Introduction
FK506-binding proteins (FKBPs) belong to the family
of immunophilins, which may bind to immunosuppressant drugs
(1). It contains a 3-unit
tetratricopeptide repeat structural motif connected to a domain
that mediates enzymatic function and peptidyl prolyl isomerase
activity. FKBP51 interacts with glucocorticoid, androgen, estrogen,
progestin and mineralocorticoid receptors (2–7) through a
number of components of the molecular chaperone machinery,
including heat shock protein 90. FKBP51 has been identified to be
upregulated or downregulated in multiple types of cancer and to be
associated with cell motility and invasion, for instance, Leach
et al (8) identified that
FKBP51 is increased in prostate cancer cells compared with normal
prostate epithelial cells and improves the ability of stromal
androgen receptor to predict prostate cancer-specific mortality.
Hou et al (9) reported that
low levels of FKBP51 promote pancreatic tumor growth. FKBP51 may
promote cancer, including melanoma, through the nuclear factor
(NF)-κB pathway (10,11) and suppress cancer through the
phosphatidylinositol-4,5-bisphosphate 3 kinase/protein kinase B
(AKT) signaling pathway (12,13). It has been demonstrated that the
activation of NF-κB contributes to growth (14) and aggressiveness of papillary thyroid
carcinoma, and lymph node metastases in papillary thyroid carcinoma
(PTC) are significantly correlated with NF-κB levels (15). Recently, studies have demonstrated
that FKBP51 serves an important role in epithelial-mesenchymal
transition (EMT) (16–18). EMT refers to the biological process by
which epithelial cells are transformed into mesenchymal phenotype
cells by certain procedures. In this procedure, expression of
epithelial cell characteristic proteins, including adhesion
molecule E-cadherin, decreased and cytokeratin cytoskeleton changed
into vimentin skeleton (19). In
colorectal carcinoma, Rotoli D was determined to have increased
microvessel density and tumor-associated macrophages in connective
tissue surrounding FKBP51 positive lesions (16).
The morbidity rate of PTC is increasing worldwide
(20,21). Surgery is the primary treatment for
PTC. The prevalence of PTC is nearly 3 times higher in females than
in males (22). This sex difference
suggests that growth and progression of PTC may be influenced by
female sex hormones, and it has been demonstrated that estrogen and
progesterone receptors are more strongly expressed in thyroid
hyperplasia diseases than in normal thyroid tissue (23).
As FKBP51 regulates the activity of sex-hormone
receptors and correlates with a number of types of cancer, and PTC
may be a sex hormone-associated cancer, in the present study, the
function of FKBP51 in PTC was investigated.
Materials and methods
Materials
The human PTC cell lines K1 and TPC-1 were purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). RPMI-1640, Dulbecco's modified Eagle's medium and fetal
bovine serum (FBS) were purchased from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Anti-FKBP51 (ab46002) used in
immunohistochemistry (IHC), anti-IκBα (ab32518), anti-N-cadherin
(ab76011), anti-Vimentin (ab92547), anti-β-catenin (ab32572) and
anti-MMP9 (ab76003) were purchased from Abcam (Cambridge, UK).
Anti-p65 (sc-8008), anti-FKBP51 (sc-13983), anti-GAPDH (sc-47724),
anti-α-tubulin (sc-53646), secondary antibody anti-mouse
IgG-horseradish peroxidase (HRP; sc-516102) and anti-rabbit IgG-HRP
(sc-2372) used in western blotting were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Anti-TGF-β1 (3709) and
anti-histone 3 (12164) was purchased from Cell Signaling Technology
(Danvers, MA, USA). Ammonium pyrrolidine dithiocarbamate (PDTC;
T3147) was purchased from Target Molecule Corp. (Wellesley Hills,
MA, USA).
IHC
IHC was used to detect expression of FKBP51. Thyroid
tissue microarrays including 31 PTC tissues and 41 adjacent normal
tissues were purchased from Shanxi Alena Biotechnology (Shanxi,
China). Following staining with the Elivision™ plus kit (cat. no.
