Introduction
Esophageal squamous cell carcinoma (ESCC) is a
prominent subtype of esophageal cancer and has the fourth-highest
incidence of cancer-associated mortality in China, 2015 (1). Besides surgery, the current treatment
options for esophageal cancer include chemoradiotherapy and
chemotherapy, which is often accompanied by high toxicity and
acquired therapeutic resistance (2).
It is therefore urgent to identify novel drugs and regimens for the
treatment of esophageal cancer.
Protein neddylation is a novel regulatory
post-translational modification pathway of, in which ubiquitin-like
molecule neural precursor cell expressed, developmentally
downregulated 8 (NEDD8) was added to substrate proteins via a
sequential enzymatic process, which includes the NEDD8-activating
enzyme E1 (NAE), NEDD8-conjugating enzyme E2 and substrate-specific
NEDD8-E3 ligases (3–5). The best-characterized substrates of
neddylation are cullin family proteins (6), which are important skeleton protein of
Cullin-RING E3 ligases (CRLs) (6).
Previous studies demonstrated that the hyperactivated neddylation
pathway and abnormally activated CRL E3 ligases serve important
functions in disease progression, including poorer patient overall
survival rates (7–11). These results indicate that the
neddylation pathway is a potential anticancer target.
MLN4924 has been shown to function as a potent and
selective inhibitor of NEDD8-activating enzyme (NAE) (3); previous studies revealed that MLN4924
inhibits cancer cell growth in vitro and in vivo by
inactivating CRLs and causing the accumulation of substrates
(12–16). Previously, MLN4924 exhibited
attractive pharmacodynamic effects in phase I studies in patients
with advanced solid tumors, relapsed/refractory lymphoma and
metastatic melanoma for its tolerable safety profile (17–19).
MLN4924 may function as a novel chemosensitizer or radiosensitizer
in multiple types of cancer (20–24). These
results indicate that targeting the neddylation pathway may
represent an attractive anticancer strategy, either alone or in
combination with other therapies.
To further explore the effect and mechanism of
MLN4924 on esophageal cancer cell growth alone or in combination
with cisplatin (CDDP), a standard chemotherapy drug for patients
with esophageal cancer, the cell cycle arrest induced by MLN4924
treatment and the stability of cell-cycle associated proteins,
including cyclin-dependent kinase inhibitor 1 (CDKN1A; also known
as p21), CDKN1B (also known as p27) and Wee1-like protein kinase
(Wee1) were investigated. In addition, MLN4924 and CDDP have been
demonstrated to induce DNA damage, therefore the combinational
effect on DNA damage and apoptosis was also examined.
Materials and methods
Cell lines, culture and reagents
Human ESCC EC1 and Kyse450 cell lines were kindly
gifted from Professor Zhao (Zhengzhou University, Zhengzhou, China)
and cultured in Dulbecco's modified Eagle's medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) containing 10% fetal
bovine serum (FBS; Biochrom, Ltd., Cambridge, UK) at 37°C with 5%
CO2. MLN4924 was synthesized and prepared as previously
described (7,25).
Cell viability
Cell viability was detected using an ATPLite
Luminescence Assay kit (PerkinElmer, Inc., Waltham, MA, USA) as
previously described (7,25). Briefly, cells were seeded in 96-well
plates (3×103 cells/well) and treated with dimethyl
sulfoxide (DMSO; 0.1%) or MLN4924 (0.0, 0.1, 0.2, 0.3, 0.4, 0.6,
0.8, 1.0 µM). Cell proliferation was determined at 24, 48 and 72 h
with indicated concentration (0.0, 0.1, 0.2, 0.3, 0.4, 0.6, 0.8,
1.0 µM) using the assay kit according to the manufacturer's
instructions.
Clonogenic assay
A total of 500 cells were seeded into 6-well plates
in triplicate, treated with DMSO (0.1%), MLN4924 (0.05 µM), CDDP
(MedChem Express Co., Ltd., Shanghai, China) (1 µg/ml) or MLN4924
(0.05 µM) + CDDP (1 µg/ml) and then incubated for 12 days. The
colonies were fixed with 4% paraformaldehyde for 30 min at room
temperature and stained with crystal violet for 30 min at room
temperature. The colonies were counted under light field of
inverted microscope at ×100 magnification, and captured using Gel
Doc™ XR+ Gel Documentation System (Bio-Rad Laboratories, Inc.,
Shanghai, China). Colonies comprising 50 cells or more were
counted.
Cell cycle analysis
Cells treated with MLN4924 at indicated
concentrations were harvested, fixed in 70% ethanol at −20°C and
then stained with 50 µg/ml propidium iodide (PI) containing 30
µg/ml RNase A (both from Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) at 37°C for 30 min and analyzed for cell-cycle profile by
flow cytometry (Becton Dickinson FACScan; Becton-Dickinson, San
Jose, CA, USA). Data were analyzed with ModFit LT 3.1 software
(Verity Software House, Inc., Topsham, ME, USA).
Western blotting
EC1 and Kyse450 were treated with MLN4924 (0.1, 0.3,
0.6 µM) or DMSO (0.1%) for 72 h. Then the protein was extracted
using radioimmunoprecipitation buffer (Beyotime Institute of
Biotechnology, Jiangsu, China) and protein concentration was
determined using BCA kit (Beyotime Institute of Biotechnology).
When detected, 50 µg proteins were loaded per lane with 2% SDS. The
proteins were transferred to polyvinylidene fluoride membrane and
blocked with 5% skim milk for 2 h at room temperature. The protein
was detected using antibodies against p21 (cat. no. 2947), p27
(cat. no. 3686), Wee1 (cat. no. 4936), cyclin B (cat. no. 12231),
Phospho-Histone H3 (Ser10; cat. no. 3377), origin recognition
complex (ORC1; cat. no. 4731), DNA replication factor Cdt1 (CDT1;
cat. no. 8064), serine/threonine-protein kinase Chk1 (CHK1; cat.
no. 2360), CHK2 (cat. no. 2662), p-histone H2AX at Ser139 (γH2AX;
cat. no. 9718), cleaved caspase-3 (cat. no. 9664), cleaved
poly(ADP-ribose) polymerase (PARP; cat. no. 5625), GAPDH (cat. no.
