Introduction
Acute myeloid leukemia (AML) is a clonal disorder
that comprises a group of clonal malignant diseases, in which the
cancer cells originate from the bone marrow and display genetic
instability, leading to the accumulation of abnormal immature
myeloid cells in the bone marrow and blood (1). The principal therapeutic strategies for
patients with AML are aggressive chemotherapeutic regimens and
hematopoietic stem cell transplantation (HSCT) (1–3). However,
AML consists of 8 subtypes, including M0 (minimally
differentiated AML), M1 (AML without maturation),
M2 (AML with maturation), M3 (acute
promyelocytic leukemia), M4 (acute myelomonocytic
leukemia), M5 [acute monocytic leukemia (AMoL)],
M6 (erythroleukemia) and M7 (acute
megakaryoblastic leukemia). Patients with AML who receive intensive
chemotherapy may achieve complete remission; however, the overall
survival rate of patients with AML is poor and the treatment is
typically associated with serious complications (4). In addition, there is no optimal
therapeutic schedule for each type of AMoL (5). Therefore, the aim of the present study
was to identify a novel drug for treating M5 (AMoL).
The development of AMoL is a complex, multistep and
multifactorial process. Features of M5 typically include
increased numbers of white blood cells, an increased rate of marrow
infiltration and an increase in chromosomal aberrations including
translocation, mutation, aneuploidy and the formation of fusion
genes (6,7). Furthermore, the clinical complete
remission rate is low with poor prognosis. A total of 87 different
translocations have been identified, with 11q23 chromosomal
abnormalities accounting for ~22% of M5 cases. There are
various chemotherapy regimens used to treat patients with AMoL
including: i) Homoharringtonine, cytarabine and etoposide; ii)
daunorubicin, Ara-c and teniposide; and iii) mitoxantrone, Ara-c
and teniposide (7). However, AMoL is
insensitive to a number of chemotherapy regimens, thus patients
with AMoL exhibit a low remission rate and decreased survival time
(2,6,7).
Therefore, novel effective drugs are required.
Since novel antitumor drugs fell under
investigation, bioactive natural products have emerged and gained
considerable attention. Artemisinin is an effective drug for
treating malaria and belongs to the family of sesquiterpene
lactones, which are produced by the Artemisia annua plant
(8). Dihydroartemisinin (DHA) is a
water-soluble semi-synthetic derivative of artemisinin (9), and it is commercially combined with
piperaquine as an effective therapy for malaria with limited side
effects (10,11). In addition, DHA has been identified to
exhibit inhibitory effects on cancer cells, including lung
carcinoma (12), osteosarcoma
(13) and ovarian cancer (14). Previous studies have demonstrated that
DHA may inhibit molt-4 acute lymphoid cell leukemia cells (15,16).
However, the effects and underlying mechanisms of action of DHA in
AMoL remain unknown.
Therefore, the aim of the present study was to
analyze the antitumor effects of DHA against AMoL and to
investigate its underlying biological mechanisms. An in
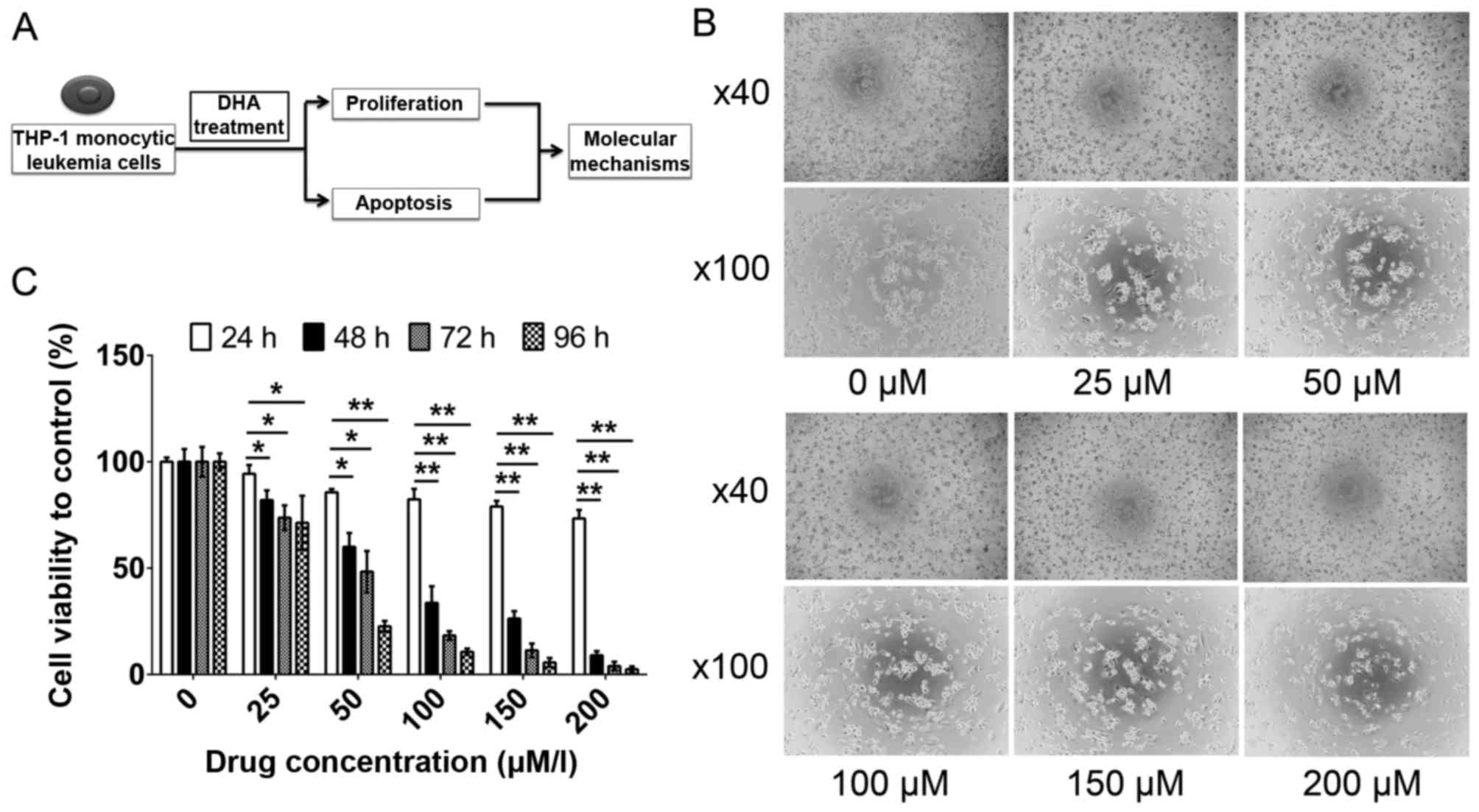
vitro approach (Fig. 1A) was used
to elucidate the molecular mechanisms underlying these effects of
DHA. The results of the present study verified that DHA exhibited
effective antitumor activity against AMoL in vitro via the
downregulation of phospho-protein kinase B (p-Akt) and
p-extracellular signal-regulated kinase (ERK) protein expression,
and the upregulation of cleaved caspase-3 levels.
