Introduction
Multiple myeloma (MM) is characterized by the
accumulation of malignant plasma cells within the bone marrow (BM)
and is the second most common type of hematological cancer,
accounting for 10–15% of all types of hematological cancer. At
present, chemotherapy is one of the primary treatments for MM,
including newly marketed proteasome inhibitors and
immunomodulators. However, MM remains incurable in the majority of
patients, with a median survival time of 3–5 years with
conventional treatments. All current drugs exhibit limited efficacy
and/or severe toxicity or adverse effects (1–4).
Therefore, novel agents and combinations are required to further
improve the survival of patients with MM.
Previous studies have indicated that the
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling
pathway and mammalian target of rapamycin (mTOR) are aberrantly
activated in myeloma cells (5,6). mTOR, a
serine (Ser)/threonine (Thr) protein kinase expressed in the PI3K
pathway, is not only a possible valuable target for the treatment
of MM (7), but also has a vital
function in regulating critical cellular processes, including
transcription, ribosomal biogenesis, protein synthesis and
cytoskeletal organization. In particular, mTOR coordinates cell
viability in response to inputs of growth factors, energy stress
and nutrient status, in addition to regulating cell cycle
progression and survival (8–10). In mammalian cells, mTOR, via its two
distinct multiprotein complexes, mTORC1 and mTORC2, has an
important function in the regulation of different biological
functions. mTORC1 is sensitive to rapamycin and phosphorylates its
downstream effectors, including p70 S6 kinase, which in turn
phosphorylates ribosomal S6 protein, thereby promoting protein
biosynthesis. Activated mTORC1 also phosphorylates eukaryotic
translation initiation factor 4E-binding protein 1 and promotes the
formation of the protein synthesis initiation complex (11). mTORC2, however, regulates the growth,
proliferation and angiogenesis of cancer cells, as a result of its
contribution to the phosphorylation of Akt at the Ser473
residue, which is crucial for the maximal activation of Akt
(12). In addition, mTORC1 is able to
negatively influence Akt activity by inhibiting mTORC2 (13–15).
Furthermore, the mTORC has a well-known function in MM cell
proliferation, tumor development and chemoresistance through the
diverse actions of mTORC1 and mTORC2 and, as such, a disturbance of
the delicate balance of this pathway may lead to MM cell apoptosis,
tumor regression and an extended survival time for patients with MM
(16). Therefore, inhibitors of the
mTOR pathway may be promising future treatment options for MM.
The majority of studies regarding the mTOR pathway
in cancer treatment primarily focus on rapamycin and its
derivatives, which are known to selectively inhibit mTORC1 and to
upregulate the Akt and extracellular signal-regulated kinase
(ERK)/mitogen-activated protein kinase (MAPK) pathways via a
PI3K-dependent feedback loop, leading to enhanced cell survival and
resistance to chemotherapy (17,18).
mTORC2 was previously believed to phosphorylate Akt at
Ser473 and to be the mediator of rapamycin-associated
drug resistance. In contrast, in a previous study, mTORC2
inhibition by rapamycin did not prevent increased phosphorylation
of Akt or drug resistance (19). Due
to the fact that mTORC2 was considered to phosphorylate Akt at
Ser473, it was initially understood to be the mediator
of rapamycin-associated drug resistance. Based upon existing data,
it appears that the efficacy of monotherapy with rapamycin or its
derivatives in the treatment of MM is highly limited (7). This may be due to the fact that
rapamycin and its derivatives do not completely block mTORC1 and
are unable to effectively inhibit mTORC2 to trigger feedback
activation of Akt signaling in order to attenuate antitumor
activity (20,21). In recent years, preclinical data have
demonstrated that mTOR inhibitors, including rapamycin,
temsirolimus and everolimus, may be potential target therapies for