2118) (all from Cell Signaling Technology, Inc., Danvers, MA, USA)
and p-CHK1 (cat. no. ab58567), p-CHK2 (cat. no. ab59408) (both from
Abcam, Cambridge, UK). All the primary antibodies were diluted in
1:1,000 and incubated at 4°C overnight. Secondary antibodies
peroxidase-conjugated goat anti-mouse IgG (cat. no. ZB-2305) and
peroxidase-conjugated goat anti-rabbit IgG (cat. no. ZB-2301) were
purchased from ZGSB-Bio, Inc., (Beijing, China). The membrane was
incubated with according secondary antibodies (1:3,000) for 2 h at
room temperature. Then the membrane was detected using an ECL Kit
(Beyotime Institute of Biotechnology).
Apoptosis detection
Cells were treated with the indicated concentration
of MLN4924 (0.3 µM), CDDP (1.6 µg/ml) and MLN4924 (0.3 µM) + CDDP
(1.6 µg/ml) for 72 h. Apoptosis was determined with the Annexin
V-Fluorescein Isothiocyanate/PI Apoptosis kit and CaspGLOW
Fluorescein Active Caspase-3 Staining kit (both from BioVision,
Inc., Milpitas, CA, USA), according to the manufacturer's
instructions. Using Annexin V-Fluorescein Isothiocyanate/PI,
apoptosis and the activity of caspase-3 were detected using a flow
cytometer, and results were analyzed using Summit 6.1 software (BD
Biosciences, Franklin Lakes, NJ, USA).
Immunofluorescence staining
EC1 and Kyse450 cells were treated with MLN4924
using 0.0, 0.3, 0.6 or 1.0 µM and fixed with 4% paraformaldehyde
for 30 min at room temperature, permeabilized using 0.2% Triton
X-100 for 10 min at room temperature, and incubated with γH2AX
primary antibody (cat. no. 9718; 1:200) at 4°C overnight and Alexa
Fluor-488 goat anti-rabbit immunoglobulin G (H+L) secondary
antibody (cat. no. A0423; 1:500) (Beyotime Institute of
Biotechnology) for 2 h at room temperature. The nuclei were stained
with DAPI (Beyotime Institute of Biotechnology). Images were
captured using a fluorescence microscope (magnification, ×200;
Olympus BX-51; Olympus Corp., Tokyo, Japan).
Cycloheximide (CHX) chase assay
Kyse450 and EC1 cells were pretreated with 0.6 µM
