Introduction
Primary hepatocellular carcinoma (HCC) is the most
common type of malignant tumor worldwide, an incidence ranked
third-highest of all types of cancer, a high degree of malignancy
and a five-year survival rate of ~30% (1). To date, no effective treatments have
been developed (2). HCC is commonly
diagnosed numerous countries with a high incidence of viral
hepatitis (3). HCC contributes 85–90%
of all primary liver cancers, with 500,000 new cases and 250,000
HCC-associated mortalities occurring worldwide every year (4). The main reasons for the high mortality
rate of HCC patients are ascribed to the lack of effective
treatments and the increasing resistance to conventional
radiotherapy and chemotherapy (5).
The proliferation of hepatoma cells is closely associated with HCC
occurrence, development, invasion and metastasis (1). Small molecule bioactive peptides have
various unique biological characteristics, including simple
structures, extensive tissue distribution and a range of biological
effects, which serve important roles in the regulation of the human
metabolism and physiological function (6). A number of cytokines and small peptides
have essential roles in the development of liver cancer. Intermedin
(IMD) belongs to the calcitonin gene-related peptide (CGRP)
superfamily (7). It was initially
identified in teleost fish in 2004, and has also been identified in
cDNA clones of other species, such as rodents and primates
(8). IMD is widely distributed in the
body, and its expression levels in adrenocortical tumors (9) and colorectal cancer (10) are high compared with those in normal
tissues. IMD may also promote angiogenesis (11), suggesting that it is associated with
the occurrence and development of tumors. The present study
explored the effect of IMD in HCC and its underlying molecular
mechanisms.
Materials and methods
HepG2 cells were purchased from Shanghai Bogoo
Biological Technology Co., Ltd. (Shanghai, China), recombinant
human IMD1-53 and its receptor antagonist (IMD17-47) were purchased
from Phoenix Pharmaceuticals, Inc. (Burlingame, CA, USA). The Cell
Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular
Technologies, Inc. (Kumamoto, Japan). The RNA extraction, reverse
transcription (RT. kit and Luciferase reporter gene assay kit were
purchased from Promega Corporation (Madison, WI, USA). The Taq
enzyme was purchased from Tiangen Biotech Co., Ltd. (Beijing,
China. and the fluorescent EvaGreen® Dye was purchased
from Biotium, Inc. (Fremont, CA, USA). The classical Wnt pathway
inhibitor (IWR-1-endo. was purchased from Cayman Chemical (Ann
Arbour, MI, USA), the TOPflash/FOPflash plasmids for Wnt signaling
pathway activity detection were purchased from Merck KGaA
(Darmstadt, Germany) and the jetPEI® transfection
reagent was purchased from Polyplus-Transfection SA (Illkirch,
France).
Cell proliferation was detected using
CCK-8
HepG2 cells in the logarithmic growth phase were
inoculated in flat-bottomed 96-well plates at a density of
1×104/100 cells/µl, and cultured in 5% CO2
incubator at 37°C. After 12 h, the medium was replaced with fresh
culture medium (Shanghai Yuan Long Biology Technology Company.
containing various concentrations (0, 1, 5, 10, 50 and 100 nM. of
IMD1-53, or with 100 nM IMD17-47 and 25 nM IWR-1-endo. The complete
culture medium without IMD and RPMI-1640 was used as the control. A
total of 48 h after inoculation, 10 µl CCK-8 was added into each
well of the plate and placed in a 37°C incubator for 1–4 h. A
microplate reader was then used to measure the absorbance value at
a wavelength of 450 nm, as previously described (12).