the treatment of MM, particularly if combined with other drugs
(22,23). Therefore, it is reasonable to
hypothesize that combined therapy with rapamycin and novel
naturally-occurring compounds that suppress the rapamycin-induced
Akt/ERK activation may improve the therapeutic efficacy of
rapamycin against a broad range of human tumor cells.
Integrative medicine presents the theory of
utilizing traditional Western medical principles synergistically
with other evidence-based biomedical concepts. Accordingly,
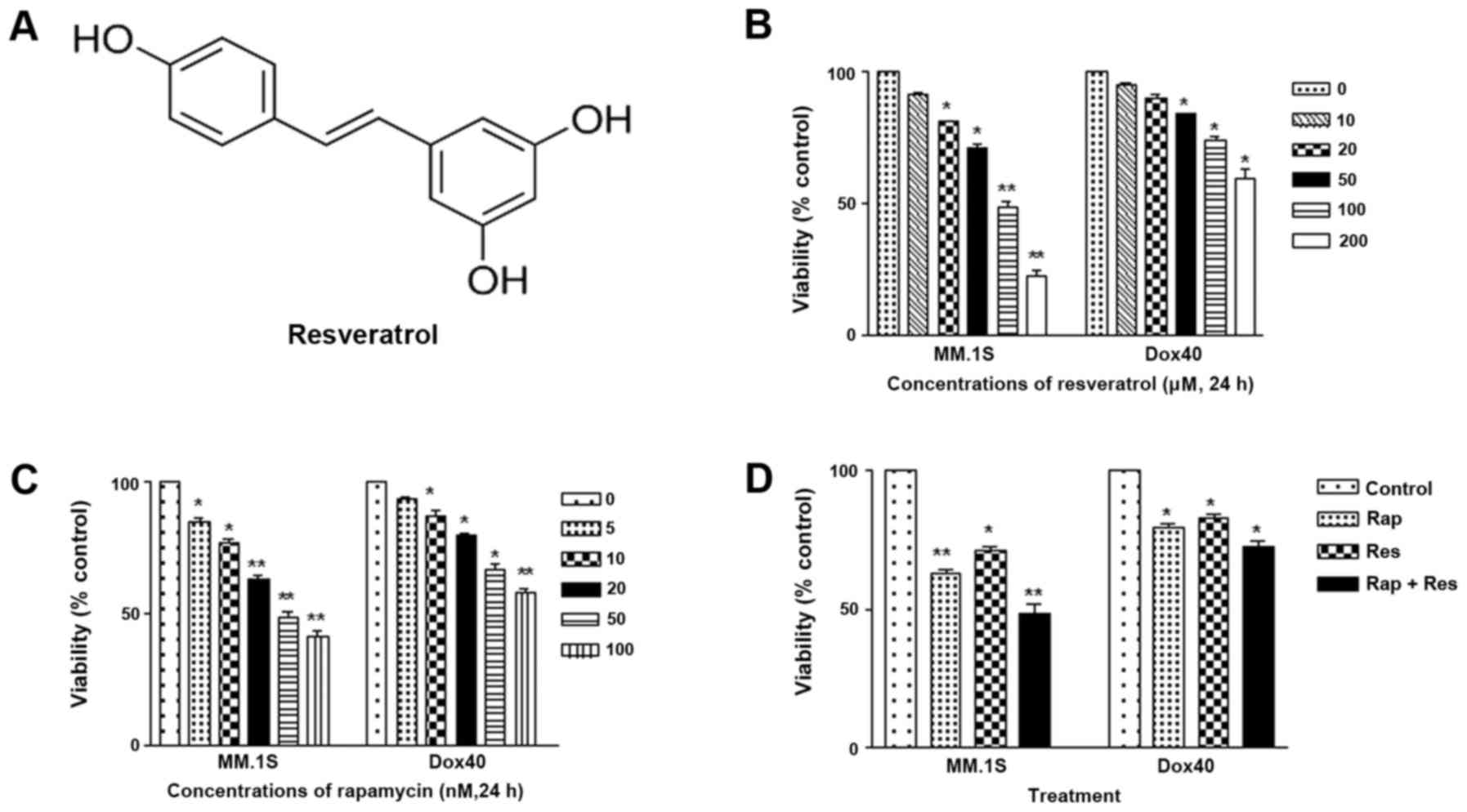
resveratrol (3,4′,5-trihydroxy-trans-stilbene; Fig. 1A), a polyphenolic compound that was
first isolated in 1940 as an ingredient of the roots of white
hellebore (Veratrum grandiflorum O.Loes), was selected for
use in the present study. Since its isolation, resveratrol has been
identified in extracts from >70 other plant species (24,25), and
demonstrates antitumor effects both in vivo and in
vitro through regulation of cell division, growth, angiogenesis
and metastasis (26). Additionally,
resveratrol has been reported to inhibit the proliferation and
induce the apoptosis of MM cells, as well as overcoming the
chemoresistance of these cells (27,28). In
human ovarian cancer cells, resveratrol induces phosphatase and
tensin homolog, in addition to reducing the levels of
phosphorylated-Akt (p-Akt) and mTOR (29,30).
Furthermore, certain studies have suggested that resveratrol may be
useful in cancer therapy when used in combination with rapamycin in
the treatment of breast cancer and chronic myeloid leukemia,
primarily due to its ability to suppress the PI3K/Akt/mTOR
signaling pathway (31,32). However, to the best of our knowledge,
whether or not MM may be treated by combined therapy with
resveratrol and rapamycin has not previously been reported.
The aim of the present study was to investigate
whether combining resveratrol with rapamycin has potential
antitumor effects in a human MM cell line and to determine whether
modulation of the PI3K/Akt/mTOR signaling pathway by resveratrol is
crucial for its anticancer effects in a human MM cell line.
Materials and methods
MM cell lines and cell culture
Dexamethasone-sensitive MM1.S and
doxorubicin-resistant RPMI-8226/DOX40 cell lines were obtained from
the American Type Culture Collection (ATCC; Manassas, VA, USA). The
two MM cell lines were cultured in RPMI-1640 medium (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany), containing 10% fetal bovine serum
(FBS; Sigma-Aldrich; Merck KGaA), 2 mM L-glutamine (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and
100 µg/ml streptomycin (both Gibco; Thermo Fisher Scientific, Inc.)
at 37°C with 5% CO2 in a humid incubator.
Reagents and antibodies
Resveratrol (Fig. 1A),
dimethyl sulfoxide (DMSO), MTT and rapamycin were purchased from
Sigma-Aldrich; Merck KGaA. Annexin V-fluorescein isothiocyanate and
propidium iodide were purchased from BD Biosciences (San Jose, CA,
USA). All primary antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The secondary horseradish
peroxidase-conjugated mouse anti-rabbit IgG polyclonal antibodies
for western blot analysis were provided by Beijing Zhongshan Golden
Bridge Biotechnology Co., Ltd. (Beijing, China).
Cell viability assay
All MM cells were cultured for 24 h at 37°C in
RPMI-1640 medium (Sigma-Aldrich; Merck KGaA) alone or with varying
concentrations of rapamycin (0, 5, 10, 20, 50 and 100 nM),
resveratrol (0, 10, 20, 50, 100 and 200 µM) or a combination of the
two drugs (concentrations of resveratrol and rapamycin were 60 µM
and 20 nM, respectively). In all the experiments, control wells
were included with DMSO at the highest concentration tested with
resveratrol or rapamycin. Cells (1×104) from 24-h
cultures were analyzed using an MTT assay. The medium was
completely removed and 200 µl DMSO was added to dissolve the MTT
formazan crystals. Absorbance readings at a wavelength of 570 nm
(OD570) were taken on a microplate reader (MQX 200; BioTek
Instruments, Inc., Winooski, VT, USA). At least three independent
experiments were performed.
