Introduction
Metastatic breast cancer is one of the most common
metastatic tumor types, and is a threat to female health globally
(1). Human breast carcinoma remains a
highly lethal disease due to local invasion and distant metastasis
at diagnosis, and the fact that >30% of patients with breast
carcinoma will develop metastasis during the development of their
disease (2,3). Although the potential function of
circulating tumor cell detection and monitoring in breast cancer
has been explored in clinical studies, the mortality and recurrence
rates remain high (4,5). A systematic review has summarized the
treatments currently available for breast cancer-associated
lymphedema, which provides potential anti-cancer strategies for
patients with breast cancer (6).
However, efficient clinical treatments for breast cancer are
required in order to improve the survival rate for patients.
Tunicamycin is a nucleotide antibiotic produced by
Streptomyces lysosuperficus. Numerous published reports have
indicated that Tunicamycin may be widely used to inhibit tumor cell
growth and aggressiveness (7,8). Evidence has revealed that Tunicamycin
may inhibit the growth of breast cancer cells through promoting
apoptosis (9,10). Hasegawa et al (11) suggested that Tunicamycin enhances the
apoptosis induced by TNF-related apoptosis-inducing ligand in
endometriotic stromal cells. The unfolded protein response is
required in the nu/nu mouse microvasculature when treating a breast
tumor with Tunicamycin, which supports the potential of Tunicamycin
to be a powerful glycotherapeutic treatment for breast cancer
(12). However, the underlying
anti-tumor mechanism mediated by Tunicamycin in breast cancer cells
has been poorly understood thus far.
In the present study, it was revealed that
Tunicamycin may be efficient for the treatment of breast cancer.
Research has demonstrated that Tunicamycin exerts anti-tumor
efficacy by inhibiting tumor cell growth and enhancing the
apoptosis of tumor cells (13,14). The
present study investigated the potential signaling mechanisms
mediated by Tunicamycin in breast cancer cells. Findings revealed
that Tunicamycin may be an efficient agent for the treatment of
breast cancer via regulation of the protein kinase B (Akt)/nuclear
factor-κB (NF-κB) signaling pathway.
Materials and methods
Ethical statement
The present study was performed according to the
recommendations in the Guide for the Care and Use of Laboratory
Animals of China (15). All animal
experiments were performed in accordance with the National
Institute of Health, and approved by the Committee on the Ethics of
The Third Affiliated Hospital of Kunming Medical University
(Kunming, China).
Cells and reagents
MCF-7 and SKBR-3 cell lines were purchased from the
American Type Culture Collection (Manassas, VA, USA). All tumor
cells were cultured in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with
10% fetal bovine serum (FBS; Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). All cells were cultured in a 37°C
humidified atmosphere of 5% CO2.
Flow cytometry
MCF-7 (1×106) and SKBR-3
(1×106) cells were cultured in 6-well plates and treated
with Tunicamycin (2 mg/ml) or phosphate-buffered saline (PBS) for
12 h at 37°C. Apoptosis of MCF-7 and SKBR-3 cells was evaluated
using an Annexin V-fluorescein isothiocyanate (FITC) and propidium
iodide (PI) apoptosis detection kit (BD Biosciences, San Jose, CA,
USA). MCF-7 and SKBR-3 cells were isolated from Tunicamycin- or
PBS-treated mice and treated with an Annexin V-FITC and PI kit,
according to the manufacturer protocol. Fluorescence was measured
with a FACScan flow cytometer (BD Biosciences) and analyzed using
FCS Express™ IVD software (version 4; De Novo Software, Los
Angeles, CA, USA).
Endogenous expression of Akt
MCF-7 and SKBR-3 cells were cultured to 90%
confluency, following which the media was removed. MCF-7 and SKBR-3
cells were transfected with lentivirus-AKT (p-AKT) or
lentivirus-vector (Control) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). MCF-7 and SKBR-3
cells with stable overexpression of Akt (OPAKT) were treated using
Tunicamycin (5 mg/ml) for 24 h at 37°C to allow analysis of the
protein expression via western blotting, as subsequently
detailed.