RNA extraction
TRIzol® reagent (Promega Corporation) was
used; the cells were aspirated from the culture medium and washed
with PBS, following which 1 ml RNAtrip reagent (Tideradar Beijing
Technology Co., Ltd., Beijing, China. was added. The cells were
triturated 30 times, transferred into an Eppendorf (EP) tube and
placed at room temperature for 10 min. A total of 0.2 ml chloroform
was added, reversal agiation was performed for 15 sec and the
samples were then incubated at room temperature for 5 min. The
samples were centrifuged at 12,000 × g for 15 min at 4°C, and the
resulting upper aqueous phase was carefully aspirated and
transferred to a clean EP tube. Subsequently, an equal volume of
isopropanol was added and mixed. The samples were allowed to
precipitate for 2 h at −70°C, prior to being centrifuged at 12,000
× g for 10 min at 4°C. The supernatant was discarded, and the cells
were precipitated and washed with 1 ml 0.75X ice-cold ethanol. They
were then centrifuged again at 12,000 × g and 4°C for 10 min,
following which the supernatant was discarded. When the whitish
pattern appeared in the center, the RNA enzyme pure water 50–100 µl
was added, and quantified with a spectrophotometer.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The reverse transcription reaction involved a 20 µl
reaction system 5-(and-6)-carboxy-2′,7′-dichlorofluorescein (pH
9.0; purchased from Beijing Jiamei Nuonuo Biological Technology
Co., Ltd.), with 2 µg RNA (protector RNase inhibitor) starting, and
used a reverse transcription kit according to the manufacturer's
protocol (purchased from Promega Corporation) for first strand cDNA
synthesis. RT-qPCR (Shanghai Solarbio Biological Technology Co.,
Ltd.. involved a 25 µl reaction system, and 1 µl reverse
transcription reaction mixture was used as a template.
Amplification conditions were as follows: Pre-degeneration for 5
min at 94°C, degeneration for 30 sec at 94°C, annealing for 30 sec
at 58°C, extension for 30 sec at 72°C then extension for 5 min at
72°C, with a total of 40 cycles. Stratagene Mx3000p software
(Stratagene; Agilent Technologies, Inc., Santa Clara, CA, USA) was
used for analysis. The 2−ΔΔCq method was used to
quantify expression levels (13).
Primer sequences were as follows: Human c-Myc upstream,
5′-TGCTCCATGAGGAGACACC-3′ and downstream,
5′-CTTTTCCACAGAAACAACATCG-3′; human cyclin D1 upstream,
5′-GAAGATCGTCGCCACCTG-3′ and downstream,
5′-GACCTCCTCCTCGCACTTCT-3′; human β-actin upstream,
5′-ATCTGGCACCACACCTTC-3′ and downstream, 5′-AGCCAGGTCCAGACGCA-3′
(Shanghai, China Solarbio Biological Technology Co., Ltd.). The
RT-qPCR protocol was repeated 5 times.
Luciferase activity measurement
HepG2 cells in the logarithmic growth phase were
seeded on 12-well plates at 1×106 cells/well, then
placed in a 5% CO2 incubator at 37°C for 12 h, and each
well was transfected with 0.5 µg various plasmids (using TOPFlash
as the reporter plasmid, FOPFlash as a negative control and pRL-TK
as an internal control) using jetPEI® reagent, and then
incubated at 37°C for 24 h. The luciferase activity was measured
with the Dual-Luciferase™ Reporter Assay System (Promega
Corporation), according to the manufacturer's protocol (14).
Statistical analysis
All results are presented as the mean ± standard
error of the mean. GraphPadPrism5.0® software (GraphPad
Software, Inc., La Jolla, CA, USA) was used for analysis and
creating the data graphs. A one-way analysis of variance was
conducted for multi-group comparison, and the Student-Newman-Keuls
method was used for post-hoc analysis and comparisons between the
groups. Student's t-test was used to compare the results between
groups. P<0.05 was considered to indicate a significantly
different difference.
