Introduction
Cisplatin (DDP) is widely used in treating ovarian
cancer and has demonstrated a marked therapeutic effect in its
clinical application. However, during the treatment progress, the
generation of DDP resistance has become an obstacle to successful
treatment (1,2). It is well-known that when DDP is
administered to patients, certain biological phenomena will
inevitably occur, such as a gradual reduction in intracellular drug
accumulation, inactivation of the general desired effects of the
treatment, improvements in the DNA repair ability of cancer cells,
alterations of the apoptosis pathway of the tumor cells, and
certain regulatory actions of activated indirect signaling
pathways, which directly results in the generation of DDP
resistance. These may increase the incidence or severity of these
events, even causing complete treatment failure (3–5). One
strategy to overcome resistance to DDP is to develop novel
chemotherapeutic drugs with less or no side effects, which has
become an important topic in the field of cancer biology,
particularly in ovarian cancer.
Previous studies examining the structure-activity
associations of natural isoflavones with antitumor responses have
demonstrated that benzene rings with hydroxyls in the seventh or
fifth carbon atom in the primary chain (C-7 or C-5), or with three
hydroxyls in whole chemical structure, possess higher rates of
pharmacological activities (6,7). The
hydroxyl of C-5 in isoflavones is the necessary chemical group that
retains the antitumor activity (8).
Isoflavones in that the hydroxyl of C-4′ is superseded by an
NO2 group will exhibit increased antitumor bioactivity
in comparison to the original isoflavones (9).
5-hydroxy-4′-nitro-7-propionyloxy-genistein (HNPG) is a novel
synthetic isoflavone derivative that possesses a nitro group in
C-4′, a hydroxyl group at C-5 and a propionyloxy group at C-7, and
it was observed that HNPG exhibited an inhibition of proliferation
in gastric and breast cancer via an MTT assay and a
5-acetylene-2′-deoxypyrimidine nucleoside staining method (10) in vitro, but its antitumor
effect has not been examined by other experimental forms, and its
molecular biological mechanism has not been investigated (11,12), its
role in ovarian cancer also remains uncharacterized.
In the present study, the anticancer effects of HNPG
in A2780/DDP cells were evaluated in vitro, and the results
demonstrated that HNPG may inhibit proliferation, clone formation,
invasion and metastasis and induce apoptosis in A2780/DDP cells,
which may attribute to its ability to increase the accumulation of
reactive oxygen species (ROS) in cells, decrease the mitochondrial
membrane potential (Δψm), regulate the ratio of B-cell lymphoma 2
(Bcl-2)/Bcl-2-associated X protein (Bax) and release Cytochrome C
(Cyt-C) from the mitochondria, triggering a caspase cascade and
inducing apoptosis, which not only demonstrated more researching
patterns of the antitumor effects of HNPG, but also highlighted the
basic molecular biological mechanism of HNPG compared with previous
reports (11,12).
Materials and methods
Reagents
HNPG was synthesized in the Department of Obstetrics
and Gynecology, The First Clinical School of Jinan University
(Guangzhou, China) as described previously (12), with minor modifications; this reagent
is a pale yellow crystalline powder with a molecular formula
C18H13O7N and a molecular weight
of 355, and its chemical structure is presented in Fig 1. HNPG was dissolved in dimethyl
sulfoxide (DMSO) into a 1 mM stock solution and stored until use,
and the maximal concentration of DMSO did not exceed 0.1% (v/v), as
this concentration does not exhibit an inhibitory effect on cell
proliferation. DMSO, PBS, MTT and Giemsa, crystal violet and
hematoxylin & eosin (H&E) stains were all obtained from
Beyotime Institute of Biotechnology, Co., Ltd. (Shanghai, China).
Matrigel™ was purchased from Shanghai Invitrogen
Biological Technology Co., Ltd. (Shanghai, China). Specific
antibodies for rabbit anti-Bcl-2 (cat. no. A0040-1; dilution,
1:1,000), rabbit anti-Bax (cat. no. A00183; dilution, 1:1,000),
rabbit anti-Cyt-C (cat. no. A03529; dilution, 1:1,000), rabbit
anti-cleaved caspase-9 (cat. no. BM4521; dilution, 1:1,000), rabbit
anti-cleaved caspase-3 (cat. no. BM3937; dilution, 1:1,000), rabbit
anti-GAPDH antibodies (cat. no. A00227-1; dilution, 1:1,000) and
horseradish peroxidase-conjugated secondary rabbit antibody used
for western blotting (cat. no. BA1082; dilution, 1:5,000) were all
obtained from Boster Biological Technology Co., Ltd. (Wuhan,
China).
Cell culture and grouping
A2780-cisR cell line was purchased from the China
Centre for Type Culture Collection (Wuhan, Hubei, China) and
cultured at 37°C in humidified 5% CO2 in Dulbecco's
modified Eagle's medium (DMEM; Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
Hangzhou, China), penicillin (100 U/ml, Qilu Pharmaceutical Co.,
Ltd., Jinan, China) and streptomycin (100 µg/ml, Shandong
Pharmaceutical Co., Ltd., Jinan, Shandong, China). The cells were
then divided in four groups: The control group (0.1% DMSO) and the
different concentrations of HNPG (5, 10 and 20 µM) groups.
