Introduction
Prostate cancer (PCa) is one of the most common
malignancies in men globally and the second leading cause of cancer
associated mortality in developed countries (1,2). Like
other cancers, PCa is considered to be a disease which caused by
age, diet and gene aberrations (3).
Accumulating evidences have demonstrated that a series of genes and
pathways involved in the occurrence, progression and metastasis of
PCa (4). At present, the underlying
mechanism of PCa occurrence is still unclear, which limits the
diagnosis and therapy. Therefore, it is urgent to identify the key
genes and pathways involved in the occurrence of PCa (2,5).
Microarray is a useful tool for analysis of gene
expression that can be applied to disease diagnosis and targeted
therapy (6,7). During the past decade, hundreds of
differentially expressed genes (DEGs) participated in biological
processes, cell component, molecular functions and pathways of PCa
were identified by microarray technology (8,9). However,
previous studies of DEGs analysis have shown relative limitations,
for example, no reliable biomarker was identified that could
distinguish tumors from normal tissues (10). Therefore, gene expression in the
occurrence of PCa needs to be further analyzed by microarray
combining bioinformatics technology at present.
In the present study, Gene expression profile
(GSE55945) was downloaded from Gene Expression Omnibus (GEO)
(http://www.ncbi.nlm.nih.gov/geo/). The
gene expression profile was analyzed, and the DEGs were identified
between PCa group and normal group. Subsequently, gene ontology
(GO) analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analysis and protein-protein interaction (PPI) analysis of
DEGs were performed. Finally, the expression of screened key genes
was verified by immunohistochemistry (IHC). The present study aimed
to identify key genes and pathways which involved in the occurrence
of PCa and explored the potential biomarker for prognosis,
diagnosis and drug targets.
Materials and methods
Expression Profile Microarray
Gene expression profile (GSE55945) was downloaded
from GEO (http://www.ncbi.nlm.nih.gov/geo/). GSE55945 was based
on GPL570 platform (Affymetrix Human Genome U133 Plus 2.0 Array),
which was submitted by Arredouani et al, and contained 21
samples, including 18 PCa samples and 8 normal prostate samples
(8).
Identification of DEGs
The gene expression profile was analyzed by Morpheus
online tools (https://software.broadinstitute.org/morpheus/) as
previous study (11). The DEGs were
identified between PCa group and normal group. The significance of
DEGs was identified by classical t-test. The change ≥twofold
and P<0.05 was considered to indicate a statistically
significant difference.
GO analysis and KEGG pathway analysis
of DEGs
In order to analyse the function and pathway of the
DEGs, DAVID database (https://david.ncifcrf.gov/) was used for GO analysis
and KEGG pathway analysis as previous study (7). P<0.05 was considered to indicate a
statistically significant difference.
PPI network and module analysis
Search Tool for the Retrieval of Interacting Genes
(STRING; http://string-db.org/cgi/input.pl) was a database that
could assess the protein-protein interaction. The DEGs were mapped
to STRING, and a score >0.4 was considered to be significant.
Then, the Cytoscape software (version 3.3.0) was used to construct
PPI networks. Finally, the modules of PPI network were screened by
the plug-in Molecular Complex Detection (MCODE). In addition, the
pathway analysis was performed in the modules. P<0.05 was
considered to indicate a statistically significant difference.
Patients and tissue samples
The tissue microarray was purchased from Alenabio
Co., Ltd (Xian, China) including 60 PCa samples from patients and
10 normal prostate tissue samples from healthy donors. The
procedures performed in this study involving human patients were in
accordance with the ethical standards of the institutional and/or
national research committee and with the 1964 Helsinki declaration
and its later amendments or comparable ethical standards. The
present study was approved by the Research Ethics Committee of
Weifang Medical University (Weifang, China).