Western blot analysis
For the analysis of mTORC1, mTORC2, caspase-3, poly
ADP ribose polymerase (PARP), cyclin D1 and retinoblastoma protein
(Rb) expression, whole cells were lysed in ice-cold lysis buffer
[50 mM Tris-HCl (pH 7.5), 150 µM NaCl, 20 µM NaF, 20 µM
b-glycerophosphate, 1% Nonidet P-40, 1% Sodium Lauryl Sulfate, 10
mg/ml phenylmethanesulfonyl fluoride, 5 mM EDTA, 10 mg/ml aprotinin
and 10 mg/ml leupeptin]. Equal quantities of protein from total
cell extracts were mixed with a sample buffer containing 100 mM
β-mercaptoethanol and were heated for 10 min at 97°C. Content of
the total protein was detected by BCA protein assay kit (Tiangen
Biotech Co., Ltd., Beijing, China). Protein (20 µg) samples were
separated by 10% SDS-PAGE and were transferred onto Immobilon
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). Polyvinylidene difluoride membranes were incubated in 5%
non-fat dry milk at room temperature for 2 h. Rabbit anti-mTORC1
(cat. no. 2587; 1:1,000), mouse anti-mTORC2 (cat. no. 13017;
1:1,000), rabbit anti-caspase-3 (cat. no. 9664; 1:1,000) and rabbit
anti-PARP (cat. no. 9532; 1:1,000) antibodies (Cell Signaling
Technology, Inc.) were added and incubated overnight at 4°C.
Peroxidase-conjugated Affinipure goat anti-rabbit IgG (cat. no.
ZB-2301, dilution 1:5,000) and rabbit anti-mouse IgG (cat. no.
ZDR-5109, dilution 1:5,000) (Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.) were then added separately and incubated
for 2 h at room temperature. Following washing, membranes were
incubated with horseradish peroxidase-conjugated secondary
antibodies at room temperature for 1 h. Blots were then developed
using an enhanced chemiluminescence kit (EMD Millipore).
Densitometric analysis of protein bands was performed using Image J
v.1.45s software (National Institutes of Health, Bethesda, MD,
USA). All western blot analysis was performed ≥3 times to ensure
reproducibility.
Statistical analysis
Data are expressed as the mean ± standard deviation.
SPSS version 17.0 software (SPSS, Inc., Chicago, IL, USA) was used
for statistical analysis. Comparisons among multiple groups were
determined using one-way analysis of variance, followed by Fisher's
least significant difference test (for equal variances assumed) or
Dunnett's T3 test (for equal variances not assumed) to compare
individual group differences. P<0.05 was considered to indicate
a statistically significant difference.
Results
Resveratrol, rapamycin and combined
treatment suppress the viability of MM cells
In the present study, an MTT assay was used to
determine the effect on cell viability of resveratrol and
rapamycin. The MM MM1.S and RPMI-8226/DOX40 cell lines were
cultured in complete medium in the absence (control) or presence of
various concentrations of resveratrol and rapamycin for indicated
time periods. Untreated cells were considered as the baseline
(100%) for analysis. The viability-impairing activity of
resveratrol, rapamycin and the combination of the two is presented
in Fig. 1B-D. Cells were initially
treated with varying concentrations of resveratrol (0, 10, 20, 50,
100 and 200 µM) and rapamycin (0, 5, 10, 20, 50 and 100 nM) for 24
h. Resveratrol and rapamycin significantly impaired the viability
of the MM1.S cells in a dose-dependent manner, which was more
significant than the effect on RPMI-8226/DOX40 cells. For the
combined treatment, 60 µM resveratrol was used with 20 nM
rapamycin. In the MM.1S cells, the reduction in the rate of cell
viability proliferation was significantly increased in the
combination treatment group compared with the control group
(P<0.01), and in RPMI-8226/DOX40 cells there was a significant
reduction in the rate of cell viability proliferation in the
combination treatment group compared with the blank group
(P<0.05). Therefore, the MM.1S cells were more sensitive to
combination therapy compared with the RPMI-8226/DOX40 cells.
Compared with resveratrol or rapamycin monotherapy, the combination
of the two drugs was much more effective in impairing the viability
of the MM1.S cells, an effect that was more notable than that on
the RPMI-8226/DOX40 cells. These results indicated that combined
drug therapy may improve the treatment of MM, as single-agent
therapy is becoming less effective as a result of the multi-drug
resistance exhibited by cancer cells.
mTOR/Akt signaling in MM cells
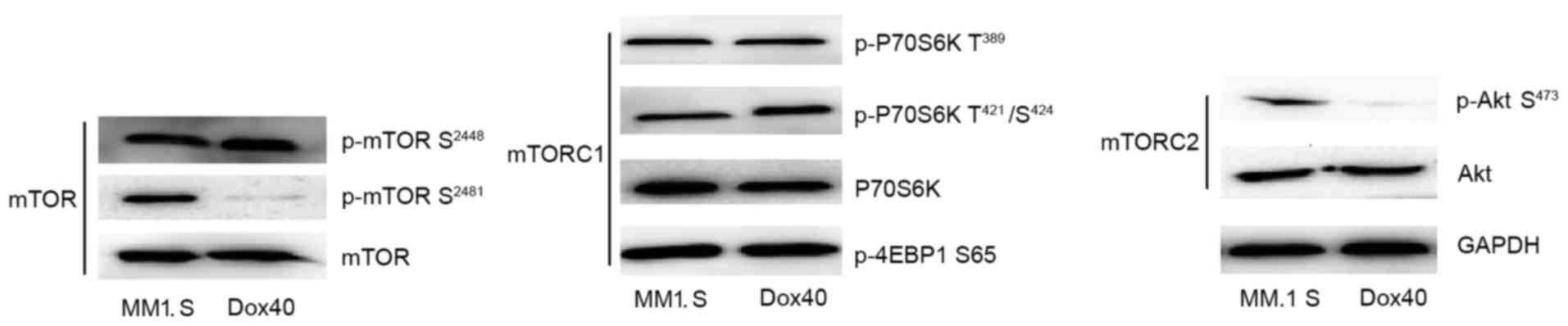
The present study initially examined whether or not
autophosphorylation of mTOR at Ser2481 may be used as
readout for mTORC2/Akt signaling in MM cells. Therefore, the
baseline phosphorylation of mTOR on Ser2481 in MM cell
lines was evaluated. Normally, mTORC1 primarily contains mTOR
phosphorylated at Ser2448, while mTORC2 predominantly
contains mTOR phosphorylated at Ser2481 (33). Western blot analysis revealed that
p-mTOR Ser2481 was present in MM1.S cells, while p-mTOR
Ser2448 was expressed in the two MM cell lines (Fig. 2). Subsequently, the expression of mTOR
phosphorylated at Ser2448 and Ser2481 was
compared with TORC1 and TORC2 readouts, respectively. The
best-characterized mTORC1 readouts are P70S6K and 4EBP, whereas the
best-characterized mTORC2 readouts are Akt. The results of the
present study demonstrated that baseline expression of p-mTOR
(Ser2448) is associated with TORC1-phosphorylatable
P70S6K and 4EBP, and that expression of p-mTOR Ser2481
is associated with TORC2-phosphorylatable Akt Ser473 in
the MM1.S cell line. p-mTOR Ser2448 is highly expressed
in the RPMI-8226/DOX40 cell line, while p-Akt Ser473 is
not. These results indicated that aberrant activation of mTORC1 and
mTORC2 signaling exists in the MM1.S cell line used in the present
study.