Cell proliferation assay
A2780/DDP cells were seeded on a 96-well plate
(Beyotime Institute of Biotechnology Co., Ltd., Shanghai, China) at
a density of 1×104 cells/well and incubated with
different concentrations of HNPG of 0.625, 1.25, 2.5, 5, 10, 20,
40, 80 and 160 µM for 24 h at 37°C in a humidified atmosphere
containing 5% CO2, which concentration intervals benefit
to screen the most sensitive range of pharmacological activities.
Subsequently, 20 µl 5 mg/ml MTT stock solution was added to each
well, and an additional incubation was performed for 6 h at 37°C. A
total of 100 µl DMSO was added to each well to halt the reaction,
and spectrophotometric absorbance was subsequently measured using a
microplate reader (ELX-800; BioTek China, Shanghai, China) at 570
nm (A570). The proliferative inhibition graph was made
based on the rate of proliferation inhibition calculated according
to the value of spectrophotometric absorbance. The three most
sensitive concentrations of HNPG were selected, then 0.1% DMSO and
different concentrations of HNPG (5, 10 and 20 µM) were introduced
into each well and sequentially A2780/DDP cells were cultured with
complete DMEM medium for 24, 48 and 72 h in humidified 5%
CO2 at 37°C. The rate of proliferation inhibition (IR)
was calculated as follows: (1-average A570 of the
experimental group/average A570 of the control group)
×100%. Experiments were performed in triplicate, and the mean value
was calculated.
Flat plate clone formation assay
A2780/DDP cells were collected and seeded onto a
6-well plate (Beyotime Institute of Biotechnology Co., Ltd.,
Shanghai, China) at a density of 3×103 cells/well and
incubated for 24 h at 37°C in a humidified atmosphere containing 5%
CO2. Then, 0.1% DMSO or different concentrations of HNPG
(5, 10 or 20 µM) were added to each well, and the cells were
continuously cultured for 48 h. Subsequently, the drug-containing
medium was removed, and the cells were washed twice with PBS.
Complete DMEM medium was then added, and the cells were cultured
for 7 days at 37°C in humidified 5% CO2 until visible
clones formed. Clones containing >50 cells were defined as one
clone, and were fixed with 95% methanol for 10 min at room
temperature and stained with 0.1% Giemsa stain for 10 min at room
temperature. Individually-stained clones in each well were counted
and the clone formation rate was calculated: The clone formation
inhibition rate (%) was calculated as follows: 1-(the mean number
of HNPG group/the mean number of control group) ×100%. The results
were representative of three independent experiments.
Assessment of the invasive
capacity
A pre-cooled 24-well Transwell plate (Beyotime
Institute of Biotechnology Co., Ltd., Shanghai, China) was covered
with 30 µl Matrigel™ at a 1:3 dilution and incubated at
37°C for 3 h. Then, A2780/DDP cells (1×105/well) were
cultured at 37°C for 20 h in the inner chamber and exposed to 100
µl 0.1% FBS/DMEM with 0.1% DMSO or different doses of HNPG (5, 10
or 20 µM), and the outer chamber was filled with 500 µl 10%
FBS/DMEM to act as a chemoattractant. Subsequent the membranes of
Transwell plate were washed with PBS three times and the invasive
cells were fixed with 95% ethanol for 15 min at room temperature,
and then stained with 0.5% H&E staining for 15 min at room
temperature. The number of invasive cells was manually counted in 5
randomly selected fields under an inverted microscope (N-STORM 4.0;
Nikon Corporation, Tokyo, Japan) with magnification, ×200. Every
group had three repeat wells and the average value was
calculated.
Examination of the metastasizing
ability
A2780/DDP cells (1×105/well) were seeded
into the inner chamber of 24-well Transwell plate, treated with 100
µl 0.1% FBS/DMEM containing 0.1% DMSO or different doses of HNPG
(5, 10 or 20 µM). The inner chamber was placed into the outer
chamber containing 500 µl 10% FBS/DMEM, and cultured for 16 h at
37°C. Following this, the membranes of the Transwell plate were
washed with PBS three times, and the metastasized cells were fixed
with 4% paraformaldehyde for 15 min at room temperature and
subsequently stained with 0.1% crystal violet stain for 15 min at
room temperature. The number of metastasized cells was manually
calculated in 5 randomly selected fields under inverted microscope
with magnification, ×200. Each group had three repeated wells and
the mean value was calculated.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) stain apoptosis assay
Groups of A2780/DDP cells were treated with 0.1%
DMSO or different concentrations of HNPG (5, 10 or 20 µM) for 48 h,
then washed with PBS twice, digested with 0.25% trypsin,
centrifuged 300 × g for 5 min at room temperature, and the
supernatant was discarded. Subsequently, the cells were stained
with PI (50 mg/ml) and Annexin V-FITC (25 mg/ml) solution (Beyotime
Institute of Biotechnology Co., Ltd., Shanghai, China) at room
temperature for 15 min in the dark, and then analyzed within 1 h
using flow cytometry. Excitation and emission wavelengths of 488
and 530 nm, respectively, were selected, and the fraction of the
cell populations in different quadrants was analyzed using quadrant
statistics. Experiments were performed in triplicate, and the mean
value was calculated.
Evaluation of ROS
A2780/DDP cells were treated with 0.1% DMSO or
different concentrations of HNPG (5, 10 or 20 µM) for 48 h, washed
by PBS twice and digested with 0.25% trypsin, centrifuged 300 × g
for 5 min at room temperature, and the supernatant was discarded.