IHC validation
IHC was performed as previous study (12). The tissue sample was blocked by 0.3%
H2O2 and blocked by 10% bovine serum albumin
(BSA) for 25 min. Then, the tissue sample was incubated overnight
with anti-ribosomal protein S21 (RPS21), anti-forkhead box O1
(FOXO1), anti-baculoviral IAP repeat containing 5 (BIRC5), anti-RNA
polymerase II subunit H (POLR2H), anti-ribosomal protein L22 like 1
(RPL22L1) and anti-nucleophosmin 1 (NPM1; 1:100 dilution;
Proteintech, Wuhan, China) at 4°C and incubated with secondary
antibody (1:1,000 dilution; Beyotime Institute of Biotechnology,
Shanghai, China) for 1.5 h at 30°C. At last, 3,3-diaminobenzidine
(DAB) was used for color visualization and hematoxylin was used for
counterstained. In this study, eight pairs of normal and cancerous
tissues were stained with each antibody. To evaluate the gene
expression, a previously described scoring system was utilized
(13). Briefly, the scores of two
parameters were multiplied by the staining intensity (range, 0–3)
and the percentage of positive cells [range, 0–4 (0, 0–10%; 1,
11–25%; 2, 26–50%; 3, 51–75%; and 4, 76–100%)]. The tissue sample
with scores of 8 or higher was classified as positive staining. In
addition, an independent sample t-test was applied to the results
of staining.
Results
Identification of DEGs
A total of 13 PCa samples and 8 normal samples were
analyzed using Morpheus online tools (https://software.broadinstitute.org/morpheus/). A
total of 2,000 DEGs were identified in PCa compared to normal
group, including 1000 upregulated and 1,000 downregulated genes
respectively. The heat map of DEGs expression (including top 40
upregulated and downregulated genes) was shown in Fig. 1.
GO analysis of DEGs
All DEGs were uploaded to DAVID database (https://david.ncifcrf.gov/), and then GO analysis was
conducted. The results showed that upregulated DEGs were enriched
in biological processes, including cellular response to growth
factor stimulus, phosphate metabolic process, cell development,
cell cycle and cell division, while downregulated DEGs were
enriched in biological processes, including signal transduction,
cell communication, response to stimulus, cell projection
organization and cell development (Table
I). For cell component, upregulated DEGs were enriched in
contractile fiber, actin cytoskeleton, anchoring junction, adherens
junction and focal adhesion, while downregulated DEGs were enriched
in membrane raft, actin cytoskeleton, adherens junction, anchoring
junction and membrane-bounded vesicle (Table I). Furthermore, for molecular
function, upregulated DEGs were enriched in enzyme binding,
cytoskeletal protein binding, enzyme regulator activity,
macromolecular complex binding and protein kinase binding, while
downregulated DEGs were enriched in RNA binding, cytoskeletal
protein binding, enzyme regulator activity, transcription factor
binding and calcium ion binding (Table
I).
 | Table I.GO annotation of DEGs in PCa. |
Table I.
