Introduction
Breast cancer is the most common malignancy in
females, accounting for ~1/3 of all types of cancer (1). Despite advances in the detection and
treatment of primary and metastatic cancers, and the application of
surgery, radiotherapy, chemotherapy, immunotherapy and drug
combinations, the mortality rate from cancer remains high, and side
effects from the aforementioned combined treatments are severe
(2). Therefore, identifying a more
effective and less dangerous therapy for cancer is imperative.
Genetically engineered mesenchymal stem cells (MSCs)
have been hypothesized to exhibit therapeutic potential in a wide
range of human diseases, including cancer. Intravenous injection of
MSCs expressing interferon (IFN)-β may inhibit the expansion of the
pulmonary metastasis of melanoma and breast cancer in mice
(3,4),
and prolong the survival of mice with glioma xenografts (5). It is well accepted that a critical
property of MSCs for cell therapies is their intrinsic homing
ability; when infused systemically, MSCs are able to home to the
sites of tumor, injury, inflammation and ischemia, although the
underlying molecular mechanisms remain unclear (6,7). Previous
studies have indicated the role of cytokines and chemokines
secreted by target tissues and tumors, including vascular
endothelial growth factor, C-C motif chemokine ligand (CCL)2 and
CCL5, in MSC rolling, arrest and transmigration along the
endothelium. Following transmigration, MSCs were able to contribute
to the antitumor effect by synthesizing the engineered proteins
(8,9).
It is hypothesized that the innate and acquired immune responses
serve crucial roles in the antitumor response, and the interactions
between the host immune system and tumor cells have been the
subject of intense research over the last decades (10). Cytokines, including transforming
growth factor-β, tumor necrosis factor-α (TNF-α), interleukin
(IL)-1, IL-12, IL-18 and IFNs, serve important roles in the immune
response (11,12). Alterations in cytokine levels are
important parameters that affect the course of disease. IL-18, a
more recently described member of the IL-1 cytokine superfamily, is
now recognized as an important regulator of innate and acquired
immune responses (13). IL-18 is
produced by macrophages and immature DC early in the acute immune
response, and serves an important role in the Th1 response,
primarily by its ability to induce IFN-γ production by T cells and
natural killer (NK) cells (14). In
addition to IFN-γ, IL-18 also induces granulocyte/macrophage
colony-stimulating factor, TNF-α and IL-1 expression, and acts in
concert with IL-12 (13,15). Nakata et al (16) demonstrated that IL-18 was able to
inhibit osteolytic growth at bone metastatic sites and suppress an
early onset of bone metastasis in a breast cancer mouse model.
Coskun et al (17)
demonstrated that serum IL-18 levels were significantly increased
in patients with breast cancer compared with controls. The increase
in IL-18 levels was identified to be associated with established
clinically used prognostic factors, including tumor size, axillary
lymph node involvement and disease stage (18).
Our previous study identified that human MSCs
derived from umbilical cord (hUMSCs) genetically modified with the
IL-18 gene (hUMSCs/IL-18) were able to significantly decrease the
proliferation, migration and invasion of breast cancer cells (MCF-7
and HCC1937 cells) in vitro (19). The underlying molecular mechanism for
this suppression of proliferation may be the G1- to S-phase arrest
of breast cancer cells induced by hUMSCs/IL-18. The transduced
hUMSCs maintained their differentiation potential and pluripotency,
and were capable of migration (19).
However, limited data exist concerning the presence of IL-18 in
human tumors in vivo, therefore, investigation of the effect
of hUMSCs/IL-18 on other types of tumor in vivo is
warranted.
The aim of the present study was to determine
whether hUMSCs/IL-18 were able to inhibit the proliferation and
metastasis of breast cancer cells in vivo. A breast cancer
mouse model was developed by injecting 4T1 cells subcutaneously
(s.c.) into BALB/c mice, and injecting hUMSCs/IL-18 at the early (1
week after injection of 4T1 cells) and late (4 weeks after
injection of 4T1 cells) stages of breast cancer, to investigate the
safety and effect of hUMSCs/IL-18 on breast cancer progression.
Materials and methods
Animals
The present study was approved by the Institutional
Animal Ethical Committee of Qingdao University (Qingdao, China) and
the Ethics Committee of the Affiliated Hospital of Qingdao
University. All experimental procedures involving animals were
performed in accordance with the Guide for the Care and Use of
Laboratory Animals (National Institutes of Health publication no.
80-23, revised 1996) and according to the institutional ethical
guidelines for animal experiments. Female BALB/c mice between 6 and
8 weeks of age, with a median weight of 20 g, were purchased from
the Laboratory Animal Center of Medical College, Tianjin University
(Tianjin, China). All mice were housed in a certified specific
pathogen-free animal facility, fed with regular rat chow and
maintained under optimal temperature (22–23°C), light (12-h
light/12-h dark cycle), oxygen, humidity (60%) and ventilation
conditions until sacrifice.
hUMSC/IL-18 preparation
The umbilical cord was obtained from a healthy
mother, aged 27 years, following the birth of a healthy term
newborn, with no family history of genetic disease, no cancer
history, and no presence of hepatitis B virus, hepatitis C virus,
human immunodeficiency virus, Epstein-Barr virus, cytomegalovirus
or syphilis in serum. Collection of the umbilical cord was approved
by the Institutional Medical Research Ethics Committee of Qingdao
Maternity Hospital (Shangdong, China). Written informed consent was
obtained from the mother 2 weeks prior to delivery.
The preparation of hUMSCs was performed in the
laminar flow laboratory, as previously reported (15). A lentivirus construct containing a
green fluorescent protein (GFP) gene and mouse IL-18 gene or
lentivirus construct containing a GFP gene only (Shanghai
GenePharma Co., Ltd., Shanghai, China) was used for the
transduction of the hUMSCs. Lentiviruses were added to the medium
at room temperature to infect the MSCs at 70 plaque-forming
units/cell, and the transfection medium was removed 24 h later.
Effective transduction was confirmed using a human IL-18 ELISA kit
(cat. no. KB1138; Shanghai Kaibo Biochemical Reagent Co., Ltd.,
Shanghai, China) to determine IL-18 levels in the culture
supernatant.
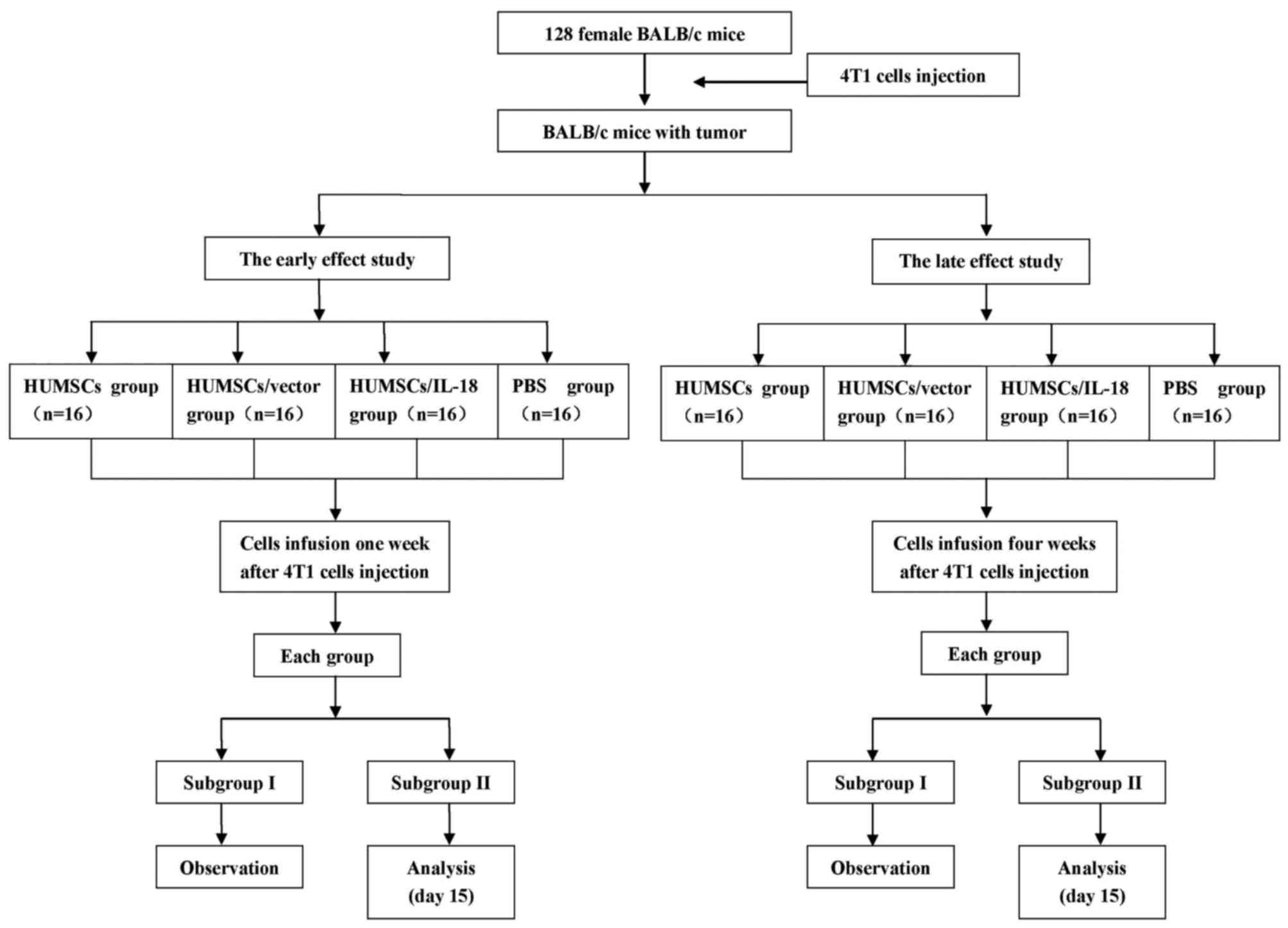
Experimental plan
Fig. 1 outlines the
plan of the experiment. 4T1 cells were administered as a suspension
of 1×106 cells in 100 µl PBS s.c. into the flank of mice
to prepare the breast cancer model. The model was considered
successful when the tumor size was >2×2 mm. In order to
investigate the early and the late effects of hUMSC/IL-18
transduction on breast cancer, suspensions of hUMSCs, hUMSCs/vector
or hUMSCs/IL-18 in 200 µl PBS, or 200 µl PBS alone, were injected
into each mouse group 1 and 4 weeks after injection of 4T1 cells.