9902; Maxim Biotech, Inc., Rockville, MD, USA), according to the
manufacturer's protocols, the specimens were incubated overnight
with primary antibody (anti-FKBP51; 1:50 dilution) at 4°C, and
washed with 0.1% Tween-20 PBS for 5 min 3 times. Subsequently,
polymer enhancer (Elivision™ plus kit), which assists horse radish
peroxidase-anti-rabbit polymer (Elivision™ plus kit) to combine
with primary antibodies, was added and incubated for 20 min at room
temperature, and then wash with 0.1% Tween-20 PBS for 5 min 3
times. Following this, the horse radish peroxidase-labeled
anti-rabbit polymer (Elivision™ plus kit) was added and incubated
at room temperature for 30 min, and then washed with 0.1% Tween-20
PBS for 5 min 3 times. Additionally, the specimens were stained
with diaminobenzidine solution (DAB-0031/1031; Maxim Biotech, Inc.)
for 5 min at room temperature, and distilled water was used to
rinse the specimens and stop the coloration.
The expression of FKBP51 was scored by two
professional pathologists as follows: 1+, 1–25% of positive tumor
cells; 2+, 25–50%; 3+, 50–74%; and 4+, ≥75%. 0, negative staining;
1+, light brown; 2+, brown; 3+, dark brown. The total score was
between 0–12 and was calculated as the positive percentage score
multiplied by the intensity score. FKBP51 expression levels were
ranked into four grades according to the total score: 0, -; 1–4, +;
5–8, ++; 9–12, +++.
The clinical stage of papillary thyroid carcinoma
was guided by American Joint Committee on Cancer (24).
Lentivirus and plasmid
transfection
The human FKBP51 gene sequence was retrieved from
the NCBI gene bank (https://www.ncbi.nlm.nih.gov/nuccore/U71321.1) and
restriction enzymes were used to cut appropriate sequences as the
target gene and amplified through polymerase chain reaction [The
target gene from chemical synthesis was denatured into a single
strand at a high temperature ~95°C, and the single nucleotide was
combined into the target gene from the 3′ end of the primer
(FKBP51: Sense, 5′-AAAAGGCCAAGGAGCACAAC-3′ and antisense
5′-TTGAGGAGGGGCCGA194GTTC-3′) to synthesize a new complementary
strand of DNA with a heat-resistant DNA polymerase (Takara Bio,
Inc., Otsu, Japan). The full thermocycling conditions can be
simplified into the following steps: Pre-denaturation (95°C for 30
sec), reaction cycles (95°C for 5 sec and 60°C for 30 sec), melting
(95°C for 5 sec and 60°C for 1 min) and cooling (50°C for 30 sec).
Subsequently, the FKBP51 cDNA was cloned into the GV303 lentiviral
expression vector. All this process was conducted by Shanghai
GeneChem (Shanghai GeneChem, Co., Ltd., Shanghai, China). Plasmids
encoding human FKBP51 shRNA (SC-35380-SH) and scramble shRNA were
purchased from Santa Cruz Biotechnology, Inc. K1 cells were seeded
in a 96-well plate (5×103 cells/well) overnight. Then,
the cells were infected with 1×106 units of the
recombinant lentiviral vector carrying wild-type human FKBP51
(concentration, 2×108 U/ml) or randomized flanking
sequences (concentration, 1×109 U/ml) as a control under
the condition of the Polybrene (5 µg/ml) as the transfection
reagent (Shanghai GeneChem, Co., Ltd.). After 8–12 h, the virus
mixture was exchanged for fresh RPMI-1640 complete medium. Green
fluorescent protein (GFP)-positive infected cells were detected by
fluorescence microscopy (×10) at 72 h after transfection. TPC-1
cells were seeded in a 6-well plate (2×105 cells/well).
The next day, shR-FKBP51 and shR-vec plasmids were added to the
cells, and 5–7 h later, the mixture was changed into fresh medium
containing 20% FBS. After 48 h, transfected cells were selected
using puromycin (12 µg/ml). Subsequent experiments were performed
following confirmation of the efficiency of infection (~72 h after
infection).
Proliferation assay
FKBP51-overexpressing or knockdown cells and control
cells were digested with trypsin and then seeded into 96-well
plates (5×103 cells/well) and incubated at 37°C under 5%
CO2. Following incubation for 24, 48 and 72 h, 10 µl of
cell counting kit-8 and 90 µl of RPMI-1640 fresh medium were added
into each well. Following incubation at 37°C in an atmosphere
containing 5% CO2 for 1 h, the absorbance was measured
at 450 nm using a SpectraMax M2 instrument. All experiments were
performed in triplicate.