MTT assays
MCF-7, SKBR-3 or Akt-overexpressing MCF-7 or SKBR-3
cells (1×106 of each) using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) were cultured in
96-well plates for 48 h at 37°C. Growing MCF-7 and SKBR-3 cells
were treated using 3, 5 or 8 mg/ml Tunicamycin (Sigma-Aldrich;
Merck KGaA) for 48 or 72 h at 37°C. Following 48 h of incubation,
20 µl MTT (5 mg/ml) in PBS solution was added to each well, and the
plate was further incubated for 4 h at 37°C. The majority of the
medium was removed and 100 µl DMSO was added into the wells to
solubilize the crystals. The optical density was measured using a
Bio-Rad (ELISA) reader (Bio-Rad Laboratories, Inc., Hercules, CA,
USA) at a wavelength of 450 nm.
Cell migration and invasion
assays
Stable Akt-overexpressing MCF-7 and SKBR-3 cells
were cultured with Tunicamycin (5 mg/ml) or PBS for 48 h at 37°C.
For migration assays, cells (1×106) suspended in
FBS-free medium were plated in the upper wells of 24-well
polycarbonate Transwell inserts (EMD Millipore, Billerica, MA,
USA). DMEM supplemented with 10% FBS was added to the lower wells.
Following incubation for 24 h, cells on the upper surface of the
inserts were scraped off, and cells on the lower surface were fixed
with formaldehyde for 5 min at 37°C, stained with 4% crystal violet
for 15 min at 37°C and counted under a light microscope (Olympus
BX51; Olympus Corporation, Tokyo, Japan). Cell invasion assays were
performed using a Cell Invasion Assay kit (ECM550; EMD Millipore),
according to the manufacturer's protocol.
Analysis of the cell cycle
To analyze the effects of Tunicamycin (5 mg/ml) on
the cell cycle stage of MCF-7 or SKBR-3 cells, flow cytometry was
performed. MCF-7 or SKBR-3 cells in the exponential phase of
growth, or Akt-overexpressing MCF-7 or SKBR-3 cells, were treated
with Tunicamycin (5 mg/ml) for 48 h at 37°C. Cells were then
washed, trypsinized for 5 min at 37°C, and rinsed with PBS. All
cells were fixed in 75% ice-cold ethanol for 15 min and then washed
with PBS three times. The fixed cells were washed with RNaseA (20
µg ml/l; Fermentas; Thermo Fisher Scientific, Inc.) and stained
using PI (20 µg ml/l; Sigma-Aldrich; Merck KGaA) for 10 min at
37°C. The percentages of cells in the G1/G1 and S phase were
analyzed using a BD FACSCalibur flow cytometer (BD Biosciences).
Cells were analyzed using the FCS Express IVD software (version
4).
Western blotting
MCF-7 or SKBR-3 cells were treated using Tunicamycin
(5 mg/ml), harvested by scraping and lysed using
radioimmunoprecipitation assay lysis buffer (Sigma-Aldrich; Merck
KGaA) at 37°C for 2 h, followed by homogenization at 4°C for 10
min. Protein concentration was measured using a BCA protein assay
kit (Thermo Fisher Scientific, Inc.). Proteins (20 µg) were
analyzed via SDS-PAGE, and then transferred onto a polyvinylidene
difluoride membrane (EMD Millipore). Following blocking with 5%
bull serum albumin in PBS (Sigma-Aldrich; Merck KGaA) at 37°C for 2
h, proteins were incubated with rabbit anti-human Akt (dilution,
1:500; cat no. ab38449), NF-κB (dilution, 1:400; cat no. ab7204) or
β-actin (dilution, 1:500; cat no. ab32572; all Abcam, Cambridge,
UK) antibodies for 12 h at 4°C. A horseradish peroxidase-conjugated
goat anti-rabbit IgG (cat no. PV-6001; ZSGB-BIO, Beijing, China)
was used at a dilution factor of 1:5,000 and used for analyzing
protein expression (Olympus BX51, Olympus; Tokyo, Japan). The
results were visualized using a chemiluminescence detection system
(GE Healthcare Life Sciences, Chicago, IL, USA). The density of the
bands was analyzed by Quantity One software version 4.62 (Bio-Rad
Laboratories, Inc.) detected using a Western Blotting Luminol
Reagent (GE Healthcare Life Sciences, Chicago, IL, USA).