Tumor progression and the behavior of the mice were recorded over
the course of the experiment.
For the early-effect and late-effect studies, mice
with tumors were randomly divided into 4 groups: The PBS group
(n=16), in which mice were injected with 200 µl PBS through the
lateral tail vein; the hUMSC group (n=16), in which mice were
injected with a suspension of 1×106 hUMSCs in 200 µl
PBS; the hUMSC/vector group (n=16), in which mice were injected
with a suspension of 1×106 hUMSCs/vector in 200 µl PBS;
and the hUMSC/IL-18 group (n=16), in which mice were injected with
a suspension of 1×106 hUMSCs/IL-18 in 200 µl PBS. Cell
suspensions were administered using a 26G needle via the lateral
tail vein.
Assessment of antitumor effect
Tumor dimensions (length, height and width) were
measured twice a week using calipers (Dwyer Instruments, Inc.,
Michigan City, IN, USA) by a researcher blinded to the treatment
group, and tumor volume was calculated according to the following
formula: Volume=lengthxwidth2/2. None of the mice had to
be sacrificed during the experiment due to tumor ulceration,
bleeding or a moribund state with excessive weight loss >25% of
the initial weight. At the endpoint of the experiment, animals were
sacrificed and tumors were excised. To detect IL-18 expression in
tissues, and alterations in immune cells and cytokines in
vivo, 8 mice from each group were sacrificed 1 week after cell
therapy to acquire blood and tissues for analysis. Each experiment
was performed at least in triplicate.
Spleen cell preparation and flow
cytometric analysis
Single-cell suspensions were obtained by passing
spleens through a 70-µm pore size cell strainer (Falcon; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), followed by lysis of
erythrocytes with red blood cell lysis buffer (Ortho-Clinical
Diagnostics GmbH, Neckargemünd, Germany). The splenocyte
suspensions (including lymphocytes and macrophages) were used for
flow cytometric analysis. Fluorescein isothiocyanate (FITC)-cluster
of differentiation (CD)4, phycoerythrin (PE)-CD8, FITC-CD3 and
PE-CD25 monoclonal antibodies (cat. no. CD3-4-8-A, a mix of
FITC-CD4, PE-CD8 and FITC-CD3; cat. no. CD3-25-A, a mix of FITC-CD3
and PE-CD25) were added to the splenocyte suspensions and incubated
for 30 min at 4°C. All antibodies were diluted at 1:200 and
purchased from eBioscience, Inc., San Diego, CA, USA. The cells
were then washed twice with PBS, fixed with 1% formaldehyde and
analyzed using a FACSCalibur flow cytometer with BD FACStation™
software version 6.1 (BD Biosciences, Franklin Lakes, NJ, USA).
Histological examination of tumor
tissue
Tissue samples from each mouse were divided into
thirds. The first third was saved for the analysis of cytokines.
Another third was fixed in 4% formaldehyde solution at room
temperature overnight and embedded in paraffin using the
conventional method, then cut into 4-µm-thick sections. Following
deparaffinization with xylene (10 min, 2 times) and rehydration
with a decreasing gradient concentration of ethanol (100, 90 and
70% for 5 min at each concentration), the sections were stained
with hematoxylin and eosin for gross histological examination and
immunohistochemistry.
The remaining third of the tumor tissues was
embedded in OCT medium (Tissue-Tek; Sakura Finetek USA, Inc.,
Torrance, CA, USA), snap-frozen immediately in liquid nitrogen and
stored at −80°C until use. For indirect immunofluorescence
analysis, 4-µm-thick cryosections were prepared, dried in air,
fixed in acetone, rehydrated in PBS and blocked using 10% goat
serum at 37°C for 60 min. Sections were incubated with primary
antibodies against CD3, CD8, CD16, CD56, CD80 and CD86 (all
eBioscience, Inc.) at 4°C overnight, followed by three washes in
PBS/1% bovine serum albumin (cat. no. 10437028; Gibco; Thermo
Fisher Scientific, Inc.). Subsequently, a FITC-conjugated goat
anti-mouse IgG secondary antibody was added (dilution, 1:100; cat.
no. 62-6511; Thermo Fisher Scientific, Inc.) for 60 min to detect
primary antibody binding, followed by three washes with PBS.
Sections were mounted using Prolong Gold anti-fade mounting medium
with DAPI (Invitrogen; Thermo Fisher Scientific, Inc.), examined
using a fluorescence microscope and acquired with SPOT software
(version 4.0.9; Diagnostic Instruments, Inc., Sterling Heights, MI,
USA).
Proliferation index assay of
proliferation marker protein Ki-67
An immunohistochemical procedure to detect Ki-67 was
performed and results were analyzed using the AxioVision Rel. 4.6
computerized image analysis system assisted by an automatic
measurement program (Zeiss AG, Oberkochen, Germany). Following
deparaffinization and rehydration, tissue sections were soaked in
3% H2O2 at 25°C for 10 min, then washed twice
in distilled water for 5 min, followed by incubation with an
anti-Ki-67 antibody (cat. no. MAB4190; dilution, 1:300;
Sigma-Aldrich; Merck KGaA) at 25°C for 60 min. Following washing
with PBS, sections were incubated with a peroxidase labeled goat
anti-mouse IgG secondary antibody (cat. no. A8924; dilution, 1:300;
Sigma-Aldrich; Merck KGaA) at 25°C for 45 min, prior to being
colored with DAB (Dako REAL™ EnVision™ Detection System),
counterstained with hematoxylin, dehydrated in a gradient
concentration of alcohol and mounted with neutral gum under a light
microscope. The stained sections were analyzed at ×200
magnification and 10 representative staining fields of each section
were analyzed to produce a mean optical density value, which
represented the strength of staining signals measured per positive
pixel. The mean absorbance data were analyzed to determine
statistical differences between groups of tissues.
CD31 examination
Sections were immunostained for CD31, to indicate
neovascularization, by incubating them with primary mouse
anti-CD31/PECAM-1 monoclonal antibody (dilution, 1:150; cat. no.
NB100-1642, Novus Biologicals, LLC, Littleton, CO, USA) and
biotinylated goat anti-mouse IgG (dilution, 1:150; cat. no.
NBP1-97590; Novus Biologicals, LLC). The sections were then
incubated with ExtrAvidin-horseradish peroxidase (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). Aminoethylcarbazole was used as a
chromogenic substrate using an Aminoethylcarbazole Staining kit
(Sigma-Aldrich; Merck KGaA). Microphotographs were captured, and 20
random fields of 3 stained sections (>4 fields/section) from
each group were observed at ×40 magnification from central healing
areas for semiquantitative analysis of microvessel density.
Negative control sections were incubated with PBS instead of the
primary antibody.
Determination of cytokine levels
Serum samples were obtained by centrifugation (5,000
× g for 7 min at room temperature) of blood (postmortem
intracardial puncture) from heparinized mice. A total of 4
cytokines, namely IL-18, IL-12, IFN-γ and TNF-α, from serum and
tumor tissue homogenates were measured using a quantitative
sandwich enzyme technique with Quantikine® ELISA kits
(BD Biosciences), according to the manufacturer's protocol. The
minimum measurable limit of each cytokine was 7.8 pg/ml for IL-12,
20 pg/ml for IFN-γ and TNF-α, and 1.0 pg/ml for IL-18. Hemolyzed
samples were excluded. Samples were assayed in duplicate and the
mean absorbance was calculated using a standard curve.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analysis was performed using GraphPad Prism software
(version 4.0; GraphPad Software, Inc., La Jolla, CA, USA).
Differences among three groups were analyzed by one-way analysis of
variance and Bonferroni's post-hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Mouse characteristics
All mice had developed tumors 1 week after injection
of 4T1 cells. Prior to cell therapy, no significant differences in
the tumor size, diet or vitality of the mice among different groups
were identified. Following cell therapy, the diet and vitality of
the mice in the hUMSC/IL-18 group were improved compared with the
PBS, hUMSC and hUMSC/vector groups.