Materials and methods
Reagents and antibodies
The chemical reagents, including DHA, used in the
present study were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany) unless stated otherwise. A stock solution (200
mM) of DHA was prepared in dimethyl sulfoxide (Sigma-Aldrich; Merck
KGaA) and stored at −20°C. Primary antibodies against p-ERK (cat.
no. 4348S), total (t)-ERK (cat. no. 4695S), p-Akt (cat. no. 4060S),
t-Akt (cat. no. 4685S), cleaved caspase-3 (cat. no. 1050S), β-actin
(cat. no. 4970S) and secondary antibodies (cat. no. 7074) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA).
Cell culture
The THP-1 human acute monocytic leukemia cell line
was purchased from the Chinese Academy of Sciences (Shanghai,
China). THP-1 cells were cultured in RPMI-1640 (HyClone; GE
Healthcare, Logan, UT, USA) supplemented with 10% fetal bovine
serum, 100 U/ml penicillin and 100 µg/ml streptomycin (all from
HyClone; GE Healthcare) at 37°C in a humidified atmosphere
containing 5% CO2. All cells were used within 20
passages.
Cell viability assay
DHA cytotoxicity was determined using a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan), which evaluated the metabolic activity of viable
cells. A total of 20,000 cells/well were seeded into 96-well plates
in RPMI-1640 medium, which contained 100 nM phorbol 12-myristate
13-acetate to induce adherence as THP-1 cells were suspending
cells, and subsequently treated with 0, 25, 50, 100, 150 or 200 µM
DHA for 24, 48, 72 and 96 h at 37°C. Following treatment with DHA
for 24, 48, 72 and 96 h, the RPMI-1640 medium was removed and the
cells were washed with PBS. Subsequently, 100 µl RPMI-1640 medium
and 10 µl CCK-8 solution was added to each well and incubated at
37°C for 2.5 h. The optical density (OD) at 450 nm was determined
daily for the following 4 days using a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA). All measurements were
repeated in triplicate. The cell viability compared with the
control group was calculated using the following equation: Cell
viability to control (%)=ODdrug-treated
group/ODcontrol group.
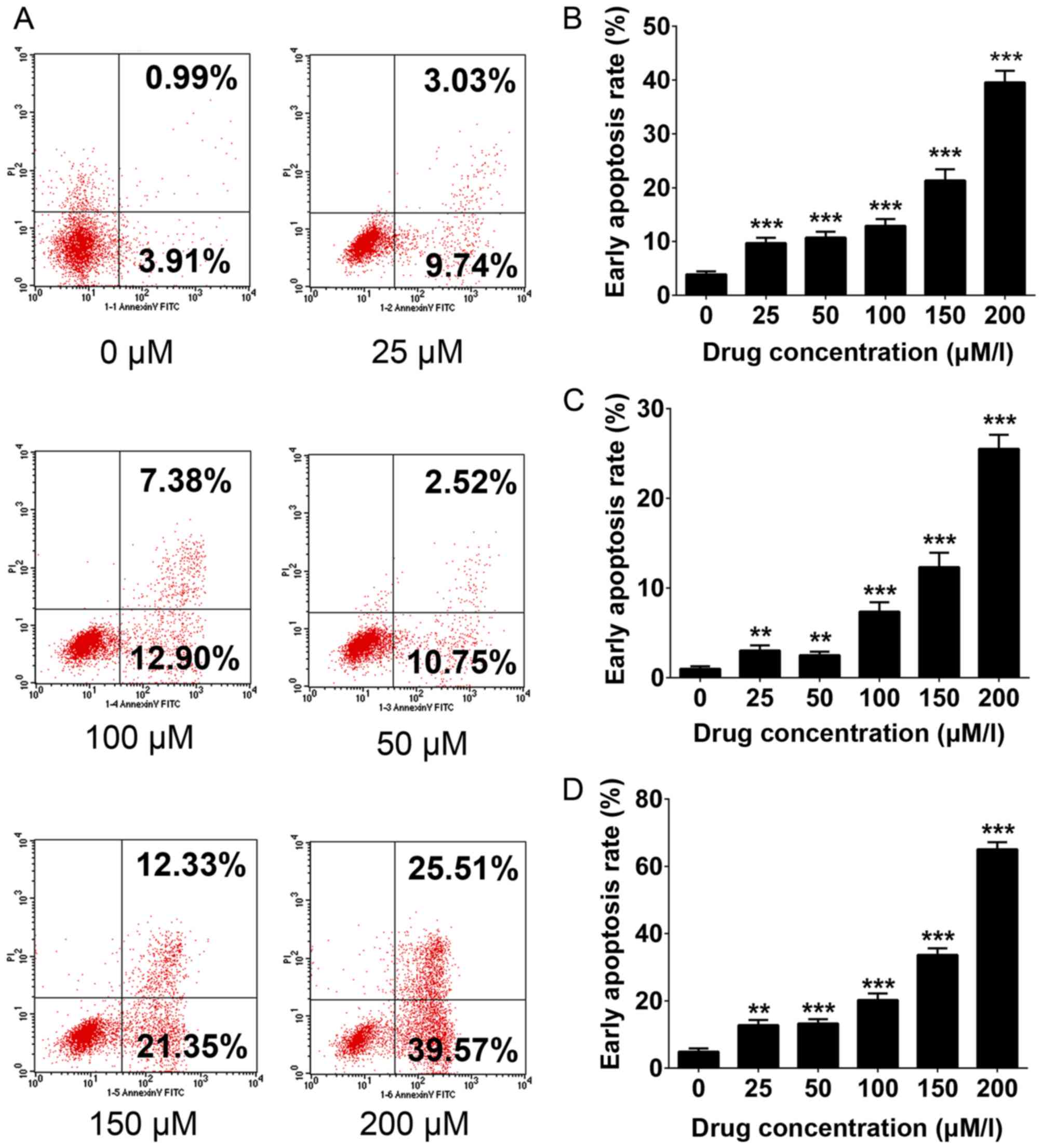
Apoptosis analysis
Cells were seeded at 1×106 cells/well in
6-well plates and harvested 24 h after treatment with 0, 25, 50,
