Introduction
Ovarian cancer results in abnormal cells that have
the ability to invade or spread to other part of the body.
Recently, numerous studies confirmed that several genes are closely
associated with human ovarian cancer (1,2). However,
the precise molecular mechanisms of ovarian cancer remain to be
clarified. To date, studies on mechanisms of OC have focused on
known genes (3–7), but microRNAs (miRNAs) might also provide
further insights into OC.
The discovery of non-coding RNA in the human genome
was an important conceptual breakthrough in the post-genomic
sequencing era (8–12). miRNAs constitute a class of small,
non-coding RNA molecules with a length of 19-22, that modulates
gene expression. As a new layer of gene-regulation mechanism,
regulation by miRNAs is achieved through imperfect pairing with
target mRNAs of protein-coding genes at the transcriptional or
post-transcriptional level (13).
Furthermore, miRNAs can regulate proliferation, differentiation and
apoptosis as well as cancer initiation and progression (14). A number of studies indicated that
miRNAs were differentially regulated in various tumors, including
colorectal (15), breast (16,17) and
lung cancer (18,19).
Recently, a number of reports show that the majority
of miRNAs are aberrantly expressed in multiple types of cancer,
including OC. For example, miR-17-5p was overexpressed in OC cells,
and the upregulation of miR-17-5p significantly promoted the
proliferation, migration, invasion and epithelial to mesenchymal
transition (EMT) of OC cells by targeting phosphatase and tensin
homolog signaling (20). Niu et
al (21) demonstrated that
miR-205 expression was associated with the motility of OC cells by
targeting zinc finger E-Box binding homeobox 1. miR-661 promoted
the proliferation of OC cells by directly targeting inositol
polyphosphate-5-phosphatase J (22).
These three miRNAs (miR-17-5p, miR-205 and miR-661) act as
oncogenes; some miRNAs with tumor suppressor functions were also
studied in OC. For example, the overexpression of miR-30d reversed
transforming growth factor-β1-induced EMT of OC cells by decreasing
snail expression (23). The level of
miR-127 was markedly downregulated in OC tissues, and the
overexpression of miR-127 inhibited the proliferation of OC cells
by targeting cyclin G1 (24).
miR-302a was also demonstrated to be involved in the suppression of
syndecan 1 to inhibit the tumorigenicity of OC cells (25).
miR-199a-5p has been indicated to be involved in the
regulation of multiple biological processes, including cell
proliferation, apoptosis, migration and invasion (26–31). In
recent years, miR-199a-5p was considered as a tumor suppressor and
downregulated in prostate, gastric and non-small cell lung cancer
(31–33). Guo et al (27) reported that miR-199a-5p functioned as
a tumor suppressor by inhibiting proliferation and inducing
apoptosis in liver cancer cells by targeting hexokinase 2.
In the present study, the downregulation of
miR-199a-5p was observed in HO-8910 and ES-2 OC cell lines. The
upregulation of miR-199a-5p was able to inhibit the proliferation
and invasion of OC cells. Furthermore, NF-κB1 was identified as a
potential target of miR-199a-5p in OC, and miR-199a-5p may function
as a tumor suppressor by downregulating NF-κB1. Therefore, these
results reveal critical roles for miR-199a-5p in the pathogenesis
of OC and miR-199a-5p may be a potential target for the treatment
of ovarian tumors.
Materials and methods
Cell culture and miRNA
transfection
Human OC cell lines, HO-8910 and ES-2, and an
immortalized normal human fallopian tube epithelial cell line,
FTE187, were purchased from the American Type Culture Collection
(ATCC, Manassas, VA, USA). All cell lines were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) 100 U/ml
penicillin and 100 µg/ml streptomycin (Lonza Group, Ltd., Basel,
Switzerland). The cells were cultured in 0.1% gelatin-coated
culture flasks at 37°C with a humidified atmosphere of 5%
CO2. To increase the expression of miR-199a-5p in cell
lines, 50 nM miR-199a-5p mimic (sequence:
5′-cccaguguucagacuaccuguuc-3′; RiboBio Co., Ltd., Guangzhou, China)
was transfected into HO-8910 and ES-2 cells. miR-negative control
(sequence: 5′-catttacgcaguguucagacuaccuguuc-3′; RiboBio Co., Ltd.)
was also transfected into HO-8910 and ES-2 cells, respectively. A
total of 24 h prior to transfection, when the cells were at ~40-60%
confluence, the cell medium was changed to antibiotic-free medium.
After 24 h, the cells were transfected with 50 nM miR-199a-5p mimic
and miR-negative control using Lipofectamine™ 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The expression of miR-199a-5p and four genes
(E-cadherin, vimentin, MMP-2 and MMP-9) was analyzed with SYBR
Green II (Takara Co., Ltd. Dalian China) and a RT-qPCR system (MJ
Research; Bio-Rad Laboratories, Inc., Hercules, CA, USA). Briefly,
the total RNA from HO-8910 and ES-2 cells was extracted using
Trizol reagent (Thermo Fisher Scientific, Inc.). A total of 2 mg
RNA was used for gene-specific RT-qPCR using a one-step RT-PCR kit
(Qiagen, Inc., Valencia, CA, USA) according to the manufacturer's
instructions. Complementary DNA (cDNA) was subjected to 40 cycles
of amplification. The sequences of the primers are as follows:
miR-199a-5p forward, 5′-CCGGGATCCGCAAACTCAGCTTTAC-3′ and reverse
5′-CGGAATTCGTGGCGACCGTGATACC-3′; E-cadherin forward,
5′-TACACTGCCCAGGAGCCAGA-3′ and reverse, 5′-TGGCACCAGTGTCCGGATTA-3′;
N-cadherin forward, 5′-CGAATGGATGAAAGACCCATCC-3′ and reverse,
5′-GGAGCCACTGCCTTCATAGTCAA-3′; vimentin, forward,
5′-GCTGAATGACCGCTTCGCCAACT'-3′ and revere,
5′-GCTCCCGCATCTCCTCCTCGTA-3′; MMP-2, forward,
5′-CTGCGGTTTTCTCGAATCCA-3′ and reverse, 5′-GGGTATCCATCGCCATGCT-3′;
MMP-9 forward, 5′-CCCTGGAGACCTGAGAACCA-3′ and revere,
5′-CCACCCGAGTGTAACCATAGC-3′; U6, forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; GAPDH forward,
5′-GAGTCAACGGATTTGGTCGTATTG-3′ and reverse,
5′-CCTGGAAGATGGTGATGGGATT-3′. SYBR Green II was used for detection
of signal fluorescence and ROX was used as the control
fluorescence. Reverse transcription was performed on a GeneAmp PCR
system 9700 (Applied Biosystems; Thermo Fisher Scientific, Inc.),
and qPCR was performed on ABI 7500 real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). All samples were
processed at the same time to avoid inter-experiment variance. The
thermocycling conditions were as follows: A holding step at 95°C
for 30 sec, and 40 cycles of 95°C for 5 sec and 60°C for 30
sec.