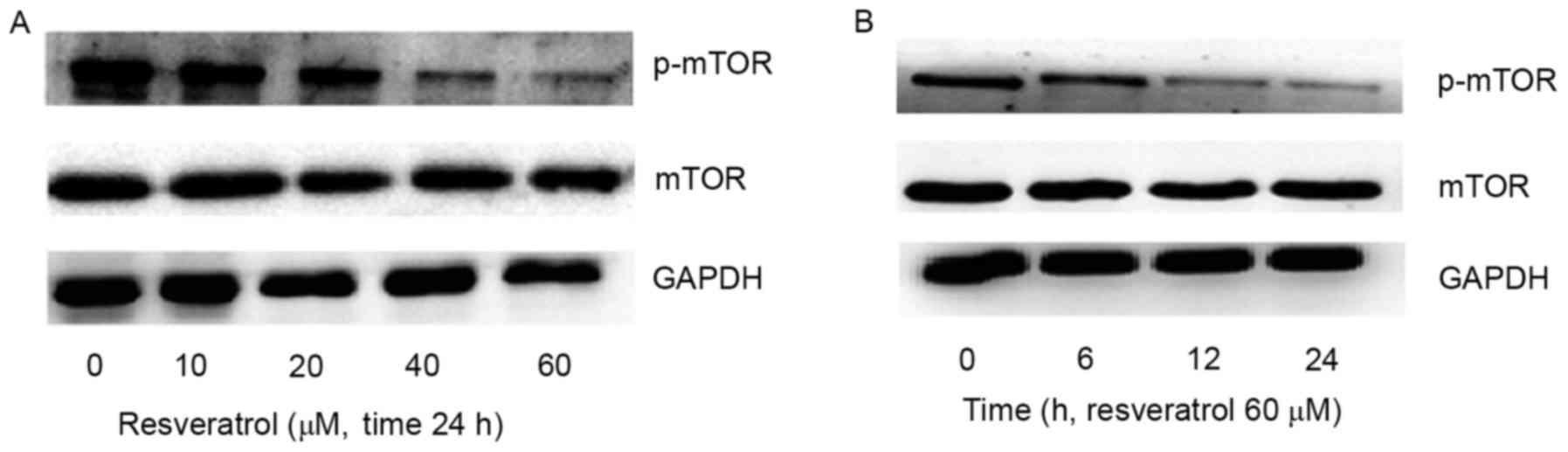
Resveratrol downregulates mTOR protein
expression in MM1.S cells
Analysis of the expression of the proteins involved
in the PI3K/Akt/mTOR signaling pathway revealed that resveratrol
reduced the expression of p-mTOR in a dose-dependent manner
(Fig. 3A). Similarly, the MM1.S cells
were subsequently treated with 60 µM resveratrol for 0, 6, 12 and
24 h and p-mTOR expression decreased following resveratrol
treatment for 12 h (Fig. 3B). These
results indicated that the specific inhibitor of mTOR enhances the
effect induced by resveratrol.
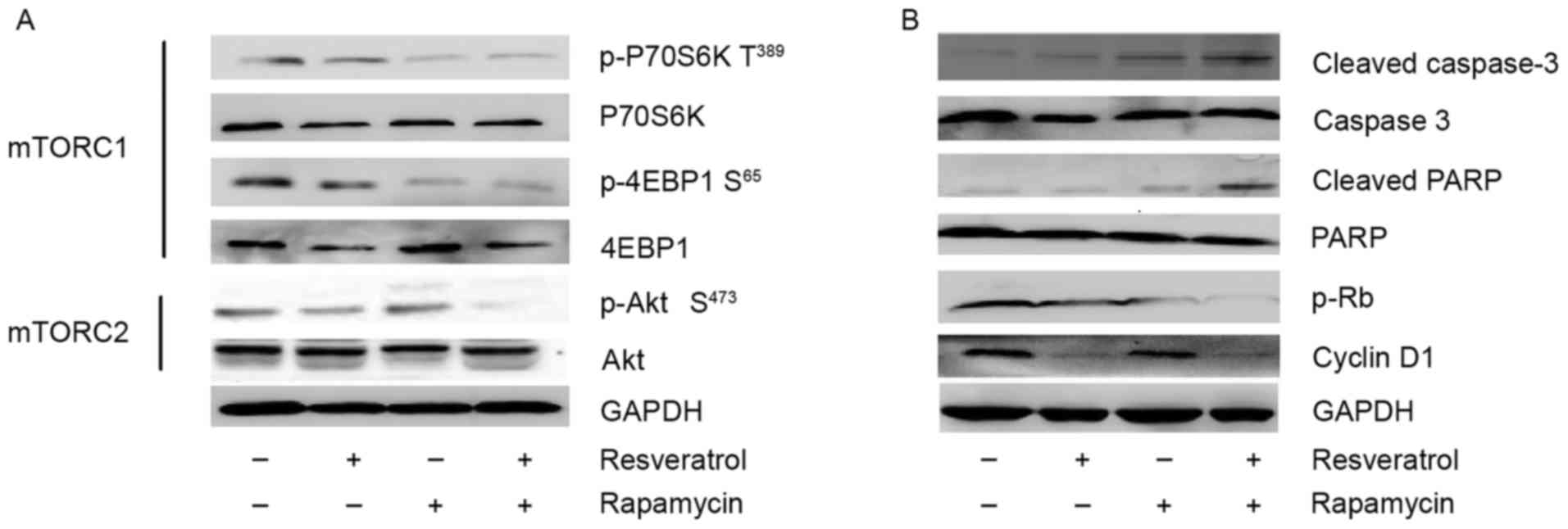
Combined treatment with resveratrol
and rapamycin is synergistic in MM1.S cell line
Following treatment with resveratrol, rapamycin, a
combination of the two or DMSO for 24 h, the MM1.S cells were
collected by trypsinization and were used in western blot analysis
or stored at −80°C until further use. Phosphorylation of Akt was
measured at Ser473 (mTORC2 substrate), phosphorylation
of P70S6K was measured at Thr389 (mTORC1 substrate) and
phosphorylation of 4EBP1 was measured at Ser65 (mTORC1
substrate). As demonstrated in Fig.