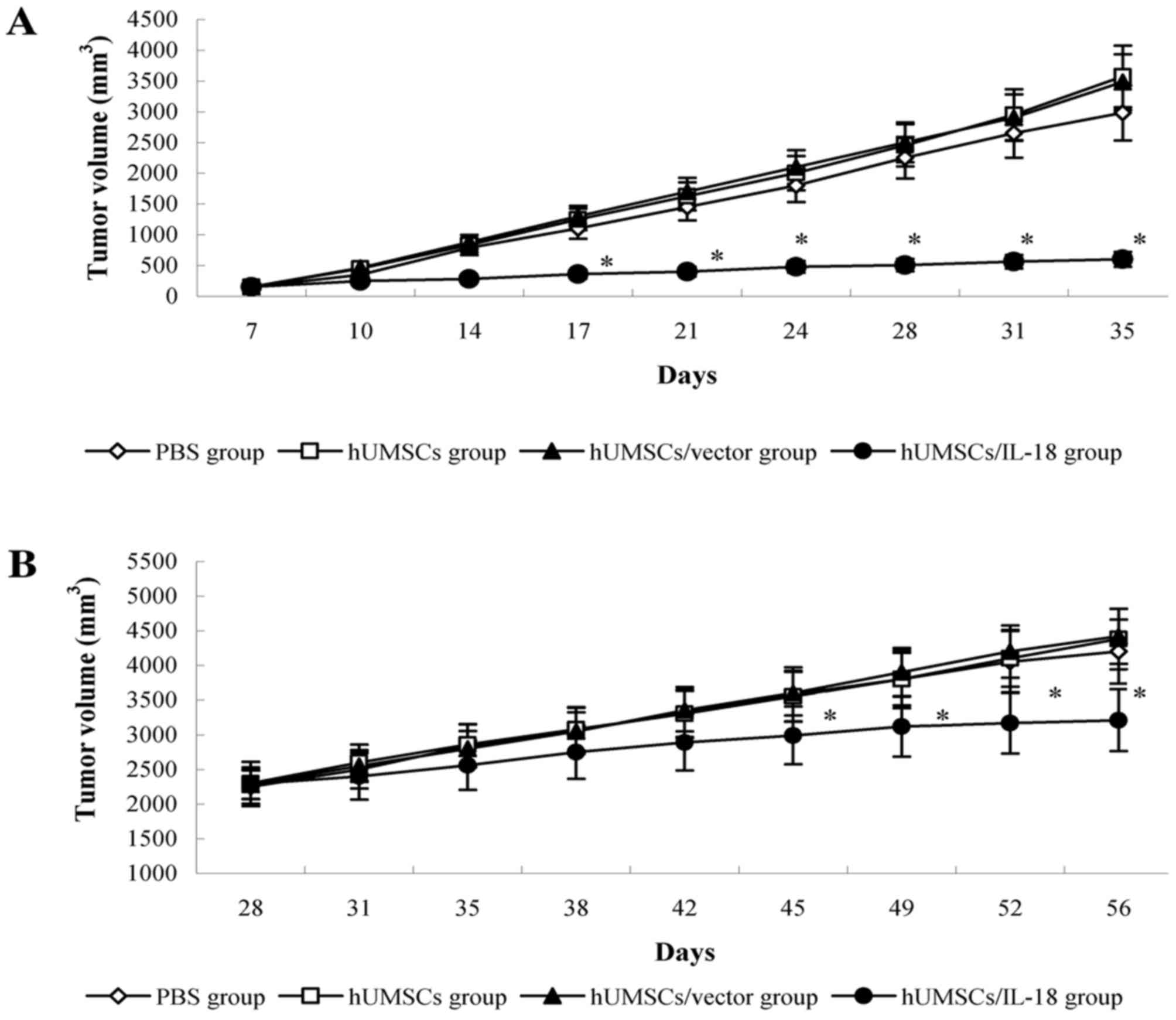
hUMSCs/IL-18 suppresses tumor
proliferation in vivo
The early effect of hUMSCs/IL-18 on breast cancer
was evaluated by transduction of hUMSCs/IL-18 1 week after
injection of 4T1 cells. Following therapy, proliferation of tumor
cells in the mice in the hUMSC/IL-18 group was markedly decreased.
There were significant differences in tumor volume between the
hUMSC/IL-18 group and the other groups from day 17 (P=0.035). For
the course of the study (day 84), the mean tumor volume in the
hUMSC/IL-18 group (0.58±0.29 mm3) was significantly
decreased compared with the other groups (P<0.01; Fig. 2). All mice in the hUMSC/IL-18 group
remained alive, whereas only 6/8 mice remained alive in the PBS
group, and only 4/8 mice remained alive in each of the hUMSC and
hUMSC/vector groups (P<0.05).
In the late-effect study, proliferation of the tumor
cells in the hUMSC/IL-18 group was slightly decreased compared with
the other groups following cell therapy. At day 45 of the study,
tumor volumes in the PBS, hUMSC and hUMSC/vector groups were
increased compared with the hUMSC/IL-18 group (P<0.05; Fig. 2). Until the end of the study (day 84),
6/8 mice remained alive in the hUMSC/IL-18 group, 5/8 mice remained
alive in the PBS group, and 4/8 mice remained alive in each of the
hUMSC and hUMSC/vector groups (P>0.05).
hUMSCs/IL-18 suppresses tumor
metastasis
The effect of transduction of hUMSCs/IL-18 on tumor
metastasis was evaluated by determining pulmonary and hepatic
metastasis in mice with breast cancer. In the early-effect study,
following cell therapy, tumor cell metastasis in the hUMSC/IL-18
group was markedly decreased. Until the end of study, 4/8 mice
exhibited pulmonary and hepatic metastasis in the hUMSC/IL-18
group, whereas all mice in the PBS, hUMSC and hUMSC/vector groups
exhibited pulmonary and hepatic metastasis (P=0.021). In the
late-effect study of hUMSC/IL-18 transduction on tumor metastasis,
no positive differences were identified in pulmonary and hepatic
metastasis in mice among the 4 groups.
Analysis of tumor tissue
In the early-effect study, using indirect
immunofluorescence analysis, lymphocytes that had infiltrated
tumors in the hUMSC/IL-18 group were significantly increased
compared with the PBS, hUMSC and hUMSC/vector groups.
CD3+ and CD8+ T cells that had infiltrated
tumors were significantly increased in the hUMSC/IL-18 group
compared with the other groups (P<0.01; Fig. 3). The number of CD16+,
CD56+, CD80+ and CD86+ cells in
the hUMSC/IL-18 group were significantly increased compared with
the other groups (P<0.05; Fig. 3).
These results indicated that hUMSCs/IL-18 effectively activated the
immunocytes to serve their antitumor roles. In the late-effect
study, numbers of CD3+ and CD8+ T cells, and
CD16+, CD56+, CD80+ and
CD86+ NK cells in the hUMSC/IL-18 group were
significantly increased compared with the other groups, but
decreased in comparison with those in the hUMSC/IL-18 group in the
early-effect study (all P<0.05; Fig.
3).