Animal studies
A total of 80 specific pathogen-free male Balb/c
(6–8 weeks old; body weight, 25–32 g) mice were purchased from
Slack Co., Ltd. (Shanghai, China, http://qiye20447702.xinlimaoyi.com/). Mice were
maintained in a 12/12 h light/dark cycle with ad libitum
food and water. Each mouse was housed individually. Experimental
mice were implanted with either MCF-7 cells (5×107) or
SKBR-3 cells (5×107) in the groin, and divided into two
groups (n=40 in each group). Treatments were initiated on the fifth
day following tumor implantation (diameter: 5–8 mm). Tumor-bearing
mice were intravenously injected with Tunicamycin (5 mg/kg), or
with PBS as the control. The treatment was performed once a day for
a total of 10 days. The tumor volumes were calculated as detailed
in a previous study (16). Mice were
sacrificed once the tumor diameter had reached 18 mm.
Terminal deoxynucleotidyl
transferase-mediated dUTP nick-end labeling (TUNEL) analysis
For the analysis of myocardial cell apoptosis, a
TUNEL assay (Bimake, Houston, TX, USA) was used to detect
TUNEL-positive cells. Tumor cells sourced from experimental mice
following treatment with Tunicamycin or PBS were fixed using 4%
paraformaldehyde solution for 60 min at 4°C. The tumor sections
were washed with PBS three times, and then permeabilized by
immersing cells slides in 0.2% Triton X-100 solution in PBS for 30
min at 4°C. Subsequently, cells were incubated with equilibration
buffer (Sigma-Aldrich; Merck KGaA) for 30 min at 4°C. The tumor
sections were then incubated with 50 µl of the reaction mixture at
37°C for 60 min, and washed 3 times with PBS. The cells nuclei were
stained with 4′,6-diamidino-2-phenylindole for 60 min at 4°C.
Finally, tumor tissue images were captured with a ZEISS LSM 510
confocal microscope at 488 nm (magnification, ×40).
Statistical analysis
Software used for statistical analysis included the
SPSS 19.0 statistical package (IBM Corp., Armonk, NY, USA) and
GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). Data
were analyzed using an unpaired Student's t-test, and comparisons
between the data of multiple groups were performed using one-way
analysis of variance, followed by Dunnett's test. Data are
presented as the mean ± the standard error of the mean obtained
from three independent experiments. P<0.05 was considered to
indicate a statistically significant difference.