4A, combined treatment effectively inhibited p-Akt
Ser473, p-P70S6K Thr389 and p-4EBP1
Ser65 in MM1.S cells, suggesting that combined treatment
inhibits mTORC1 and mTORC2 signaling in the MM1.S cell line.
However, rapamycin markedly inhibited p-P70S6K Thr389
and p-4EBP1 Ser65, but did not result in the inhibition
of p-Akt Ser473, suggesting that rapamycin only inhibits
mTORC1 signaling and is unable to suppress mTORC2 signaling in
MM1.S cells. In addition, rapamycin treatment resulted in a
significant increase in p-Akt Ser473, demonstrating that
rapamycin treatment induced feedback activation of Akt signaling in
MM1.S cells. Notably, resveratrol treatment effectively inhibited
p-Akt Ser473, p-P70S6K Thr389 and p-4EBP1
Ser65 in MM1.S cells, indicating that resveratrol may
inhibit mTORC2 and mTORC1 signaling in MM1.S cell lines.
Resveratrol, rapamycin and a
combination of the two induce activation of caspase-3 and PARP in
MM1.S cells
The aforementioned results demonstrated that
combining resveratrol with rapamycin may inhibit mTOR signaling in
MM1.S cells. In order to analyze whether treatments with rapamycin,
resveratrol or a combination of the two had an effect on the
apoptosis of MM1.S cells, cleavage of PARP and caspase-3, the key
mediators of the apoptotic process, was detected by western blot
analysis. As demonstrated in Fig. 4B,
rapamycin induced gentle cleavage of PARP and caspase-3, consistent
with its cytostatic effect. Resveratrol induced minimal cleavage of
PARP and caspase-3, whereas the combination of rapamycin and
resveratrol induced a higher degree of PARP and caspase-3 cleavage
compared with the effect observed with each agent alone.
Furthermore, the results demonstrated that PARP had undergone
cleavage while the expression of caspase-3 diminished. The
expression of these proteins following combined treatment decreased
markedly. The results of the present study indicated that
resveratrol-, rapamycin- and combined therapy-induced apoptosis in
MM1.S cells is associated with caspase-3 and PARP.
Resveratrol combined with rapamycin
suppresses cyclin D1 expression and Rb phosphorylation in MM1.S
cells
In order to further investigate the synergistic
mechanism of combining resveratrol with rapamycin, the present
study examined the effect of these agents on cyclin D1/Rb
expression. As demonstrated in Fig.
4B, cyclin D1 and phosphorylated Rb (p-Rb) were detected in
MM1.S cells treated with rapamycin, resveratrol or a combination of
the two, using immunoblotting. In MM1.S cells, monotherapy with
resveratrol inhibited cyclin D1 protein expression, while rapamycin
barely inhibited cyclin D1 expression. Cyclins interact with
cyclin-dependent kinases (CDKs) to phosphorylate Rb. Since
D-cyclins are major determinants of CDK4/6 holoenzyme activity,
decreased expression of cyclin D1 may prevent Rb phosphorylation in
an Akt-dependent fashion. In order to confirm this hypothesis, the
Rb phosphorylation status in MM1.S cells treated with rapamycin,
resveratrol or a combination of the two was also detected. The
results indicated that the combination of rapamycin and resveratrol
induced downregulation of phosphorylated Rb.
Discussion
Combinations of novel agents that target various
signaling pathways in MM are required in order to overcome
resistance to treatment and to achieve higher response and survival
rates in patients with MM. A previous study demonstrated that the
combination of rapamycin with other agents, including 17-AAG,
CC-5013 and dexamethasone, leads to a synergistic effect on MM
cells and sensitizes the cells to apoptosis (34). Resveratrol is a naturally occurring
phytoalexin and a polyphenolic compound. The molecule was revealed
to be present in various fruits and vegetables and is abundant in
grapes and red wine (24,25). Previous studies have demonstrated that
resveratrol not only has no known cytotoxicity in animal models,
but also is relatively pharmacologically safe in humans (35,36).
Although resveratrol has been reported to have a wide range of
potential targets during the inhibition of proliferation and the
induction of apoptosis in a variety of tumor cell types (37), the underlying molecular mechanisms of
its anticancer effects are not well understood, particularly in MM
which is often difficult to treat as multiple signaling pathways
are involved (38). Contradictory
results have previously been reported regarding the inhibition of
proliferation and the induction of apoptosis resulting from
treatment with resveratrol. A previous study reported that
resveratrol treatment induces apoptosis in various tumor cells
(39). However, an additional study
reported that resveratrol induces differentiation, but not
apoptosis, in certain types of cancer cells (40). In the present study, it was revealed
that the combination of rapamycin and resveratrol, tested at
pharmacologically achievable doses, resulted in a strong
synergistic effect on MM cell viability in vitro. This
combination led to the viability impairment of MM cells and the
induction of apoptosis in the two MM cell lines. However, the
effects on MM1.S cells treated with a combination of rapamycin and
resveratrol were more significant than the effects on the
RPMI-DOX40 cells and therefore, the focus was primarily on the
effect of combined treatment on MM1.S cells.