The cells were then resuspended using 5 mM
2′,7′-dichlorodihydro-fluorescein diacetate (DCFH-DA; Bioluminor
Biotechnology Co., Ltd., Xiamen, Fujian, China), incubated for 30
min at 37°C in the dark and washed with serum-free DMEM medium
three times. Subsequently, the samples were analyzed by flow
cytometry using excitation and emission wavelengths of 488 and 530
nm, respectively. The independent experiments were repeated three
times, and the average value was calculated.
Measurement of Δψm
A2780/DDP cells were exposed to 0.1% DMSO or
different concentrations of HNPG (5, 10, 20 µM) for 48 h, washed
twice with cold PBS, digested with 0.25% trypsin, centrifuged 300 ×
g for 5 min at room temperature, and the supernatant was discarded,
resuspended in 10 mM lipophilic cationic dye
2-(6-Amino-3-imino-3H-xanthen-9-yl) benzoic acid methyl ester
(Rh123; Yeasen Biological Technology Co., Ltd., Shanghai, China)
for 30 min at 37°C in the dark, and then analyzed by flow
cytometry. Excitation and emission wavelengths were 475 and 525 nm,
respectively. Experiments were performed in triplicate, and the
mean value was calculated.
Western blot analysis
A2780/DDP cells that were treated with 0.1% DMSO or
different concentrations of HNPG (5, 10 and 20 µM) for 48 h were
washed with cold PBS twice and lysed in 5% lysis buffer (Beyotime
Institute of Biotechnology Co., Ltd., Shanghai, China). The cell
lysate was incubated on ice for 15 min and then centrifuged at
20,000 × g for 30 min at 4°C. The amount of total cell protein was
determined by BCA kit (Beyotime Institute of Biotechnology Co.,
Ltd., Shanghai, China). Protein aliquots (50 µg) were separated by
12% SDS-PAGE and transferred to nitrocellulose membranes.
Non-specific binding sites were blocked by incubating the
nitrocellulose membrane for 1 h at 37°C with 5% non-fat dried milk
in TBS containing 0.05% Tween-20 (TBST). The membranes were
incubated for 3 h at 37°C with primary antibodies (rabbit
anti-Bcl-2, rabbit anti-Bax, rabbit anti-Cyt-C, rabbit anti-cleaved
caspase-9, rabbit anti-cleaved caspase-3 and rabbit anti-GAPDH
antibodies, then washed using TBST buffer three times and 15
minutes every time and incubated with a horseradish
peroxidase-conjugated secondary rabbit antibody for 2 h at 37°C.
The dilutions of all the primary antibodies were 1:1,000, and the
dilution of horseradish peroxidase-conjugated secondary rabbit
antibody was 1:5,000. Bands were visualized using an enhanced
chemiluminescence kit (cat no. AR1170; Boster Biological Technology
Co., Ltd., Wuhan, China) and analyzed using Image J (version 1.8.0;
National Institutes of Health, Bethesda, MD, USA). Each experiment
was repeated three times and the mean value was obtained.
Statistical analysis
SPSS 15.0 software package (SPSS Inc., Chicago, IL,
USA) was used for analysis. Data are presented as the mean ±
standard deviation. The means of multiple groups were compared with
one-way analysis of variance, after analyzing means using the test
for homogeneity of variance, the comparisons among the means were