Migration and invasion assays
In total, 2×105 K1 cells or
3×104 TPC-1 cells in serum-free RPMI-1640 medium were
added into Transwell upper chambers with 8-mm pore-size membranes,
and complete medium was added to the lower chambers. To detect cell
invasion ability, membranes were coated with Matrigel (diluted 1:6
in 1× PBS; BD Biosciences, Franklin Lakes, NJ, USA). The plate was
incubated at 37°C in an atmosphere containing 5% CO2 for
24 h. Cells on the upper membrane surface were removed using cotton
swabs and washed with 1× PBS, and the cells attached to the bottom
surface of the membrane were stained using crystal violet at room
temperature for 15 min. Images (×20) were captured using a Nikon
fluorescence microscope (Nikon Corporation, Tokyo, Japan).
Western blotting
Cells were harvested and total protein was extracted
using radioimmunoprecipitation assay buffer (Thermo Fisher
Scientific, Inc.). The protein concentration was determined using a
bicinchoninic acid assay. A total of 30 µg of protein was separated
via 10% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes (EMD Millipore, Billerica, MA, USA). Following blocking
for 1 h with 5% nonfat milk (Yili Dairy Corporation Group, Co.,
Ltd., Hohhot, China) diluted with PBS, the membranes were incubated
with the following primary antibodies at 4°C overnight: Anti-FKBP51
(1:1,000), anti-p65 (1:250), anti-IκBα (1:1,000), anti-TGF-β1
(1:1,000), anti-N-cadherin (1:1,000), Vimentin (1:1,000),
anti-β-catenin (1:1,000) and anti-MMP9 (1:1,000). Following washing
three times with 1X TBS with Tween-20, the membranes were incubated
with horseradish peroxidase-conjugated secondary antibodies
(1:1,000) for 1 h at room temperature and then washed three times
with 1X TBS with Tween-20. The resulting signal was detected using
Scion Image software version 4.03 (Scion Corporation, Frederick,
MD, USA), and the quantity of protein was normalized to that of
GAPDH (1:1,000), histone 3 (1:1,000), or α-tubulin (1:1,000) (for
normalization to total protein, nuclear protein, or cytoplasmic
protein, respectively).
Immunofluorescence microscopy
Cells on a slide were fixed with 3.7% formaldehyde
for 10 min and washed with PBS containing 0.1% Triton X-100 twice.
Tubulin-Tracker Red (Beyotime Institute of Biotechnology, Haimen,
China) was diluted 1:250 in PBS containing 5% bovine serum albumin
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) and 0.1% Triton X-100, and the slides were incubated with
this solution for 30 min at room temperature. The slides were then
washed three times with PBS for 5 min each.
PDTC inhibition test
PDTC powder was dissolved in dimethyl sulfoxide
(DMSO) as a storage solution (20 mM), and the working fluid
concentration was 20 µM. In the Transwell test, 0.2 µl PDTC storage
solution was added into 200 µl cell suspension in the upper
chambers and 0.2 µl DMSO was added in the control group. Migrated
and invaded cells were counted after 24 h. In the western blot
assay, FKBP51-overexpressing K1 cells were seeded into 6-well plate
(1×106 cells/well). The following day, 2 µl PDTC storage
solution was added into 2 ml fresh RPMI-1640 medium in the plate
and 2 µl DMSO was added in the control group. The protein of the
two cell groups were extracted after culturing for 24 h at 37°C in
an atmosphere containing 5% CO2. The migration/invasion
and western blotting assays were similar to that aforementioned,
with the difference being whether PDTC or DMSO was added.