MLN4924 for 18 h to increase the basal protein level of proteins.
Cells were then washed with PBS to remove residual drug and divided
into 2 groups, which were further treated with 50 µg/ml CHX
(Beijing Solarbio Science and Technology Co., Ltd., Beijing, China)
or CHX+MLN4924 (0.6 µM) for indicated times and then collected for
western blot analysis, as aforementioned.
Statistical analysis
For Figs. 1A, 3B and 4A, the
statistical significance of differences between groups was assessed
using SPSS 17 software (SPSS, Inc., Chicago, IL, USA). One-way
analysis of variance followed by a least-significant difference
post hoc test was used to compare between multiple groups. For
Fig. 4B, the statistical significance
of differences between CDDP and MLN+CDDP was assessed using
GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla, CA,
USA. Unpaired Student's t-test was used for the comparison of
parameters between groups. P<0.05 was considered to indicate a
statistically significant difference. Data is presented as the mean
± standard.
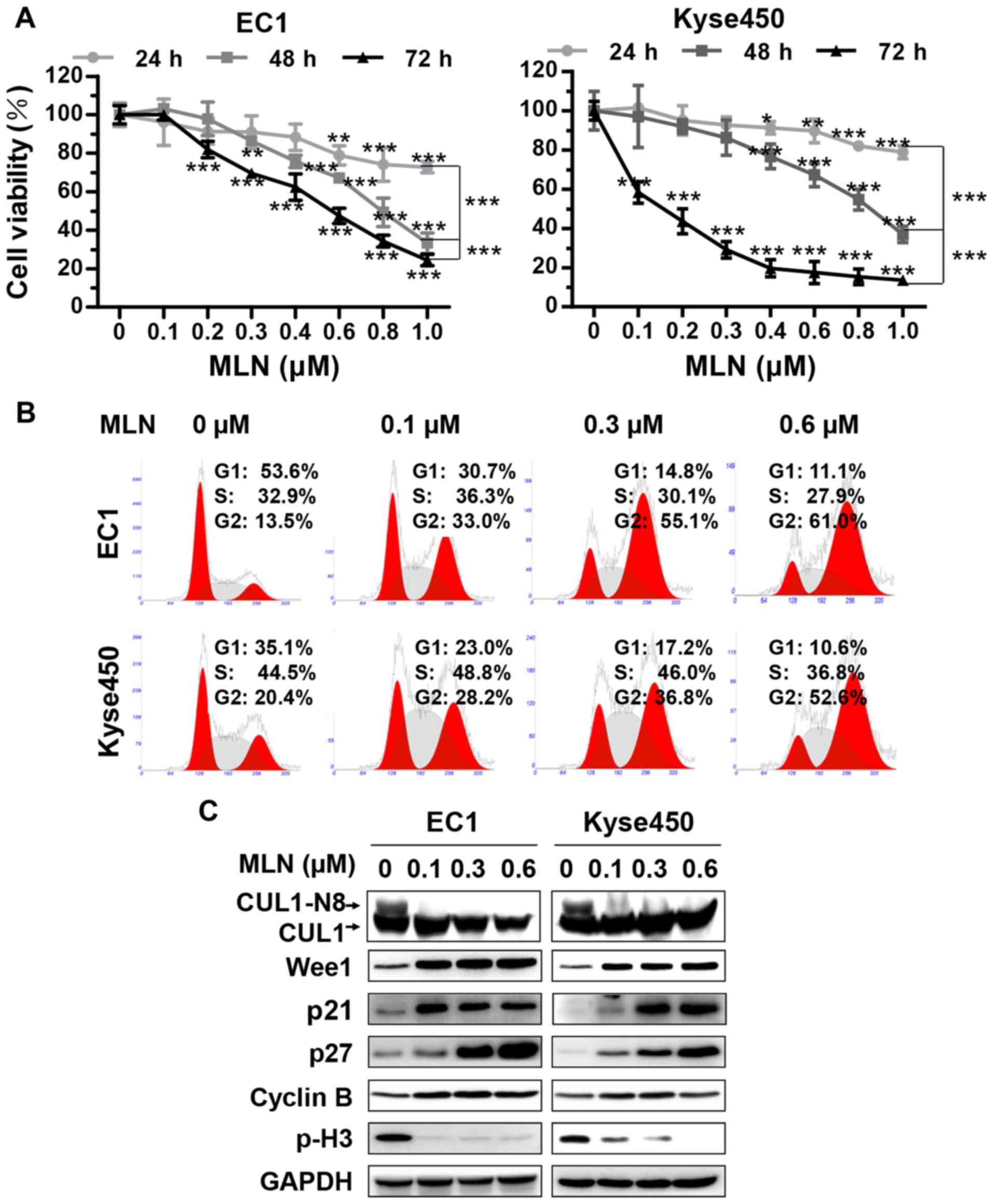 | Figure 1.MLN triggered G2 cell
cycle arrest in esophageal cancer cells. (A) MLN suppressed the
proliferation of esophageal cancer cells. Cells were seeded on a
96-well plate and treated with MLN at indicated concentrations.
Cell viability was measured at the time-points indicated using the
ATPLite assay. *P<0.05, **P<0.01, ***P<0.001 vs. 0 µM
unless marked otherwise; n=3. Error bars, standard deviation. (B)
MLN induced G2/M cell cycle arrest. EC1 and Kyse450
cells were treated with MLN at different concentration for 24 h,
followed by propidium iodide staining and FACS analysis for cell
cycle profile. (C) MLN induced accumulation of Wee1, p21, p27,
cyclin B and decrease of p-H3. EC1 and Kyse450 cells were treated
with MLN at indicated concentration for 72 h, and subjected to
western blotting. GAPDH served as a loading control. p21,
cyclin-dependent kinase inhibitor 1A; p27, cyclin-dependent kinase