There is a large volume of evidence supporting the
critical function of the PI3K/Akt/mTOR signaling pathway in cancer
proliferation, tumor genesis and metastasis, and numerous studies
have demonstrated that PI3K/Akt/mTOR activity is increased in a
variety of tumor cell lines, including MM cells (41–43).
Dysregulation of mTOR function via physiological or mutational
activation of upstream pathways is a common event in tumors. mTOR
activation has been observed in numerous types of cancer and
therefore, mTOR has become an attractive target for cancer therapy
(7). As a catalytic subunit, mTOR
exists in two complexes, mTORC1 (Ser2448) and mTORC2
(Ser2481), which function as specific scaffold proteins.
Therefore, decreasing the expression of these two proteins may lead
to reduced mTOR activity as mTOR activity is dependent upon the
integrity of the complex. Impairment of any single component of the
mTOR complex may reduce mTOR activity. The results of the present
study demonstrated that p-mTOR Ser2481 was present in
MM1.S cells, while p-mTOR Ser2448 was revealed to be
expressed in the two MM cell lines by western blot analysis
(Fig. 2), indicating that the mTORC1
and mTORC2 signaling pathways were activated in the MM1.S cell
lines. Rapamycin inhibits basal p70S6K activity and induces
dephosphorylation of P70S6K and 4EBP-1, thereby inhibiting cap
structure-dependent protein synthesis and cell proliferation
(44), indicating that rapamycin may
directly inhibit mTORC1 and lead to tumor cell apoptosis.
Furthermore, the results of a previous study demonstrated that
mTORC2 is rapamycin-insensitive, although it may be inhibited by
prolonged rapamycin treatment (45).
Activated mTORC1 phosphorylates P70S6K and 4EBP1, two
best-characterized substrates of mTORC1, to positively regulate the
protein synthesis that drives cell growth and proliferation.
Therefore, p-P70S6K and p-4EBP1 are frequently used in research as
biomarkers of mTORC1 activity. mTORC2 has been considered to
function primarily through activating Akt by phosphorylating it at
Ser473, which is important for its maximal activation
(33). Therefore, the level of p-Akt
Ser473 is widely used to measure mTORC2 activity. The
results of the present study demonstrated that the mTOR inhibitor,
rapamycin, may inhibit mTORC1 signaling as p-P70S6K
Thr389 and p-4EBP1 Ser65 levels were reduced,
and may be unable to effectively inhibit mTORC2 signaling and
result in feedback activation of Akt signaling as p-Akt
Ser473 levels increased, thereby attenuating antitumor
activity in MM1.S cells (Fig. 4A).
The results of the present study were consistent with those of a
previous study (19). By contrast,
resveratrol treatment significantly decreased the phosphorylation
of Akt at Ser473 in MM1.S cells, indicating that
resveratrol is effective in attenuating mTORC2 function and
preventing the feedback activation of Akt. These results indicated
that resveratrol may inhibit mTORC2 signaling and may not suppress
mTORC1 signaling in MM1.S cells. Therefore, combined treatment with
rapamycin and resveratrol effectively inhibited p-Akt
Ser473, p-P70S6K Thr389 and p-4EBP1
Ser65 in MM1.S cells, suggesting that combined treatment
inhibited mTORC1 and mTORC2 signaling in the MM1.S cells.
The results of the present study demonstrated that
the combination of rapamycin and resveratrol induced a significant
effect on the cleavage of apoptosis-associated proteins, including
PARP and caspase-3. The downstream signaling cascades targeted by
rapamycin and resveratrol were subsequently delineated. The results
of the present study demonstrated that rapamycin induced gentle
cleavage of PARP and caspase-3, and that resveratrol induced
minimal cleavage of PARP and caspase-3, whereas the combination of
rapamycin and resveratrol induced a higher degree of PARP and
caspase-3 cleavage compared with the effect observed following
monotherapy with each agent alone. Furthermore, the results also
demonstrated that PARP had undergone cleavage as the expression of
caspase-3 decreased. Additionally, the expression of these proteins
following combined treatment decreased markedly. The results of the
present study indicated that resveratrol-, rapamycin- and combined
therapy-induced apoptosis of MM1.S cells may be associated with
caspase-3 and PARP.
In the present study, cyclin D1 and phosphorylated
Rb were revealed to be inhibited in MM1.S cells treated with
rapamycin and resveratrol, indicating another possible mechanism of
synergy. A previous study demonstrated that low Akt activity in MM
cell lines induced resistance to rapamycin by allowing continued
cap-independent protein synthesis of cyclin D1 (46). In the present study, it was
demonstrated that cyclin D1 was downregulated in response to
resveratrol in the MM1.S cell line, indicating that cyclin D1 may
be a useful marker of response to resveratrol in MM. Notably,
phosphorylated Rb, another G1 regulatory protein downstream of
cyclin D1, was inhibited in response to rapamycin, resveratrol and
a combination of the two agents in MM1.S cells. Further studies are
required to investigate the role of other cyclins and CDKs in the
inhibition of phosphorylated Rb in cell lines with lower Akt
activity. These results suggested mechanisms for synergy of
rapamycin and resveratrol through targeting of multiple proteins of
the PI3K/Akt/mTOR pathway and G1 regulatory proteins.
In conclusion, the results of the present study
suggested that resveratrol is well-tolerated and exhibits low
toxicity and is therefore safe for preclinical research. Notably,
it was revealed that the newly-developed compound, resveratrol, may
inhibit mTORC2 signaling. Additionally, the combination of
rapamycin with resveratrol is safe and effective in inhibiting the
proliferation and increasing the apoptosis of MM1.S cells. Taken
together, the results of the present study indicated that combined
therapy with rapamycin and resveratrol may be an attractive
approach for the treatment and prevention of MM. However, these
observations require further investigation in preclinical
trials.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YGB made contributions to experimental design,
revised the manuscript critically for important intellectual
content and provided final approval of the version to be published;
HGJ made substantial contributions to the acquisition of data, and
analysis and interpretation of data, and was involved in drafting
the manuscript. GZW was involved in performing the experiments. GHW
conducted the MTT assays and analyzed cell viability. GHW was also
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of any part of the work are
appropriately resolved. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This experiment only involves cell level research,
approved by Ethics Committee of Affiliated Hospital of Inner
Mongolia University for the Nationalities (Ethics Committee no.