GO annotation of DEGs in PCa.
| A, Upregulated
genes |
|---|
|
|---|
| Category | Term/gene
function | Count | P-value |
|---|
| GOTERM_BP_FAT | Cellular response
to growth factor stimulus | 27 | 4.7×10-6 |
| GOTERM_BP_FAT | Regulation of
phosphate metabolic process | 25 | 6.2×10-6 |
| GOTERM_BP_FAT | Cell
development | 54 | 6.3×10-5 |
| GOTERM_BP_FAT | Regulation of cell
cycle | 28 | 3.1×10-3 |
| GOTERM_BP_FAT | Cell division | 19 | 3.7×10-3 |
| GOTERM_CC_FAT | Contractile
fiber | 20 | 5.4×10-9 |
| GOTERM_CC_FAT | Actin
cytoskeleton | 25 | 9.6×10-7 |
| GOTERM_CC_FAT | Anchoring
junction | 31 | 2.5×10-6 |
| GOTERM_CC_FAT | Adherens
junction | 30 | 4.6×10-6 |
| GOTERM_CC_FAT | Focal adhesion | 21 | 9.4×10-6 |
| GOTERM_MF_FAT | Enzyme binding | 53 | 6.1×10-6 |
| GOTERM_MF_FAT | Cytoskeletal
protein binding | 31 | 9.0×10-3 |
| GOTERM_MF_FAT | Enzyme regulator
activity | 30 | 9.5×10-4 |
| GOTERM_MF_FAT | Macromolecular
complex binding | 34 | 3.9×10-3 |
| GOTERM_MF_FAT | Protein kinase
binding | 18 | 4.4×10-3 |
|
| B, Downregulated
genes |
|
|
Category | Term/gene
function | Count | P-value |
|
| GOTERM_BP_FAT | Regulation of
signal transduction | 25 | 9.2×10-6 |
| GOTERM_BP_FAT | Regulation of cell
communication | 25 | 3.9×10-5 |
| GOTERM_BP_FAT | Regulation of
response to stimulus | 27 | 4.6×10-5 |
| GOTERM_BP_FAT | Cell projection
organization | 26 | 5.4×10-5 |
| GOTERM_BP_FAT | Cell
development | 33 | 1.0×10-4 |
| GOTERM_CC_FAT | Membrane raft | 11 | 2.6×10-4 |
| GOTERM_CC_FAT | Actin
cytoskeleton | 12 | 2.6×10-3 |
| GOTERM_CC_FAT | Adherens
junction | 14 | 7.5×10-3 |
| GOTERM_CC_FAT | Anchoring
junction | 14 | 9.1×10-3 |
| GOTERM_CC_FAT | Membrane-bounded
vesicle | 44 | 1.2×10-3 |
| GOTERM_MF_FAT | RNA binding | 43 | 9.3×10-6 |
| GOTERM_MF_FAT | Cytoskeletal
protein binding | 17 | 1.1×10-4 |
| GOTERM_MF_FAT | Enzyme regulator
activity | 18 | 2.3×10-3 |
| GOTERM_MF_FAT | Transcription
factor binding | 11 | 7.7×10-3 |
| GOTERM_MF_FAT | Calcium ion
binding | 12 | 2.9×10-3 |
KEGG pathway analysis of DEGs
The KEGG pathway analysis results showed that
upregulated DEGs were enriched in proteoglycans in cancer,
endocytosis, focal adhesion, hippo signaling pathway and cGMP-PKG
signaling pathway, whereas downregulated DEGs were enriched in
proteoglycans in cancer, endocytosis, hippo signaling pathway,
thyroid hormone signaling pathway and sulfur relay system (Table II).
| Table II.KEGG pathway analysis of DEGs in
PCa. |
Table II.
KEGG pathway analysis of DEGs in
PCa.
| A, Upregulated
genes |
|---|
|
|---|
| KEGG terms | Count | P-value | Genes |
|---|
| Proteoglycans in
cancer | 13 | 5.0×10-5 | IQGAP1, ROCK2,
ARHGEF12, TIMP3, CAV1, CAV2, FZD1, HOXD10, PAK1, PPP1R12A,
PPP1R12B, SDC4, TGFB2 |
| Endocytosis | 12 | 1.9×10-4 | SH3GLB1, VPS37A,
ARRB1, CAV1, CAV2, CHMP1B, CHMP7, SNX2, SPG20, TGFB2, TGFB3,
VPS36 |
| Focal adhesion | 10 | 4.2×10-3 | ROCK2, CAV1, CAV2,
COL4A6, CCND2, MYLK, PAK1, PPP1R12A, PPP1R12B, VCL |
| Hippo signaling