Results
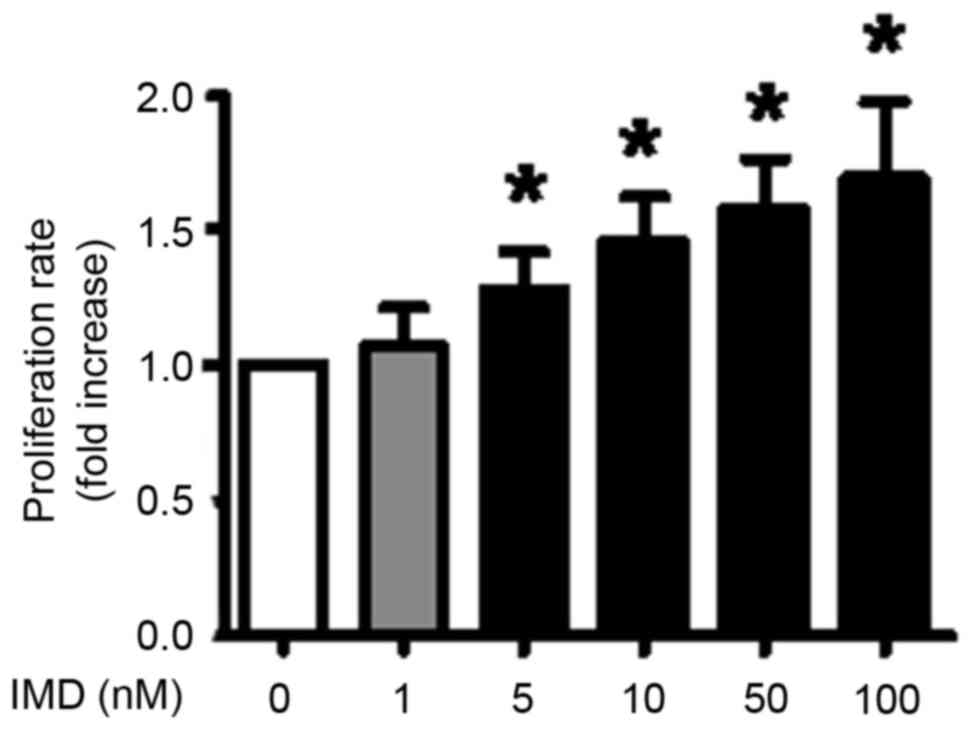
IMD induces HepG2 cell proliferation
in a dose-dependent manner
The present study explored the direct effect of IMD
on HepG2 cell proliferation. The results of the CCK-8 test
demonstrated that the administration of 5–100 nM IMD for 48 h
significantly promoted HepG2 cell proliferation (Fig. 1; P<0.05, compared with the
control).
IMD promotes HepG2 cell proliferation
in a dose-dependent manner
Additional analysis revealed that treatment with 100
nM IMD17-47 (competitive antagonist of the IMD receptor)
significantly inhibited the promotion effect of 10 nM IMD on cell
proliferation (P<0.05). Furthermore, IMD was observed to promote
HepG2 cell proliferation in a dose-dependent manner, and exhibited
a receptor-dependent manner (Fig.
2).
IMD activates the classical Wnt
signaling pathway in HepG2 cells
The possible underlying mechanisms associated with
IMD-promoted HepG2 cell proliferation were also investigated. The
Wnt signaling pathway serves an important role in tumor cell
proliferation. TOPflash was used as a reporter plasmid, FOPflash
was used as the negative control, and a thymidine kinase
promoter-Renilla luciferase reporter plasmid was used as the
internal reference control. Subsequent to transfection for 6 h and
treatment with 10 nM IMD for 24 h, a dual luciferase activity assay
was conducted. The results demonstrated that the TOPflash/FOPflash
ratio following IMD stimulation was significantly higher compared
with in the control group (P<0.05; Fig. 3A). The mRNA levels of the downstream
target genes c-Myc and cyclin D1 were identified to
be significantly increased (P<0.05; Fig. 3B), suggesting that the classical Wnt
signaling pathway was activated following IMD stimulation, and that
specific transcriptional activity was significantly enhanced
(P<0.05; Fig. 3).