0010/2016).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Palumbo A and Anderson K: Multiple
myeloma. N Engl J Med. 364:1046–1060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kyle RA, Gertz MA, Witzig TE, Lust JA,
Lacy MQ, Dispenzieri A, Fonseca R, Rajkumar SV, Offord JR, Larson
DR, et al: Review of 1027 patients with newly diagnosed multiple
myeloma. Mayo Clin Proc. 78:21–33. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cao B, Li J and Mao X: Dissecting
bortezomib: Development, application, adverse effects and future
direction. Curr Pharm Des. 19:3190–3200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Herndon TM, Deisseroth A, Kaminskas E,
Kane RC, Koti KM, Rothmann MD, Habtemariam B, Bullock J, Bray JD,
Hawes J, et al: U.S. Food and drug administration approval:
Carfilzomib for the treatment of multiple myeloma. Clin Cancer Res.
19:4559–4563. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang F, Zhang W, Guo L, Bao W, Jin N, Liu
R, Liu P, Wang Y, Guo Q and Chen B: Gambogic acid suppresses
hypoxia-induced hypoxia-inducible factor-1α/vascular endothelial
growth factor expression via inhibiting phosphatidylinositol
3-kinase/Akt/mammalian target protein of rapamycin pathway in
multiple myeloma cells. Cancer Sci. 105:1063–1070. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Adachi M, Hoshino Y, Izumi Y, Sakai H and
Takagi S: Effects of inhibitors of vascular endothelial growth
factor receptor 2 and downstream pathways of receptor tyrosine
kinases involving phosphatidylinositol 3-kinase/Akt/mammalian
target of rapamycin or mitogen-activated protein kinase in canine
hemangiosarcoma cell lines. Can J Vet Res. 80:209–216.
2016.PubMed/NCBI
|
|
7
|
Cirstea D, Santo L, Hideshima T, Eda H,
Mishima Y, Nemani N, Mahindra A, Yee A, Gorgun G, Hu Y, et al:
Delineating the mTOR Kinase pathway using a dual TORC1/2 inhibitor,
AZD8055, in multiple myeloma. Mol Cancer Ther. 13:2489–2500. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gera J and Lichtenstein A: The mammalian
target of rapamycin pathway as a therapeutic target in multiple
myeloma. Leuk Lymphoma. 52:1857–1866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dowling RJ, Topisirovic I, Fonseca BD and
Sonemberg N: Dissecting the role of mTOR: Lessons from mTOR
inhibitor. Biochim Biophys Acta. 1804:433–439. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2010. View Article : Google Scholar
|
|
11
|
Hara K, Yonezawa K, Weng QP, Kozlowski MT,
Belham C and Avruch J: Amino acid sufficiency and mTOR regulate p70
S6 kinase and eIF-4E BP1 through a common effector mechanism. J
Biol Chem. 273:14484–14494. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Serrano M: Dissecting the role of mTOR
complexes in cellular senescence. Cell Cycle. 11:2231–2232. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lang SA, Hackl C, Moser C, Fichtner-Feigl
S, Koehl GE, Schlitt HJ, Geissler EK and Stoeltzing O: Implication
of RICTOR in the mTOR inhibitor-mediated induction of insulin-like
growth factor-I receptor (IGF-IR) and human epidermal growth factor
receptor-2 (Her2) expression in gastrointestinal cancer cells.
Biochim Biophys Acta. 1803:435–442. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wander SA, Hennessy BT and Slingerland JM:
Next-generation mTOR inhibitors in clinical oncology: How pathway
complexity informs therapeutic strategy. J Clin Invest.
121:1231–1241. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Zhu J, Cao B and Mao X: The mTOR
signaling pathway is an emerging therapeutic target in multiple
myeloma. Curr Pharm Des. 20:125–135. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Willems L, Tamburini J, Chapuis N, Lacombe
C, Mayeux P and Bouscary D: PI3K and mTOR signaling pathways in
cancer: New data on targeted therapies. Curr Oncol Rep. 14:129–138.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Benjamin D, Colombi M, Moroni C and Hall
MN: Rapamycin passes the torch: A new generation of mTOR
inhibitors. Nat Rev Drug Discov. 10:868–880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feldman ME, Apsel B, Uotila A, Loewith R,
Knight ZA, Ruggero D and Shokat KM: Active-site inhibitors of mTOR
target rapamycin-resistant outputs of mTORC1 and mTORC2. PLoS Biol.
7:e382009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu DM, Zhao L, Liu TT, Jiao PL, Zhao DD,
Shih MS, Tao B, Sun LH, Zhao HY and Liu JM: Rictor/mTORC2 loss in
osteoblasts impairs bone mass and strength. Bone. 90:50–58. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsieh AC, Costa M, Zollo O, Davis C,
Feldman ME, Testa JR, Meyuhas O, Shokat KM and Ruggero D: Genetic
dissection of the oncogenic mTOR pathway reveals druggable
addiction to translational control via 4EBP-eIF4E. Cancer Cell.
17:249–261. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yan H, Frost P, Shi Y, Hoang B, Sharma S,
Fisher M, Gera J and Lichtenstein A: Mechanism by which mammalian
target of rapamycin inhibitors sensitize multiple myeloma cells to
dexamethasone-induced apoptosis. Cancer Res. 66:2305–2313. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Francis LK, Alsayed Y, Leleu X, Jia X,
Singha UK, Anderson J, Timm M, Ngo H, Lu G, Huston A, et al:
Combination mammalian target of rapamycin inhibitor rapamycin and
HSP90 inhibitor 17-allylamino-17-demetoxiygeldanamycin has
synergistic activity in multiple myeloma. Clin Cancer Res.