pathway | 8 | 8.3×10-3 | MOB1A, WWTR1, YAP1,
CCND2, FZD1, SNAI2, TGFB2, TGFB3 |
| cGMP-PKG signaling
pathway | 8 | 1.4×10-3 | GNAI2, ROCK2,
CALM1, CALM3, MEF2A, MYLK, NPR2, PPP1R12A |
|
| B, Downregulated
genes |
|
| KEGG
terms | Count | P-value | Genes |
|
| Proteoglycans in
cancer | 8 | 1.6×10-3 | IQGAP1, ARHGEF12,
TIMP3, CAV1, CAV2, FZD1, HOXD10, SDC4 |
| Endocytosis | 6 | 7.2×10-3 | SH3GLB1, VPS37A,
ARRB1, CAV1, CAV2, SPG20 |
| Hippo signaling
pathway | 5 | 4.2×10-2 | MOB1A, WWTR1, YAP1,
FZD1, SNAI2 |
| Thyroid hormone
signaling pathway | 4 | 7.6×10-2 | SIN3A, FOXO1, RXRA,
SLC16A2 |
| Sulfur relay
system | 2 | 8.4×10-2 | MOCS1, MOCS2 |
Module screening from the PPI
network
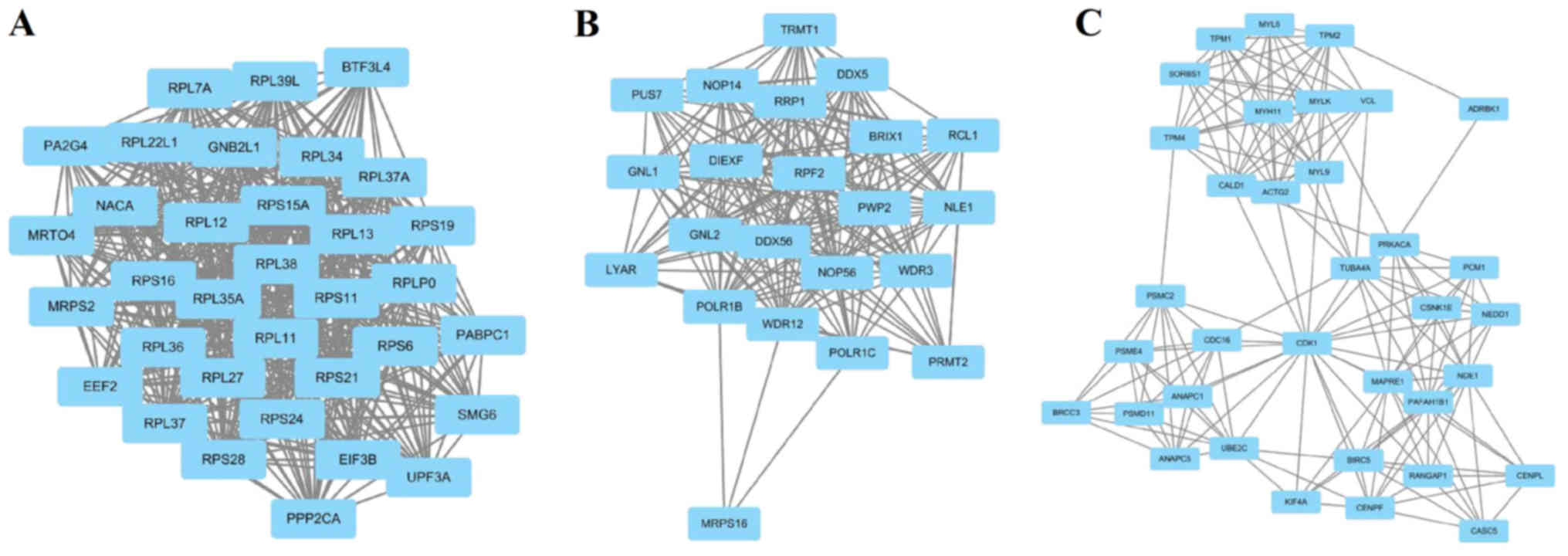
The top 6 hub nodes with high degrees were screened
by the STRING database. These hub genes included RPS21, FOXO1,
BIRC5, POLR2H, RPL22L1 and NPM1. In addition, total nodes were
analyzed by plug-ins MCODE, and the top three significant modules
were selected (Fig. 2). The results
showed that a total of 91 genes of functional annotation were
involved in the modules that associated with cell cycle, oocyte
meiosis and ribosome biogenesis in eukaryotes (data not shown).
IHC validation of key genes in PCa
samples
To verify the bioinformatics analysis data, the
expression of key genes including RPS21, FOXO1, BIRC5, POLR2H,
RPL22L1 and NPM1 were examined by IHC in PCa samples and normal
samples. As shown in Fig. 3, compared
with the normal tissues, the expression of RPS21, BIRC5, POLR2H,
RPL22L1 and NPM1 were significantly upregulated in the cancer cells
from tumor tissues, while the expression of FOXO1 was significantly
downregulated in the cancer cells from tumor tissues (P<0.05).
The IHC results were matched with the bioinformatics analysis.
 | Figure 3.The expression of DEGs in PCa
compared to normal tissues. (A1) RPS21 of PCa; (A2) RPS21 of
control; (B1) FOXO1 of PCa; (B2) FOXO1 of control; (C1) BIRC5 of
PCa; (C2) BIRC5 of control; (D1) POLR2H of PCa; (D2) POLR2H of
control; (E1) RPL22L1 of PCa; (E2) RPL22L1 of control; (F1) NPM1 of
PCa; (F2) NPM1 of control. IHC, magnification, ×400. Scale bar, 50
µm. Control: Normal tissues. DEGs, differentially expressed genes;
PCa, prostate cancer; IHC, immunohistochemistry; FOXO1, forkhead
box O1; BIRC5, baculoviral IAP repeat containing 5; RPS21,
ribosomal protein S21; POLR2H, RNA polymerase II subunit H;
RPL22L1, ribosomal protein L22 like 1; NPM1, nucleophosmin 1. |
Discussion
In the present study, we uploaded GSE21815 and
identify 2,000 DEGs (upregulated and downregulated) between PCa and
normal tissues by bioinformatics analysis. Go analysis and KEGG
pathway analysis showed that the DEGs were mainly involved in cell
cycle, cell division, cell development and cell junction. The
results of PPI analysis showed that some key genes might play an
important role in the occurrence, progression and metastasis of PCa
that could provide the potential biomarker for prognosis, diagnosis
and drug targets.