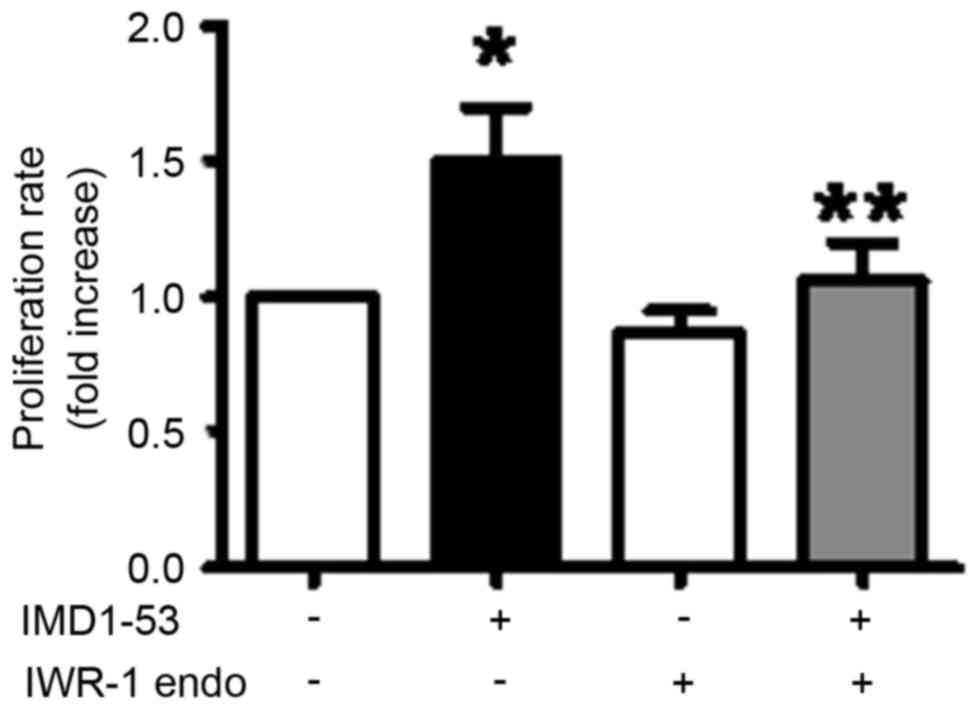
IWR-1-endo is able to partially
inhibit IMD-induced HepG2 cell proliferation
Pre-treatment with 25 µM IWR-1-endo for 1 h
significantly inhibited 10 nM IMD-induced HepG2 cell proliferation
(P<0.05, compared with the control). Therefore, IMD may promote
HepG2 cell proliferation via the Wnt signaling pathway (Fig. 4).
Discussion
The results of the present study have indicated that
IMD is able promote HepG2 cell proliferation via the classical Wnt
signaling pathway, according to the data that suggest the
following: IMD treatment may directly induce HepG2 cell
proliferation; IMD17-47 may block HepG2 cell proliferation; IMD may
activate the classical Wnt signaling pathway transcriptional
activity and affect downstream target gene mRNA expression levels;
blocking the Wnt signaling pathway may inhibit IMD-induced HepG2
cell proliferation.
IMD is a member of the CGRP superfamily, which
exhibits ~30% similarity with the amino acid structure of
adrenomedullin (AM), and is therefore also known as AM2 (7). Its precursor consists of 148 amino acid
residues with multiple protease cleavage sites, and may be cut into
three active fragments: IMD 1–47; IMD 8–47; IMD1-53 (15). CGRP may serve its role through binding
with calcitonin receptor like receptors/receptor activity modifying
protein receptor complexes (RAMPs) (16). CGRP primarily acts on RAMP1, AM
principally acts on RAMP2/3 and IMD non-selectively acts on
RAMP1/2/3; therefore, IMD may have a wider range of biological
effects (8). A number of studies have
investigated the effect of IMD on homeostasis regulation,
hypertension, myocardial ischemia, heart failure and renal failure
(15,17,18).
Although previous studies have revealed that the expression of IMD
in pancreatic cancer and breast cancer was higher than the control
group (19,20), the effect of IMD in tumors,
particularly in the occurrence and development of liver cancer,
remains unclear. The results of the present study demonstrated that
IMD may promote HepG2 cell proliferation (Fig. 1), and this effect is achieved through
receptor-dependence (Fig. 2). The
autocrine hypothesis proposed that numerous tumor cells may react
with self-secreted growth factors, such as basic fibroblast growth
factor, and thereby escape from growth inhibition (21,22). The
present study also observed that the IMD receptor blocker IMD17-47
can still inhibit the proliferation of HepG2 cells without
exogenous IMD stimulation (Fig. 2),
indicating that HepG2 cell may express and secrete IMD. This result
is concordant with the data obtained by Guo et al (23).
The Wnt signaling pathway has an important role in
the proliferation of tumor cells. Activation of the classical Wnt
signaling pathway is dependent on the abnormal accumulation of
β-catenin in the cytoplasm, which then enters the nucleus and binds
with TCF/LEF to form transcription factor complexes, these
complexes then regulate the expression of various downstream genes,
including cyclin D1 and c-Myc (24,25).