100, 150 and 200 µM DHA. Cells were centrifuged at 500 × g for 5
min at 4°C and the supernatants were discarded. Subsequently, the
cells were resuspended in 1X annexin-binding buffer (BD
Biosciences, Franklin Lakes, NJ, USA). Apoptotic cells were
determined using annexin V-fluorescein isothiocyanate and propidium
iodide staining (BD Biosciences, Franklin Lakes, NJ, USA) and
quantified using a flow cytometer (BD Biosciences). FlowJo software
(version 7.6.1; FlowJo LLC, Ashland, OR, USA) was used to analyze
the rate of apoptosis. The cell proliferation results presented in
Fig. 1B and C demonstrate the
inhibitory effect of DHA on THP-1 cell proliferation. Therefore,
further analysis was used to determine whether DHA is able to
induce apoptosis. In Fig. 2A, the
lower right quadrant represents early apoptosis, whereas the upper
right quadrant represents late apoptosis. The proportion of
apoptotic cells identified includes cells in early and late
apoptosis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
THP-1 cells (1×106 cells/well) were
seeded at in 6-well plates and harvested following treatment with
0, 25, 50, 100, 150 and 200 µM DHA for 24 h. Cells were centrifuged
at 500 × g for 5 min at 4°C and the supernatants were discarded.
Total RNA was isolated from THP-1 cells treated with 0, 25, 50,
100, 150 and 200 µM DHA using the AxyPrep™ Multisource Total RNA
Miniprep kit (Axygen Scientific, Inc., Union City, CA, USA).
Equivalent amounts of RNA were converted into cDNA using the
PrimeScript™ RT Reagent kit (Takara Bio, Inc., Otsu,
Japan) at 37°C for 15 min and 85°C for 5 sec. cDNA was measured
using a Nanodrop 2000 instrument (Thermo Fisher Scientific,
Waltham, MA, USA). qPCR was performed using an ABI 7500 Sequencing
Detection System and SYBR® Premix Ex Taq (Takara Bio,
Inc.). All procedures were performed according to the
manufacturers' protocols. Cycling conditions included 40 cycles of
95°C for 5 sec and 60°C for 34 sec. Gene expression was quantified
using the 2−∆∆Cq method (17). β-actin was used as the reference gene
and all primer sequences are listed in Table I.
 | Table I.Sequences of primers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Sequences of primers used in reverse
transcription-quantitative polymerase chain reaction.