In this study, the gene expression profile of
GSE55945 was downloaded from GEO which including 18 PCa samples and
8 normal prostate samples. The results showed that a total of 2,000
DEGs were identified in PCa compared to normal group, including
1,000 upregulated and 1,000 downregulated genes respectively.
Previous studies have demonstrated that co-expression genes were
frequently involved in similar biological function and signal
pathway (2,9). Therefore, GO analysis and KEGG pathway
analysis was further performed.
The GO analysis showed that the DEGs were mainly
participated in cell cycle, cell division, cell development and
cell junction (Table I). This result
was consistent with other studies, and the dysregulation of cell
function lead to the occurrence, progression and metastasis of PCa
(2,8).
The KEGG pathway analysis showed that the DEGs were mainly enriched
in proteoglycans in cancer, endocytosis, focal adhesion and hippo
signaling pathway (Table II).
Previous study demonstrated that dysregulation of the hippo pathway
exerts a significant impact on cancer development. For example,
activated hippo signaling pathway was observed in many types of
cancers, including colon, liver, breast, lung and ovary (14). Recent study implied that proteoglycans
exert diverse functions in the occurrence of cancer. For instance,
proteoglycans contributed to the formation of provisional matrix
for tumor growth affecting cell-cell and cell-matrix interactions
and signal transduction of tumor cells. Proteoglycans also
regulated the phenotype of tumor cells and tumor stroma
angiogenesis (15). In addition,
other studies showed that endocytosis and focal adhesion were
closely related to tumorigenesis (16,17).
Bibens-Laulan et al demonstrated that the high expression of
galectin-7 in ovarian and breast cancer cells was due to the
endocytosis (16). Kanteti et
al indicated that focal adhesion kinase play an important role
in tumor cell phenotype such as survival, proliferation, migration
and invasion (17).
Finally, PPI analysis was performed and the key
genes were identified (Fig. 2).
Subsequently, the result was confirmed by IHC. As shown in Fig. 3, the expression of RPS21, BIRC5,
POLR2H, RPL22L1 and NPM1 were significantly upregulated in PCa,
while the expression of FOXO1 was significantly downregulated in
PCa. The IHC results were matched with the bioinformatics
analysis.
RPS21 was the first identified key gene, belonging
to ribosomal proteins (RPS) family. RPS are the pivotal components
of ribosome, which associated with proliferation, differentiation,
DNA repair and apoptosis of cell (18). Arthurs et al reported that
RPS21 was upregulated in PCa and might serve to be a possible
biomarker (19), which was correspond
to our study. To our knowledge, this was the only study of RPS21 in
cancer up to now. Moreover, Huang et al reported that RPS27L
may be a useful index for predicting prognoses in colorectal cancer
(20). Li et al also reported
that knockdown of RPSL26 or RPSL29 significantly inhibits cell
proliferation in pancreatic cancer (21). The second key gene was FOXO1,
belonging to the FOXOs family, which involved in cell
proliferation, differentiation and apoptosis by the regulation of
multiple genes (22). Consistent with
our study, previous studies demonstrated the expression of FOXO1
was downregulated in PCa (23,24).