performed using Least-significant Difference, Student-Newman-Keuls
and Bonferroni. Multiple post-hoc tests were used in order to
demonstrate the statistical significance from multiple-aspect.
P<0.05 was considered to indicate a statistically significant
difference.
Results
HNPG inhibits A2780/DDP cell
proliferation
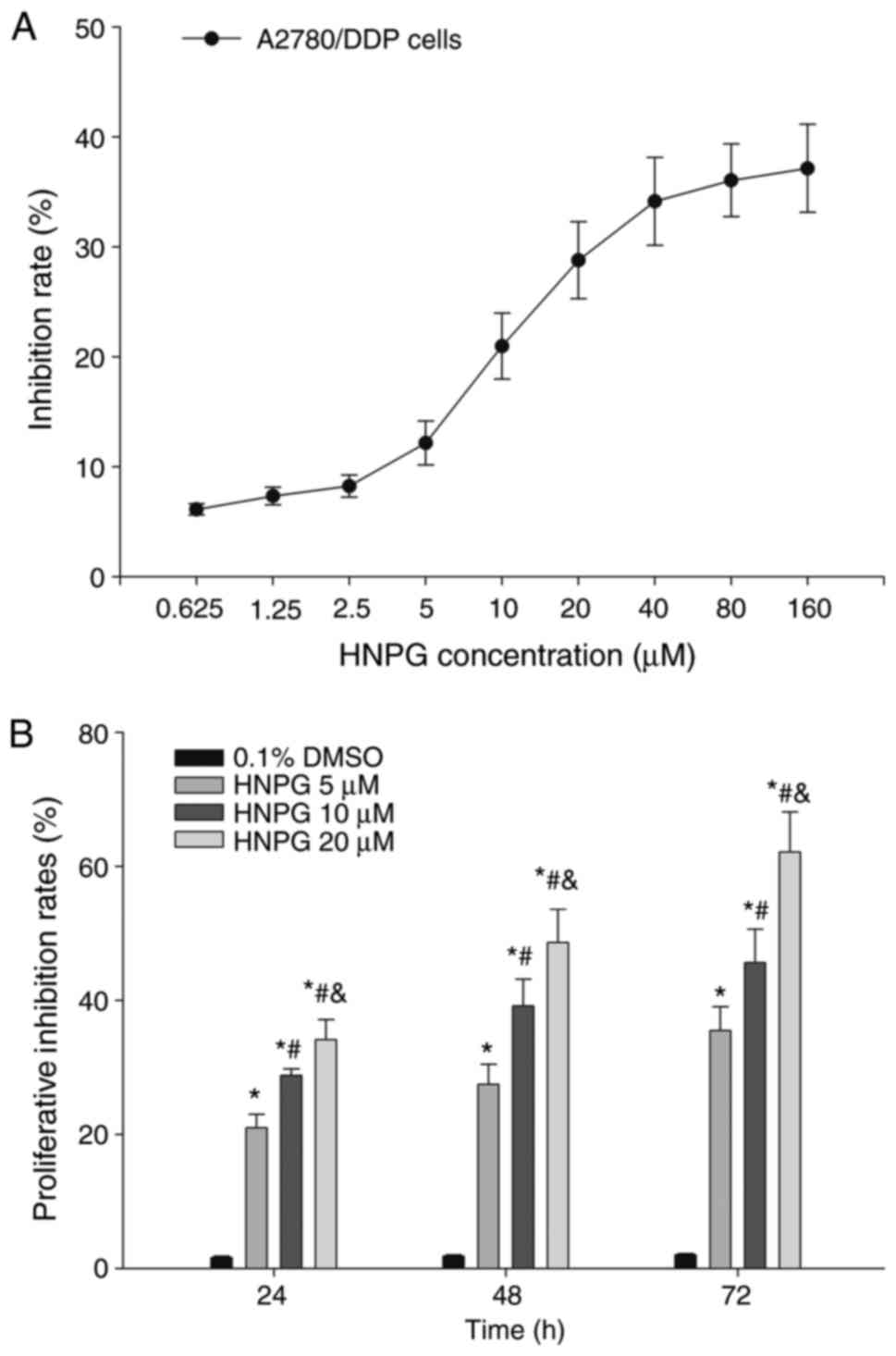
A2780/DDP cells were treated with different
concentrations of HNPG ranging from 0.625–160 µM for 24 h, and the
proliferation of A2780/DDP cells was inhibited by HNPG in a
dose-dependent manner. The inhibition rate of HNPG concentrations
ranging from 0.625–2.5 µM or from 40–160 µM was non-significant,
but the inhibition ratio was markedly increased by concentrations
ranging between 2.5–40 µM. A2780/DDP cells were exposed to
different doses of HNPG (5, 10 or 20 µM) for 24, 48 or 72 h,
respectively. The proliferation of A2780/DDP cells was markedly
inhibited in a dose-and time-dependent manner; the inhibition rate
of every HNPG-treated group was significantly different compared
with the control group (P5 µM/0.1% DMSO<0.05,
P10 µM/0.1% DMSO<0.05, P20 µM/0.1%
DMSO<0.05), and there was statistical difference among
each HNPG-treated group (P5/10 µM<0.05, P5/20
µM<0.05, P10/20 µM<0.05). The value of half
maximal inhibitory concentrations (IC50) were 16.32,
12.48 and 8.64 µM for 24, 48 and 72 h, respectively, as
demonstrated in Fig. 2A and B.
HNPG suppresses A2780/DDP cell clone
formation
A2780/DDP cells were incubated at 37°C with 0.1%
DMSO or different concentrations of HNPG (5, 10 or 20 µM) for 48 h.
Subsequently, the drug-containing medium was removed, and cells
were incubated with complete DMEM medium for 7 days until visible
clones formed. The rate of clone formation was markedly reduced and
the cell number inside the clones was significantly decreased. The
inhibition rate of clone formation was significantly increased in a
dose-dependent manner; every HNPG-treated group demonstrated a
marked statistical difference compared with the control group
(P5 µM/0.1% DMSO<0.05, P10 µM/0.1%
DMSO<0.05, P20 µM/0.1% DMSO<0.05). In
addition, there was statistical difference among each HNPG-treated
group (P5/10 µM<0.05, P5/20 µM<0.05,
P10/20 µM<0.05). The IC50 of clone
formation was 9.36 µM for 7 days, as demonstrated in Fig. 3A and B.
HNPG inhibits A2780/DDP cell invasion
ability
A2780/DDP cells were cultured with 0.1% DMSO or
different concentrations of HNPG (5, 10 or 20 µΜ) for 20 h, and the
invasive ability was markedly decreased in a dose-dependent manner.
The results demonstrated that the average cell numbers of control,
5, 10 and 20 µM groups invading through the Matrigel were
54.25±4.68, 36.75±3.22, 26.64±2.15 and 15.76±1.26, respectively.
There was a statistical difference between each HNPG-treated group
and the control (P5 µM/0.1% DMSO<0.05, P10
µM/0.1% DMSO<0.05, P20 µM/0.1% DMSO<0.05),
and there was statistical difference among each HNPG-treated group
(P5/10 µM<0.05, P5/20 µM<0.05,
P10/20 µM<0.05), as indicated in Fig. 4A and B.