| Gene | Direction | Primer sequence
(5′-3′) |
|---|
| Akt1 | Forward |
ATGAGCGACGTGGCTATTGTGAAG |
|
| Reverse |
GAGGCCGTCAGCCACAGTCTGGATG |
| Akt2 | Forward |
ATGAATGAGGTGTCTGTCATCAAAGAAGGC |
|
| Reverse |
TGCTTGAGGCTGTTGGCGACC |
| Akt3 | Forward |
CAGTCTGTCTGCTACAGCCTGGATA |
|
| Reverse |
ATGAGCGATGTTACCATTGT |
| Bcl-2 | Forward |
GAACTGGGGGAGGATTGTGG |
|
| Reverse |
CCGTACAGTTCCACAAAGGC |
| Bax | Forward |
CCAGAGGCGGGGTTTCAT |
|
| Reverse |
GGAAAAAGACCTCTCGGGGG |
| β-actin | Forward |
CCAACCGCGAGAAGATGA |
|
| Reverse |
CCAGAGGCGTACAGGGATAG |
Western blot analysis
THP-1 cells were analyzed using qPCR and cells were
lysed in lysis buffer containing complete EDTA-free tablets which
are a protease inhibitor cocktail containing phenylmethylsulfonyl
fluoride, aprotinin, bestatin, E-64, leupeptin and pepstatin A
(Roche Applied Science, Penzbery, Germany). The protein
concentration was quantified using the bicinchoninic acid protein
assay kit (Santa Cruz Biotechnology, Inc., Dallas, TX, USA). For
western blot analysis, 20 µg total protein were boiled and
subsequently separated by SDS-PAGE using a 10% gel for p-Akt, total
(t-) Akt, p-ERK, t-ERK and β-actin and a 12.5% gel for cleaved
caspase-3. Following electrophoresis, proteins were blotted onto
polyvinylidene difluoride membranes and blocked by 5% skimmed milk
suspended in Tris-buffered saline with Tween-20 for 1 h at room
temperature. Each membrane was incubated with appropriate primary
antibodies against p-Akt, t-Akt, p-ERK, t-ERK, cleaved caspase-3
and β-actin, with a dilution ratio of 1:1,000 at 4°C overnight.
Blots were subsequently incubated with a horseradish
peroxidase-conjugated secondary antibody for 1 h at room
temperature. Protein bands were visualized using X-ray films and an
enhanced chemiluminescence detection system (GE Healthcare Life
Sciences, Little Chalfont, UK). Positive immunoreactive bands were
densitometrically quantified and normalized to β-actin. Adobe
Photoshop (Creative Suite 5; Adobe Systems, Inc., San Jose, CA,
USA) was used for densitometry.
Statistical analysis
SPSS software (version 19.0; IBM Corp., Armonk, NY,
USA) was used to analyze the data. The differences between the
experimental groups and controls were assessed using the Student's
t-test or one-way analysis of variance as appropriate. The data are
expressed as the mean ± standard deviation. All data were obtained
from at least three independent experiments. P<0.05 was
considered to indicate a statistically significant difference.
Results
THP-1 cell viability decreases
following DHA treatment
To investigate the effects of DHA, a cell viability
assay using CCK-8 was performed on THP-1 cells. The viability of
THP-1 cells was inhibited following treatment with DHA, accompanied
by the appearance of morphological characteristics of apoptosis
(Fig. 1B). The inhibitory effect of
DHA on the viability of THP-1 cells was markedly increased at
higher DHA concentrations following 96 h of treatment (P<0.05;
Fig. 1C). DHA may inhibit the
proliferation of THP-1 cells in a dose- and time-dependent
manner.
DHA induces the apoptosis of THP-1
cells
As presented in Fig.
2, flow cytometry analysis was used to evaluate DHA-induced
apoptosis. DHA significantly increased the total apoptosis rate