inhibitor 1B; Wee1, Wee1-like protein kinase; CHX, cyclohexamide;
MLN, MLN4924. |
Results
Targeting neddylation by MLN4924
inhibited the proliferation of ESCC cells
To assess the anticancer function of MLN4924 on
esophageal cancer cells, the present study assessed the effect of
MLN4924 on the proliferation of the ESCC EC1 and Kyse450 cell lines
by ATPLite cell viability assay. MLN4924 inhibited the cell
proliferation of the 2 cell lines in a time- and dose-dependent
manner (Fig. 1A), which indicated
that MLN4924 could serve as an attractive anticancer drug for
patients with ESCC.
Neddylation inhibition with MLN4924
triggered G2 cell cycle arrest of ESCC cells
To investigate the mechanism of cell growth
inhibition, the effect of MLN4924 on cell cycle progression was
evaluated using PI staining. MLN4924 induced a marked
G2-M cell cycle arrest in the 2 treated cell lines in a
dose-dependent manner (Fig. 1B).
MLN4924 also induced accumulation of p21, p27, inhibitor of
G2-M phase transition Wee1 (26) and an increase in cyclin B, accompanied
by a decrease in the level of phosphohistone H3 (p-H3), a hallmark
of M-phase cells (27), indicating
that MLN4924-treated cells were arrested at the G2 phase
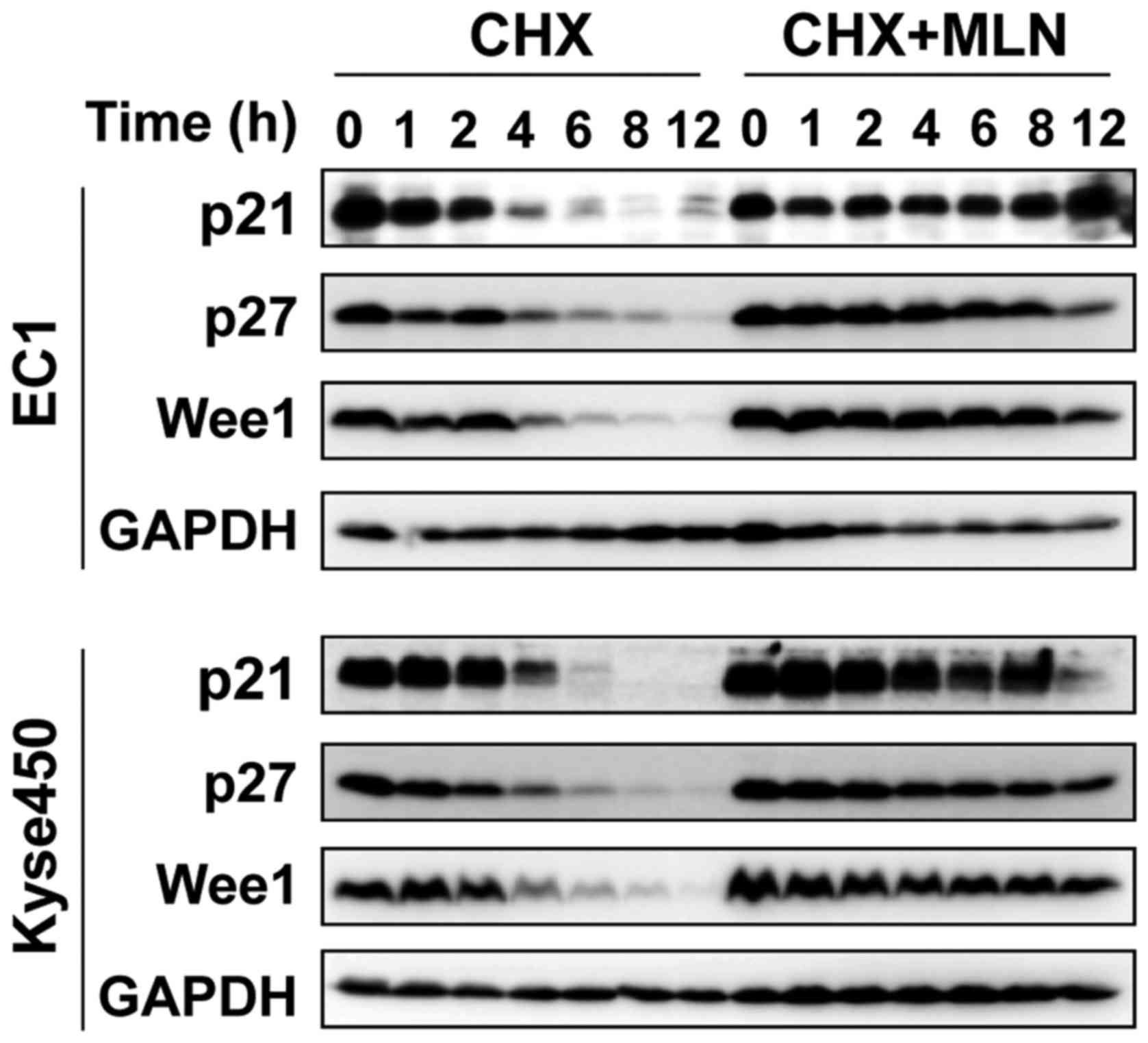
(Fig. 1C). Furthermore, following the
blocking of protein synthesis using CHX, the protein stability of
p21, p27 and Wee1 were markedly increased by MLN4924 treatment
(Fig. 2), indicating that
G2-phase cell cycle arrest may be attributable to the
accumulation of cell cycle-associated proteins.
MLN4924 induces DNA damage
response
Activation of the DNA damage response is an
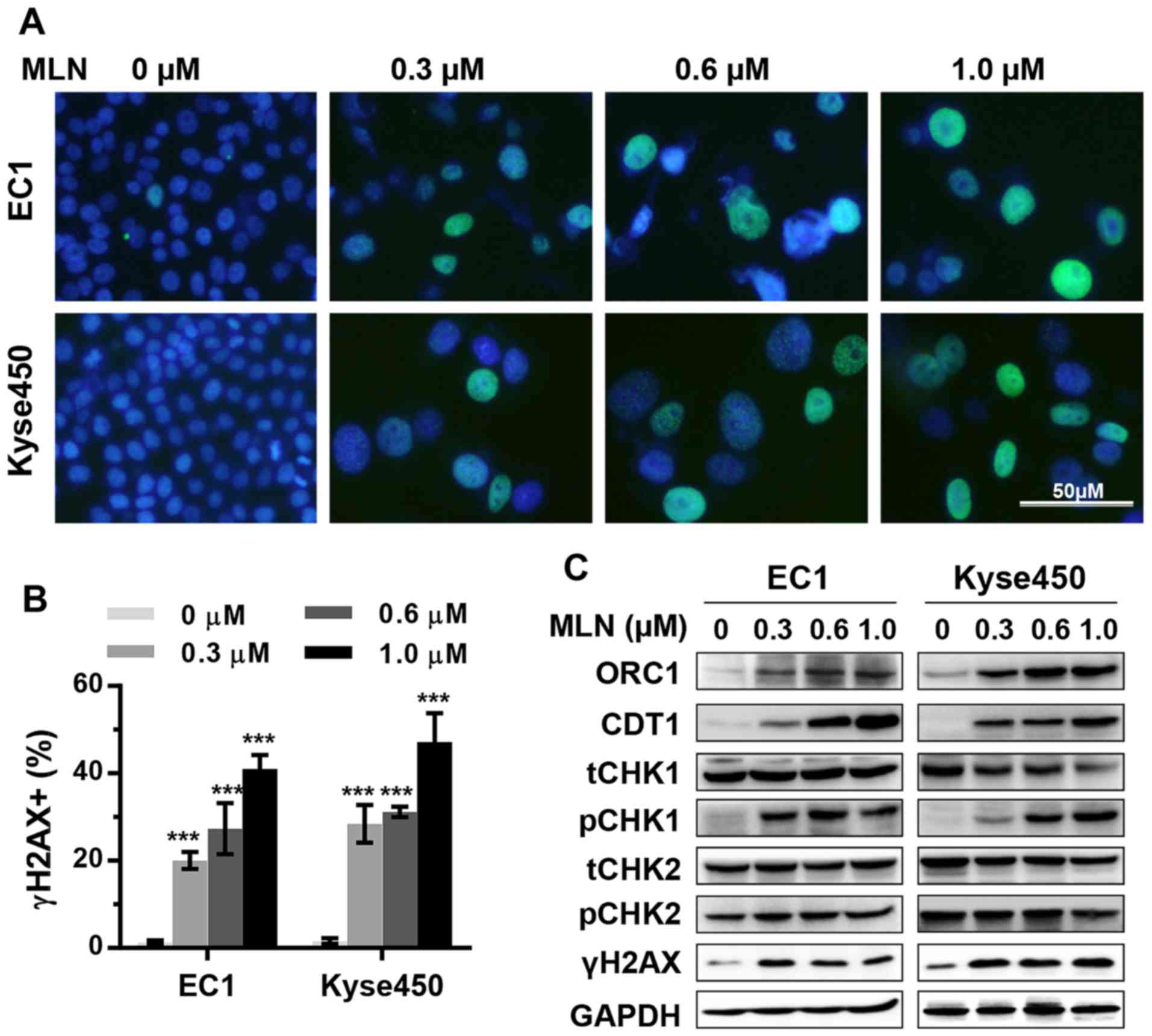
important property of cytotoxic drugs; the expression of γH2AX was
therefore determined to detect DNA double-strand breaks by
immunofluorescence and western blotting. MLN4924 induced sustained
γH2AX foci in the EC1 and Kyse450 cells in a dose-dependent manner
(Fig. 3A and B), which was further
confirmed by western blotting (Fig.