HNPG affects A2780/DDP cells
metastasis ability
A2780/DDP cells were cultured with 0.1% DMSO or
different concentrations of HNPG (5, 10 or 20 µΜ) for 16 h, and the
metastasizing ability of A2780/DDP cells was significantly
decreased in a concentration-dependent manner. The results
demonstrated that the average cell numbers of control, 5, 10 and 20
µM groups that metastasized through polycarbonate membrane were
50.36±4.33, 32.45±3.05, 24.12±2.12 and 13.86±1.16, respectively.
There was a statistical difference among every HNPG-treated group
(P5/10 µM<0.05, P5/20 µM<0.05,
P10/20 µM<0.05), and each HNPG-treated group
exhibited a statistical difference compared with the control group
(P5 µM/0.1% DMSO<0.05, P10 µM/0.1%
DMSO<0.05, P20 µM/0.1% DMSO<0.05), as
demonstrated in Fig. 5A and B.
HNPG induces A2780/DDP cells
apoptosis
A2780/DDP cells were exposed to 0.1% DMSO and
different concentrations of HNPG (5, 10 or 20 µΜ) for 48 h, and the
apoptotic rate of A2780/DDP cells was significantly enhanced. The
results indicated that HNPG may markedly induce A2780/DDP cells
apoptosis in a dose-dependent manner; the apoptotic rates of
control, 5, 10 and 20 µM groups were 0.93±0.12, 21.09±2.15,
33.69±3.52 and 41.39±4.54%, respectively. Every HNPG-treated group
possessed statistical difference compared with control group
(P5 µM/0.1% DMSO<0.05, P10 µM/0.1%
DMSO<0.05, P20 µM/0.1% DMSO<0.05), and there
were significant differences among each HNPG-treated group
(P5/10 µM<0.05, P5/20 µM<0.05,
P10/20 µM<0.05), as exhibited in Fig. 6A and B.
HNPG affects A2780/DDP cells ROS
level
A2780/DDP cells were cultured with 0.1% DMSO and
different concentrations of HNPG (5, 10 or 20 µΜ) for 48 h, the
average intensity of fluorescence of DCFH-DA was increased in a
dose-dependent manner, which directly represented the level of
intracellular ROS. The results exhibited that HNPG may cause
oxidative injury in A2780/DDP cells, increase the level of
intracellular ROS of A2780/DDP cells; the mean fluorescence
intensities of DCFH-DA of control, 5, 10 and 20 µM groups were
2.72±0.25, 3.30±0.29, 4.29±0.40 and 5.05±0.51, respectively. There
were statistical differences among each HNPG-treated group
(P5/10 µM<0.05, P5/20 µM<0.05,
P10/20 µM<0.05), and all HNPG-treated groups
demonstrated statistical difference compared with the control group
(P5 µM/0.1% DMSO<0.05, P10 µM/0.1%
DMSO<0.05, P20 µM/0.1% DMSO<0.05), as
indicated in Fig. 7A and B.
HNPG changes A2780/DDP cell Δψm
A2780/DDP cells were treated to 0.1% DMSO and
different concentrations of HNPG (5, 10 or 20 µΜ) for 48 h, the
average intensity of fluorescence of Rh123 was decreased in
concentration-dependent manner, which were directly on behalf of
the level of Δψm. The results indicated that HNPG may markedly
decrease the Δψm of A2780/DDP cells and the mean fluorescence
intensities of Rh123 of control, 5, 10 and 20 µM groups were
19.4±2.16, 13.7±1.58, 8.38±0.82 and 4.61±0.52, respectively. There
were statistical differences among every HNPG-treated group
(P5 & 10 µM<0.05, P5 & 20
µM<0.05, P10 & 20 µM<0.05), and all of
HNPG-treated groups demonstrated statistical difference compared
with the control group (P5 µM & 0.1 % DMSO <0.05,
P10 µM & 0.1% DMSO<0.05, P20 µM & 0.1%
DMSO<0.05), as demonstrated in Fig. 8A and B.
HNPG regulates Bcl-2 family proteins
expression, releases Cyt-C from the mitochondria and activates a
caspase cascade in A2780/DDP cells
A2780/DDP cells were exposed to 0.1% DMSO and
different concentrations of HNPG (5, 10 or 20 µΜ) for 48 h, and the
average relative density of Bcl-2, Bax, Cyt-C, cleaved caspase-9
and cleaved caspase-3 were analyzed. The Bcl-2 expression level
exhibited a decreasing trend, while Bax, Cyt-C, cleaved caspase-9
and cleaved caspase-3 expression levels increased. The ratio of
Bcl-2/Bax also decreased. All HNPG-treated groups exhibited a
statistical difference compared with the control group (P5
µM/0.1% DMSO <0.05, P10 µM/0.1% DMSO<0.05,
P20µM/0.1% DMSO<0.05) in Bcl-2, Bax, Cyt-C, cleaved
caspase-9, cleaved caspase-3 expression levels and the ratio of
Bcl-2/Bax. There were statistical differences among every
HNPG-treated group in Bax, Cyt-C, cleaved caspase-9, cleaved
caspase-3 expression levels and the ratio of Bcl-2/Bax (P5/10
µM<0.05, P5/20 µM<0.05, P10/20
µM<0.05). There were significant differences between the 5
µΜ HNPG-treated group and the 10 or 20 µΜ HNPG-treated groups in
Bcl-2 expression (P5/10 µM<0.05 or P5/20
µM<0.05), but there were no difference between the 10 and
20 µΜ HNPG-treated groups in Bcl-2 expression (P10/20
µM>0.05), as exhibited in Fig.