Moreover, other studies further proposed that FOXO1 was a key tumor
suppressor in cancer, including PCa (22,25). The
third identified key gene was BIRC5, which play an important role
in the occurrence and progression of cancer (26). Wang et al reported that BIRC5
was involved in the tumorigenesis of colorectal cancer (27). In addition, BIRC5 was reported to be
associated with microtubule-kinetochore attachment, interacting
with cell adhesion (28,29). Therefore, BIRC5 might play a critical
role in metastasis of PCa. The fourth identified key gene was
POLR2H, which was the necessary subunit of RNA polymerase II, which
was essential for transcription of DNA (30). To our knowledge, the related study of
POLR2H was extremely rare. This study indicated that POLR2H was
involved in the occurrence and progression of PCa for the first
time, and the mechanism need to be further explored. The fifth key
gene was RPL22L1, which was identified as a trace component of
ribosome (31,32). Wu et al reported that RPL22L1
could promote ovarian cancer metastasis by inducing
epithelial-to-mesenchymal transition (33). Moreover, recent study also
demonstrated that RPL22L1 could play vital and definite roles in
hematopoietic development (34). The
sixth key gene was NPM1, which play an crucial role in cell growth
and proliferation (35). Leotoing ea
al reported that NPM1 was significantly upregulated in prostate
tumour cells, indicating that NPM1 might be an enhancer in
progression of PCa (36,37). Further study showed that NPM1 was
critical for migration and invasion of PCa, and knockdown of NPM1
resulted in a decrease in the growth of the tumor cell (35).
Module analysis of PPI indicated that the occurrence
of PCa was associated with cell cycle, oocyte meiosis and ribosome
biogenesis. It is well known that cell cycle is involved in the
occurrence, progression and metastasis of cancer (38–40).
Moreover, previous studies demonstrated that ribosome biogenesis
was closely related to tumorigenesis, and suppressing ribosome
biogenesis could inhibit cancer development (41,42).
Therefore, drugs targeted ribosome biogenesis was proposed recently
for cancer therapy, which could repress tumor cell proliferation
without genotoxic activity (43).
In conclusion, our study identified some key genes
and pathways by bioinformatics analysis, which might be involved in
the occurrence of PCa. The molecular mechanism of these key genes
in the occurrence of PCa need to be further studied.
Acknowledgements
Not applicable.
Funding
The present study was supported by Natural Science
Foundation of Shandong Province (grant nos. ZR2013CM032,
ZR2014CL034, ZR2015HM028 and ZR2017MH103), Science and Technology
Development Plan of Shandong Province (grant no. 2015GSF118178) and
Medical and Health Development Plan of Shandong Province (grant no.
2017WS058).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the GSE55945 repository, https://www.ncbi.nlm.nih.gov/geo/.
Authors' contributions
WF and ZFP designed the study. SF and ZL performed
the data analysis and wrote the paper. ZG and ZWP identified the
DEGs. SH and XL carried out the GO analysis, KEGG pathway analysis
and PPI analysis. CZ and WY carried out immunohistochemical
analysis. All the authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The procedures performed in this study involving
human patients were in accordance with the ethics standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. The present study was approved by the Research Ethics
Committee of Weifang Medical University. Written informed consent
was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Djulbegovic M, Beyth RJ, Neuberger MM,
Stoffs TL, Vieweg J, Djulbegovic B and Dahm P: Screening for
prostate cancer: Systematic review and meta-analysis of randomised
controlled trials. BMJ. 341:c45432010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu L, Guo K, Liang Z, Li F and Wang H:
Identification of candidate genes that may contribute to the
metastasis of prostate cancer by bioinformatics analysis. Oncol
Lett. 15:1220–1228. 2018.PubMed/NCBI
|
|
3
|
Abate-Shen C and Shen MM: Molecular
genetics of prostate cancer. Genes Dev. 14:2410–2434. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cancer genome atlas research network: The
molecular taxonomy of primary prostate cancer. Cell. 163:1011–1025.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hafeez BB, Zhong W, Fischer JW, Mustafa A,
Shi X, Meske L, Hong H, Cai W, Havighurst T, Kim K and Verma AK:
Plumbagin, a medicinal plant (Plumbago zeylanica)-derived
1,4-naphthoquinone, inhibits growth and metastasis of human
prostate cancer PC-3M-luciferase cells in an orthotopic xenograft
mouse model. Mol Oncol. 7:428–439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nannini M, Pantaleo MA, Maleddu A, Astolfi
A, Formica S and Biasco G: Gene expression profiling in colorectal
cancer using microarray technologies: Results and perspectives.