Each experiment was performed in triplicate, and the
mean value was used for analysis. The expression level was analyzed
by using the ΔΔCq method. ΔCq values represent normalized target
genes levels with respect to the internal control. ΔΔCq values were
calculated as the ΔCq of each test sample minus the mean ΔCq of the
calibrator samples for each target gene. U6 small nuclear RNA and
GAPDH mRNA were used for normalization. The fold change was
calculated using the method 2−ΔΔCq (34). Each treatment was performed in
triplicate in three independent experiments.
Cell proliferation and invasion
To study the role of miR-199a-5p mimic in the
proliferation of HO-8910 and ES-2 cells, 5×104 cells
were seeded in a 96-well plate and allowed to grow for 24 h in DMEM
at 37°C with 10% FBS. The medium was subsequently removed, and the
cells were transfected with 50 nM miR-199a-5p mimic or miR-NC for
24 h at 37°C. Cell proliferation 5-bromo-2-deoxyuridine
(BrdU)-ELISA colorimetric kit (Roche Diagnostics, Indianapolis, IN,
USA) was used to assess cell proliferation according to the
manufacturer's instructions. Optical densities were determined at a
wavelength of 530 nm using a spectrophotometric plate reader.
Transwell Matrigel invasion assay
The effect of miR-199a-5p on the invasive ability of
OC cells was also investigated using Transwell chambers (8-mm pore
size; Corning, USA) pre-coated with Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA) that included extracellular matrix
proteins. In brief, after serum-starvation for 24 h,
2×105 cells were suspended in 100 µl serum-free DMEM,
and seeded in the top chamber, and 600 µl DMEM containing 10% FBS
was added to the lower chamber. After 24 h incubation at 37°C in a
5% CO2 atmosphere, cells that remained on the upper
surface of the membrane were removed by cotton swabs and
penetrating cells were fixed in methanol, and then stained with
0.1% crystal violet at 25°C for 12 h. Cell invasion was quantified
by counting cells on the lower surface using phase contrast
microscopy with ×40 magnification for eight fields of view.
Western blot analysis
For protein extraction, HO-8910 and ES-2 cells were
washed twice in cold phosphate-buffered saline and subsequently
lysed in radioimmunoprecipitation assay buffer (Beyotime Institute
of Biotechnology, Haimen, China) with a protease inhibitor-PMSF
(Beyotime Institute of Biotechnology). The protein concentration of
cell lysates was quantified by using the OCA kit (Beyotime
Institute of Biotechnology). Equal quantities (50 µg) of total
protein were separated by 10% SDS-PAGE, and transferred to a
polyvinylidene fluoride membrane (Merck Millipore). The membranes
were blocked in 5% skimmed milk diluted with Tris-buffered saline
Tween-20 at room temperature for 1 h and incubated overnight at 4°C
with primary anti-nuclear factor-κB1 (NF-κB1) antibody (cat. no.
13586, 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA).
The membranes were subsequently incubated with a goat anti-rabbit
horseradish peroxidase-conjugated immunoglobulin G secondary
antibody (cat. no. A0208, 1:2,000; Beyotime Institute of
Biotechnology) for 2 h. The proteins were visualized using enhanced
chemiluminescence-plus reagents (Beyotime Institute of
Biotechnology). The density of the bands was measured using Image J
software (version 1.5b; National Institutes of Health USA), and the
values were normalized to the densitometric values of GAPDH
(1:1,000; Cell Signaling Technology, Inc.) in each sample.
Measurement of MMP-2 and MMP-9
levels
ELISA kits (USCN Life Science, Wuhan, China) was
used to determine the levels of MMP-2 and −9 in the cells culture
supernatants based on the manufacturer's instructions.
Luciferase reporter assay
NF-κB1 was predicted to be a direct target of
miR-199a-5p by using online prediction software RNA22 (https://cm.jefferson.edu/rna22/Interactive/),
TargetScan (http://www.targetscan.org/vert_71/) and PITA
(https://genie.weizmann.ac.il/pubs/mir07/mir07_data.html).
Then, HO-8910 and ES-2 cells (2×106/well) were seeded in
24-well plates and incubated overnight at 37°C prior to
transfection. The cells were co-transfected with the 20 ng of
NF-κB1-3′ untranslated region (UTR) wild-type (WT) or mutant (MUT)
reporter plasmid, 50 nm of miR-199a-5p mimic or miR-negative
control (NC) (Promega Corporation, Madison, WI, USA) using
Lipofectamine™ 2000. Firefly and Renilla
luciferase activities were quantified using a dual luciferase
reporter system (Promega Corporation) according to the
manufacturer's instructions 48 h following co-transfection. Each
treatment was performed in triplicate in three independent
experiments.