9A-D.
 | Figure 9.The protein expression changes of
A2780/DDP cells incubated with different concentrations of HNPG (5,
10 or 20 µM) for 48 h. (A and B) Electrophoretograms demonstrating
the expression of Bcl-2, Bax, Cyt-C, cleaved caspase-3, cleaved
caspase-9. (C) Histogram demonstrating the mean relative gray
density value of Bcl-2, Bax, and the ratio of Bcl-2/Bax. (D)
Histogram indicating the average relative gray density value of
Cyt-C, cleaved caspase-3 and cleaved caspase-9. The data are
represented as the mean ± standard deviation for three independent
experiments. *P<0.05 vs. 0.1% DMSO group, #P<0.05
vs. 5 µM HNPG group, &P<0.05 vs. 10 µM HNPG.
HNPG, 5-hydroxy-4′-nitro-7-propionyloxy-genistein; DMSO, dimethyl
sulfoxide; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2 associated X
protein; Cyt-C, cytochrome C. |
Discussion
Proliferation, invasion and metastasis are basic
characteristics of tumors and these features have also become the
normal indices of measuring pharmacological activity (13,14). The
speed of proliferation and clone formation, and the capacities for
invasion and metastasis directly reflect the malignant degree of
tumors. Malignant tumors not only germinate in the primary site via
infiltrating and damaging adjacent organs and tissues, but also
metastasize to other areas to proliferate and grow (15,16).
Therefore, the fundamental methods of evaluating the
pharmacological effects of chemotherapeutics are detecting their
ability to inhibit the proliferation, clone formation, invasion and
metastasis of cancer cells. previous studies have suggested that
HNPG demonstrated inhibition of proliferation in gastric and breast
cancer cells in vitro (11,12), but
its molecular mechanism of this inhibition of proliferation has not
yet been elucidated. In the present study, the data demonstrated
that HNPG suppressed proliferation, clone formation, invasion and
metastasis, and induced apoptosis of human ovarian cancer A2780/DDP
cells in a concentration-and time-dependent manner, which
demonstrated novel antitumor effects of HNPG compared with previous
studies (11,12). These experimental results suggested
HNPG may be an excellent novel candidate for therapy in human
ovarian cancer A2780/DDP cells.
Apoptosis is a type of programmed cell death that
occurs in small areas or small numbers of cells at a time, and
serves an important role in the genesis and development of embryos,
alternation of new and old mature cells, biological degradation,
inflammation, atrophy, aging, autoimmune diseases and tumors
(17). In the early phase of
apoptosis, cell membrane phospholipids are asymmetrically lost,
which results in phosphatidylserine exposure at the cell surface.
These exposed phosphatidylserine molecules at the cell surface
exhibit a strong binding ability with Annexin V in the presence of
calcium (18). During the late phase
of apoptosis, propidium iodide (PI) enters the cytoplasm through
the cell membrane, and combines with the nucleus (19). In the results of the present study,
the numbers of Annexin V/PI-positive A2780/DDP cells were markedly
enhanced, in a dose-dependent manner, following HNPG treatment for
48 h. Therefore, it was suggested that the HNPG-mediated inhibition
of proliferation, clone formation, invasion and metastasis of
A2780/DDP cells may occur via an apoptotic pathway.
ROS include a series of molecules that directly or
indirectly originate from oxygen molecules and possess more
biological activities than oxygen molecules in cells that are
regarded as signaling molecules that regulate cell proliferation,
differentiation, survival and immune responses (19). Numerous in vitro studies have
revealed that the death of cancer cells was accompanied by a marked
accumulation of intracellular ROS, significant increases in
metabolic activity and markedly damaged mitochondrial function
(20). The damage to mitochondria may
promote the creation of ROS in cells, while the generation of
intracellular ROS may conversely cause a lipid peroxidation
reaction, which lead to various cellular events inducing cell
apoptosis or necrosis (21). In the
present study, the content of ROS in human ovarian cancer A2780/DDP
cells was markedly increased following HNPG treatment for 48 h.
Therefore, it was suggested that the HNPG-mediated inhibition of
proliferation, clone formation, invasion, metastasis and induction
of apoptosis may be through ROS accumulation in A2780/DDP
cells.
It is well-known that mitochondria serve a crucial
function in the extrinsic and intrinsic pathways of apoptosis; the
structural integrity and normal function of mitochondrial membranes
are critical for cell survival (22).
Notably, if the structure and function of mitochondrial membranes
sustain damage, for example through ultraviolet irradiation,
genotoxic agents or oxidative stress, this will trigger a series of
cellular events that will affect the basic characteristics of
malignant tumors, such as the proliferation, invasion and
metastasis, and even induce cells apoptosis or necrosis (23). Bcl-2 and Bax are the most important
apoptosis-inducing factors that function in the mitochondrial
membrane, jointly constituting certain ion channels that regulate
mitochondrial permeability transition (MPT). Once the mitochondrial
membrane is subjected to damage, Bcl-2, Bax and the ratio of
Bcl-2/Bax will be altered, which will trigger a series of cellular
events releasing Cyt-C from mitochondria (24). In the present study, in A2780/DDP
cells incubated with different concentrations of HNPG, it was
demonstrated that the ROS content was notably increased, along with
marked decreases in Δψm, a downregulation of Bcl-2, upregulation of
Bax and decreases in the Bcl-2/Bax ratio. These results suggest
that HNPG-mediated apoptosis may occur through the mitochondrial
pathway.