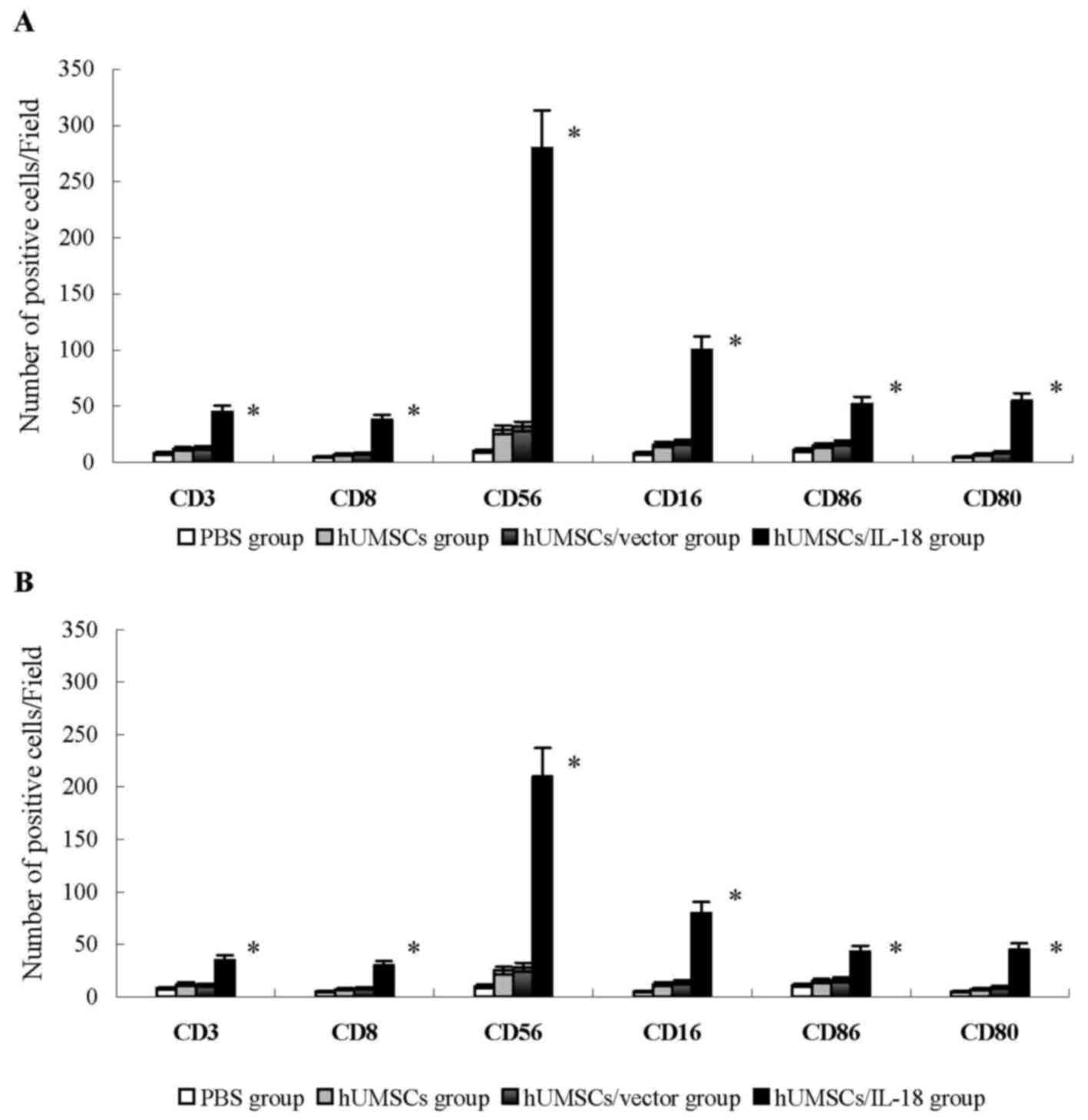 | Figure 3.Analysis of lymphocyte infiltration
into tumor tissues. Tumor tissues were snap-frozen, and 4-µm thick
sections were prepared, then stained with fluorescently labeled
antibodies. The number of CD3+ and CD8+ T
cells, and CD16+, CD56+, CD80+ and
CD86+ NK cells were quantified in 4 sections randomly
selected for each group. (A) In the early-effect study, the
proportions of CD3+ and CD8+ T cells, and
CD16+, CD56+, CD80+ and
CD86+ NK cells in the hUMSC/IL-18 group were
significantly increased compared with the other groups. (B) In the
late-effect study, the proportions of CD3+ and
CD8+ T cells, and CD16+, CD56+,
CD80+ and CD86+ NK cells in the hUMSC/IL-18
group were significantly increased compared with those in the other
groups, but were decreased compared with those in the hUMSC/IL-18
group in the early-effect study (P=0.039). *P<0.05 vs. all other
groups. CD, cluster of differentiation; hUMSCs, human mesenchymal
stem cells derived from umbilical cord; IL-18, interleukin 18; NK,
natural killer. |
The levels of cytokines, including IFN-γ, TNF-α,
IL-18 and IL-12, were examined in the tumor tissues of the mice 1
week after therapy. Compared with the PBS, hUMSC and hUMSC/vector
groups, the levels of these cytokines in the hUMSC/IL-18 group were
significantly increased (P<0.05; Fig.
4). Similarly, in the late-effect study, levels of IFN-γ,
TNF-α, IL-18 and IL-12 in the hUMSC/IL-18 group were significantly
increased compared with those in the other groups (P<0.05;
Fig. 4). No significant differences
in the levels of cytokines in the hUMSC/IL-18 group between the
early-effect study and the late-effect study were identified. These
results suggested that hUMSCs/IL-18 may induce the release of
cytokines to promote the antitumor effect.
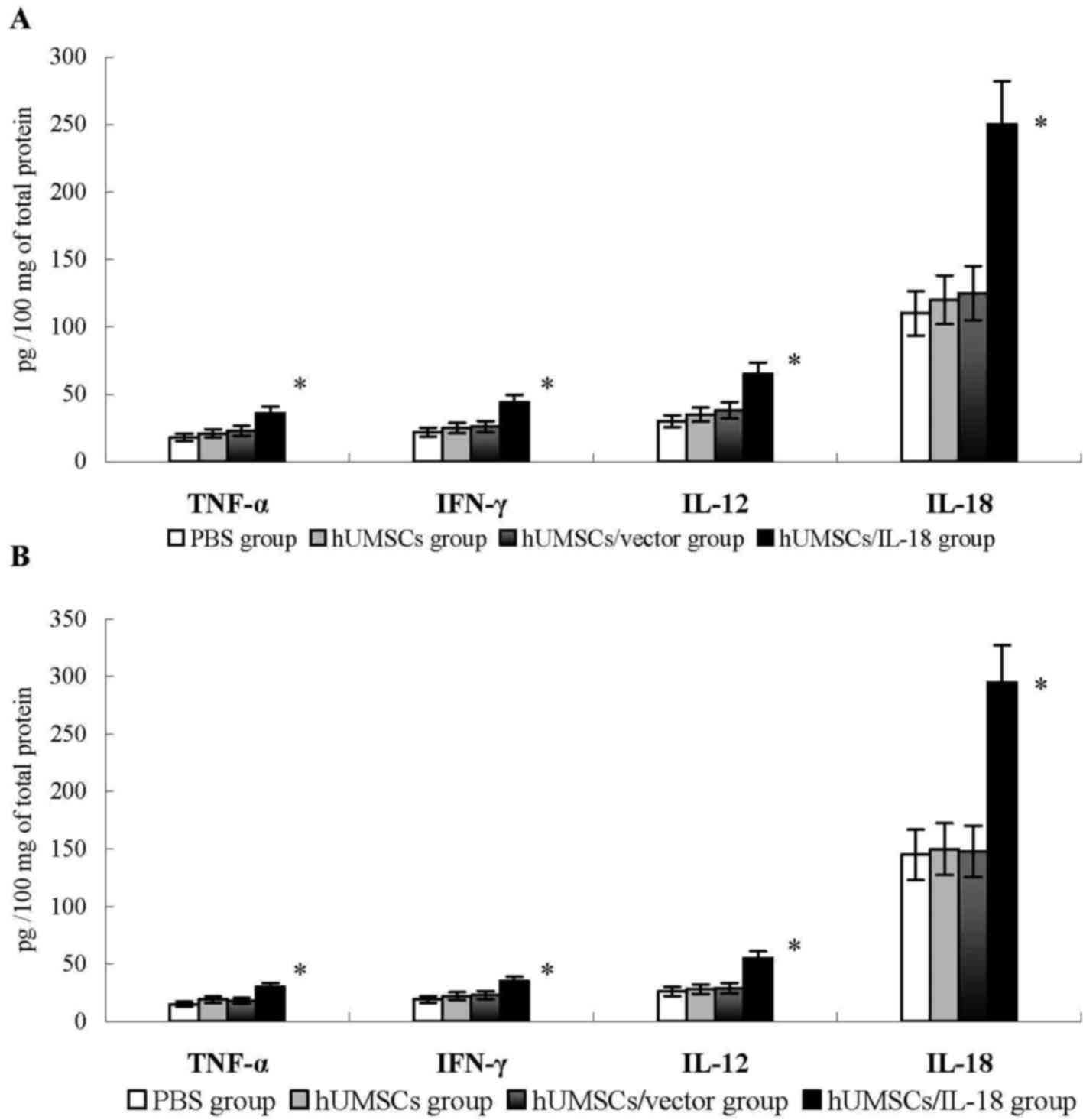 | Figure 4.Cytokine levels in tumor tissues. At
1 week after cell therapy, levels of cytokines in tumor tissues,
including IFN-γ, TNF-α, IL-18 and IL-12, were determined using
ELISA. (A) In the early-effect study, the levels of IFN-γ, TNF-α,
IL-18 and IL-12 in the hUMSC/IL-8 group were significantly
increased compared with levels in the PBS, hUMSC and hUMSC/vector
groups. (B) In the late-effect study, levels of IFN-γ, TNF-α, IL-18
and IL-12 in hUMSCs/IL-18 group in late infusion study were
significantly increased compared with levels in the PBS, hUMSC and
hUMSC/vector groups. No significant differences in the levels of
cytokines in the hUMSC/IL-18 group between the early-effect and the
late-effect study were identified. *P<0.05 vs. all other groups.