Statistical analysis
All statistical analyses were performed using SPSS
(version 18.0; SPSS, Inc., Chicago, IL, USA). Data from each group
are expressed as the mean ± standard error of the mean. One-way
analysis of variance was used to compare data sets. Bonferroni test
was used to compare the variances at the same time with different
means and the Tamhane's T2 test was used to compare variances.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-199a-5p level is decreased in OC
cell lines
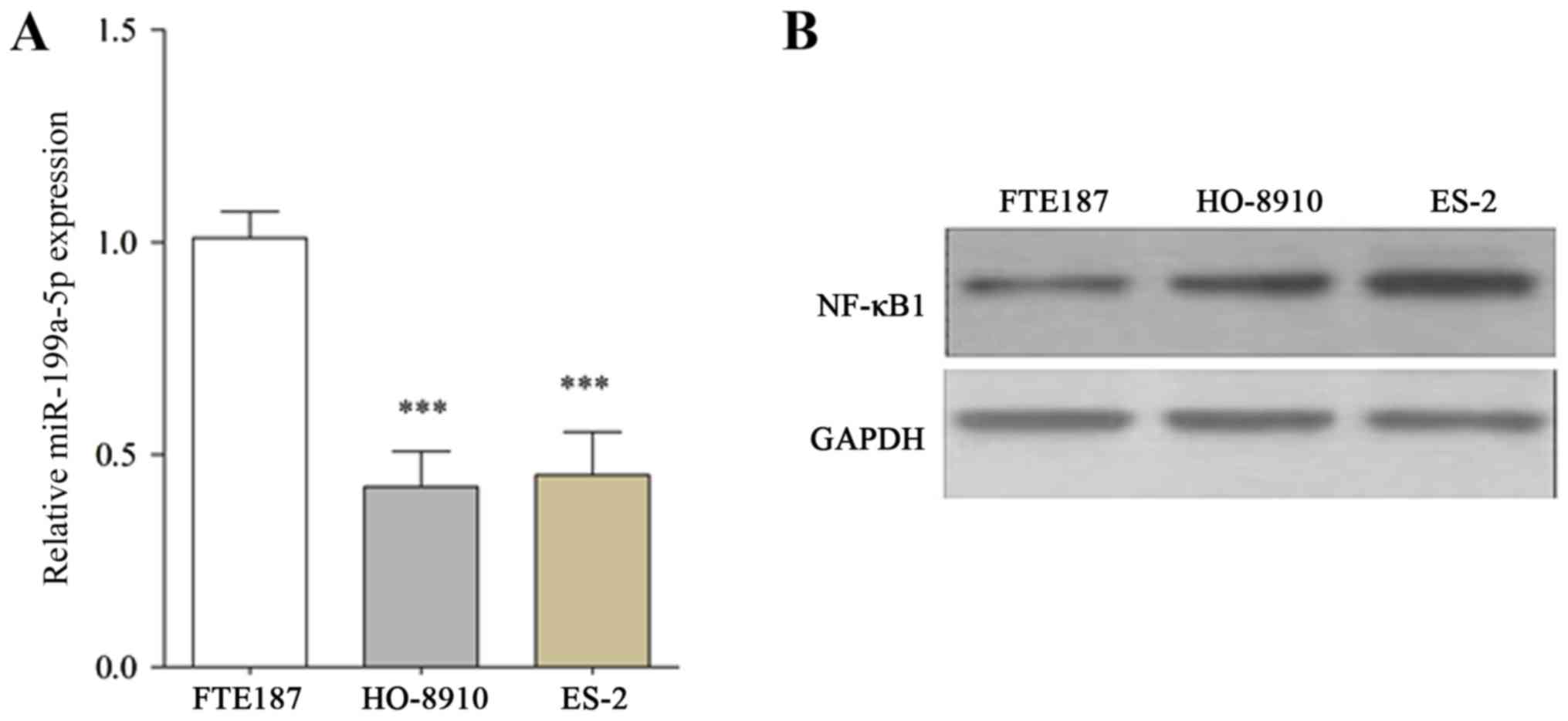
The findings of the present study indicated that
miR-199a-5p expression was significantly reduced in HO-8910 and
ES-2 cells compared with FTE187 cells (Fig. 1A). Furthermore, NF-κB1 was predicted
to be a direct target of miR-199a-5p by using online prediction
software RNA22, TargetScan and PITA. The protein expression of
NF-κB1, a target of miR-199a-5p, was further assessed in HO-8910
and ES-2 cell lines. The results indicate that NF-κB1 expression in
HO-8910 and ES-2 cells was markedly increased compared with FTE187
cells (Fig. 1B).
Cell proliferation and invasion are
inhibited by miR-199a-5p
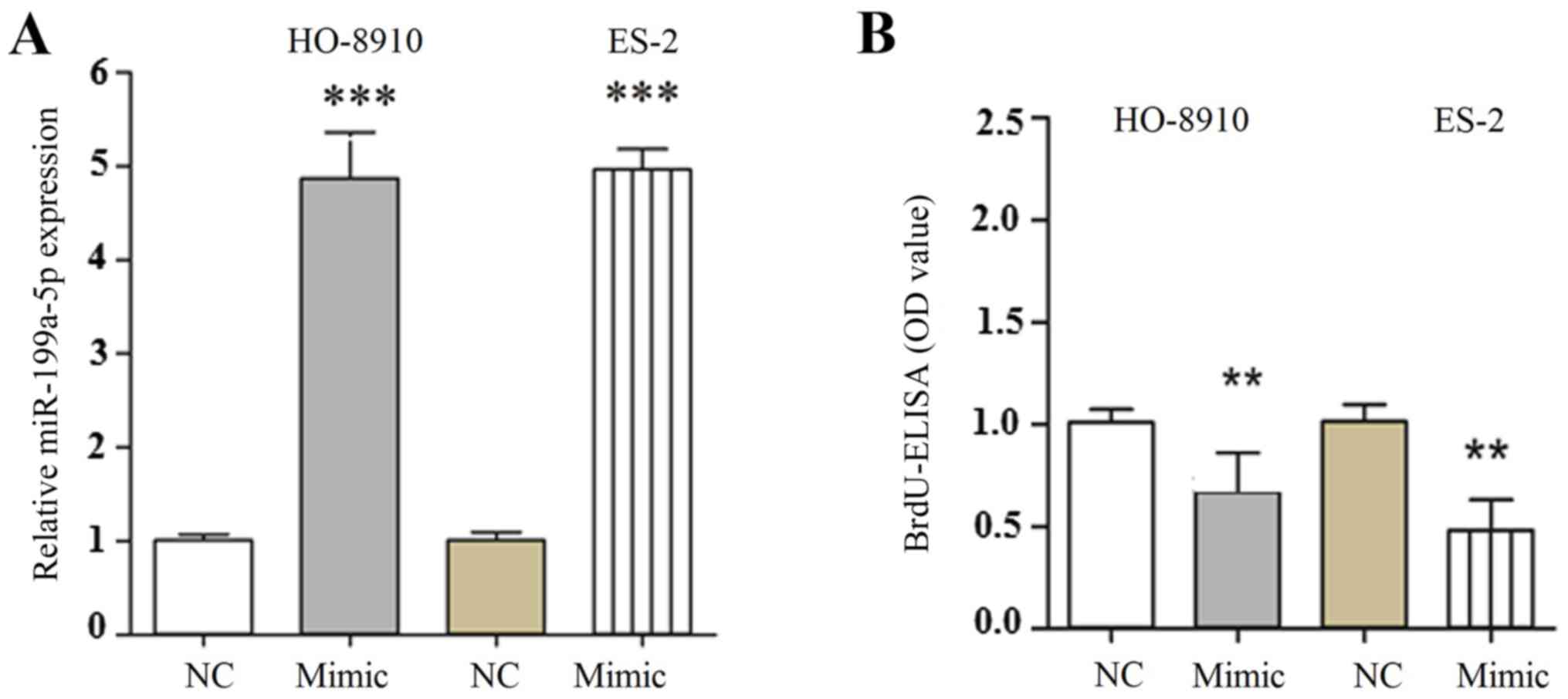
The level of miR-199a-5p mRNA was significantly
upregulated following transfection with miR-199a-5p mimic compared
with the miR-NC group (Fig. 2A). To
investigate the role of miR-199a-5p in the proliferation of OC