The caspase family serves a central role in
regulating apoptosis (25). It has
been established that caspase-8 or caspase-9 is activated by
apoptosis stimulating factors, for example Cyt-C or the Fas
Ligand-Fas-Fas-associated protein with death domain complex, which
will trigger the downstream caspase-3, and the activated caspase-3
will directly cause the loss of DNA repair function and activation
of endonuclease and DNA fragmentation, resulting in cell apoptosis
(26,27). In the present study, it was observed
that the A2780/DDP cells exposed to different concentrations of
HNPG underwent apoptosis in a dose-dependent manner, accompanied by
the upregulation of Cyt-C, cleaved caspase-9 and cleaved caspase-3.
The observed caspase-mediated properties of HNPG were in agreement
with the basic properties and functions of the caspase family
(27). Therefore, these results
suggested that HNPG-triggered apoptosis was potentially mediated,
at least in part, by this caspase cascade.
In conclusion, HNPG demonstrated significant
cytotoxic activity in human ovarian cancer A2780/DDP cells. HNPG
inhibited the rates of proliferation, clone formation, invasion and
metastasis, and induced apoptosis in vitro. Simultaneously,
the levels of intracellular ROS and Δψm were increased and
decreased, respectively. Additionally, HNPG downregulated the
expression of Bcl-2, upregulated the expression of Bax, led to
Cyt-C release from mitochondria, activated caspase-9 and caspase-3
and caused cell apoptosis. Compared with previous studies examining
HNPG (11,12), the present study not only detected its
anti-proliferative effects, but also examined the inhibitory
effects on clone formation, invasion and metastasis, induction of
apoptosis, accumulation of ROS and alteration of Δψm. The present
study demonstrated the antitumor effect of HNPG from multiple
perspectives and additionally investigated the basic molecular
biological mechanism of HNPG. The results not only provided
additional data concerning the antitumor effects of HNPG, but also
aimed to elucidate the basic molecular biological mechanisms
involved. In summary, HNPG indicated a marked cytotoxic activity in
human ovarian cancer A2780/DDP cells via the ROS-mediated
mitochondrial dysfunction pathway, which suggests that HNPG may be
a novel candidate for chemotherapeutic drug development. Although
the antitumor effect of HNPG and its basic molecular biological
mechanism were detected in vitro, there are several
limitations requiring additional investigation, such as the absence
of data of the antitumor mechanism on A2780 cells which may assist
to elucidate the underlying resistance mechanisms of HNPG on A2780
cells, the lack of data on normal cells that may explain the side
effects of HNPG on normal cells and tissues and organs, the absence
of data on positive control groups, for example 5-fluoracil,
paclitaxel and methotrexate, which may assist in illuminating the
pharmacological effects of HNPG and the effective pharmacological
ratio between HNPG and clinical common drugs. Subsequent studies
will investigate the metabolism of HNPG in experimental animal
models, detect its blood drug concentration and its half-life and
the side effects on the brain, heart, lung, liver and kidney
cells.
Acknowledgements
The present study was supported by The First
Clinical School of Jinan University (grant no. FRPR201601-04). The
authors would like to thank Professor Wanyu Xie (the First
Affiliated Hospital of University of South China, Hengyang, Hunan,
China) for her technical assistance.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wijdeven RH, Pang B, Assaraf YG and
Neefjes J: Old drugs, novel ways out: Drug resistance toward
cytotoxic chemotherapeutics. Drug Resist Updat. 28:65–81. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bergamini A, Pisano C, Di Napoli M,
Arenare L, Della Pepa C, Tambaro R, Facchini G, Gargiulo P,
Rossetti S, Mangili G, et al: Cisplatin can be safely administered
to ovarian cancer patients with hypersensitivity to carboplatin.
Gynecol Oncol. 144:72–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brozovic A: The relationship between
platinum drug resistance and epithelial-mesenchymal transition.
Arch Toxicol. 91:605–619. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jin L, Xu M, Luo XH and Zhu XF: Stephania
tetrandra and ginseng-containing Chinese herbal formulation NSENL
reverses cisplatin resistance in lung cancer xenografts. Am J Chin
Med. 45:385–401. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Henklewska M, Pawlak A, Pruchnik H and
Obminska-Mrukowicz B: Complex of platinum(II) with
tris(2-carboxyethyl) phosphine induces apoptosis in canine
lymphoma/leukemia cell lines. Anticancer Res. 37:539–546. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Y, Cass SL, Kutty SK, Yee EM, Chan
DS, Gardner CR, Vittorio O, Pasquier E, Black DS and Kumar N:
Synthesis, biological evaluation and structure-activity
relationship studies of isoflavene based mannich bases with potent
anti-cancer activity. Bioorg Med Chem Lett. 25:5377–5383. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Uesawa Y, Sakagami H, Kagaya H, Yamashita
M, Takao K and Sugita Y: Quantitative structure-cytotoxicity
relationship of 3-benzylidenechromanones. Anticancer Res.