Previous studies have demonstrated that 30–40% of liver cancer
cases also exhibit a disorder of the Wnt signaling pathway or a
β-catenin gene mutation; β-catenin expression may be observed in
the cytoplasm and nucleus of 62–70% of liver cancer cells, and has
been observed to be marked in poorly differentiated HCC cell nuclei
(26–28). c-Myc is the first discovered
downstream target gene of the β-catenin/TCF complex, and c-Myc
serves an important role in various biological process, including
cell apoptosis, proliferation, metabolism, DNA repair and
angiogenesis (29). The cyclin
D1 gene is an additional direct target gene of the
β-catenin/TCF pathway; sustained high expression levels of
cyclin D1 may cause cells to remain in the S phase of
growth, resulting in excessive proliferation and eventual malignant
transformation (30). In the present
study, the results of the TOPflash/FOPflash luciferase assay
indicated that IMD may enhance the transcriptional activity of the
β-catenin/TCF complex; the expression of the Wnt downstream genes
cyclin D1 and c-Myc were also significantly
upregulated, suggesting that IMD may activate the Wnt signaling
pathway, thus promoting HepG2 cell proliferation.
In conclusion, the present study demonstrated that,
as a member of the CGRP superfamily, IMD binds with receptor, and
then activates the Wnt signaling pathway, thus, it serves a role in
promoting the proliferation of liver cancer cells, which presents a
novel strategy for the treatment of liver cancer. In addition, IMD
antagonists may be used as a potential anti-liver cancer agent.
Acknowledgements
The present study was supported by the Project of
Shenyang Municipal Science and Technology Bureau (grant no.
F12-93-9-56).
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IMD
|
intermedin
|
|
HCC
|
primary hepatocellular carcinoma
|
References
|
1
|
Tang B, Liang X, Tang F, Zhang J, Zeng S,
Jin S, Zhou L, Kudo Y and Qi G: Expression of USP22 and Survivin is
an indicator of malignant behavior in hepatocellular carcinoma. Int
J Oncol. 47:2208–2216. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhuang PH, Xu L, Gao L, Lu W, Ruan LT and
Yang J: Correlations of microvascular blood flow of
contrast-enhanced ultrasound and HGF/c-Met signaling pathway with
clinicopathological features and prognosis of patients with
hepatocellular carcinoma. Onco Targets Ther. 10:847–857. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dai L, Peng XX, Tan EM and Zhang JY:
Tumor-associated antigen CAPERα and microvessel density in
hepatocellular carcinoma. Oncotarget. 7:16985–16995. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bozkaya G, Korhan P, Cokakli M, Erdal E,
Sağol O, Karademir S, Korch C and Atabey N: Cooperative interaction
of MUC1 with the HGF/c-Met pathway during hepatocarcinogenesis. Mol
Cancer. 11:642012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Whittaker S, Marais R and Zhu AX: The role
of signaling pathways in the development and treatment of
hepatocellular carcinoma. Oncogene. 29:4989–5005. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang C: Review on nutrition function of
bioactive pephides. Food Science. 24:153–154. 2003.
|
|
7
|
Holmes D, Campbell M, Harbinson M and Bell
D: Protective effects of intermedin on cardiovascular, pulmonary
and renal diseases: Comparison with adrenomedullin and CGRP. Curr
Protein Pept Sci. 14:294–329. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takei Y, Inoue K, Ogoshi M, Kawahara T,
Bannai H and Miyano S: Identification of novel adrenomedullin in
mammals: A potent cardiovascular and renal regulator. FEBS Lett.
556:53–58. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morimoto R, Satoh F, Murakami O, Hirose T,
Totsune K, Imai Y, Arai Y, Suzuki T, Sasano H, Ito S and Takahashi
K: Expression of adrenomedullin 2/intermedin in human adrenal
tumors and attached non-neoplastic adrenal tissues. J Endocrinol.