12:6826–6835. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gupta SC, Kannappan R, Reuter S, Kim JH
and Aggarwal BB: Chemosensitization of tumors by resveratrol. Ann N
Y Acad Sci. 1215:150–160. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Baur JA and Sinclair DA: Therapeutic
potential of resveratrol: The in vivo evidence. Nat Rev Drug
Discov. 5:493–506. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bishayee A: Cancer prevention and
treatment with resveratrol: From rodent studies to clinical trials.
Cancer Prev Res (Phila). 2:409–418. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bhardwaj A, Sethi G, Vadhan-Raj S,
Bueso-Ramos C, Takada Y, Gaur U, Nair AS, Shishodia S and Aggarwal
BB: Resveratrol inhibits proliferation, induces apoptosis, and
overcomes chemoresistance through down-regulation of STAT3 and
nuclear factor-kappaB-regulated antiapoptotic and cell survival
gene products in human multiple myeloma cells. Blood.
109:2293–2302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Boissy P, Andersen TL, Abdallah BM, Kassem
M, Plesner T and Delaissé JM: Resveratrol inhibits myeloma cell
growth, prevents osteoclast formation, and promotes osteoblast
differentiation. Cancer Res. 65:9943–9952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kueck A, Opipari AW Jr, Griffith K, Tan L,
Choi M, Huang J, Wahl H and Liu JR: Resveratrol inhibits glucose
metabolism in human ovarian cancer cells. Gynecol Oncol.
107:450–457. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Waite KA, Sinden MR and Eng C:
Phytoestrogen exposure elevates PTEN levels. Hum Mol Genet.
14:1457–1463. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He X, Wang Y, Zhu J, Orloff M and Eng C:
Resveratrol enhances the anti-tumor activity of the mTOR inhibitor
rapamycin in multiple breast cancer cell lines mainly by
suppressing rapamycin-induced AKT signaling. Cancer Lett.
301:168–176. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sui T, Ma L, Bai X, Li Q and Xu X:
Resveratrol inhibits the phosphatidylinositide 3-kinase/protein
kinase B/mammalian target of rapamycin signaling pathway in the
human chronic myeloid leukemia K562 cell line. Oncol Lett.
7:2093–2098. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Copp J, Manning G and Hunter T:
TORC-specific phosphorylation of mammalian target of rapamycin
(mTOR): Phospho-Ser2481 is a marker for intact mTOR signaling
complex 2. Cancer Res. 69:1821–1827. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Raje N, Kumar S, Hideshima T, Ishitsuka K,
Chauhan D, Mitsiades C, Podar K, Le Gouill S, Richardson P, Munshi
NC, et al: Combination of the mTOR inhibitor rapamycin and CC-5013
has synergistic activity in multiple myeloma. Blood. 104:4188–4193.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cao Y, Fu ZD, Wang F, Liu HY and Han R:
Anti-angiogenic activity of resveratrol, a natural compound from
medicinal plants. J Asian Nat Prod Res. 7:205–213. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gusman J, Malonne H and Atassi G: A
reappraisal of the potential chemopreventive and chemotherapeutic
properties of resveratrol. Carcinogenesis. 22:1111–1117. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang B, Liu J and Gong Z: Resveratrol
induces apoptosis in K562 cells via the regulation of mitochondrial
signaling pathways. Int J Clin Exp Med. 8:16926–16933.
2015.PubMed/NCBI
|
|
38
|
Jang J, Jeong SJ, Kwon HY, Jung JH, Sohn
EJ, Lee HJ, Kim JH and Kim SH, Kim JH and Kim SH: Decursin and
doxorubicin are in synergy for the induction of apoptosis via STAT3
and/or mTOR pathways in human multiple myeloma cells. Evid Based
Complement Alternat Med. 2013:5063242013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Trung Quoc L, Espinoza JL, Takami A and
Nakao S: Resveratrol induces cell cycle arrest and apoptosis in
malignant NK cells via JAK2/STAT3 pathway inhibition. PLoS One.
8:e551832013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen Q, Ganapathy S, Singh KP, Shankar S
and Srivastava RK: Resveratrol induces growth arrest and apoptosis
through activation of FOXO transcription factors in prostate cancer
cells. PLoS One. 5:e152882010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li H, Hu J, Wu S, Wang L, Cao X, Zhang X,
Dai B, Cao M, Shao R, Zhang R, et al: Auranofin-mediated inhibition
of PI3K/AKT/mTOR axis and anticancer activity in non-small cell
lung cancer cells. Oncotarget. 7:3548–3558. 2016.PubMed/NCBI
|
|
42
|
Que W, Chen J, Chuang M and Jiang D:
Knockdown of c-Met enhances sensitivity to bortezomib in human
multiple myeloma U266 cells via inhibiting Akt/mTOR activity.
APMIS. 120:195–203. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Feng N, Luo J and Guo X: Silybin
suppresses cell proliferation and induces apoptosis of multiple
myeloma cells via the PI3K/Akt/mTOR signaling pathway. Mol Med Rep.
13:3243–3248. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rousseau D, Gingras AC, Pause A and
Sonenberg N: The eIF4E-binding proteins 1 and 2 are negative
regulators of cell growth. Oncogene. 13:2415–2420. 1996.PubMed/NCBI
|
|
45
|
Sarbassov DD, Ali SM, Kim DH, Guertin DA,
Latek RR, Erdjument-Bromage H, Tempst P and Sabatini DM: Rictor, a
novel binding partner of mTOR, defines a rapamycin-insensitive and
raptor-independent pathway that regulates the cytoskeleton. Curr
Biol. 14:1296–1302. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gera JF, Mellinghoff IK, Shi Y, Rettig MB,
Tran C, Hsu JH, Sawyers CL and Lichtenstein AK: AKT activity
determines sensitivity tomammalian target of rapamycin (mTOR)
inhibitors by regulating cyclin D1 and c-myc expression. J Biol
Chem. 279:2737–2746. 2004. View Article : Google Scholar : PubMed/NCBI
|