36:5803–5812. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wootten D, Simms J, Koole C, Woodman OL,
Summers RJ, Christopoulos A and Sexton PM: Modulation of the
glucagon-like peptide-1 receptor signaling by naturally occurring
and synthetic flavonoids. J Pharmacol Exp Ther. 336:540–550. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagib DA and MacMillan DW:
Trifluoromethylation of arenes and heteroarenes by means of
photoredox catalysis. Nature. 480:224–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lv H, Yang J, Wang C, Yu F, Huang D and Ye
L: The WNT7B protein promotes the migration and differentiation of
human dental pulp cells partly through WNT/beta-catenin and c-Jun
N-terminal kinase signalling pathways. Arch Oral Biol. 87:54–61.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang JH, Gao CJ and Meng Lk: Study on the
synthesis and antitumor effects of
5-Hydroxy-4′-nitro-7-propionyloxy-isoflavone. China Pharmacist.
15:1378–1385. 2012.(In Chinese).
|
|
12
|
Jin YS, Liu CM, Wu QY, Yao B, Dai Y, Zhang
LR and Shen XL: Design and synthesis of genistein derivatives
5-hydroxy-4′-nitro-7-substituted acyloxy isoflavone and their
antitumor effects. Acad J Mil Med Univ. 26:182–185. 2005.(In
Chinese).
|
|
13
|
Singh S, Guetzko M and Resnick K:
Preoperative predictors of delay in initiation of adjuvant
chemotherapy in patients undergoing primary debulking surgery for
ovarian cancer. Gynecol Oncol. 143:241–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Z, Mei J and Tan Y: Baicalin attenuates
DDP (cisplatin) resistance in lung cancer by downregulating MARK2
and p-Akt. Int J Oncol. 50:93–100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Solmaz Hasdemir P and Guvena T: Borderline
ovarian tumors A contemporary review of clinicopathological
characteristics, diagnostic methods and therapeutic options. J
Buon. 21:780–786. 2016.PubMed/NCBI
|
|
16
|
Erdogan S, Turkekul K, Serttas R and
Erdogan Z: The natural flavonoid apigenin sensitizes human CD44+
prostate cancer stem cells to cisplatin therapy. Biomed
Pharmacother. 88:210–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mirzaei MR, Mahmoodi M, Hassanshahi G and
Ahmadi Z: Down-regulation of anti-apoptotic genes in tumor cell
lines is facilitated by suppression of OCT4B1. Adv Med Sci.
62:97–102. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jian KL, Zhang C, Shang ZC, Yang L and
Kong LY: Eucalrobusone C suppresses cell proliferation and induces
ROS-dependent mitochondrial apoptosis via the p38 MAPK pathway in
hepatocellular carcinoma cells. Phytomedicine. 25:71–82. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Han X, Zhen S, Ye Z, Lu J, Wang L, Li P,
Li J, Zheng X, Li H, Chen W, et al: A feedback loop between
miR-30a/c-5p and DNMT1 mediates cisplatin resistance in ovarian
cancer cells. Cell Physiol Biochem. 41:973–986. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bauer G: Central signaling elements of
intercellular reactive oxygen/nitrogen species-dependent induction
of apoptosis in malignant cells. Anticancer Res. 37:499–513. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pluchino LA, Choudhary S and Wang HC:
Reactive oxygen species-mediated synergistic and preferential
induction of cell death and reduction of clonogenic resistance in
breast cancer cells by combined cisplatin and FK228. Cancer Lett.
381:124–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bauer D, Werth F, Nguyen HA, Kiecker F and
Eberle J: Critical role of reactive oxygen species (ROS) for
synergistic enhancement of apoptosis by vemurafenib and the
potassium channel inhibitor TRAM-34 in melanoma cells. Cell Death
Dis. 8:e25942017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zajac J, Kostrhunova H, Novohradsky V,
Vrana O, Raveendran R, Gibson D, Kasparkova J and Brabec V:
Potentiation of mitochondrial dysfunction in tumor cells by
conjugates of metabolic modulator dichloroacetate with a Pt (IV)
derivative of oxaliplatin. J Inorg Biochem. 156:89–97. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vijayarathna S, Oon CE, Chen Y, Kanwar JR
and Sasidharan S: Polyalthia longifolia methanolic leaf extracts
(PLME) induce apoptosis, cell cycle arrest and mitochondrial
potential depolarization by possibly modulating the redox status in
hela cells. Biomed Pharmacother. 89:499–514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nasser MI, Masood M, Wei W and Li X, Zhou
Y, Liu B, Li J and Li X: Cordycepin induces apoptosis in SGC-7901
cells through mitochondrial extrinsic phosphorylation of PI3K/Akt
by generating ROS. Int J Oncol. 50:911–919. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsin IL, Wang SC, Li JR, Ciou TC, Wu CH,
Wu HM and Ko JL: Immunomodulatory proteins FIP-gts and chloroquine
induce caspase-independent cell death via autophagy for
resensitizing cisplatin-resistant urothelial cancer cells.
Phytomedicine. 23:1566–1573. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leekha A, Gurjar BS, Tyagi A, Rizvi MA and
Verma AK: Vitamin C in synergism with cisplatin induces cell death
in cervical cancer cells through altered redox cycling and p53
upregulation. J Cancer Res Clin Oncol. 142:2503–2514. 2016.
View Article : Google Scholar : PubMed/NCBI
|