198:175–183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hikosaka T, Tsuruda T, Nagata S, Kuwasako
K, Tsuchiya K, Hoshiko S, Inatsu H, Chijiiwa K and Kitamura K:
Adrenomedullin production is increased in colorectal
adenocarcinomas; its relation to matrix metalloproteinase-9.
Peptides. 32:1825–1831. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smith RS Jr, Gao L, Bledsoe G, Chao L and
Chao J: Intermedin is a new angiogenic growth factor. Am J Physiol
Heart Circ Physiol. 297:H1040–H1047. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu Y, Pang X, Dong M, Wen F and Zhang Y:
Nesfatin-1 inhibits ovarian epithelial carcinoma cell proliferation
in vitro. Biochem Biophys Res Commun. 440:467–472. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Zhang S, Shang H, Pang X and Zhao
Y: Basic fibroblast growth factor upregulates adrenomedullin
expression in ovarian epithelial carcinoma cells via JNK-AP-1
pathway. Regul Pept. 157:44–50. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bell D and McDermott BJ: Intermedin
(adrenomedullin-2): A novel counter-regulatory peptide in the
cardiovascular and renal systems. Br J Pharmacol. 153 Suppl
1:S247–S262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roh J, Chang CL, Bhalla A, Klein C and Hsu
SY: Intermedin is a calcitonin/calcitonin gene-related peptide
family peptide acting through the calcitonin receptor-like
receptor/receptor activity-modifying protein receptor complexes. J
Biol Chem. 279:7264–7274. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong F, Taylor MM, Samson WK and Ren J:
Intermedin (adrenomedullin-2. enhances cardiac contractile function
via a protein kinase C- and protein kinase A-dependent pathway in
murine ventricular myocytes. J Appl Physiol (1985). 101:778–784.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren H, Ren H, Li X, Yu D, Mu S, Chen Z and
Fu Q: Effects of intermedin on proliferation, apoptosis and the
expression of OPG/RANKL/M-CSF in the MC3T3-E1 osteoblast cell line.
Mol Med Rep. 12:6711–6717. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hollander LL, Guo X, Salem RR and Cha CH:
The novel tumor angiogenic factor, adrenomedullin-2, predicts
survival in pancreatic adenocarcinoma. J Surg Res. 197:219–224.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu YM, Zhong JB, Wang HY, Yu XF and Li ZQ:
The prognostic value of intermedin in patients with breast cancer.
Dis Markers. 2015:8621582015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Billottet C, Janji B, Thiery JP and
Jouanneau J: Rapid tumor development and potent vascularization are
independent events in carcinoma producing FGF-1 or FGF-2. Oncogene.
21:8128–8139. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Di Blasio AM, Cremonesi L, Viganó P,
Ferrari M, Gospodarowicz D, Vignali M and Jaffe RB: Basic
fibroblast growth factor and its receptor messenger ribonucleic
acids are expressed in human ovarian epithelial neoplasms. Am J
Obstet Gynecol. 169:1517–1523. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo X, Schmitz JC, Kenney BC, Uchio EM,
Kulkarni S and Cha CH: Intermedin is overexpressed in
hepatocellular carcinoma and regulates cell proliferation and
survival. Cancer Sci. 103:1474–1480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moon RT, Bowerman B, Boutros M and
Perrimon N: The promise and perils of Wnt signaling through
beta-catenin. Science. 296:1644–1646. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wong CM, Fan ST and Ng IO: Beta-catenin
mutation and overexpression in hepatocellular carcinoma:
Clinicopathologic and prognostic significance. Cancer. 92:136–145.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Devereux TR, Stern MC, Flake GP, Yu MC,
Zhang ZQ, London SJ and Taylor JA: CTNNB1 mutations and
beta-catenin protein accumulation in human hepatocellular
carcinomas associated with high exposure to aflatoxin B1. Mol
Carcinog. 31:68–73. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hsu HC, Jeng YM, Mao TL, Chu JS, Lai PL
and Peng SY: Beta-catenin mutations are associated with a subset of
low-stage hepatocellular carcinoma negative for hepatitis B virus
and with favorable prognosis. Am J Pathol. 157:763–770. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bienz M and Clevers H: Linking colorectal
cancer to Wnt signaling. Cell. 103:311–320. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tetsu O and McCormick F: Beta-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|