Cancer Treat Rev. 35:201–209. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang B, Li C and Zhao J: Identification
of key pathways and genes in colorectal cancer using bioinformatics
analysis. Med Oncol. 33:1112016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arredouani MS, Lu B, Bhasin M, Eljanne M,
Yue W, Mosquera JM, Bubley GJ, Li V, Rubin MA, Libermann TA and
Sanda MG: Identification of the transcription factor single-minded
homologue 2 as a potential biomarker and immunotherapy target in
prostate cancer. Clin Cancer Res. 15:5794–5802. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marín-Aguilera M, Codony-Servat J, Kalko
SG, Fernández PL, Bermudo R, Buxo E, Ribal MJ, Gascón P and Mellado
B: Identification of docetaxel resistance genes in
castration-resistant prostate cancer. Mol Cancer Ther. 11:329–339.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lascorz J, Hemminki K and Försti A:
Systematic enrichment analysis of gene expression profiling studies
identifies consensus pathways implicated in colorectal cancer
development. J Carcinog. 10:72011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mi B, Liu G, Zhou W, Lv H, Liu Y and Liu
J: Identification of genes and pathways in the synovia of women
with osteoarthritis by bioinformatics analysis. Mol Med Rep.
17:4467–4473. 2018.PubMed/NCBI
|
|
12
|
Feng W, Zhou D, Meng W, Li G, Zhuang P,
Pan Z, Wang G and Cheng Z: Growth retardation induced by avian
leukosis virus subgroup J associated with down-regulated
Wnt/β-catenin pathway. Microb Pathog. 104:48–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Zhang J, Zhan X, Lin T, Yang M, Hu
J, Han B and Hu S: SOX4 is associated with poor prognosis in
cholangiocarcinoma. Biochem Biophys Res Commun. 452:614–621. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu FX, Zhao B and Guan KL: Hippo pathway
in organ size control, tissue homeostasis, and cancer. Cell.
163:811–828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Theocharis AD and Karamanos NK:
Proteoglycans remodeling in cancer: Underlying molecular
mechanisms. Matrix Biol pii: S0945-053X. 30313-X:2017.
|
|
16
|
Bibens-Laulan N and St-Pierre Y:
Intracellular galectin-7 expression in cancer cells results from an
autocrine transcriptional mechanism and endocytosis of
extracellular galectin-7. PLoS One. 12:e01871942017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanteti R, Mirzapoiazova T, Riehm JJ,
Dhanasingh I, Mambetsariev B, Wang J, Kulkarni P, Kaushik G,
Seshacharyulu P, Ponnusamy MP, et al: Focal adhesion kinase a
potential therapeutic target for pancreatic cancer and malignant
pleural mesothelioma. Cancer Biol Ther. 19:316–327. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang W, Nag S, Zhang X, Wang MH, Wang H,
Zhou J and Zhang R: Ribosomal proteins and human diseases:
Pathogenesis, molecular mechanisms, and therapeutic implications.
Med Res Rev. 35:225–285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arthurs C, Murtaza BN, Thomson C, Dickens
K, Henrique R, Patel HRH, Beltran M, Millar M, Thrasivoulou C and
Ahmed A: Expression of ribosomal proteins in normal and cancerous
human prostate tissue. PLoS One. 12:e01860472017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang CJ, Yang SH, Lee CL, Cheng YC, Tai
SY and Chien CC: Ribosomal protein S27-like in colorectal cancer: A
candidate for predicting prognoses. PLoS One. 8:e670432013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li C, Ge M, Yin Y, Luo M and Chen D:
Silencing expression of ribosomal protein L26 and L29 by RNA
interfering inhibits proliferation of human pancreatic cancer
PANC-1 cells. Mol Cell Biochem. 370:127–139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Duan X, Kong Z, Liu Y, Zeng Z, Li S, Wu W,
Ji W, Yang B, Zhao Z and Zeng G: β-arrestin2 contributes to cell
viability and proliferation via the down-regulation of FOXO1 in
castration-resistant prostate cancer. J Cell Physiol.
230:2371–2381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong XY, Chen C, Sun X, Guo P, Vessella
RL, Wang RX, Chung LW, Zhou W and Dong JT: FOXO1A is a candidate
for the 13q14 tumor suppressor gene inhibiting androgen receptor
signaling in prostate cancer. Cancer Res. 66:6998–7006. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu JJ, Wu YX, Zhao FJ and Xia SJ: miR-96
promotes cell proliferation and clonogenicity by down-regulating of
FOXO1 in prostate cancer cells. Med Oncol. 31:9102014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu Z and Tindall DJ: FOXOs, cancer and
regulation of apoptosis. Oncogene. 27:2312–2319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Altieri DC: Survivin, cancer networks and
pathway-directed drug discovery. Nat Rev Cancer. 8:61–70. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang H, Zhang X, Wang L, Zheng G, Du L,
Yang Y, Dong Z, Liu Y, Qu A and Wang C: Investigation of cell free
BIRC5 mRNA as a serum diagnostic and prognostic biomarker for
colorectal cancer. J Surg Oncol. 109:574–579. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vitale I, Galluzzi L, Senovilla L, Criollo
A, Jemaà M, Castedo M and Kroemer G: Illicit survival of cancer
cells during polyploidization and depolyploidization. Cell Death
Differ. 18:1403–1413. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kwon M, Godinho SA, Chandhok NS, Ganem NJ,
Azioune A, Thery M and Pellman D: Mechanisms to suppress multipolar
divisions in cancer cells with extra centrosomes. Genes Dev.
