of the epidermal growth factor receptor pathway.
BMC Med Genomics. 6:432013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shields RL, Lai J, Keck R, O'Connell LY,
Hong K, Meng YG, Weikert SH and Presta LG: Lack of fucose on human
IgG1 N-linked oligosaccharide improves binding to human Fcgamma
RIII and antibody-dependent cellular toxicity. J Biol Chem.
277:26733–26740. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shinkawa T, Nakamura K, Yamane N,
Shoji-Hosaka E, Kanda Y, Sakukrada M, Uchida K, Anazawa H, Satoh M,
Yamasaki M, et al: The absence of fucose but not the presence of
galactose or bisecting N-acetylglucosamine of human IgG1
complex-type oligosaccharides shows the critical role of enhancing
antibody-dependent cellular cytotoxicity. J Biol Chem.
278:3466–3473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Niwa R, Shoji-Hosaka E, Sakurada M,
Shinkawa T, Uchida K, Nakamura K, Matsushima K, Ueda R, Hanai N and
Shitara K: Defucosylated chimeric anti-CC chemokine receptor 4 IgG1
with enhanced antibody-dependent cellular cytotoxicity shows potent
therapeutic activity to T-cell leukemia and lymphoma. Cancer Res.
64:2127–2133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Okazaki A, Shoji-Hosaka E, Nakamura K,
Wakitani M, Uchida K, Kakita S, Tsumoto K, Kumagai I and Shitara K:
Fucose depletion from human IgG1 oligosaccharide enhances binding
enthalpy and association rate between IgG1 and FcgammaRIIIa. J Mol
Biol. 336:1239–1249. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Niwa R, Hatanaka S, Shoji-Hosaka E,
Sakurada M, Kobayashi Y, Uehara A, Yokoi H, Nakamura K and Shitara
K: Enhancement of the antibody-dependent cellular cytotoxicity of
low-fucose IgG1 Is independent of FcgammaRIIIa functional
polymorphism. Clin Cancer Res. 10:6248–6255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki E, Niwa R, Saji S, Muta M, Hirose
M, Iida S, Shiotsu Y, Satoh M, Shitara K, Kondo M and Toi M: A
nonfucosylated anti-HER2 antibody augments antibody-dependent
cellular cytotoxicity in breast cancer patients. Clin Cancer Res.
13:1875–1882. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR and Freshney RI: Check your cultures! A list of
cross-contaminated or misidentified cell lines. Int J Cancer.
127:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shiraishi K, Mimura K, Izawa S, Inoue A,
Shiba S, Maruyama T, Watanabe M, Kawaguchi Y, Inoue M, Fujii H and
Kono K: Lapatinib acts on gastric cancer through both
antiproliferative function and augmentation of trastuzumab-mediated
antibody-dependent cellular cytotoxicity. Gastric Cancer.
16:571–580. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carter P, Presta L, Gorman CM, Ridgway JB,
Henner D, Wong WL, Rowland AM, Kotts C, Carverr ME and Shepard HM:
Humanization of an anti-p185HER2 antibody for human cancer therapy.
Proc Natl Acad Sci USA. 89:pp. 4285–4289. 1992; View Article : Google Scholar : PubMed/NCBI
|
|
20
|
DrugBANK. https://www.drugbank.ca/drugs/DB00002
|
|
21
|
Yamane-Ohnuki N, KInoshita S,
Inoue-Urakubo M, Kusunoki M, Iida S, Nakano R, Wakitani M, Niwa R,
Sakurada M, Uchida K, et al: Establishment of FUT8 knockout Chinese
hamster ovary cells: An ideal host cell line for producing
completely defucosylated antibodies with enhanced
antibody-dependent cellular cytotoxicity. Biotechnol Bioeng.