Results
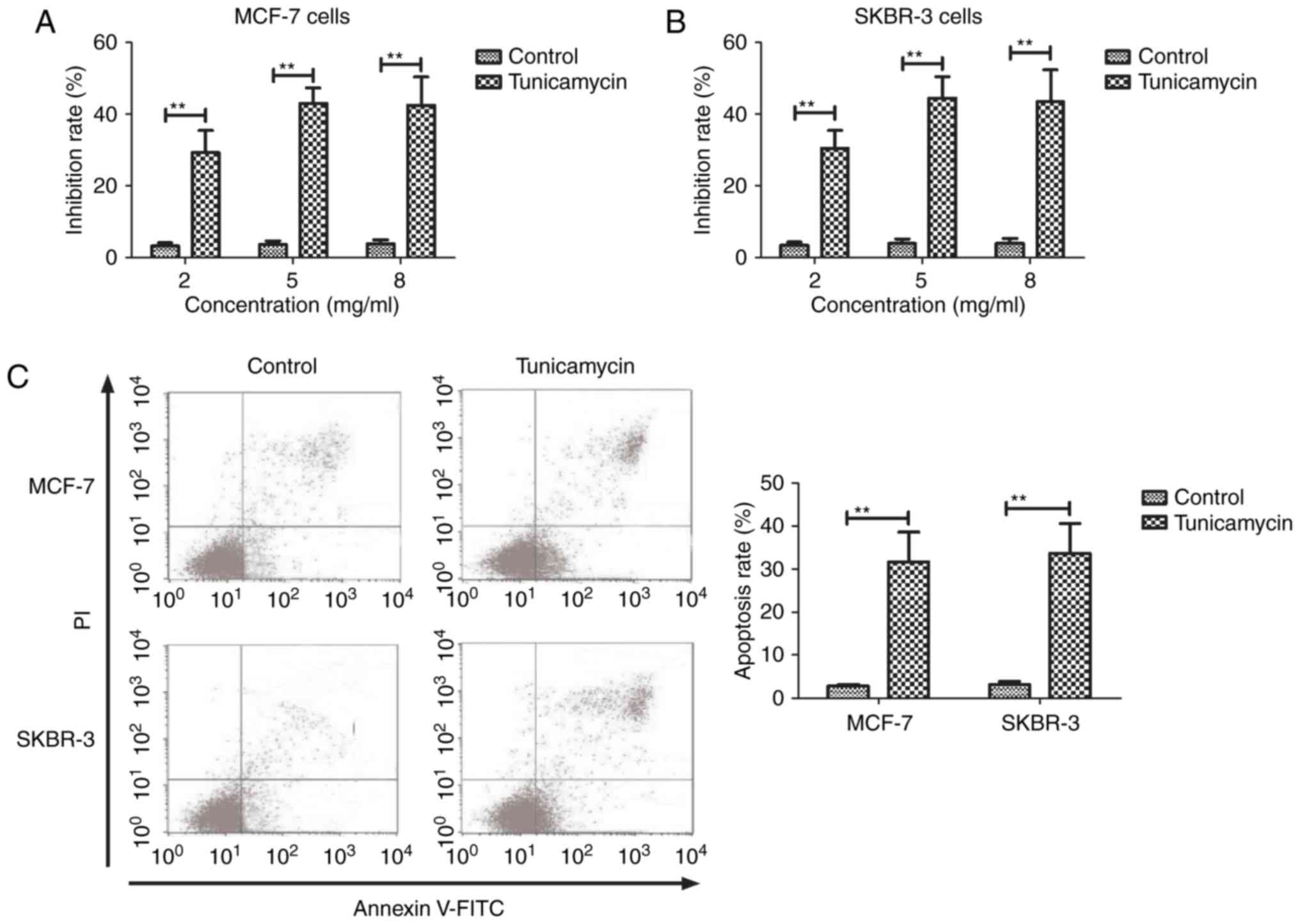
Tunicamycin treatment inhibits the
growth of and promotes the apoptosis of breast cancer cells
The inhibitory effects of Tunicamycin on breast
cancer cells were investigated in the present study. As presented
in Fig. 1A and B, Tunicamycin
significantly inhibited the growth of MCF-7 and SKBR-3 cells in a
dose-dependent manner (2, 5 and 8 mg/ml), compared with the control
cells (P<0.01). A dose of 5 mg/ml maximally inhibited the growth
of MCF-7 and SKBR-3 cells. It was revealed that Tunicamycin
significantly induced the apoptosis of MCF-7 and SKBR-3 cells in
vitro compared with control cells (P<0.01; Fig. 1C). These data suggest that Tunicamycin
treatment may inhibit the growth and promote the apoptosis of
breast cancer cells.