22:2189–2203. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Du YJ, Hou YL and Hou WR: Molecular
characterization of a gene POLR2H encoded an essential subunit for
RNA polymerase II from the Giant Panda (Ailuropoda Melanoleuca).
Mol Biol Rep. 40:1495–1498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sugihara Y, Honda H, Iida T, Morinaga T,
Hino S, Okajima T, Matsuda T and Nadano D: Proteomic analysis of
rodent ribosomes revealed heterogeneity including ribosomal
proteins L10-like, L22-like 1, and L39-like. J Proteome Res.
9:1351–1366. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
O'Leary MN, Schreiber KH, Zhang Y, Duc AC,
Rao S, Hale JS, Academia EC, Shah SR, Morton JF, Holstein CA, et
al: The ribosomal protein Rpl22 controls ribosome composition by
directly repressing expression of its own paralog, Rpl22l1. PLoS
Genet. 9:e10037082013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu N, Wei J, Wang Y, Yan J, Qin Y, Tong D,
Pang B, Sun D, Sun H, Yu Y, et al: Ribosomal L22-like1 (RPL22L1)
promotes ovarian cancer metastasis by inducing
epithelial-to-mesenchymal transition. PLoS One. 10:e01436592015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Duc AC, Rao S, Sun XL, Bilbee AN,
Rhodes M, Li Q, Kappes DJ, Rhodes J and Wiest DL: Control of
hematopoietic stem cell emergence by antagonistic functions of
ribosomal protein paralogs. Dev Cell. 24:411–425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Loubeau G, Boudra R, Maquaire S,
Lours-Calet C, Beaudoin C, Verrelle P and Morel L: NPM1 silencing
reduces tumour growth and MAPK signalling in prostate cancer cells.
PLoS One. 9:e962932014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Léotoing L, Meunier L, Manin M, Mauduit C,
Decaussin M, Verrijdt G, Claessens F, Benahmed M, Veyssière G,
Morel L and Beaudoin C: Influence of nucleophosmin/B23 on DNA
binding and transcriptional activity of the androgen receptor in
prostate cancer cell. Oncogene. 27:2858–2867. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dai L, Li J, Xing M, Sanchez TW, Casiano
CA and Zhang JY: Using serological proteome analysis to identify
serum anti-nucleophosmin 1 autoantibody as a potential biomarker in
european-american and african-american patients with prostate
cancer. Prostate. 76:1375–1386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yun SJ, Moon SK and Kim WJ:
Investigational cell cycle inhibitors in clinical trials for
bladder cancer. Expert Opin Investig Drugs. 22:369–377. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao Z, Liu J, Wang C, Wang Y, Jiang Y and
Guo M: MicroRNA-25 regulates small cell lung cancer cell
development and cell cycle through cyclin E2. Int J Clin Exp
Pathol. 7:7726–7734. 2014.PubMed/NCBI
|
|
40
|
Soták M, Sumová A and Pácha J: Cross-talk
between the circadian clock and the cell cycle in cancer. Ann Med.
46:221–232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gentilella A, Kozma SC and Thomas G: A
liaison between mTOR signaling, ribosome biogenesis and cancer.
Biochim Biophys Acta. 1849:812–820. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang C, Yin C, Wang L, Zhang S, Qian Y,
Ma J, Zhang Z, Xu Y and Liu S: HSPC111 governs breast cancer growth
by regulating ribosomal biogenesis. Mol Cancer Res. 12:583–594.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brighenti E, Treré D and Derenzini M:
Targeted cancer therapy with ribosome biogenesis inhibitors: A real
possibility? Oncotarget. 6:38617–38627. 2015. View Article : Google Scholar : PubMed/NCBI
|