IFN-γ, interferon γ; TNF-α, tumor necrosis factor α; IL,
interleukin; hUMSCs, human mesenchymal stem cells derived from
umbilical cord. |
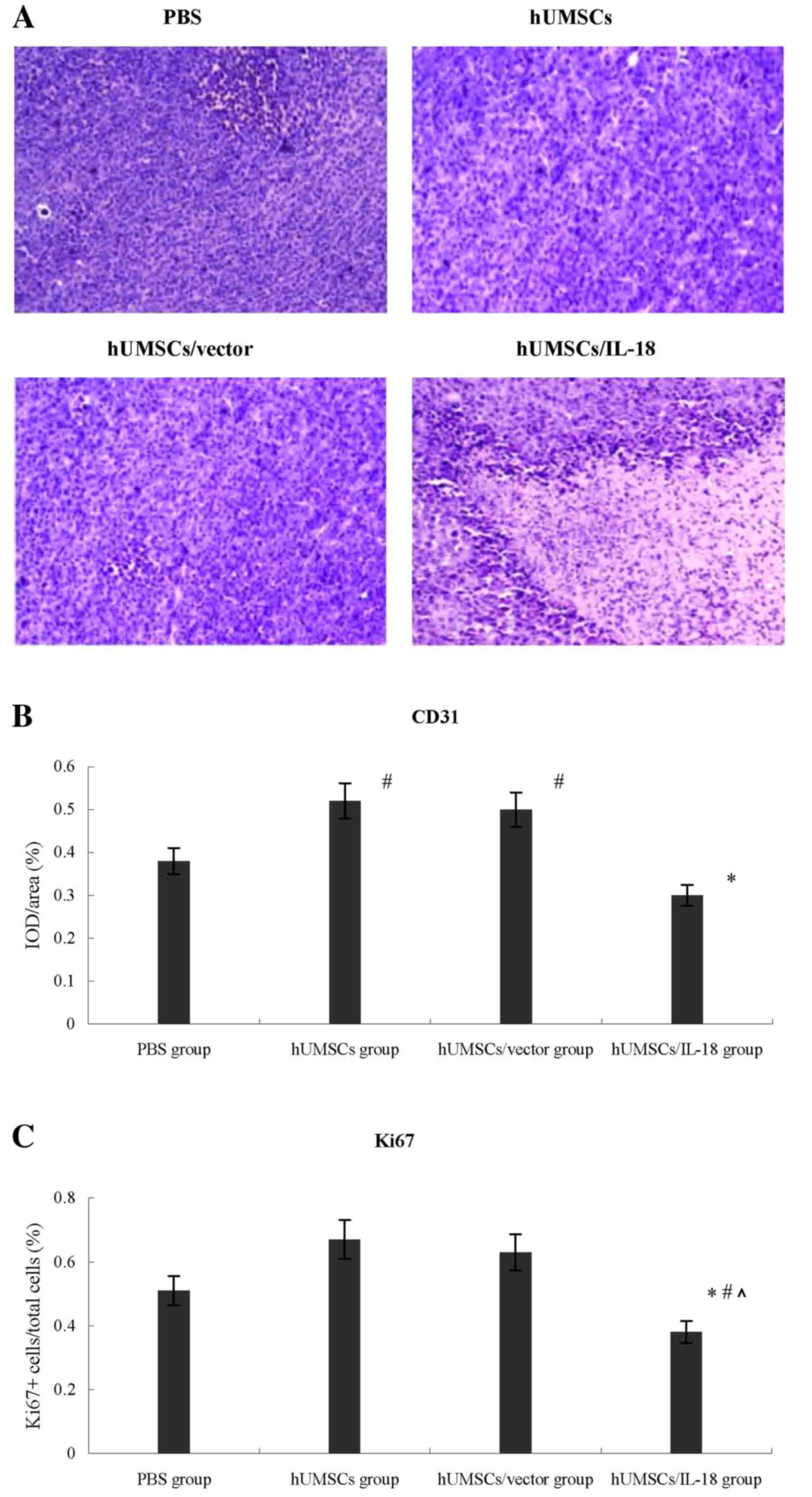
Lymphocyte infiltration was greater in the tumors of
the hUMSCs/IL-18 group compared with those in the other 3 groups,
which indicated a large number of tumor cells and minimal
lymphocyte infiltration (Fig. 5). The
effect of hUMSCs/IL-18 on tumor angiogenesis was investigated
further by examining the expression of CD31 in tumor tissues using
immunohistochemistry. The expression of CD31 in the hUMSC and
hUMSC/vector groups was significantly increased compared with the
PBS and hUMSC/IL-18 groups in the early-effect and the late-effect
studies, and expression of CD31 in the hUMSC/IL-18 group was
significantly decreased compared with that in the PBS group
(P<0.05; Fig. 5).
In order to evaluate the effect of hUMSCs/IL-18 on
the proliferation of breast cancer cells in vivo, the Ki-67
proliferation index was determined using immunohistochemistry. As
presented in Fig. 5, the median
percentage of proliferating Ki-67+ cells within the
tumors in the hUMSC/IL-18 group mice was decreased significantly
compared with the PBS group [0.38 (range, 0.35–0.67)
Ki-67+ cells/total cells in the hUMSC/IL-18 group vs.
0.51 (range, 0.42–0.75) in the PBS group, P=0.037], hUMSC group and
hUMSC/vector group [0.67 (range, 0.48–0.90) in the hUMSC group,
P=0.009; and 0.63 (range, 0.38–0.83) in the hUMSC/vector group,
P=0.004].
Immune system analysis
In order to compare the difference of levels of
cytokines between serum and tumor tissues, the serum levels of
cytokines, including IFN-γ, TNF-α, IL-18 and IL-12, were determined
in mice 1 week after therapy. Compared with the PBS, hUMSC and
hUMSC/vector groups, the serum levels of IL-18 and IL-12 in the
hUMSC/IL-18 group were significantly increased (P<0.05), whereas
no significant differences in the levels of IFN-γ and TNF-α among
the 4 groups were identified (Fig.
6). No significant differences in serum levels of IFN-γ, TNF-α,
IL-18 and IL-12 in the hUMSC/IL-18 group were identified between
the late-effect and the early-effect study (P>0.05; Fig. 6).
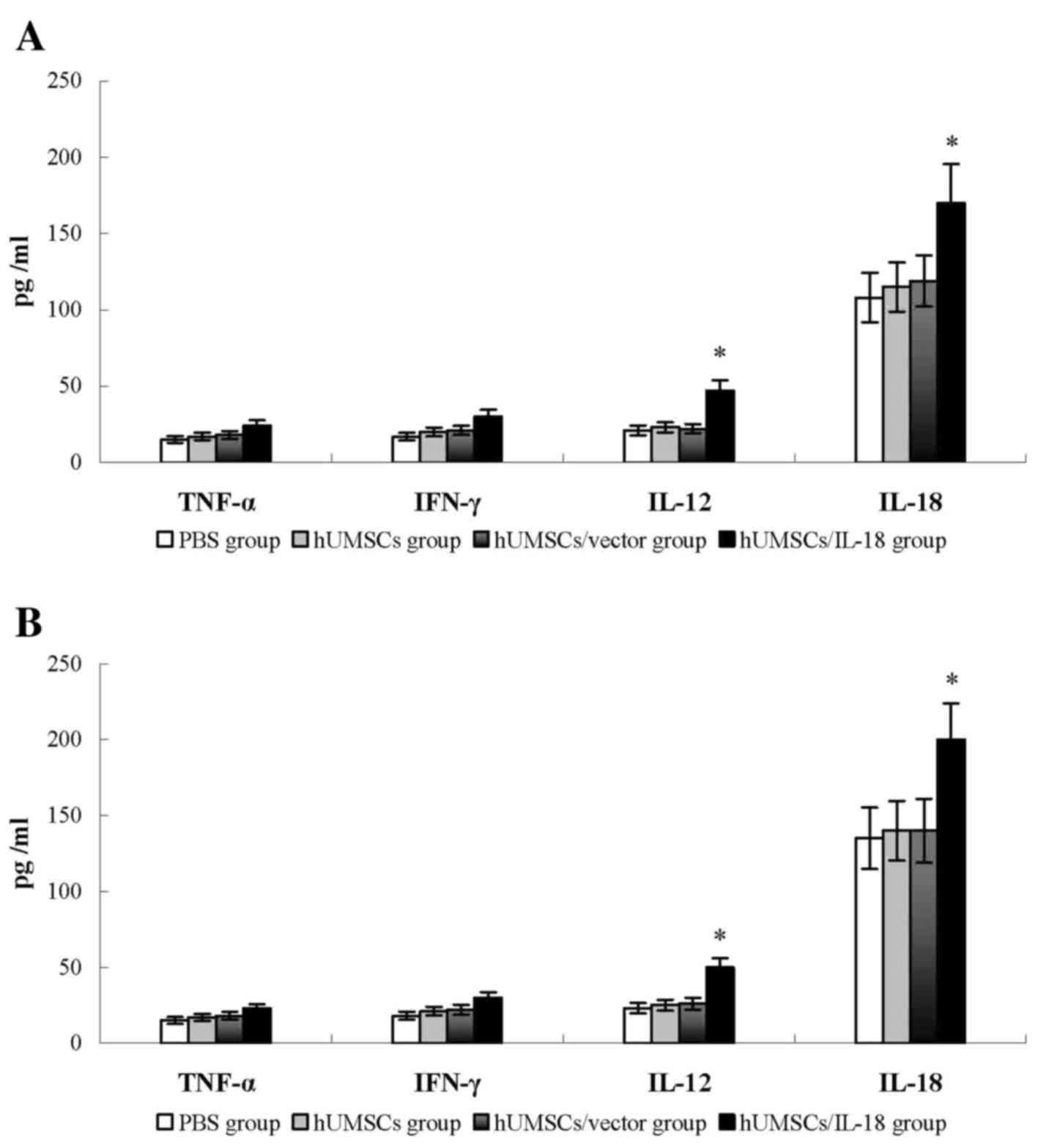 | Figure 6.Cytokine levels in peripheral blood.