cells, HO-8910 and ES-2 cell were transfected with miR-199a-5p
mimic or miR-NC. Results from the BrdU-ELISA assay indicated that
exogenous expression of miR-199a-5p significantly suppressed the
proliferation of HO-8910 and ES-2 cells (Fig. 2B). These findings indicated that the
upregulation of miR-199a-5p was able to inhibit the proliferation
of OC cells. To understand whether the exogenous expression of
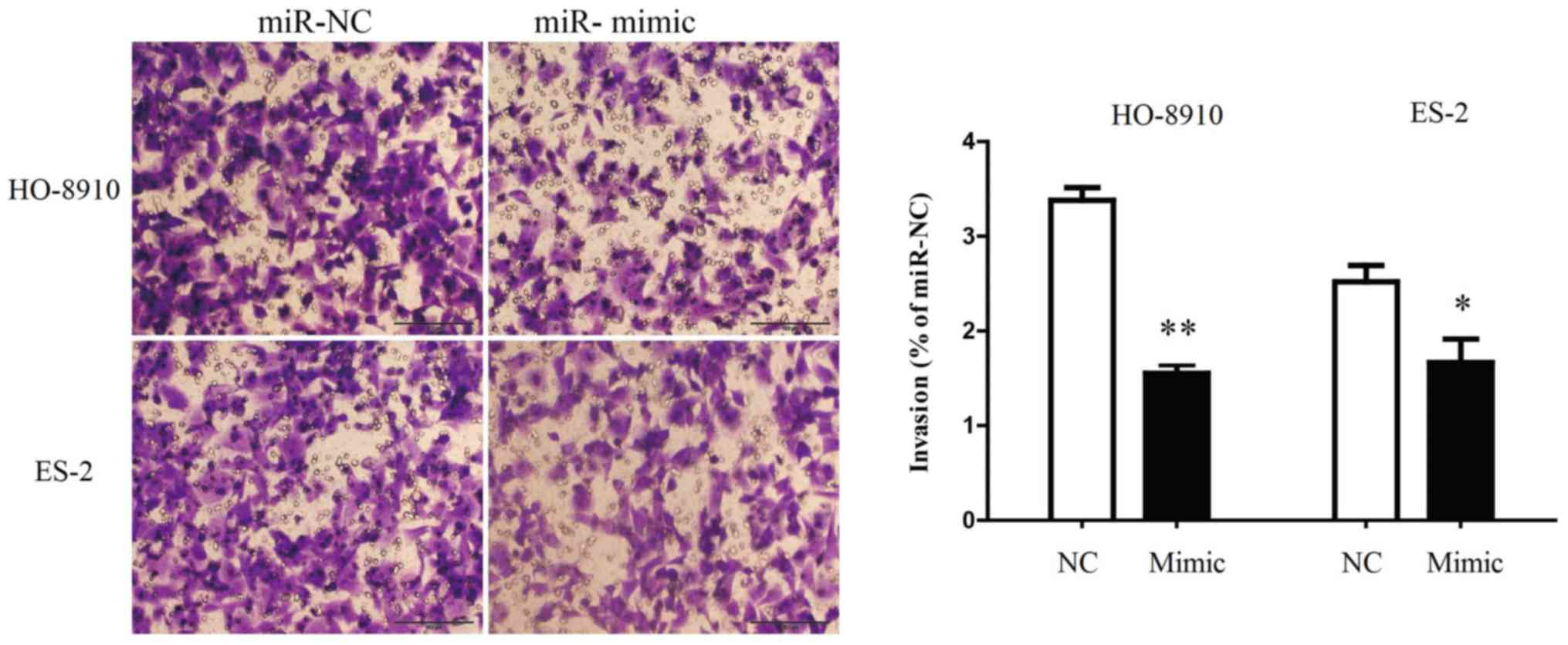
miR-199a-5p was able to inhibit the invasion of OC cells,
miR-199a-5p mimic was transfected into HO-8910 and ES-2 cells and
the invasive ability of the cells was evaluated by Transwell
invasion assay. It was demonstrated that the number of HO-8910 and
ES-2 cells migrating through the Transwell membrane was
significantly decreased in the miR-199a-5p mimic group compared
with the miR-NC group (Fig. 3).
Therefore, these findings indicated that the upregulation of
miR-199a-5p suppressed the invasion of OC cells.
Effect of miR-199a-5p overexpression
on MMP-2 and 9 expression in OC cells
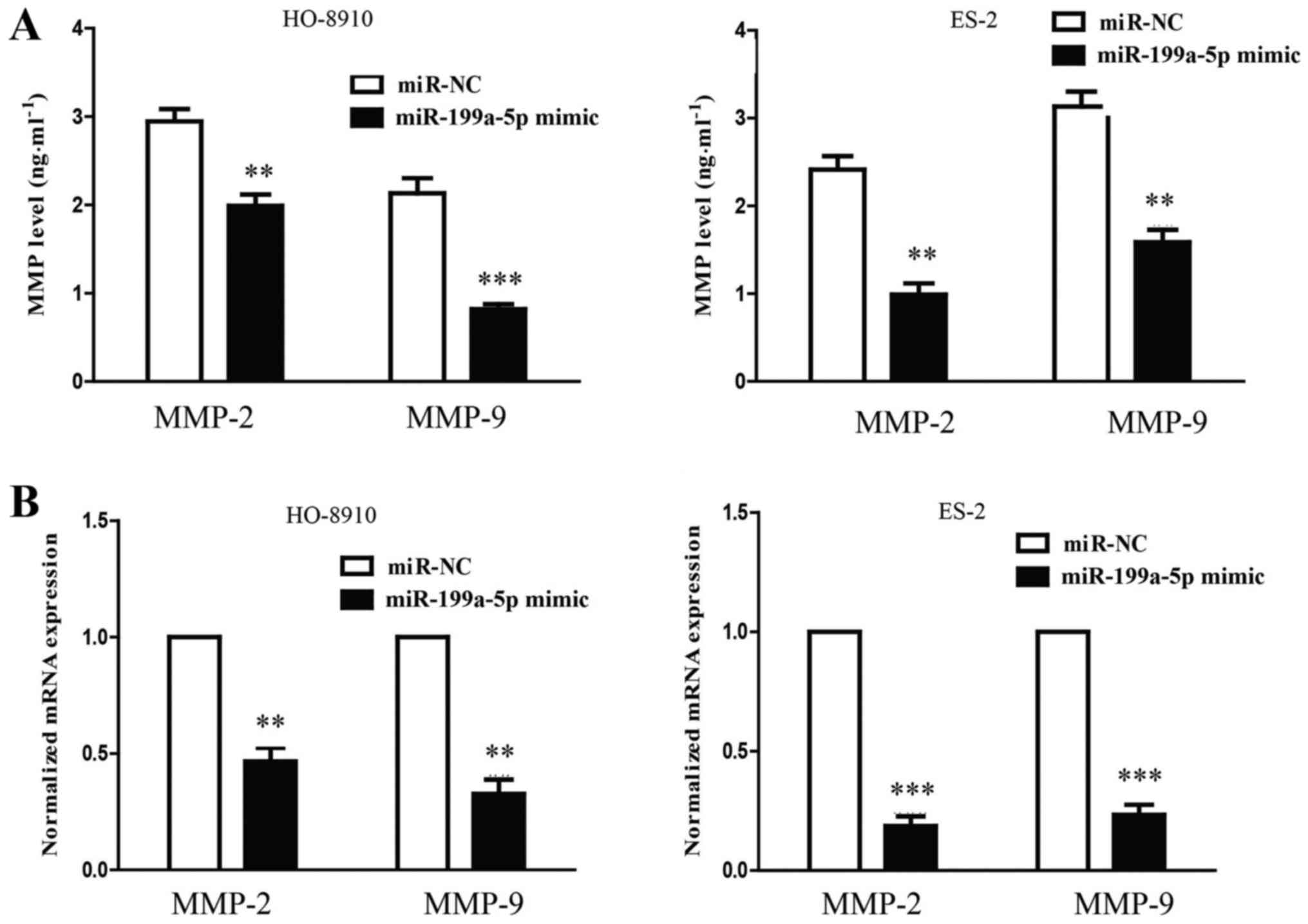
MMPs may be responsible for the impaired invasion of
the cells that were transfected with miR-199a-5p mimic. To confirm
this hypothesis, the mRNA levels of MMP-2 and 9 were detected by
RT-qPCR. The overexpression of miR-199a-5p resulted in significant
reductions in the levels of MMP-2 and −9 mRNA (Fig. 4). The results suggested that the