3C). MLN4924 treatment increased the level of CDT1 and ORC1
proteins in a dose-dependent manner (Fig.
3C). These results implied that the accumulation of
DNA-replication licensing proteins CDT1 and ORC1, known CRL
substrates (28), partially
contributed to the induction of DNA damage by MLN4924
treatment.
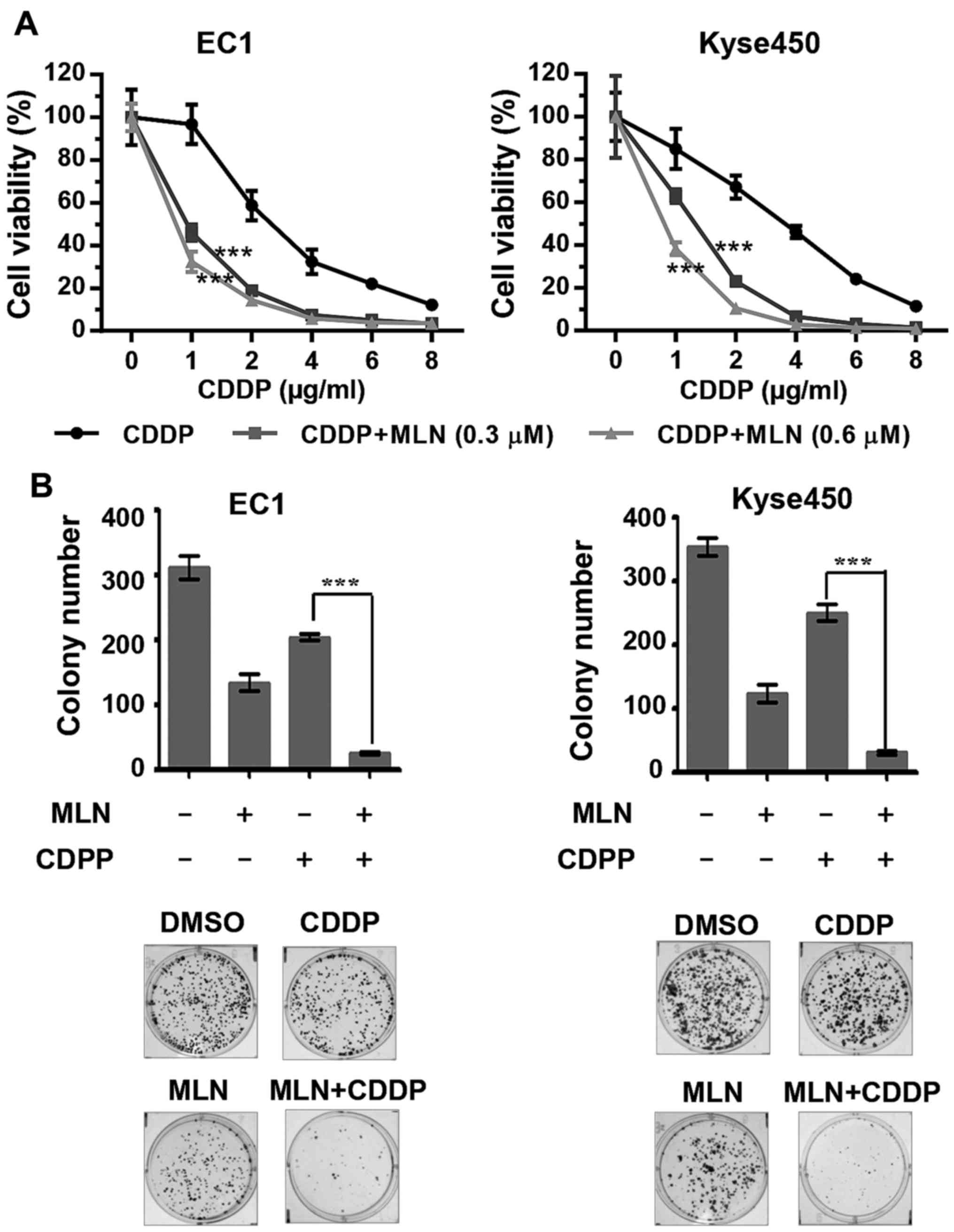
MLN4924 enhances the cytotoxicity of
CDDP to ESCC cells
To assess the function of MLN4924 as a novel
chemosensitizer to increase the anti-ESCC activity of CDDP, ESCC
EC1 and Kyse450 cell lines were treated with MLN4924, CDDP or the 2
together. Results demonstrated that MLN4924 significantly enhanced
the cytotoxicity of CDDP and inhibited cell viability (Fig. 4A; P<0.001) and clonogenic cell
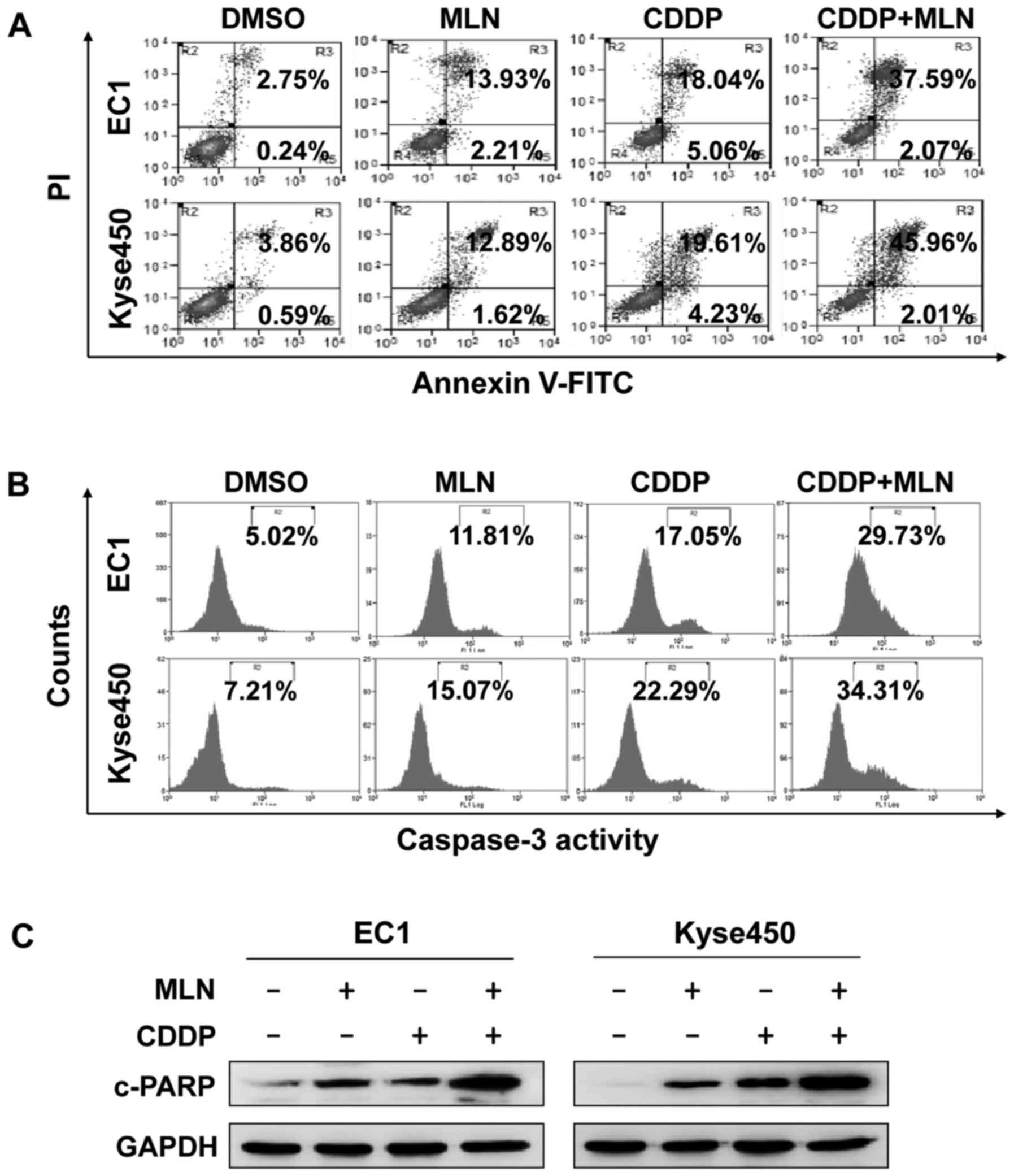
survival (Fig. 4B). The combination
of MLN4924 with CDDP markedly enhanced apoptosis compared with
either single agent, which is evident by the increased percentage
of Annexin V-positive (Fig. 5A) or
caspase-3 active cells (Fig. 5B) and
evaluated expression of cleaved-PARP (Fig. 5C).
Discussion
Previous studies demonstrated that MLN4924 inhibited
the growth of a broad panel of cancer cells in vitro and
in vivo (3,8,15,29–31). On
the basis of its promising anticancer efficacy and tolerated
toxicity, MLN4924 has undergone preclinical or phase I clinical
trials for multiple human malignancies (17–19).
Mechanistically, MLN4924 blocks cullin neddylation, inactivates
CRL, induces the accumulation of tumor-suppressive CRL substrates
and causes DNA damage, cell cycle arrest, cellular senescence and
apoptosis in a tumor cell-specific manner (3,7,15,29,31). To
date, the detailed mechanism of MLN4924 on cell proliferation
inhibition in human ESCC cells requires further investigation. The
present study demonstrated that MLN4924 could induce G2
cell cycle arrest in ESCC cells. At the same time, MLN4924
significantly enhanced the protein stability of p21, p27 and Wee1,
which is in accordance with previous studies (9,15), where
they indicate that all of the 3 proteins serve an important
function in MLN4924-induced G2 cell cycle arrest
(23).
G2/M-phase arrest is a crucial response
to DNA damage in the majority of cancer cells (32). The present study found that MLN4924
treatment triggered DNA damage response in ESCC cells, as
demonstrated by the appearance of γH2AX. Further analysis revealed
that 2 well-known CRL substrates, CDT1 and ORC1, were accumulated
in MLN4924-treated ESCC cells; their overexpression was known to
trigger DNA damage response (3,11,29).