(4.9 control vs. 12.77, 13.24, 20.28, 33.68 and 65.08% when treated
with 25, 50, 100, 150 and 200 µM DHA, respectively; P<0.05;
Fig. 2A and D), and quantitative data
regarding the early and late apoptosis rate were consistent with
this (Fig. 2B and C). The results of
the present study validated the inhibitory function of DHA
functioned via the activation of the apoptosis pathway in THP-1
cells.
Downstream gene expression
To investigate the molecular mechanisms underlying
the inhibitory effects of DHA in THP-1 human monocytic leukemia
cells, RT-qPCR was used to examine variations in gene expression.
It was demonstrated that the gene expression levels of Akt1, Akt2,
Akt3 and B-cell lymphoma 2 (Bcl-2) were decreased in DHA-treated
THP-1 cells in a dose-dependent manner, whereas the expression of
Bcl-2-associated X protein (Bax) was upregulated (Fig. 3). The results indicated that these
genes are potential downstream targets of DHA in monocytic leukemia
treatment.
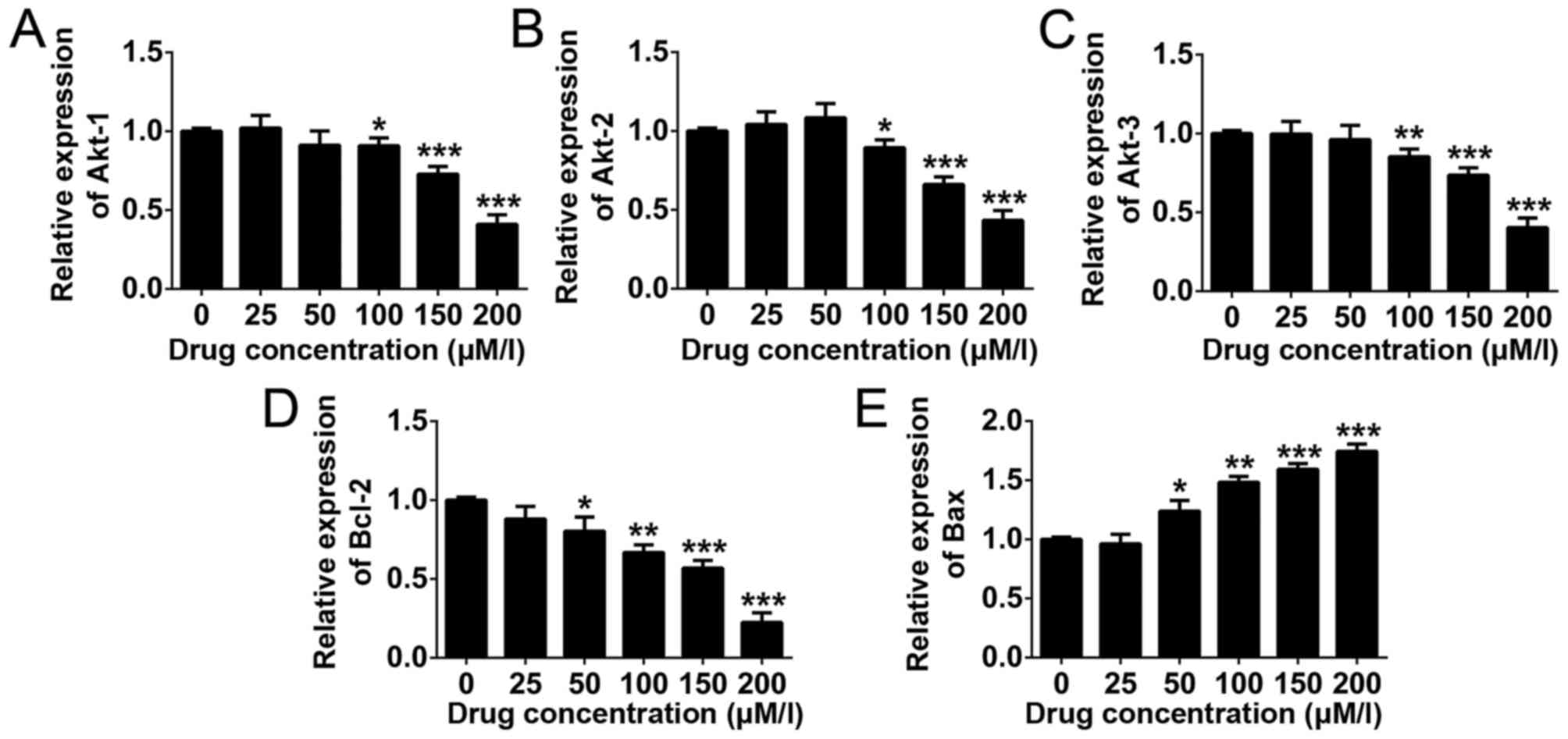 | Figure 3.Apoptosis-associated gene expression
in THP-1 cells following DHA treatment. RT-qPCR was employed to
detect the expression of (A) Akt1, (B) Akt2, (C) Akt3, (D) Bcl-2
and (E) Bax in DHA-treated THP-1 cells. Data are presented as the
mean ± standard deviation. ***P<0.001, **P<0.01, *P<0.05.
All data were obtained from at least three independent experiments.
RT-qPCR, reverse transcription quantitative polymerase chain
reaction; Akt, protein kinase B; Bcl-2, B cell lymphoma 2; Bax,
Bcl-2-associated X protein; DHA, dihydroartemisinin. |
Akt, ERK and cleaved caspase-3 are
potential downstream targets of DHA
Western blot analysis was used to determine the
protein expression levels of phospho (p)-Akt/total-Akt,
p-ERK/total-ERK and cleaved caspase-3 in THP-1 cells, following
different concentrations of DHA treatment. The results revealed
that the levels of p-Akt and p-ERK with increasing concentrations
of DHA (Fig. 4A-C). However, DHA
increased the levels of cleaved caspase-3 in a dose-dependent
manner (Fig. 4A and D). Therefore,
the results suggested that Akt, ERK and cleaved caspase-3 are
potential downstream targets of DHA-induced apoptosis in THP-1
cells.
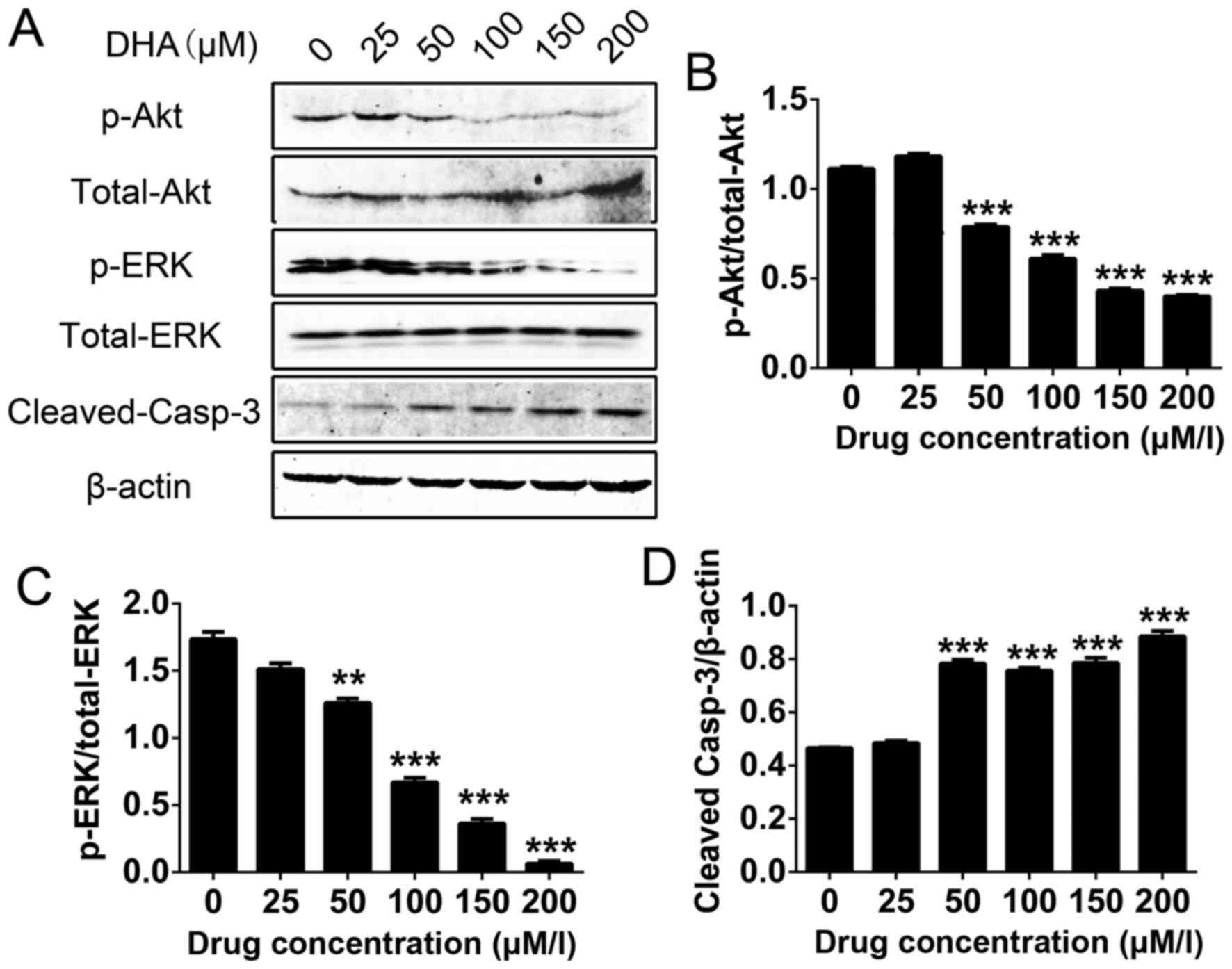 | Figure 4.Western blots of p-t-Akt, p-ERK/t-ERK
and cleaved caspase-3 levels following the DHA-induced apoptosis of
THP-1 cells. (A) Expression of p/total-Akt, p-ERK/total-ERK and
cleaved caspase-3 in THP-1 cells following DHA treatment. Levels of
(B) p-t-Akt, (C) p-ERK/t-ERK and (D) cleaved caspase-3 in THP-1
cells. Results are expressed as the ratio of cleaved caspase-3 to
β-actin, p-Akt to t-Akt and p-ERK to t-ERK. Data are presented as
the mean ± standard deviation. ***P<0.001, **P<0.01. All data
were obtained from at least three independent experiments. p-,
phosphorylated; t-, total; DHA, dihydroartemisinin; Akt, protein
kinase B; ERK, extracellular-signal-regulated kinase. |
Discussion
Induction failure and a high incidence of relapse
due to drug resistance are the principal problems surrounding AML
treatment (2,18). The use of novel drugs is one approach
for treating patients who are resistant to standard therapies
(19). Clinical evaluation of