Statistical analysis
All results were analyzed using SPSS software
(version 20.0; IBM Corp., Armonk, NY, USA) and data are presented
as the mean ± standard error of at least three independent
experiments. The differences between two groups were evaluated
using Student's t-test. Differences among multiple groups were
assessed using one-way analysis of variance and the Student's
Newman Keuls-q test was used for post-hoc analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
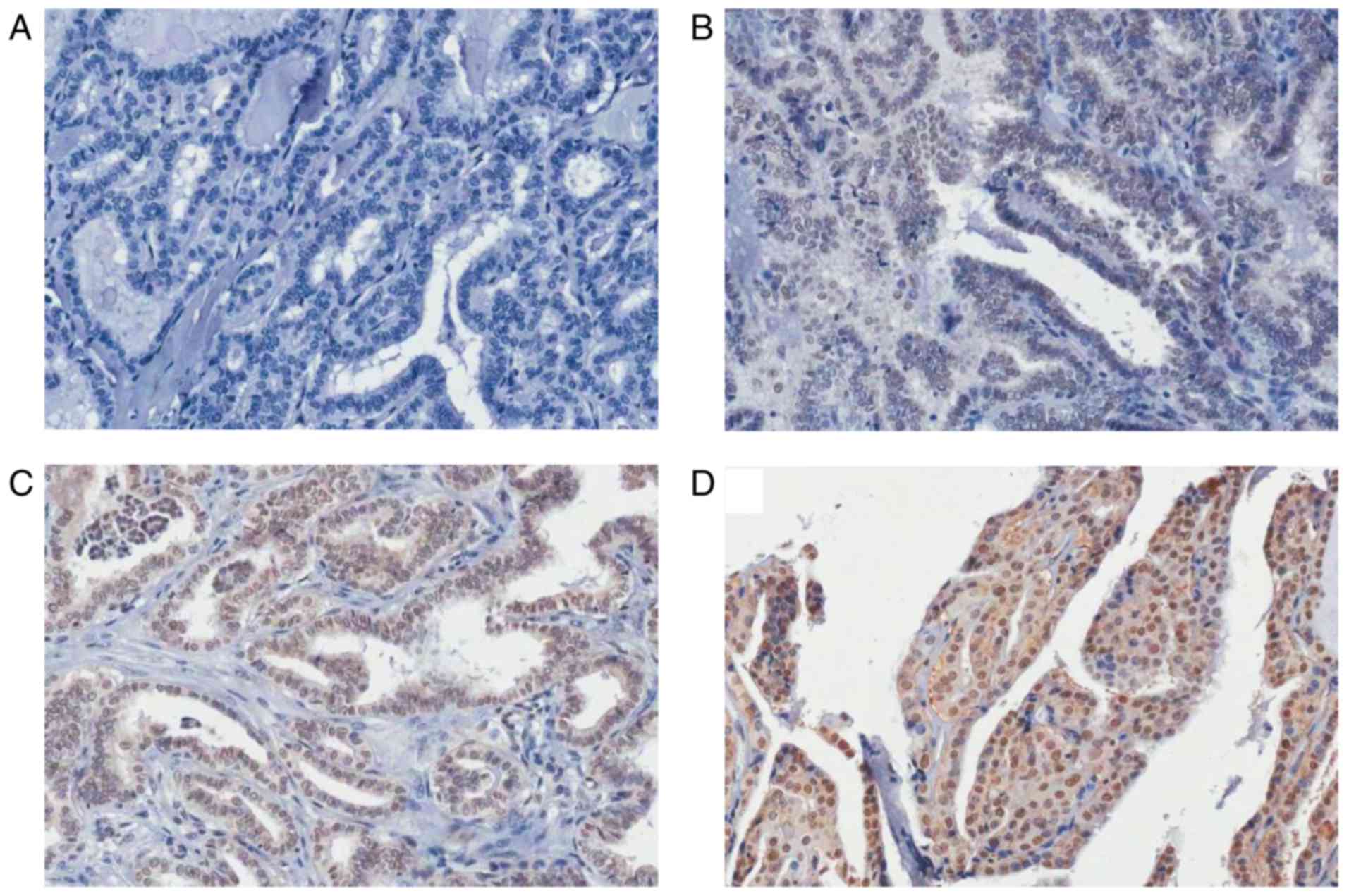
FKBP51 is highly expressed in PTC
IHC analysis revealed higher FKBP51 expression in
PTC than in the adjacent normal tissues. Furthermore, the FKBP51
expression level was associated with clinical tumor, node and
metastasis (TNM) stage, with stronger FKBP51 expression indicating
poorer TNM stage (Fig. 1; Table I).
 | Table I.FKBP51 expression in different
patients and tissues. |
Table I.