upregulation of hsa-miR-199a-5p was able to suppress the invasive
ability of HO-8910 and ES-2 cells by potentially downregulating
MMP-2 and 9.
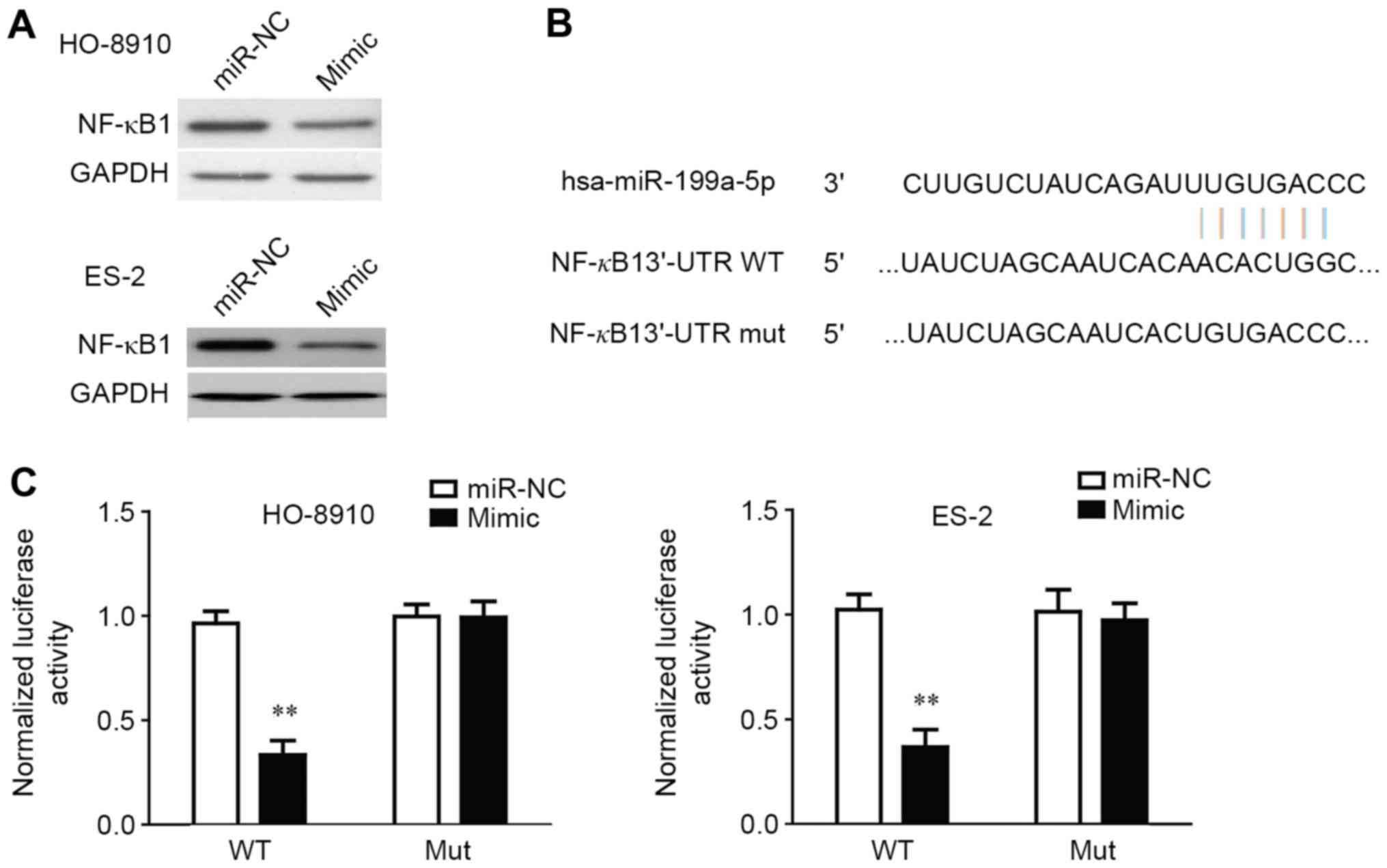
NF-κB1 is a direct target of
miR-199a-5p in OC cells
As NF-κB1 was predicted to be a potential direct
target of miR-199a-5p by prediction software (RNA22, TargetScan and
PITA), the expression of NF-κB1 protein in HO-8910 and ES-2 cells
that were transfected with miR-199a-5p mimic was determined by
western blotting (Fig. 5A). The
results demonstrated that NF-κB1 expression was decreased in the
cells that were transfected with miR-199a-5p mimic compared with
the cells that were transfected with miR-199a-5p NC (Fig. 5A). To further confirm whether NF-κB1
is a direct target of miR-15a-5p, the 3′-UTR of NF-κB1 was cloned
into a luciferase reporter vector, and the putative miR-199a-5p
binding site in the NF-κB1 3′-UTR was mutated (Fig. 5B). Luciferase reporter assay was used
to determine whether the miR-15a-5p mimic is able to interact with
the NF-κB1 3′-UTR. The exogenous expression of the miR-199a-5p
mimic was able to significantly suppress the luciferase activity of
NF-κB1 3′-UTRWT (Fig. 5C). The effect
of miR-199a-5p was abolished by mutating the miR-199a-5p-binding
site in the NF-κB1 3′-UTR, which suggested that miR-199a-5p was
able to directly negatively regulate NF-κB1.