Tunicamycin treatment inhibits the
aggressiveness of and arrests the cell cycle of breast cancer
cells
The efficacy of Tunicamycin on the aggressiveness of
breast cancer cells was analyzed. As presented in Fig. 2A and B, it was observed that
Tunicamycin (5 mg/ml) significantly inhibited the migration and
invasion of MCF-7 and SKBR-3 cells after 48 h of incubation, as
compared with the control cells (P<0.01). Representative flow
cytometry images revealed that Tunicamycin treatment affected the
cell cycle of MCF-7 and SKBR-3 cells (Fig. 2C). It was additionally revealed that
Tunicamycin significantly arrested the cell cycle of MCF-7 and
SKBR-3 cells at the G1/G2 and S phase, compared with the control
cells (P<0.01; Fig. 2D and E).
These results suggest that Tunicamycin treatment may inhibit the
aggressiveness of and arrest the cell cycle of breast cancer
cells.
Tunicamycin regulates the growth and
aggressiveness of breast cancer cells through the Akt/NF-κB
signaling pathway
The potential molecular mechanism mediated by
Tunicamycin and involved in the inhibition of breast cancer cell
growth and aggressiveness was further analyzed. As presented in
Fig. 3A, Tunicamycin treatment
suppressed Akt and NF-κB expression in MCF-7 and SKBR-3 cells,
relative to in the control cells. OPAKT increased NF-κB expression
and cancelled out the effect of Tunicamycin-inhibited (OPAKT/TC)
NF-κB expression in MCF-7 and SKBR-3 cells (Fig. 3B). Overexpression of Akt significantly
cancelled out the effect of Tunicamycin-inhibited growth on MCF-7
and SKBR-3 cells compared with cells treated with Tunicamycin
(P<0.01; Fig. 3C).
Tunicamycin-inhibited migration and invasion in MCF-7 and SKBR-3
cells was also significantly abolished by the overexpression of
Akt, compared with in cells treated Tunicamycin (P<0.01;
Fig. 3D and E). These results suggest
that Tunicamycin regulates the growth and aggressiveness of breast
cancer cells through the Akt/NF-κB signaling pathway.
In vivo efficacy of Tunicamycin in
tumor-bearing mice
The in vivo effects of Tunicamycin were
investigated in MCF-7- and SKBR-3-bearing mice. As presented in
Fig. 4A and B, Tunicamycin treatment
(5 mg/kg) significantly inhibited MCF-7 (P<0.01), and
non-significantly inhibited SKBR-3 tumor growth compared with the
control. It was additionally observed that Tunicamycin treatment
significantly increased the number of apoptotic cells in MCF-7 and
SKBR-3 tumor sections compared with tumor sections from PBS-treated
mice (P<0.01; Fig. 4C and D).
Observations revealed that Tunicamycin treatment significantly
prolonged the survival of MCF-7- and SKBR-3-bearing mice, compared
with PBS-treated mice (P<0.01; Fig. 4E
and F). Western blot analysis revealed that Tunicamycin
downregulated Akt expression and NF-κB protein expression in MCF-7
and SKBR-3 tumor sections (Fig. 4G and
H). These results suggest that Tunicamycin treatment may
significantly inhibit breast cancer growth in vivo.
Discussion
Breast cancer is one of the most common female
malignant tumor types to occur in the mammary gland epithelial
tissue (17). Tunicamycin has been
widely used to enhance the antitumor activity of trastuzumab on
breast cancer in vitro and in vivo (18). In the present study, the efficacy and
potential mechanism of Tunicamycin against breast cancer growth,
apoptosis and aggressiveness in vitro and in vivo
were explored. Results revealed that Tunicamycin treatment
significantly inhibited the growth, migration and invasion of MCF-7
and SKBR-3 cells. In addition, Tunicamycin treatment induced the
apoptosis of MCF-7 and SKBR-3 cells, and arrested the cell cycle.
Notably, it was observed that Tunicamycin treatment decreased Akt
and NF-κB expression, whilst the overexpression of Akt cancelled
out the effect of Tunicamycin-inhibited cell growth and
aggressiveness, suggesting that Tunicamycin regulates cell growth
and aggressiveness through the Akt/NF-κB signaling pathway.