At 1 week after cell therapy, serum levels of cytokines, including
IFN-γ, TNF-α, IL-18 and IL-12, were determined using ELISA. (A) In
the early-effect study, the serum levels of IL-18 and IL-12, but
not of TNF-α or IFN-γ, in the hUMSC/IL-18 group were significantly
increased compared with levels in the PBS, hUMSC and hUMSC/vector
groups. (B) In the late-effect study, the serum levels of IL-18 and
IL-12, but not of TNF-α or IFN-γ, in the hUMSC/IL-18 group were
significantly increased compared with levels in the PBS, hUMSC and
hUMSC/vector groups. No significant differences in serum levels of
IFN-γ, TNF-α, IL-18 and IL-12 in the hUMSC/IL-18 group between the
late-effect and the early-effect study were identified. *P<0.05,
vs. all other groups. IFN-γ, interferon γ; TNF-α, tumor necrosis
factor α; IL, interleukin; hUMSCs, human mesenchymal stem cells
derived from umbilical cord. |
Spleen lymphocytes of mice in the hUMSC/IL-18 group
were markedly activated following therapy (Fig. 7). At 1 week after cell therapy, in the
early-effect study, the proportion of CD3+ T cells in
the spleen lymphocytes of mice in the PBS, hUMSC, hUMSC/vector and
hUMSC/IL-18 groups was 69, 71, 72 and 97%, respectively. However,
in the late-effect study, the proportions of CD3+ T
cells in the spleen lymphocytes of mice in the PBS, hUMSC,
hUMSC/vector and hUMSC/IL-18 groups were 52.8, 56.6, 58.7 and
85.7%, respectively. Thus, the activation of immunocytes in the
hUMSC/IL-18 group in the early-effect study was increased compared
with that in the late-effect study (P=0.039). The percentage of
CD3+ T cells was markedly increased in the hUMSC/IL-18
group compared with the PBS group (P=0.031; Fig. 7). Subpopulations of CD4+
and CD8+ T cells in the 4 groups were analyzed. Compared
with the other groups, the percentage of
CD3+CD8+ T cells in the hUMSC/IL-18 group was
significantly increased (P=0.001; Fig.
7). For CD4+CD25+ T cells, the proportion
was significantly decreased in the hUMSC/IL-18 group compared with
the PBS, hUMSC and hUMSC/vector groups (P=0.032; Fig. 7). In the late-effect study, the
proportion of CD3+ T cells in the hUMSC/IL-18 group was
increased slightly compared with the other groups; however, no
significant difference was identified (P>0.05; Fig. 7).
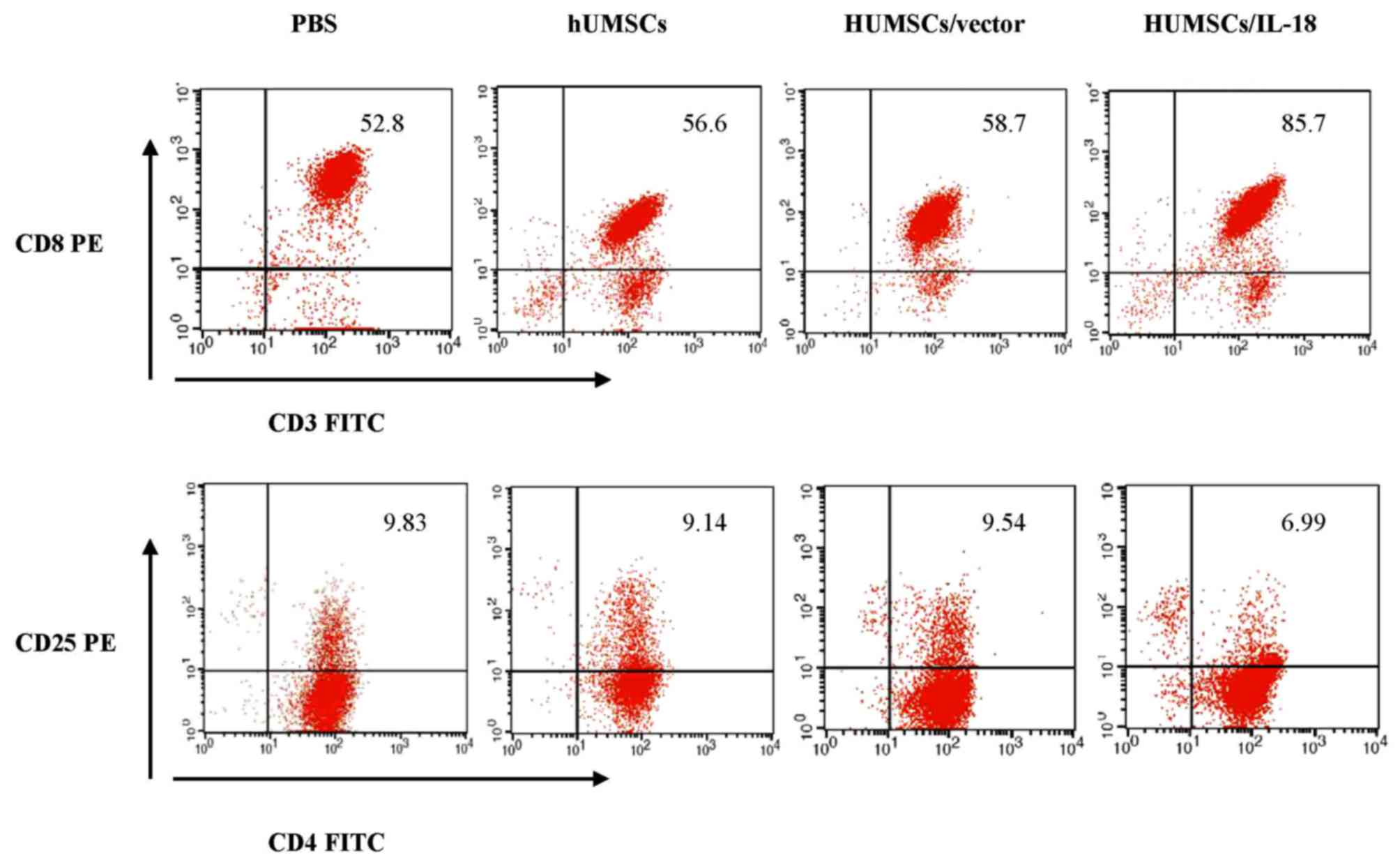 | Figure 7.Flow cytometric analysis of spleen
lymphocyte subpopulations. At 1 week after cell therapy, 8 mice in
each group were sacrificed to collect spleen lymphocytes for
analysis. The proportion of CD3+CD8+ T cells
in the hUMSC/IL-18 group was significantly increased compared with
the PBS, hUMSC and hUMSC/vector groups (P=0.007, 0.023 and 0.039,
respectively). Conversely, the proportion of
CD4+CD25+ T cells in the hUMSC/IL-18 group
was significantly decreased compared with the PBS, hUMSC and
hUMSC/vector groups (P=0.009, 0.035 and 0.021, respectively). CD,
cluster of differentiation; hUMSCs, human mesenchymal stem cells
derived from umbilical cord; IL-18, interleukin 18; PE,
phyocerythrin; FITC, fluorescein isothiocyanate. |
Side effects
During the therapy and observation, no abnormality
was identified at the sites of injection by macroscopic
observations. No acute or chronic side effects were observed in the
mice in the hUMSC/IL-18, hUMSC or hUMSC/vector groups compared with
the PBS group.
Discussion
IL-18 serves important roles in cancer progression
and metastasis. hUMSCs/IL-18 were able to suppress the
proliferation, migration and invasion of MCF-7 cells and HCC1937
cells in vitro in our previous study (19). In the present study, the effect of
hUMSCs/IL-18 on breast cancer in a mouse model was further
investigated, and it was identified that hUMSCs/IL-18 served
antitumor roles in vivo, including the expression of IL-18
protein, the suppression of tumor proliferation and metastasis by
activating immunocytes and immune cytokines, decreasing the Ki-67
proliferation index of tumor cells and inhibiting tumor
angiogenesis.
It has been demonstrated that MSCs are able to be
recruited from the systemic circulation to the stroma of a number
of types of growing tumors, including melanoma, glioma, breast
cancer and pancreatic cancer (20–23). In
tumor sites, MSCs are able to enhance tumor proliferation,
invasiveness and the formation of metastases through multiple
mechanisms. In addition to differentiating into tumor endothelial
cells, pericytes, smooth muscle cells and cancer-associated
fibroblasts, MSCs may assist in promoting tumor proliferation by
stimulating tumor cells directly in a paracrine fashion (20,22). In
the present study, hUMSCs and hUMSCs/vector were able to promote
tumor proliferation and metastasis in BALB/c mice with breast
cancer. However, the tropism of MSCs to growing tumors also
represented a powerful tool to deliver therapeutic substances to
these tumors. The transduction of hUMSCs/IL-18 into mice led to
significantly reduced tumor proliferation as compared with mice
receiving hUMSCs or hUMSCs/vector alone. In the present study, in
the hUMSC/IL-18 group, tumor regression was significant from day
17; this result was consistent with the study by Müller et
al (24), which demonstrated that
IL-18-encoding plasmid DNA exhibited antitumor effects in B16
melanoma and that tumor regression was significant on day 15 of the
treatment cycle in the IL-18 group. Furthermore, recombinant murine
IL-18 was demonstrated to be effective in B16 melanoma even when
the mice were depleted of T cells and NK cells (25,26).