Discussion
Studies have demonstrated that miRNAs are
dysregulated in multiple types of cancer (35–37).
Therefore, it is critical for the identification of cancer-specific
miRNAs and their targets in order to understand their role in
oncogenesis and to identify novel therapeutic targets (38–40). The
present study focused on the function of miR-199a-5p in the
pathogenesis of OC. Initially, the level of miR-199a-5p was
detected in OC cells by RT-qPCR. It was demonstrated that the level
of miR-199a-5p was significantly reduced in ovarian cancer cells
compared with FTE187 normal human fallopian tube epithelial cells.
The results indicated in the present study are consistent with
other reports, which have demonstrated that miR-199a-5p was
significantly downregulated in colorectal cancer, breast cancer,
glioblastoma and oral squamous cell carcinoma (41,42).
Therefore, the downregulation of miR-199a-5p is exhibited by
multiple types of cancer cells and tissues, indicating that
decreased expression of miR-199a-5p may have a critical role in
tumorigenesis.
As numerous miRNAs are decreased in various cancer
types, it was hypothesized that miRNAs may normally function as
tumor suppressor genes (43–45). Therefore, the authors hypothesized
that miR-199a-5p was an inhibitory factor of growth in OC cells. As
the level of miR-199a-5p was downregulated in cancer cells and
tissues, it was hypothesized that the upregulation of miR-199a-5p
would lead to cell growth arrest. Using the BrdU-ELISA assay, it
was demonstrated that HO-8910 and ES-2 cells that were transfected
with the miR-199a-5p mimic exhibited decreased growth compared with
the cells transfected with miR-NC.
It has been reported that miR-199a-5p affects the
growth, metastasis and apoptosis of cancer cells potentially by
targeting cyclin dependent kinase 6, Rho associated coiled-coil
containing protein kinase 1 and cMet (30,31,46–49).
Although bioinformatic tools might help to reveal the putative mRNA
targets of miRNAs, experimental procedures are required for
validation. In the resent paper, it was demonstrated that
miR-199a-5p was able to target the NF-κB1 miRNA, therefore
revealing a potential mechanism associated with ovarian
oncogenesis.
As a member of the Rel/NF-κB transcription factor
family, NF-κB1 has critical roles in the regulation of immune
responses, embryo and cell lineage development, cell-cycle
progression, cell apoptosis and tumorigenesis (50–52). The
data in the present study indicated that NF-κB1 was a target of
miR-199a-5p. First, using western blotting, it was confirmed that
the transfection of miR-199a-5p mimic was able to cause a marked
decrease in the levels of NF-κB1 protein. In addition, it was
indicated that miR-199a-5p was able to directly regulate NF-κB1
expression, as the seed region of miR-199a-5p is able to bind with
the 3′ UTR of NF-κB1 mRNA. Furthermore, the luciferase activity of
the NF-κB1 3′-UTR was specifically responsive to miR-199a-5p
upregulation. Indeed, mutation of the miR-199a-5p binding site
abolished the effect of miR-199a-5p on the regulation of luciferase
activity.
NF-κB1 facilitates invasion and metastasis of cancer
cells partly by transcriptional regulation of MMP-2 and MMP-9
(53). Ferrer-Marin et al
(53) have demonstrated that miR-9
suppressed the migration and invasion of uveal melanoma cells
partly by targeting NF-κB1 expression and downregulating, MMP-2 and
−9, its downstream molecules. An additional study has indicated
that miR-9 was able to inhibit the metastasis of melanoma cells by
suppressing E-cadherin expression via targeting NF-κB1 (54). In the present study, it was
demonstrated that the expression of MMP-2 and MMP-9 was altered in
the same pattern as NF-κB1 expression upon the exogenous expression
of miR-199a-5p. Therefore, the regulation of MMP-2 and MMP-9 by
NF-κB1 may contribute to the invasion and metastasis of OC cells
that is mediated by NF-κB1.
In summary, the present study indicated that
miR-199a-5p was downregulated in OC cells compared with the normal
human fallopian tube epithelial cell line. The transfection of
miR-199a-5p inhibited the proliferation and invasion of OC cells by
directly targeting NF-κB1. This novel miR-199a-5p/NF-κB1 axis might
provide new insights into the molecular mechanisms underlying the
progression and metastasis of OC, and the overexpression of
miR-199a-5p might be a potential therapeutic strategy for the
treatment of OC in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Basic Natural
Science Foundation of Xinchang People's Hospital of Zhejiang
Province (Zhejiang, China; grant no. 81201101).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and BY contributed to experiments and data
analyses. ZW contributed to the design of the project and writing
of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhou X, Hu Y, Dai L, Wang Y, Zhou J, Wang
W, Di W and Qiu L: MicroRNA-7 inhibits tumor metastasis and
reverses epithelial-mesenchymal transition through AKT/ERK1/2
inactivation by targeting EGFR in epithelial ovarian cancer. PLoS
One. 9:e967182014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu T, Hou L and Huang Y: EZH2-specific
microRNA-98 inhibits human ovarian cancer stem cell proliferation
via regulating the pRb-E2F pathway. Tumour Biol. 35:7239–7247.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
James PA, Sawyer S, Boyle S, Young MA,
Kovalenko S, Doherty R, McKinley J, Alsop K, Beshay V, Harris M, et
al: Large genomic rearrangements in the familial breast and ovarian
cancer gene BRCA1 are associated with an increased frequency of
high risk features. Fam Cancer. 14:287–295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koensgen D, Bruennert D, Ungureanu S,
Sofroni D, Braicu EI, Sehouli J, Sümnig A, Delogu S, Zygmunt M,
Goyal P, et al: Polymorphism of the IL-8 gene and the risk of
ovarian cancer. Cytokine. 71:334–338. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xing BL, Li T, Tang ZH, Jiao L, Ge SM,
Qiang X and OuYang J: Cumulative methylation alternations of gene
promoters and protein markers for diagnosis of epithelial ovarian
cancer. Genet Mol Res. 14:4532–4540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu Q, Ding YY, Song LX and Xu JF:
Correlation of UGT1A1 and ERCC1 gene polymorphisms with the outcome
of combined irinotecan plus cisplatin treatment in recurrent
ovarian cancer. Genet Mol Res. 14:7241–7247. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yan H, Tong J, Lin X, Han Q and Huang H:
Effect of the WWOX gene on the regulation of the cell cycle and
apoptosis in human ovarian cancer stem cells. Mol Med Rep.