FKBP51 expression in different
patients and tissues.
|
| Expression of
FKBP51 |
|
|---|
|
|
|
|
|---|
|
Characteristics | Cases | − | + | ++ | +++ | P-value |
|---|
| Group |
|
|
|
|
| 0.015 |
|
PTC | 31 | 14 | 4 | 9 | 4 |
|
|
Normal | 41 | 23 | 15 | 3 | 0 |
|
| Age |
|
|
|
|
| 0.140 |
|
<45 | 46 | 27 | 11 | 6 | 2 |
|
|
>45 | 26 | 11 | 7 | 5 | 3 |
|
| TNM |
|
|
|
|
| 0.023 |
| I | 19 | 11 | 4 | 3 | 1 |
|
| II | 9 | 2 | 1 | 5 | 1 |
|
|
III | 2 | 1 | 0 | 1 | 0 |
|
| IV | 1 | 0 | 0 | 0 | 1 |
|
FKBP51 promotes migration and invasion
of PTC without affecting cell proliferation
FKBP51-overexpressing and control K1 cells were
successfully constructed, alongside FKBP51-knockdown and control
TPC-1 cells (Fig. 2A). To determine
the effect of FKBP51 on PTC cell migration and invasion, a
transwell assay was performed. Fig.
2B presents that FKBP51 overexpression promoted cell migration
and invasion as compared with the control group in K1 cells. The
counts of migrated cells were 210±61.2 and 102.0±9.1 (P<0.05),
and those of invaded cells were 75.3±11.5 and 48.8±6.2 (P<0.05),
in overexpressing and control cells, respectively. FKBP51 knockdown
inhibited cell migration and invasion in TPC-1 cells as compared
with the control group. The counts of migrated cells were
158.5±11.4 and 50.25±7.4 (P<0.05), and those of invaded cells
were 81.8±6.3 and 29.5±6.0 (P<0.05), respectively. There was no
significant difference in proliferation between
FKBP51-overexpressing and control K1 cells, and FKBP51-knockdown
and control TPC-1 cells (Fig.
2C).
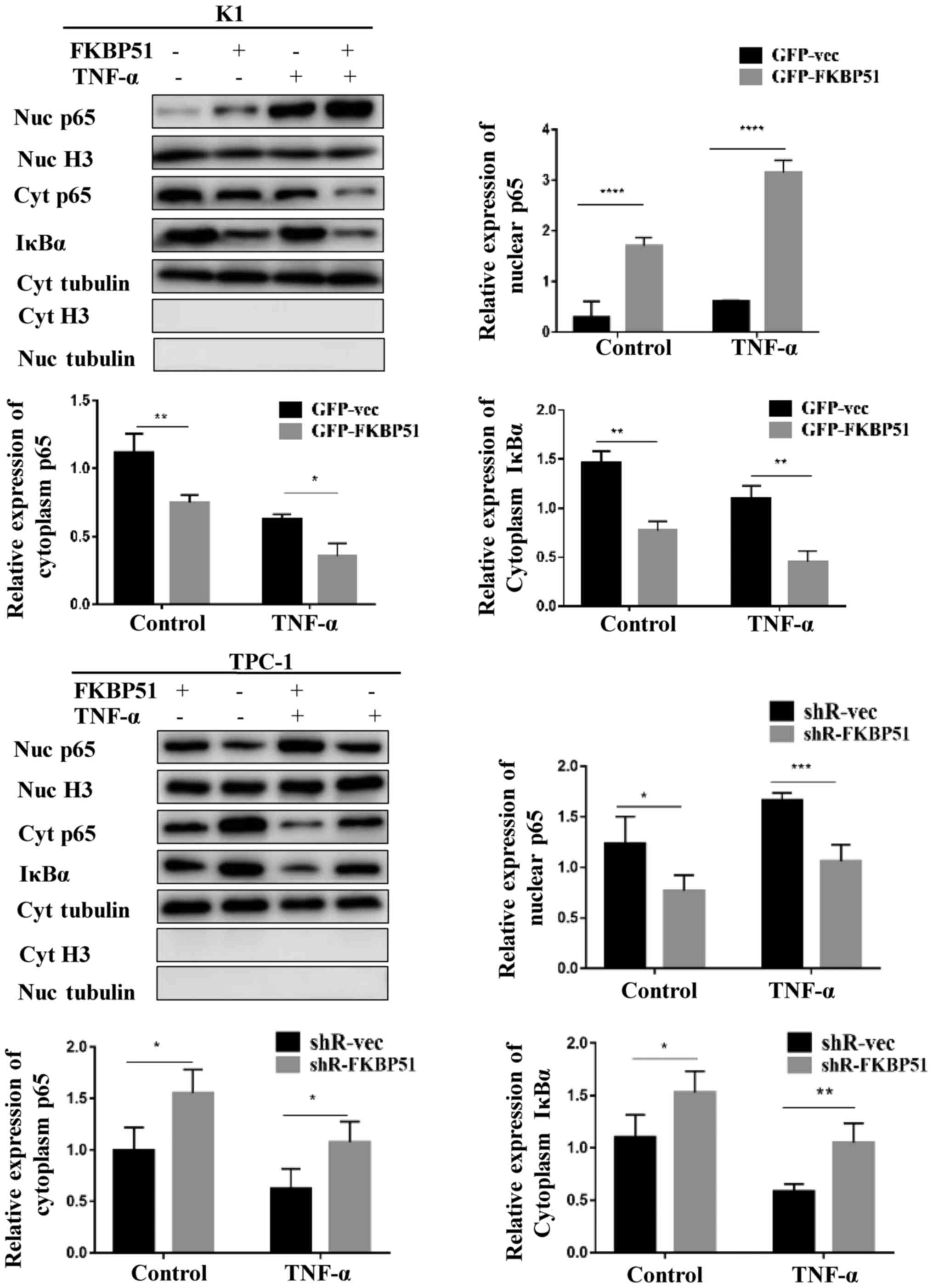
FKBP51 regulates the NF-κB
pathway
The molecular mechanism underlying the promoting
effect of FKBP51 on migration and invasion was investigated. The
results demonstrated that FKBP51 activated the NF-κB pathway. The
activity of the NF-κB pathway was investigated at the basic level
and at the TNF-α-stimulated level. Western blotting results
revealed that both at the basic and the stimulated level,
cytoplasmic IκBα and cytoplasmic P65 were decreased and nuclear P65
was increased in FKBP51-overexpressing K1 cells, while cytoplasmic
IκBα and P65 were increased and nuclear P65 decreased in
FKBP51-knockdown TPC-1 cells as compared with the control groups
(Fig. 3).