The quantity and quality of immunocyte infiltration
into the tumor environment, including the critical balance between
effector and regulatory T cells, have been recognized as vital
components of spontaneous and therapy-induced antitumor immune
control (27,28). In the present study, following
transduction of hUMSCs/IL-18 into mice, the number of immune cells
that infiltrated into the tumor site and were activated increased.
The proportions of CD3+ and CD8+ T cells, and
CD16+, CD56+, CD80+ and
CD86+ NK cells in the hUMSC/IL-18 group were increased
compared with the other groups. These results indicated that
hUMSCs/IL-18 were able to serve antitumor roles by stimulating the
cytotoxic activity of NK and T cells. Furthermore, levels of
cytokines, including IFN-γ, TNF-α, IL-18 and IL-12, in tumor
tissues were also increased in the hUMSC/IL-18 group. IL-18 alone
or in synergy with other cytokines, including IL-12, modulated the
differentiation, proliferation, function and survival of immune
cells (29,30). NK cells activated by IL-18 were able
to induce the activation of dendritic cells (DCs) and potentiate
DC-mediated induction of tumor-specific Th1 and cytotoxic T
lymphocyte adaptive immune responses through an IFN-γ- and
TNF-α-dependent mechanism (31,32). NK
cells may act as the inducers of local immune cell accumulation,
promoting the CCR5-dependent attraction of immature DCs and driving
subsequent DC production of the effector CD8+ T
cell-recruiting chemokines C-X-C motif chemokine ligand (CXCL)9,
CXCL10 and CCL5, in cells isolated from the blood of healthy donors
and in tumor-associated cells isolated from the malignant ascites
of patients with advanced (stage III–IV) cancer (33).
It was previously reported that IL-18 was able to
inhibit the proliferation of tumor cells by inhibiting tumor
angiogenesis (34). CD31 is
considered a marker that indicates the degree of tumor angiogenesis
(35). In the present study,
immunohistochemical staining of CD31 indicated that treatment with
hUMSCs/IL-18 resulted in reduced vessel formation compared with
treatment with hUMSCs or hUMSCs/vector. Furthermore, the treatment
of mice with hUMSCs/IL-18 led to a significantly decreased Ki-67
proliferation index in tumors compared with that in mice receiving
hUMSCs or hUMSCs/vector alone. In our previous in vitro
study (19), it was demonstrated that
hUMSCs/IL-18 were able to induce the G1- to S-phase arrest of
breast cancer cells. These results indicated that hUMSCs/IL-18 were
able to suppress tumor proliferation by inhibiting tumor
angiogenesis and decreasing Ki-67 proliferation indices in
tumors.
As a main cause of cancer mortality, metastasis is a
complex process involving a range of overlapping events, including
cancer cell attachment to the extracellular matrix, and cell
invasion and migration (36). Yamada
et al (37) demonstrated that
IL-18 suppressed the pulmonary metastasis of osteosarcoma cells,
independently of T and NK cells, by inducing one or more host
factors that suppressed cell mobility to inhibit the settlement of
osteosarcoma cells in the lung. Furthermore, previous studies have
demonstrated that increased IL-18 serum levels accompany tumor
progression and have a negative prognostic impact on patients with
cancer (38–40). In the present early-effect study,
transduction of hUMSCs/IL-18 into mice significantly suppressed the
pulmonary metastasis and hepatic metastasis of breast cancer,
whereas in the late-effect study, the suppressing effect on tumor
metastasis was not evident. The underlying molecular mechanisms
remain unclear; however, it has been demonstrated previously that
IL-18 alone accelerated tumor progression in the absence of
Th1-like cytokines, in part through cell autonomous effects on
cancer and endothelial cells (41).
In addition, low doses of IL-18 were able to mediate
immunosuppression on the NK cell arm of immunity (42,43).
In the present study, the difference between
cytokines and immunocytes that had infiltrated into tumor tissues
and into the systemic circulation was compared. The levels of
cytokines and immunocytes that had infiltrated into tumor tissues
were increased compared with those that had infiltrated into the
systemic circulation. The levels of cytokines in tumor tissues were
increased ~1.5-fold compared with those in the systemic
circulation. This result may be due to the locally increased
concentration of IL-18 and partial release into the bloodstream,
and also indicated that increased numbers of hUMSCs/IL-18 migrated
to the tumor sites to serve this role. The early effect and the
late effect of hUMSCs/IL-18 on breast cancer were also
investigated. For the early effect, hUMSCs/IL-18 significantly
suppressed tumor proliferation and tumor metastasis, whereas for
the late effect, the therapeutic effect was not as marked as the
early effect, particularly for tumor metastasis. This result
indicated that the early effect of treatment offered an improvement
over the late effect of treatment.
The results of the present study identified that
hUMSCs/IL-18 were able to inhibit the proliferation and metastasis
of tumor cells in a mouse model of breast cancer. hUMSCs/IL-18
suppressed the proliferation of tumor cells by activating
immunocytes and immune cytokines, decreasing the Ki-67
proliferation index of tumor cells and inhibiting tumor
angiogenesis. Furthermore, hUMSCs/IL-18 were able to induce an
improved therapeutic effect in the tumor sites, particularly in
early tumors. However, the present study represents a preliminary
study, and further investigations are required to identify how to
increase the expression of IL-18, how to determine the optimal
number and duration of cell infusion for cancer therapy, and how to
combine cell therapy with chemotherapy or radiotherapy,
particularly for advanced tumors.
Acknowledgements
The present study was supported by the Department of
Central Laboratory, The Affiliated Hospital of Qingdao University,
and by the Shandong Provincial Natural Science Foundation (grant
no. 2013ZRB01426).
References
|
1
|
Schrijver WAME, Schuurman K and van Rossum
A: Dutch Distant Breast Cancer Metastases Consortium, Peeters T,
Ter Hoeve N, Zwart W, van Diest PJ and Moelans CB: Loss of steroid
hormone receptors is common in malignant pleural and peritoneal
effusions of breast cancer patients treated with endocrine therapy.
Oncotarget. 8:55550–55561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zeidan AM, Long JB, Wang R, Hu X, Yu JB,
Huntington SF, Abel GA, Mougalian SS, Podoltsev NA, Gore SD, et al:
Risk of myeloid neoplasms after radiotherapy among older women with
localized breast cancer: A population-based study. PLoS One.
12:e01847472017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahn JO, Lee HW, Seo KW, Kang SK, Ra JC and
Youn HY: Anti-tumor effect of adipose tissue derived-mesenchymal
stem cells expressing interferon-β and treatment with cisplatin in
a xenograft mouse model for canine melanoma. PLoS One.
8:e748972013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ryu H, Oh JE, Rhee KJ, Baik SK, Kim J,
Kang SJ, Sohn JH, Choi E, Shin HC, Kim YM, et al: Adipose
tissue-derived mesenchymal stem cells cultured at high density
express IFN-β and suppress the growth of MCF-7 human breast cancer
cells. Cancer Lett. 352:220–227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee DH, Ahn Y, Kim SU, Wang KC, Cho BK,
Phi JH, Park IH, Black PM, Carroll RS, Lee J and Kim SK: Targeting
rat brainstem glioma using human neural stem cells and human
mesenchymal stem cells. Clin Cancer Res. 15:4925–4934. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Knoop K, Schwenk N, Dolp P, Willhauck MJ,
Zischek C, Zach C, Hacker M, Göke B, Wagner E, Nelson PJ and
Spitzweg C: Stromal targeting of sodium iodide symporter using
mesenchymal stem cells allows enhanced imaging and therapy of
hepatocellular carcinoma. Hum Gene Ther. 24:306–316. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Payne NL, Sun G, McDonald C, Layton D,
Moussa L, Emerson-Webber A, Veron N, Siatskas C, Herszfeld D, Price
J and Bernard CC: Distinct immunomodulatory and migratory
mechanisms underpin the therapeutic potential of human mesenchymal
stem cells in autoimmune demyelination. Cell Transplant.
22:1409–1425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Molloy AP, Martin FT, Dwyer RM, Griffin
TP, Murphy M, Barry FP, O'Brien T and Kerin MJ: Mesenchymal stem
cell secretion of chemokines during differentiation into
osteoblasts, and their potential role in mediating interactions
with breast cancer cells. Int J Cancer. 124:326–332. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dwyer RM, Potter-Beirne SM, Harrington KA,
Lowery AJ, Hennessy E, Murphy JM, Barry FP, O'Brien T and Kerin MJ:
Monocyte chemotactic protein-1 secreted by primary breast tumors
stimulates migration of mesenchymal stem cells. Clin Cancer Res.
13:5020–5027. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Macho-Fernandez E, Cruz LJ, Ghinnagow R,
Fontaine J, Bialecki E, Frisch B, Trottein F and Faveeuw C:
Targeted delivery of α-galactosylceramide to CD8α+ dendritic cells
optimizes type I NKT cell-based antitumor responses. J Immunol.