12:1783–1788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu Z, Yu YQ, Ge YZ, Zhu JG, Zhu M, Zhao
YC, Xu LW, Yang XB, Geng LG, Dou QL and Jia RP: MicroRNA expression
profiles in muscle-invasive bladder cancer: Identification of a
four-microRNA signature associated with patient survival. Tumour
Biol. 36:8159–8166. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Itesako T, Seki N, Yoshino H, Chiyomaru T,
Yamasaki T, Hidaka H, Yonezawa T, Nohata N, Kinoshita T, Nakagawa M
and Enokida H: The microRNA expressions signature of bladder cancer
by deep sequencing: The functional significance of the miR-195/497
cluster. PLoS One. 9:e843112014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pignot G, Cizeron-Clairac G, Vacher S,
Susini A, Tozlu S, Vieillefond A, Zerbib M, Lidereau R, Debre B,
Amsellem-Ouazana D and Bieche I: microRNA expression profile in a
large series of bladder tumors: Identification of a 3-miRNA
signature associated with aggressiveness of muscle-invasive bladder
cancer. Int J Cancer. 132:2479–2491. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang AM, Ma K, Song Y, Wang B, Feng Y,
Liu L and Xia X: Genetic polymorphisms of the IFNλ genes are
associated with biochemical features in Han Chinese with HCV
infection from Yunnan Province, China. Infect Genet Evol.
21:161–165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang AM, Ma K, Song Y, Feng Y, Duan H,
Zhao P, Wang B, Xu G, Li Z and Xia X: Mitochondrial DNAs decreased
and correlated with clinical features in HCV patients from Yunnan,
China. Mitochondrial DNA A DNA Mapp Seq Anal. 27:2516–2519.
2016.PubMed/NCBI
|
|
13
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jabbari N, Reavis AN and McDonald JF:
Sequence variation among members of the miR-200 microRNA family is
correlated with variation in the ability to induce hallmarks of
mesenchymal-epithelial transition in ovarian cancer cells. J
Ovarian Res. 7:122014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maqbool R, Ismail R and Hussain M:
Mutations in MicroRNA genes and they binding sites are infrequently
associated with human colorectal cancer in the kashmiri population.
Microrna. 2:219–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
De M, attos-Arruda L, Bottai G, Nuciforo
PG, Di Tommaso L, Giovannetti E, Peg V, Losurdo A, Pérez-Garcia J,
Masci G, Corsi F, et al: MicroRNA-21 links
epithelial-to-mesenchymal transition and inflammatory signals to
confer resistance to neoadjuvant trastuzumab and chemotherapy in
HER2-positive breast cancer patients. Oncotarget. 6:37269–37280.
2015.PubMed/NCBI
|
|
17
|
Wang X, Li Y, Qi W, Zhang N, Sun M, Huo Q,
Cai C, Lv S and Yang Q: MicroRNA-99a inhibits tumor aggressive
phenotypes through regulating HOXA1 in breast cancer cells.
Oncotarget. 6:32737–32747. 2015.PubMed/NCBI
|
|
18
|
Sestini S, Boeri M, Marchiano A, Pelosi G,
Galeone C, Verri C, Suatoni P, Sverzellati N, La Vecchia C, Sozzi G
and Pastorino U: Circulating microRNA signature as liquid-biopsy to
monitor lung cancer in low-dose computed tomography screening.
Oncotarget. 6:32868–32877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui R, Kim T, Fassan M, Meng W, Sun HL,
Jeon YJ, Vicentini C, Tili E, Peng Y, Scarpa A, et al: MicroRNA-224
is implicated in lung cancer pathogenesis through targeting
caspase-3 and caspase-7. Oncotarget. 6:21802–21815. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fang Y, Xu C and Fu Y: MicroRNA-17-5p
induces drug resistance and invasion of ovarian carcinoma cells by
targeting PTEN signaling. J Biol Res (Thessalon). 22:122015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Niu K, Shen W, Zhang Y, Zhao Y and Lu Y:
MiR-205 promotes motility of ovarian cancer cells via targeting
ZEB1. Gene. 574:330–336. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu T, Yuan J, Wang Y, Gong C, Xie Y and
Li H: MiR-661 contributed to cell proliferation of human ovarian
cancer cells by repressing INPP5J expressions. Biomed Pharmacother.
75:123–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ye Z, Zhao L, Li J, Chen W and Li X:
miR-30d blocked transforming growth factor β1-induced
epithelial-mesenchymal transition by targeting snail in ovarian
cancer cells. Int J Gynecol Cancer. 25:1574–1581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu X, Ma L, Rao Q, Mao Y, Xin Y, Xu H, Li
C and Wang X: MiR-1271 inhibits ovarian cancer growth by targeting
cyclin G1. Med Sci Monit. 21:3152–3158. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo T, Yu W, Lv S, Zhang C and Tian Y:
MiR-302a inhibits the tumorigenicity of ovarian cancer cells by
suppression of SDC1. Int J Clin Exp Pathol. 8:4869–4880.
2015.PubMed/NCBI
|
|
26
|
Mussnich P, Rosa R, Bianco R, Fusco A and
D'Angelo D: MiR-199a-5p and miR-375 affect colon cancer cell
sensitivity to cetuximab by targeting PHLPP1. Expert Opin Ther
Targets. 19:1017–1026. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo W, Qiu Z, Wang Z, Wang Q, Tan N, Chen
T, Chen Z, Huang S, Gu J, Li J, et al: MiR-199a-5p is negatively
associated with malignancies and regulates glycolysis and lactate
production by targeting hexokinase 2 in liver cancer. Hepatology.
62:1132–1144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim BK, Yoo HI, Kim I, Park J and Kim Yoon
S: FZD6 expression is negatively regulated by miR-199a-5p in human
colorectal cancer. BMB Rep. 48:360–366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee JM, Heo MJ, Lee CG, Yang YM and Kim
SG: Increase of miR-199a-5p by protoporphyrin IX, a photocatalyzer,
directly inhibits E2F3, sensitizing mesenchymal tumor cells to
anti-cancer agents. Oncotarget. 6:3918–3931. 2015.PubMed/NCBI
|
|
30
|
Zhao X, He L, Li T, Lu Y, Miao Y, Liang S,
Guo H, Bai M, Xie H, Luo G, et al: SRF expedites metastasis and
modulates the epithelial to mesenchymal transition by regulating
miR-199a-5p expression in human gastric cancer. Cell Death Differ.