potentially effective drugs is essential to cancer chemotherapy,
and may improve the prognosis of patients with refractory leukemia
(20). AMoL is a rare but distinct
disease, the increasing number of monocytes throughout its clinical
course characterizes the disease, and cytogenetic characterization
is required for diagnosis and prognosis stratification (21,22). Thus,
there is a requirement to identify less toxic and more efficacious
treatment alternatives. As a result, increasing attention has been
focused on the application of natural products in the treatment of
AMoL (3).
DHA, one of the bioactive derivatives of
artemisinin, has been investigated for the treatment of certain
tumor types; Professor Tu YouYou, who discovered DHA, was awarded
the Nobel Prize in 2015 (23,24). A number of additional pharmacological
effects of DHA have been identified, including antitumor activity
towards hepatocellular carcinoma in vitro and in vivo
(25). Furthermore, DHA was
identified to prevent breast cancer-induced osteolysis via
inhibiting breast cancer cells and osteoclasts (26). For leukemia treatment, DHA can induce
autophagy and inhibit the growth of iron-loaded human myeloid
leukemia K562 cells via reactive oxygen species toxicity (27); DHA and its derivative induce apoptosis
in acute myeloid leukemia cells through the Noxa-mediated pathway,
requiring iron and an endoperoxide moiety (28). However, the effect of DHA on AMoL has
yet to be fully elucidated.
The aim of the present study was to investigate
whether DHA had antitumor activity against human AMoL. It was
identified that DHA has a strong anti-leukemia effect in the THP-1
AMoL cell line in vitro. The results indicated that DHA
inhibited the spontaneous growth of THP-1 cells in a time- and
dose-dependent manner. To further investigate the mechanisms of DHA
anti-leukemia activity, the effect of DHA was analyzed with regards
to apoptosis and the activation of Akt/ERK survival signaling and
Bcl-2/Bax/caspase-3 apoptosis pathways that are constitutively
expressed in THP-1 cells. Results of the present study indicated
that DHA-induced apoptosis was caused by downregulating Akt/ERK
signaling and activating the caspase-3 pathway, following the
balance of the Bcl-2/Bax axis.
Distinct from normal cells, leukemia cells often
express constitutively active survival-signaling pathways, such as
Akt and ERK among others, due to gene mutations, rearrangements and
chromosomal translocations; these survival signaling pathways serve
vital roles in tumorigenesis, proliferation, anti-apoptosis and
drug resistance (29–31). The higher the number of constitutively
active growth signaling pathways in acute myelogenous leukemia, the
poorer the prognosis. Results indicated that, in the acute
monoblastic leukemia cell line THP-1, the Akt and ERK signal
transduction pathways were suppressed simultaneously, and that the
caspase-3 apoptosis signaling pathway was activated by the
downregulation of the Bcl-2/Bax ratio following DHA treatment. It
was speculated that DHA-mediated inhibition of Akt/ERK pathway
activation and the promotion of caspase-3 activation would result
in a more effective response to anti-leukemia therapy, as the
inhibitors would simultaneously target three pathways in THP-1
cells.