87:614–622. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mimura K, Kamiya T, Shiraishi K, Kua LF,
Shabbir A, So J, Yong WP, Suzuki Y, Yoshimoto Y, Nakano T, et al:
Therapeutic potential of highly cytotoxic natural killer cells for
gastric cancer. Int J Cancer. 135:1390–1398. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kono K, Takahashi A, Ichihara F, Sugai H,
Fujii H and Matsumoto Y: Impaired antibody-dependent cellular
cytotoxicity mediated by herceptin in patients with gastric cancer.
Cancer Res. 62:5813–5817. 2002.PubMed/NCBI
|
|
24
|
Mimura K, Kono K, Hanawa M, Kanzaki M,
Nakao A, Ooi A and Fujii H: Trastuzumab-mediated antibody-dependent
cellular cytotoxicity against esophageal squamous cell carcinoma.
Clin Cancer Res. 11:4898–4904. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moretta L, Locatelli F, Pende D, Marcenaro
E, Mingari MC and Moretta A: Killer Ig-like receptor-mediated
control of natural killer cell alloreactivity in haploidentical
hematopoietic stem cell transplantation. Blood. 117:764–771. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bae DS, Hwang YK and Lee JK: Importance of
NKG2D-NKG2D ligands interaction for cytolytic activity of natural
killer cell. Cell Immunol. 276:122–127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Okita R, Mougiakakos D, Ando T, Mao Y,
Sarhan D, Wennerberg E, Seliger B, Lundqvist A, Mimura K and
Kiessling R: HER2/HER3 signaling regulates NK cell-mediated
cytotoxicity via MHC class I chain-related molecule A and B
expression in human breast cancer cell lines. J Immunol.
188:2136–2145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Watanabe Y, Asano R, Arai K, Shimomura I,
Ogata H, Kawaguchi H, Hayashi H, Ohtsuka H, Yoshida H, Katayose Y,
et al: In vitro and in vivo antitumor effects of recombinant
bispecific antibodies based on humanized anti-EGFR antibody. Oncol
Rep. 26:949–955. 2011.PubMed/NCBI
|
|
29
|
Baselga J, Carbonell X, Castañeda-Soto NJ,
Clemens M, Green M, Harvey V, Morales S, Barton C and Ghahramani P:
Phase II study of efficacy, safety, and pharmacokinetics of
trastuzumab monotherapy administered on a 3-weekly schedule. J Clin
Oncol. 23:2162–2171. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hudis CA: Trastuzumab-mechanism of action
and use in clinical practice. N Engl J Med. 357:39–51. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Slamon DJ, Leyland-Jones B, Shak S, Fuchs
H, Paton V, Bajamonde A, Fleming T, Eiermann W, Wolter J, Pegram M,
et al: Use of chemotherapy plus a monoclonal antibody against HER2
for metastatic breast cancer that overexpresses HER2. N Engl J Med.
344:783–792. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kawaguchi Y, Kono K, Mimura K, Mitsui F,
Sugai H, Akaike H and Fujii H: Targeting EGFR and HER-2 with
cetuximab- and trastuzumab-mediated immunotherapy in oesophageal
squamous cell carcinoma. Br J Cancer. 97:494–501. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kawaguchi Y, Kono K, Mimura K, Sugai H,
Akaike H and Fujii H: Cetuximab induce antibody-dependent cellular
cytotoxicity against EGFR-expressing esophageal squamous cell
carcinoma. Int J Cancer. 120:781–787. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Omori S, Hida M, Fujita H, Takahashi H,
Tanimura S, Kohno M and Awazu M: Extracellular signal-regulated
kinase inhibition slows disease progression in mice with polycystic
kidney disease. J Am Soc Nephrol. 17:1604–1614. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fujiwara Y, Hosokawa Y, Watanabe K,
Tanimura S, Ozaki K and Kohno M: Blockade of the
phosphatidylinositol-3-kinase-Akt signaling pathway enhances the
induction of apoptosis by microtubule-destabilizing agents in tumor
cells in which the pathway is constitutively activated. Mol Cancer
Ther. 6:1133–1142. 2007. View Article : Google Scholar : PubMed/NCBI
|