21:1900–1913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu Y, Liu J, Jiang B, Chen J, Fu Z, Bai F,
Jiang J and Tang Z: MiR-199a-5p loss up-regulated DDR1 aggravated
colorectal cancer by activating epithelial-to-mesenchymal
transition related signaling. Dig Dis Sci. 59:2163–2172. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He XJ, Ma YY, Yu S, Jiang XT, Lu YD, Tao
L, Wang HP, Hu ZM and Tao HQ: Up-regulated miR-199a-5p in gastric
cancer functions as an oncogene and targets klotho. BMC Cancer.
14:2182014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yi H, Liang B, Jia J, Liang N, Xu H, Ju G,
Ma S and Liu X: Differential roles of miR-199a-5p in
radiation-induced autophagy in breast cancer cells. FEBS Lett.
587:436–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Saito Y, Nakaoka T and Saito H:
microRNA-34a as a therapeutic agent against human cancer. J Clin
Med. 4:1951–1959. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ono S, Lam S, Nagahara M and Hoon DS:
Circulating microRNA Biomarkers as liquid biopsy for cancer
patients: Pros and cons of current assays. J Clin Med. 4:1890–1907.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miyazaki T, Ikeda K, Sato W, Horie-Inoue
K, Okamoto K and Inoue S: MicroRNA library-based functional
screening identified androgen-sensitive miR-216a as a player in
bicalutamide resistance in prostate cancer. J Clin Med.
4:1853–1865. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
An F, Olaru AV, Mezey E, Xie Q, Li L,
Piontek KB and Selaru FM: MicroRNA-224 induces G1/S checkpoint
release in liver cancer. J Clin Med. 4:1713–1728. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Horsham JL, Kalinowski FC, Epis MR, Ganda
C, Brown RA and Leedman PJ: Clinical potential of microRNA-7 in
cancer. J Clin Med. 4:1668–1687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kelly BD, Miller N, Sweeney KJ, Durkan GC,
Rogers E, Walsh K and Kerin MJ: A Circulating MicroRNA signature as
a biomarker for prostate cancer in a high risk group. J Clin Med.
4:1369–1379. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stiuso P, Potenza N, Lombardi A,
Ferrandino I, Monaco A, Zappavigna S, Vanacore D, Mosca N,
Castiello F, Porto S, et al: MicroRNA-423-5p promotes autophagy in
cancer cells and is increased in serum from hepatocarcinoma
patients treated with sorafenib. Mol Ther Nucleic Acids.
4:e2332015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Slattery ML, Mullany LE, Sakoda LC,
Samowitz WS, Wolff RK, Stevens JR and Herrick JS: Expression of
Wnt-signaling pathway genes and their associations with miRNAs in
colorectal cancer. Oncotarget. 9:6075–6085. 2017.PubMed/NCBI
|
|
43
|
Nunez Lopez YO, Victoria B, Golusinski P,
Golusinski W and Masternak MM: Characteristic miRNA expression
signature and random forest survival analysis identify potential
cancer-driving miRNAs in a broad range of head and neck squamous
cell carcinoma subtypes. Rep Pract Oncol Radiother. 23:6–20. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Deb B, Uddin A and Chakraborty S: miRNAs
and ovarian cancer: An overview. J Cell Physiol. 233:3846–3854.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Weiner-Gorzel K, Dempsey E, Milewska M,
McGoldrick A, Toh V, Walsh A, Lindsay S, Gubbins L, Cannon A,
Sharpe D, et al: Overexpression of the microRNA miR-433 promotes
resistance to paclitaxel through the induction of cellular
senescence in ovarian cancer cells. Cancer Med. 4:745–758. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou J, Liu R, Wang Y, Tang J, Tang S,
Chen X, Xia K, Xiong W, Xu D, Wang S, et al: miR-199a-5p regulates
the expression of metastasis-associated genes in B16F10 melanoma
cells. Int J Clin Exp Pathol. 7:7182–7190. 2014.PubMed/NCBI
|
|
47
|
Lin HS, Gong JN, Su R, Chen MT, Song L,
Shen C, Wang F, Ma YN, Zhao HL, Yu J, et al: miR-199a-5p inhibits
monocyte/macrophage differentiation by targeting the activin A type
1B receptor gene and finally reducing C/EBPα expression. J Leukoc
Biol. 96:1023–1035. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Raimondi L, Amodio N, Di Martino MT,
Altomare E, Leotta M, Caracciolo D, Gullà A, Neri A, Taverna S,
D'Aquila P, et al: Targeting of multiple myeloma-related
angiogenesis by miR-199a-5p mimics: In vitro and in vivo anti-tumor
activity. Oncotarget. 5:3039–3054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu ZY, Lu L, Liang J, Guo XR, Zhang PH and
Luo SJ: Keloid microRNA expression analysis and the influence of
miR-199a-5p on the proliferation of keloid fibroblasts. Genet Mol
Res. 13:2727–2738. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Oltulu YM, Coskunpinar E, Ozkan G, Aynaci
E, Yildiz P, Isbir T and Yaylim I: Investigation of NF-κB1 and
NF-κBIA gene polymorphism in non-small cell lung cancer. Biomed Res
Int. 2014:5303812014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Huang H, Ma L, Li J, Yu Y, Zhang D, Wei J,
Jin H, Xu D, Gao J and Huang C: NF-κB1 inhibits c-Myc protein
degradation through suppression of FBW7 expression. Oncotarget.
5:493–505. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yu Y, Zhang D, Huang H, Li J, Zhang M, Wan
Y, Gao J and Huang C: NF-κB1 p50 promotes p53 protein translation
through miR-190 downregulation of PHLPP1. Oncogene. 33:996–1005.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ferrer-Marin F, Gutti R, Liu ZJ and
Sola-Visner M: MiR-9 contributes to the developmental differences
in CXCR-4 expression in human megakaryocytes. J Thromb Haemost.
12:282–285. 2014. View Article : Google Scholar
|
|
54
|
Chen P, Price C, Li Z, Li Y, Cao D, Wiley
A, He C, Gurbuxani S, Kunjamma RB, Huang H, et al: miR-9 is an
essential oncogenic microRNA specifically overexpressed in mixed
lineage leukemia-rearranged leukemia. Proc Natl Acad Sci USA.
110:11511–11516. 2013. View Article : Google Scholar : PubMed/NCBI
|