A number of specific inhibitors target a single
signaling molecule in the treatment of leukemia; however, DHA may
be more effective compared with common inhibitors as drug
resistance frequently emerges following the hyperactivation of
alternative signaling pathways under treatment of a single target.
It is predicted that DHA-resistance in AMoL may rarely occur, the
response to DHA may be increased and response duration may be
longer as DHA exhibited multi-targeting characteristics. Inhibition
of multiple signaling pathways increases the therapeutic ability of
DHA.
In the present study, the role of DHA in restricting
the proliferation and inducing apoptosis in AMoL cells was
investigated. DHA may directly or indirectly affect the activation
of Akt/ERK survival signaling and caspase-3 apoptosis signaling
pathways, all of which are key regulators of cell survival and
apoptosis, particularly during AMoL treatment (32,33).
However, additional in-depth investigations must follow this
preliminary study. First, the intermediate molecular mechanisms
underlying DHA-mediated changes to cell survival and apoptosis
signaling pathways must be clarified. Secondly, in vivo
experiments must be carried out to verify the treatment effects of
DHA. Finally, the results must be validated through clinical trials
and applications.
In conclusion, the present study indicated that DHA
exerted effective antitumor activities against AMoL in
vitro. Furthermore, DHA downregulated the expression of p-Akt
and p-ERK, whilst also upregulating the protein expression of
cleaved caspase-3 through increasing the Bcl-2/Bax ratio. This may
be one mechanism by which DHA exerts its effects in AMoL. However,
further and more comprehensive studies are required to confirm
this.
Acknowledgements
The present study was supported by grants from the
Shanghai Health Development Planning Commission (grant no.
ZY3-CCCX-3-3006), the National Natural Science Foundation of China
(grant nos. 81500392 and 31201010), the Shanghai Committee of
Science and Technology of China (grant nos. 12ZR1419500 and
114119a8700) and the Shanghai Health Bureau (grant no.
ZYSNXD-CCZDYJ029).
References
|
1
|
Ommen HB: Monitoring minimal residual
disease in acute myeloid leukaemia: A review of the current
evolving strategies. Ther Adv Hematol. 7:3–16. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peng H, Wang H, Xue P, Hou Y, Dong J, Zhou
T, Qu W, Peng S, Li J, Carmichael PL, et al: Suppression of
NRF2-ARE activity sensitizes chemotherapeutic agent-induced
cytotoxicity in human acute monocytic leukemia cells. Toxicol Appl
Pharmacol. 292:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guo Y, Shan Q, Gong Y, Lin J, Shi F, Shi R
and Yang X: Curcumin induces apoptosis via simultaneously targeting
AKT/mTOR and RAF/MEK/ERK survival signaling pathways in human
leukemia THP-1 cells. Pharmazie. 69:229–233. 2014.PubMed/NCBI
|
|
4
|
Ozpolat B, Akar U, Steiner M,
Zorrilla-Calancha I, Tirado-Gomez M, Colburn N, Danilenko M,
Kornblau S and Berestein GL: Programmed cell death-4 tumor
suppressor protein contributes to retinoic acid-induced terminal
granulocytic differentiation of human myeloid leukemia cells. Mol
Cancer Res. 5:95–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murashige N, Tabanda R and Zalusky R:
Occurrence of acute monocytic leukemia in a case of untreated
Waldenstrom's macroglobulinemia. Am J Hematol. 71:94–97. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cline MJ: Histiocytes and histiocytosis.
Blood. 84:2840–2853. 1994.PubMed/NCBI
|
|
7
|
Kumar RK, Basu S, Lemke HD, Jankowski J,
Kratz K, Lendlein A and Tetali SD: Effect of extracts of poly(ether
imide) microparticles on cytotoxicity, ROS generation and
proinflammatory effects on human monocytic (THP-1) cells. Clin
Hemorheol Microcirc. 61:667–680. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Y: Qinghaosu (artemisinin): Chemistry
and pharmacology. Acta Pharmacol Sin. 33:1141–1146. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin AJ, Klayman DL and Milhous WK:
Antimalarial activity of new water-soluble dihydroartemisinin
derivatives. J Med Chem. 30:2147–2150. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ashley EA, McGready R, Hutagalung R,
Phaiphun L, Slight T, Proux S, Thwai KL, Barends M, Looareesuwan S,
White NJ and Nosten F: A randomized, controlled study of a simple,
once-daily regimen of dihydroartemisinin-piperaquine for the
treatment of uncomplicated, multidrug-resistant falciparum malaria.
Clin Infect Dis. 41:425–432. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gordi T and Lepist EI: Artemisinin
derivatives: Toxic for laboratory animals, safe for humans? Toxicol
Lett. 147:99–107. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mi YJ, Geng GJ, Zou ZZ, Gao J, Luo XY, Liu
Y, Li N, Li CL, Chen YQ, Yu XY and Jiang J: Dihydroartemisinin
inhibits glucose uptake and cooperates with glycolysis inhibitor to
induce apoptosis in non-small cell lung carcinoma cells. PLoS One.
10:e01204262015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu W, Wang DW, Yu SY, Cao Y, Yang L, E
XQ, Yao GJ and Bi ZG: The effect of dihydroartemisinin on the
proliferation, metastasis and apoptosis of human osteosarcoma cells
and its mechanism. J Biol Regul Homeost Agents. 29:335–342.