CDDP is a first-line chemotherapeutic agent for the
treatment of ESCC (2). However, the
toxicity and the development of resistance often limit its efficacy
(33). Previously, a series of
studies revealed that MLN4924 could be used as a sensitizer to
chemotherapy: MLN4924 has been demonstrated to overcome resistance
to a range of chemotherapeutics, including CDDP (20,21),
carboplatin (21,34), bleomycin (35), doxorubicin (35) and etoposide (35) resistance in ovarian tumor cell lines
in vitro and in vivo. MLN4924 significantly augmented
the cytotoxicity of CDDP against CDDP-resistant cells in cervical
carcinoma xenografts (22). The
results of the present study revealed that the addition of MLN4924
significantly improved the efficacy of CDDP by enhancing the
incidence of apoptosis in ESCC cells. These results indicate that
MLN4924 could be used alone or in combination for the treatment of
human ESCC in future studies.
The present study demonstrated that inactivating the
neddylation pathway by MLN4924 suppressed the growth of esophageal
cancer cells by triggering G2 cell cycle arrest and
inducing DNA damage. Furthermore, the combination of MLN4924 and
CDDP more effectively inhibited cell growth compared with treatment
with either single drug by enhancing apoptosis in esophageal cancer
cells. These results extend the understanding of the function of
MLN4924 and propose its use in the treatment of esophageal cancer
in single treatment or in combination.
Acknowledgements
The authors thank Professor Guoqiang Zhao from
Zhengzhou University (Zhengzhou, China) for kindly providing the
EC1 and Kyse450 cell lines. The present study was supported by the
National Natural Science Foundation Grant of China (grant nos.
81001102, 81101894 and 81672421), the Outstanding Young Talent
Research Fund of Zhengzhou University (grant no. 51999223), the
Research Foundation of Education Bureau of Henan, China (grant no.
15A310024) and the Student's Platform for Innovation and
Entrepreneurship Training Program of Zhengzhou University (grant
no. 2016xjxm341).
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yam PC, Tong D and Law S: Comparisons of
sixth and seventh edition of the American Joint Cancer Committee
staging systems for esophageal cancer. Ann Surg Oncol. 21:583–588.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Soucy TA, Smith PG, Milhollen MA, Berger
AJ, Gavin JM, Adhikari S, Brownell JE, Burke KE, Cardin DP,
Critchley S, et al: An inhibitor of NEDD8-activating enzyme as a
new approach to treat cancer. Nature. 458:732–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Enchev RI, Schulman BA and Peter M:
Protein neddylation: Beyond cullin-RING ligases. Nat Rev Mol Cell
Biol. 16:30–44. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abidi N and Xirodimas DP: Regulation of
cancer-related pathways by protein NEDDylation and strategies for
the use of NEDD8 inhibitors in the clinic. Endocr Relat Cancer.
22:T55–T70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Siergiejuk E, Scott DC, Schulman BA,
Hofmann K, Kurz T and Peter M: Cullin neddylation and
substrate-adaptors counteract SCF inhibition by the CAND1-like
protein Lag2 in Saccharomyces cerevisiae. EMBO J.
28:3845–3856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen P, Hu T, Liang Y, Li P, Chen X, Zhang
J, Ma Y, Hao Q, Wang J, Zhang P, et al: Neddylation inhibition
activates the extrinsic apoptosis pathway through ATF4-CHOP-DR5
axis in human esophageal cancer cells. Clin Cancer Res.
22:4145–4157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hua W, Li C, Yang Z, Li L, Jiang Y, Yu G,
Zhu W, Liu Z, Duan S, Chu Y, et al: Suppression of glioblastoma by
targeting the overactivated protein neddylation pathway. Neuro
Oncol. 17:1333–1343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li L, Wang M, Yu G, Chen P, Li H, Wei D,
Zhu J, Xie L, Jia H, Shi J, et al: Overactivated neddylation
pathway as a therapeutic target in lung cancer. J Natl Cancer Inst.
106:dju0832014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xie P, Zhang M, He S, Lu K, Chen Y, Xing
G, Lu Y, Liu P, Li Y, Wang S, et al: The covalent modifier Nedd8 is
critical for the activation of Smurf1 ubiquitin ligase in
tumorigenesis. Nat Commun. 5:37332014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao Q, Yu GY, Shi JY, Li LH, Zhang WJ,
Wang ZC, Yang LX, Duan M, Zhao H, Wang XY, et al: Neddylation
pathway is up-regulated in human intrahepatic cholangiocarcinoma
and serves as a potential therapeutic target. Oncotarget.
5:7820–7832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuo KL, Ho IL, Shi CS, Wu JT, Lin WC, Tsai
YC, Chang HC, Chou CT, Hsu CH, Hsieh JT, et al: MLN4924, a novel
protein neddylation inhibitor, suppresses proliferation and
migration of human urothelial carcinoma: In vitro and in vivo
studies. Cancer Lett. 363:127–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wan J, Zhu J, Li G and Zhang Z:
Radiosensitization of human colorectal cancer cells by MLN4924: An
inhibitor of NEDD8-activating enzyme. Technol Cancer Res Treat.
15:527–534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lan H, Tang Z, Jin H and Sun Y:
Neddylation inhibitor MLN4924 suppresses growth and migration of
human gastric cancer cells. Sci Rep. 6:242182016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Y, Luo Z, Pan Y, Wang W, Zhou X,
Jeong LS, Chu Y, Liu J and Jia L: Targeting protein neddylation
with an NEDD8-activating enzyme inhibitor MLN4924 induced apoptosis
or senescence in human lymphoma cells. Cancer Biol Ther.