A previous study had demonstrated that
Tunicamycin-induced ER stress may regulate C-C motif chemokine
ligand 5 expression and secretion via signal transducer and
activator of transcription 3, followed by the decreased
transmigration of MCF-7 breast cancer cells (19). The results of the present study
indicate that Tunicamycin inhibited growth and aggressiveness
through the Akt/NF-κB signaling pathway. Kim et al (20) demonstrated that Tunicamycin may induce
paraptosis potentiated by inhibition of the BRAFV600E
mutation in the FRO anaplastic thyroid carcinoma cell line.
However, it was revealed in the present study that Tunicamycin may
induce apoptosis and arrest the cell cycle at the G1/G1 and S phase
in MCF-7 and SKBR-3 cells. Han et al (18) indicated that Tunicamycin enhances the
antitumor activity of trastuzumab against breast cancer in
vitro and in vivo, and the results revealed that
combinations of trastuzumab with the N-glycosylation inhibitor
Tunicamycin may be a promising approach for improving the clinical
efficacy of trastuzumab. Furthermore, Tunicamycin-induced
endoplasmic reticulum stress resulted in reduction of the in
vitro subpopulation and invasion of
CD44+/CD24− phenotype breast cancer stem
cells (21). Findings of the present
study indicate that Tunicamycin inhibits the growth of breast
cancer cells through increasing the number of apoptotic bodies in
tumor tissues in tumor-bearing mice.
A systematic review and new research directions have
highlighted the association between cell migration and female
breast cancer across various ethnicities (22). In the present study, the potential
signaling pathway underlying the Tunicamycin-mediated growth and
aggressiveness of MCF-7 and SKBR-3 cells was explored. Findings
indicated that Tunicamycin is an efficient anti-cancer agent,
suggesting that the Akt/NF-κB signaling pathway may be a potential
target for the treatment of breast cancer.
References
|
1
|
Benson R, Madan R, Julka PK and Rath GK:
Metaplastic carcinoma of breast: A case series of seven patients
from a tertiary care center and review of literature. Gulf J
Oncolog. 1:74–76. 2016.PubMed/NCBI
|
|
2
|
Conlon N, Sadri N, Corben AD and Tan LK:
Acinic cell carcinoma of breast: Morphologic and
immunohistochemical review of a rare breast cancer subtype. Hum
Pathol. 51:16–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Conlon N, Howard J, Catalano J, Gallagher
M, Tan LK and Corben AD: Breast carcinoma in young women: No
evidence of increasing rates of metastatic breast carcinoma in a
single tertiary center review. Breast J. 22:287–292. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang L, Shi JF, Huang HY, Zhu J, Li J,
Fang Y and Dai M: Economic evaluation on breast cancer screening in
mainland China: A systematic review. Zhonghua Liu Xing Bing Xue Za
Zhi. 37:1662–1669. 2016.(In Chinese). PubMed/NCBI
|
|
5
|
Banys-Paluchowski M, Krawczyk N and Fehm
T: Potential role of circulating tumor cell detection and
monitoring in breast cancer: A review of current evidence. Front
Oncol. 6:2552016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wei JL, Buza J III and Liu CJ: Does
progranulin account for the opposite effects of etanercept and
infliximab/adalimumab in osteoarthritis?: Comment on Olson et al:
‘Therapeutic opportunities to prevent post-traumatic arthritis:
Lessons from the natural history of arthritis after articular
fracture’. J Orthop Res. 34:12–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang CC, Chen LH, Gillespie S, Kiejda KA,
Mhaidat N, Wang YF, Thorne R, Zhang XD and Hersey P: Tunicamycin
sensitizes human melanoma cells to tumor necrosis factor-related
apoptosis-inducing ligand-induced apoptosis by up-regulation of
TRAIL-R2 via the unfolded protein response. Cancer Res.