2015.PubMed/NCBI
|
|
14
|
Feng X, Li L, Jiang H, Jiang K, Jin Y and
Zheng J: Dihydroartemisinin potentiates the anticancer effect of
cisplatin via mTOR inhibition in cisplatin-resistant ovarian cancer
cells: Involvement of apoptosis and autophagy. Biochem Biophys Res
Commun. 444:376–381. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park J, Lai HC, Singh M, Sasaki T and
Singh NP: Development of a dihydroartemisinin-resistant Molt-4
leukemia cell line. Anticancer Res. 34:2807–2810. 2014.PubMed/NCBI
|
|
16
|
Chan HW, Singh NP and Lai HC: Cytotoxicity
of dihydroartemisinin toward Molt-4 cells attenuated by
N-tert-butyl-alpha-phenylnitrone and deferoxamine. Anticancer Res.
33:4389–4393. 2013.PubMed/NCBI
|
|
17
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang D, Wang Q, Zhu T, Cao J, Zhang X,
Wang J, Wang X, Li Y, Shen B and Zhang J: RACK1 promotes the
proliferation of THP1 acute myeloid leukemia cells. Mol Cell
Biochem. 384:197–202. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zeng C, Wang W, Yu X, Yang L, Chen S and
Li Y: Pathways related to PMA-differentiated THP1 human monocytic
leukemia cells revealed by RNA-Seq. Sci China Life Sci.
58:1282–1287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Evans FJ and Hilton JH: Polymyositis
associated with acute Nonocytic leukemia: Case report and review of
the literature. Can Med Assoc J. 91:1272–1275. 1964.PubMed/NCBI
|
|
21
|
Wang Y, Ma L, Wang C, Sheng G, Feng L and
Yin C: Autocrine motility factor receptor promotes the
proliferation of human acute monocytic leukemia THP-1 cells. Int J
Mol Med. 36:627–632. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jain SK, Sahu R, Walker LA and Tekwani BL:
A parasite rescue and transformation assay for antileishmanial
screening against intracellular Leishmania donovani amastigotes in
THP1 human acute monocytic leukemia cell line. Journal of
visualized experiments. 70:e40542012.
|
|
23
|
Tu YY: TU You-you won Lasker Debakey
clinical medical research award-for her outstanding achievements in
studies on artemisinin. Zhongguo Zhong Xi Yi Jie He Za Zhi.
31:13012011.(In Chinese). PubMed/NCBI
|
|
24
|
Dong YJ, Li WD and Tu YY: Effect of
dihydro-qinghaosu on auto-antibody production, TNF alpha secretion
and pathologic change of lupus nephritis in BXSB mice. Zhongguo
Zhong Xi Yi Jie He Za Zhi. 23:676–679. 2003.(In Chinese).
PubMed/NCBI
|
|
25
|
Zhang CZ, Zhang H, Yun J, Chen GG and Lai
PB: Dihydroartemisinin exhibits antitumor activity toward
hepatocellular carcinoma in vitro and in vivo. Biochem Pharmacol.
83:1278–1289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng MX, Hong JX, Wang Q, Fan YY, Yuan CT,
Lei XH, Zhu M, Qin A, Chen HX and Hong D: Dihydroartemisinin
prevents breast cancer-induced osteolysis via inhibiting both
breast cancer cells and osteoclasts. Sci Rep. 6:190742016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Hu W, Zhang JL, Wu XH and Zhou HJ:
Dihydroartemisinin induces autophagy and inhibits the growth of
iron-loaded human myeloid leukemia K562 cells via ROS toxicity.
FEBS Open Bio. 2:103–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao X, Zhong H, Wang R, Liu D, Waxman S,
Zhao L and Jing Y: Dihydroartemisinin and its derivative induce
apoptosis in acute myeloid leukemia through Noxa-mediated pathway
requiring iron and endoperoxide moiety. Oncotarget. 6:5582–5596.
2015.PubMed/NCBI
|
|
29
|
Lim SG, Suk K and Lee WH: Reverse
signaling from LIGHT promotes pro-inflammatory responses in the
human monocytic leukemia cell line, THP-1. Cell Immunol. 285:10–17.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teng CL, Han SM, Wu WC, Hsueh CM, Tsai JR,
Hwang WL and Hsu SL: Mechanistic aspects of lauryl gallate-induced
differentiation and apoptosis in human acute myeloid leukemia
cells. Food Chem Toxicol. 71:197–206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi D, Xu Y, Du X, Chen X, Zhang X, Lou J,
Li M and Zhuo J: Co-treatment of THP-1 cells with naringenin and
curcumin induces cell cycle arrest and apoptosis via numerous
pathways. Mol Med Rep. 12:8223–8228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Olesen LH, Aggerholm A, Andersen BL,
Nyvold CG, Guldberg P, Nørgaard JM and Hokland P: Molecular typing
of adult acute myeloid leukaemia: Significance of translocations,
tandem duplications, methylation, and selective gene expression
profiling. Br J Haematol. 131:457–467. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ruvolo PP, Qiu Y, Coombes KR, Zhang N,
Neeley ES, Ruvolo VR, Hail N Jr, Borthakur G, Konopleva M, Andreeff
M and Kornblau SM: Phosphorylation of GSK3α/β correlates with
activation of AKT and is prognostic for poor overall survival in
acute myeloid leukemia patients. BBA Clin. 4:59–68. 2015.
View Article : Google Scholar : PubMed/NCBI
|