193:961–969. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Keyhani A, Riazi-Rad F, Pakzad SR and
Ajdary S: Human polymorphonuclear leukocytes produce cytokines in
response to Leishmania major promastigotes. APMIS. 122:891–897.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun H, Sun C and Xiao W: Expression
regulation of co-inhibitory molecules on human natural killer cells
in response to cytokine stimulations. Cytokine. 65:33–41. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim HX, Hong HJ, Cho D and Kim TS: IL-18
enhances immunosuppressive responses by promoting differentiation
into monocytic myeloid-derived suppressor cells. J Immunol.
193:5453–5460. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jarry A, Malard F, Bou-Hanna C, Meurette
G, Mohty M, Mosnier JF, Laboisse CL and Bossard C: Interferon-alpha
promotes Th1 response and epithelial apoptosis via inflammasome
activation in human intestinal mucosa. Cell Mol Gastroenterol
Hepatol. 3:72–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Serti E, Werner JM, Chattergoon M, Cox AL,
Lohmann V and Rehermann B: Monocytes activate natural killer cells
via inflammasome-induced interleukin 18 in response to hepatitis C
virus replication. Gastroenterology. 147:209–220.e3. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakata A, Tsujimura T, Sugihara A, Okamura
H, Iwasaki T, Shinkai K, Iwata N, Kakishita E, Akedo H and Terada
N: Inhibition by interleukin 18 of osteolytic bone metastasis by
human breast cancer cells. Anticancer Res. 19:4131–4138.
1999.PubMed/NCBI
|
|
17
|
Coskun U, Gunel N, Sancak B, Onuk E,
Bayram M and Cihan A: Effect of tamoxifen on serum IL-18, vascular
endothelial growth factor and nitric oxide activities in breast
carcinoma patients. Clin Exp Immunol. 137:546–551. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye ZB, Ma T, Li H, Jin XL and Xu HM:
Expression and significance of intratumoral interleukin-12 and
interleukin-18 in human gastric carcinoma. World J Gastroenterol.
13:1747–1751. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Hu J, Sun S, Li F, Cao W, Wang YU,
Ma Z and Yu Z: Mesenchymal stem cells expressing interleukin-18
suppress breast cancer cells in vitro. Exp Ther Med. 9:1192–1200.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Doucette T, Rao G, Yang Y, Gumin J,
Shinojima N, Bekele BN, Qiao W, Zhang W and Lang FF: Mesenchymal
stem cells display tumor-specific tropism in an RCAS/Ntv-a glioma
model. Neoplasia. 13:716–725. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sage EK, Kolluri KK, McNulty K, Lourenco
Sda S, Kalber TL, Ordidge KL, Davies D, Lee Gary YC, Giangreco A
and Janes SM: Systemic but not topical TRAIL-expressing mesenchymal
stem cells reduce tumour growth in malignant mesothelioma. Thorax.
69:638–647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Esposito M and Kang Y: Targeting
tumor-stromal interactions in bone metastasis. Pharmacol Ther.
141:222–233. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moniri MR, Sun XY, Rayat J, Dai D, Ao Z,
He Z, Verchere CB, Dai LJ and Warnock GL: TRAIL-engineered
pancreas-derived mesenchymal stem cells: Characterization and
cytotoxic effects on pancreatic cancer cells. Cancer Gene Ther.
19:652–658. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Müller J, Feige K, Wunderlin P, Hödl A,
Meli ML, Seltenhammer M, Grest P, Nicolson L, Schelling C and
Heinzerling LM: Double-blind placebo-controlled study with
interleukin-18 and interleukin-12-encoding plasmid DNA shows
antitumor effect in metastatic melanoma in gray horses. J
Immunother. 34:58–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Choi IK, Lee JS, Zhang SN, Park J, Sonn
CH, Lee KM and Yun CO: Oncolytic adenovirus co-expressing IL-12 and
IL-18 improves tumor-specific immunity via differentiation of T
cells expressing IL-12Rβ2 or IL-18Rα. Gene Ther. 18:898–909. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang J, Jin G, Liu X and Liu S:
Therapeutic effect of pEgr-IL18-B7.2 gene radiotherapy in B16
melanoma-bearing mice. Hum Gene Ther. 18:323–332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Galon J, Costes A, Sanchez-Cabo F,
Kirilovsky A, Mlecnik B, Lagorce-Pagès C, Tosolini M, Camus M,
Berger A, Wind P, et al: Type, density, and location of immune
cells within human colorectal tumors predict clinical outcome.
Science. 313:1960–1964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sato E, Olson SH, Ahn J, Bundy B,
Nishikawa H, Qian F, Jungbluth AA, Frosina D, Gnjatic S, Ambrosone
C, et al: Intraepithelial CD8 tumor-infiltrating lymphocytes and a
high CD8t/regulatory T cell ratio are associated with favorable
prognosis in ovarian cancer. Proc Natl Acad Sci USA.
102:18538–18543. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rodrigues DR, Fernandes RK, Balderramas
Hde A, Penitenti M, Bachiega TF, Calvi SA, Dias-Melicio LA, Ikoma
MR and Soares ÂM: Interferon-gamma production by human neutrophils
upon stimulation by IL-12, IL-15 and IL-18 and challenge with
Paracoccidioides brasiliensis. Cytokine. 69:102–109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ruiz C, Pérez E, García-Martínez O,
Díaz-Rodríguez L, Arroyo-Morales M and Reyes-Botella C: Expression
of cytokines IL-4, IL-12, IL-15, IL-18, and IFNgamma and modulation
by different growth factors in cultured human osteoblast-like
cells. J Bone Miner Metab. 25:286–292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mailliard RB, Alber SM, Shen H, Watkins
SC, Kirkwood JM, Herberman RB and Kalinski P: IL-18-induced CD83+
CCR7+ NK helper cells. J Exp Med. 202:941–953. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mailliard RB, Son YI, Redlinger R, Coates
PT, Giermasz A, Morel PA, Storkus WJ and Kalinski P: Dendritic
cells mediate NK cell help for Th1 and CTL responses: Two-signal
requirement for the induction of NK cell helper function. J
Immunol. 171:2366–2373. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wong JL, Muthuswamy R, Bartlett DL and
Kalinski P: IL-18-based combinatorial adjuvants promote the
intranodal production of CCL19 by NK cells and dendritic cells of
cancer patients. Oncoimmunology. 2:e262452013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhong L, Roybal J, Chaerkady R, Zhang W,
Choi K, Alvarez CA, Tran H, Creighton CJ, Yan S, Strieter RM, et
al: Identification of secreted proteins that mediate cell-cell
interactions in an in vitro model of the lung cancer
microenvironment. Cancer Res. 68:7237–7245. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ribatti D, Belloni AS, Nico B, Di Comite
M, Crivellato E and Vacca A: Leptin-leptin receptor are involved in
angiogenesis in human hepatocellular carcinoma. Peptides.
29:1596–1602. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gray-Schopfer V, Wellbrock C and Marais R:
Melanoma biology and new targeted therapy. Nature. 445:851–857.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamada N, Hata M, Ohyama H, Yamanegi K,
Kogoe N, Nakasho K, Futani H, Okamura H and Terada N: Immunotherapy
with interleukin-18 in combination with preoperative chemotherapy
with ifosfamide effectively inhibits postoperative progression of
pulmonary metastases in a mouse osteosarcoma model. Tumour Biol.
30:176–184. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pappa CA, Alexandrakis MG, Boula A,
Psarakis FE, Kolovou A, Bantouna V, Stavroulaki E and Tsirakis G:
Emerging roles of endoglin/CD105 and angiogenic cytokines for
disease development and progression in multiple myeloma patients.
Hematol Oncol. 31:201–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dwivedi S, Goel A, Natu SM, Mandhani A,
Khattri S and Pant KK: Diagnostic and prognostic significance of
prostate specific antigen and serum interleukin 18 and 10 in
patients with locally advanced prostate cancer: A prospective
study. Asian Pac J Cancer Prev. 12:1843–1848. 2011.PubMed/NCBI
|
|
40
|
Goto N, Tsurumi H, Kasahara S, Kanemura N,
Hara T, Yasuda I, Shimizu M, Murakami N, Sawada M, Yamada T, et al:
Serum interleukin-18 level is associated with the outcome of
patients with diffuse large B-cell lymphoma treated with CHOP or
R-CHOP regimens. Eur J Haematol. 87:217–227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vidal-Vanaclocha F, Mendoza L, Telleria N,
Salado C, Valcárcel M, Gallot N, Carrascal T, Egilegor E,
Beaskoetxea J and Dinarello CA: Clinical and experimental
approaches to the pathophysiology of interleukin-18 in cancer
progression. Cancer Metastasis Rev. 25:417–434. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Terme M, Ullrich E, Aymeric L, Meinhardt
K, Desbois M, Delahaye N, Viaud S, Ryffel B, Yagita H, Kaplanski G,
et al: IL-18 induces PD-1-dependent immunosuppression in cancer.
Cancer Res. 71:5393–5399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Terme M, Ullrich E, Aymeric L, Meinhardt
K, Coudert JD, Desbois M, Ghiringhelli F, Viaud S, Ryffel B, Yagita
H, et al: Cancer-induced immunosuppression: IL-18-elicited
immunoablative NK cells. Cancer Res. 72:2757–2767. 2012. View Article : Google Scholar : PubMed/NCBI
|