67:5880–5888. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shiraishi T, Yoshida T, Nakata S, Horinaka
M, Wakada M, Mizutani Y, Miki T and Sakai T: Tunicamycin enhances
tumor necrosis factor-related apoptosis-inducing ligand-induced
apoptosis in human prostate cancer cells. Cancer Res. 65:6364–6370.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Delom F, Emadali A, Cocolakis E, Lebrun
JJ, Nantel A and Chevet E: Calnexin-dependent regulation of
tunicamycin-induced apoptosis in breast carcinoma MCF-7 cells. Cell
Death Differ. 14:586–596. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takahashi N, Iwahori A, Breitman TR and
Fukui T: Tunicamycin in combination with retinoic acid
synergistically inhibits cell growth while decreasing
palmitoylation and enhancing retinoylation of proteins in the human
breast cancer cell line MCF-7. Oncol Res. 9:527–533.
1997.PubMed/NCBI
|
|
11
|
Hasegawa A, Osuga Y, Hirota Y, Hamasaki K,
Kodama A, Harada M, Tajima T, Takemura Y, Hirata T, Yoshino O, et
al: Tunicamycin enhances the apoptosis induced by tumor necrosis
factor-related apoptosis-inducing ligand in endometriotic stromal
cells. Hum Reprod. 24:408–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Banerjee A, Lang JY, Hung MC, Sengupta K,
Banerjee SK, Baksi K and Banerjee DK: Unfolded protein response is
required in nu/nu mice microvasculature for treating breast tumor
with tunicamycin. J Biol Chem. 286:29127–29138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Giordano E, Davalos A, Nicod N and Visioli
F: Hydroxytyrosol attenuates tunicamycin-induced endoplasmic
reticulum stress in human hepatocarcinoma cells. Mol Nutr Food Res.
58:954–962. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carlisle RE, Brimble E, Werner KE, Cruz
GL, Ask K, Ingram AJ and Dickhout JG: 4-Phenylbutyrate inhibits
tunicamycin-induced acute kidney injury via CHOP/GADD153
repression. PLoS One. 9:e846632014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davey G and Wu Z: Attitudes in China
toward the use of animals in laboratory research. Altern Lab Anim.
35:313–316. 2007.PubMed/NCBI
|
|
16
|
Bai FL, Yu YH, Tian H, Ren GP, Wang H,
Zhou B, Han XH, Yu QZ and Li DS: Genetically engineered Newcastle
disease virus expressing interleukin-2 and TNF-related
apoptosis-inducing ligand for cancer therapy. Cancer Biol Ther.
15:1226–1238. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li L, Yuan L, Chen X, Wang Q, Tian J, Yang
K and Zhou E: Current treatments for breast cancer-related
lymphoedema: A systematic review. Asian Pac J Cancer Prev.
17:4875–4883. 2016.PubMed/NCBI
|
|
18
|
Han X, Zhang X, Li H, Huang S, Zhang S,
Wang F and Shi Y: Tunicamycin enhances the antitumor activity of
trastuzumab on breast cancer in vitro and in vivo. Oncotarget.
6:38912–38925. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Liao S, Fan W, Wei W, Wang C and
Sun S: Tunicamycin-induced ER stress regulates chemokine CCL5
expression and secretion via STAT3 followed by decreased
transmigration of MCF-7 breast cancer cells. Oncol Rep.
32:2769–2776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim SH, Shin HY, Kim YS, Kang JG, Kim CS,
Ihm SH, Choi MG, Yoo HJ and Lee SJ: Tunicamycin induces paraptosis
potentiated by inhibition of BRAFV600E in FRO anaplastic thyroid
carcinoma cells. Anticancer Res. 34:4857–4868. 2014.PubMed/NCBI
|
|
21
|
Nami B, Donmez H and Kocak N:
Tunicamycin-induced endoplasmic reticulum stress reduces in vitro
subpopulation and invasion of CD44+/CD24-phenotype breast cancer
stem cells. Exp Toxicol Pathol. 68:419–426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Andreeva VA, Unger JB and Pentz MA: Breast
cancer among immigrants: A systematic review and new research
directions. J Immigr Minor Health. 9:307–322. 2007. View Article : Google Scholar : PubMed/NCBI
|