16:420–429. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han K, Wang Q, Cao H, Qiu G, Cao J, Li X,
Wang J, Shen B and Zhang J: The NEDD8-activating enzyme inhibitor
MLN4924 induces G2 arrest and apoptosis in T-cell acute
lymphoblastic leukemia. Oncotarget. 7:23812–23824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bhatia S, Pavlick AC, Boasberg P, Thompson
JA, Mulligan G, Pickard MD, Faessel H, Dezube BJ and Hamid O: A
phase I study of the investigational NEDD8-activating enzyme
inhibitor pevonedistat (TAK-924/MLN4924) in patients with
metastatic melanoma. Invest New Drugs. 34:439–449. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shah JJ, Jakubowiak AJ, O'Connor OA,
Orlowski RZ, Harvey RD, Smith MR, Lebovic D, Diefenbach C, Kelly K,
Hua Z, et al: Phase I study of the novel investigational
NEDD8-activating enzyme inhibitor pevonedistat (MLN4924) in
patients with relapsed/refractory multiple myeloma or lymphoma.
Clin Cancer Res. 22:34–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sarantopoulos J, Shapiro GI, Cohen RB,
Clark JW, Kauh JS, Weiss GJ, Cleary JM, Mahalingam D, Pickard MD,
Faessel HM, et al: Phase I study of the investigational
NEDD8-activating enzyme inhibitor pevonedistat (TAK-924/MLN4924) in
patients with advanced solid tumors. Clin Cancer Res. 22:847–857.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nawrocki ST, Kelly KR, Smith PG, Espitia
CM, Possemato A, Beausoleil SA, Milhollen M, Blakemore S, Thomas M,
Berger A and Carew JS: Disrupting protein NEDDylation with MLN4924
is a novel strategy to target cisplatin resistance in ovarian
cancer. Clin Cancer Res. 19:3577–3590. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jazaeri AA, Shibata E, Park J, Bryant JL,
Conaway MR, Modesitt SC, Smith PG, Milhollen MA, Berger AJ and
Dutta A: Overcoming platinum resistance in preclinical models of
ovarian cancer using the neddylation inhibitor MLN4924. Mol Cancer
Ther. 12:1958–1967. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin WC, Kuo KL, Shi CS, Wu JT, Hsieh JT,
Chang HC, Liao SM, Chou CT, Chiang CK, Chiu WS, et al: MLN4924, a
novel NEDD8-activating enzyme inhibitor, exhibits antitumor
activity and enhances cisplatin-induced cytotoxicity in human
cervical carcinoma: In vitro and in vivo study. Am J Cancer Res.
5:3350–3362. 2015.PubMed/NCBI
|
|
23
|
Wang X, Zhang W, Yan Z, Liang Y, Li L, Yu
X, Feng Y, Fu S, Zhang Y, Zhao H, et al: Radiosensitization by the
investigational NEDD8-activating enzyme inhibitor MLN4924
(pevonedistat) in hormone-resistant prostate cancer cells.
Oncotarget. 7:38380–38391. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ho IL, Kuo KL, Liu SH, Chang HC, Hsieh JT,
Wu JT, Chiang CK, Lin WC, Tsai YC, Chou CT, et al: MLN4924
synergistically enhances cisplatin-induced cytotoxicity via JNK and
Bcl-xL pathways in human urothelial carcinoma. Sci Rep.
5:169482015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen P, Hu T, Liang Y, Li P, Chen X, Zhang
J, Ma Y, Hao Q, Wang J, Zhang P, et al: Neddylation inhibition
activates the extrinsic apoptosis pathway through ATF4-CHOP-DR5
axis in human esophageal cancer cells. Clin Cancer Res.
22:4145–4157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sarcar B, Kahali S, Prabhu AH, Shumway SD,
Xu Y, Demuth T and Chinnaiyan P: Targeting radiation-induced G(2)
checkpoint activation with the Wee-1 inhibitor MK-1775 in
glioblastoma cell lines. Mol Cancer Ther. 10:2405–2414. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fischle W, Tseng BS, Dormann HL,
Ueberheide BM, Garcia BA, Shabanowitz J, Hunt DF, Funabiki H and
Allis CD: Regulation of HP1-chromatin binding by histone H3
methylation and phosphorylation. Nature. 438:1116–1122. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Emanuele MJ, Elia AE, Xu Q, Thoma CR,
Izhar L, Leng Y, Guo A, Chen YN, Rush J, Hsu PW, et al: Global
identification of modular cullin-RING ligase substrates. Cell.
147:459–474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oladghaffari M, Islamian JP, Baradaran B
and Monfared AS: MLN4924 therapy as a novel approach in cancer
treatment modalities. J Chemother. 28:74–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Paiva C, Godbersen JC, Berger A, Brown JR
and Danilov AV: Targeting neddylation induces DNA damage and
checkpoint activation and sensitizes chronic lymphocytic leukemia B
cells to alkylating agents. Cell Death Dis. 6:e18072015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao Y, Morgan MA and Sun Y: Targeting
Neddylation pathways to inactivate cullin-RING ligases for
anticancer therapy. Antioxid Redox Signal. 21:2383–2400. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stark GR and Taylor WR: Analyzing the G2/M
checkpoint. Methods Mol Biol. 280:51–82. 2004.PubMed/NCBI
|
|
33
|
Florea AM and Büsselberg D: Cisplatin as
an anti-tumor drug: Cellular mechanisms of activity, drug
resistance and induced side effects. Cancers. 3:1351–1371. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
de Sousa GF, Mde A Lima, Custodio DF,
Freitas VM and Monteiro G: Chemogenomic study of carboplatin in
saccharomyces cerevisiae: Inhibition of the NEDDylation process
overcomes cellular resistance mediated by HuR and cullin proteins.
PLoS One. 10:e01453772015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pan WW, Zhou JJ, Yu C, Xu Y, Guo LJ, Zhang
HY, Zhou D, Song FZ and Fan HY: Ubiquitin E3 ligase
CRL4(CDT2/DCAF2) as a potential chemotherapeutic target for ovarian
surface epithelial cancer. J Biol Chem. 288:29680–29691. 2013.
View Article : Google Scholar : PubMed/NCBI
|