Further, the NF-κB pathway inhibitor PDTC (20 µM)
suppressed migration and invasion of GFP-FKBP51 K1 (Fig. 4A). This confirmed that FKBP51
inhibited PTC cell migration and invasion via the NF-κB
pathway.
 | Figure 4.FKBP51 activated EMT through the
NF-κB pathway. (A) GFP-FKBP51 K1 cells were treated with NF-κB
pathway inhibitor PDTC (20 µM) for 24 h and the cells' migration
and invasive capacities were determined using the transwell assay.
(B) EMT-associated proteins TGF-β1, β-catenin, N-cadherin, MMP9 of
K1 cells and TGF-β1, Vimentin, N-cadherin, MMP9 of TPC-1 cells were
detected by western blotting. (C) FKBP51-overexpressing K1 cells
were treated with NF-κB inhibitor PDTC (20 mM) for 24 h and
expression of TGF-β1, β-catenin, N-cadherin and MMP9 was compared
with control group was detected. *P<0.05; **P<0.01;
***P<0.001. FKBP51, FK506 binding protein 51; GFP, green
fluorescent protein; vec, vector; MMP, matrix metalloproteinase;
EMT, epithelial mesenchymal transition. |
FKBP51 activates NF-κB-dependent
EMT
EMT-associated protein expression was altered with
FKBP51 expression level. TGF-β1, β-catenin, N-cadherin, and MMP9
were increased in FKBP51-overexpressing K1 cells, while TGF-β1,
Vimentin, N-cadherin and MMP9 were decreased in FKBP51-knockdown
TPC-1 cells, as presented in Fig. 4B.
Subsequently, it was assessed whether EMT is regulated by the NF-κB
pathway. FKBP51-overexpressing K1 cells were treated with PDTC (20
µM) for 24 h. TGF-β1, N-cadherin, β-catenin and MMP9 were decreased
as compared with control group by this treatment (Fig. 4C).
FKBP51 does not alter the formation of
tubulin
Further, it was assessed whether FKBP51 promotes
migration and invasion of PTC cells through promoting the formation
of tubulin cytoskeleton, using immunofluorescence staining. No
difference in fluorescent signal was observed between
FKBP51-overexpressing and control K1 cells, and FKBP51-knockdown
and control TPC-1 cells (Fig. 5).
Discussion
PTC accounted for 60–70% of thyroid cancer between
1992–2002 in Liaoning, China, mostly in adults aged between 20 and
50 years (25). There are numerous
factors affecting PTC, including environmental factors and genetic
mutations (26). However, gender is
universally recognized as one of the risk factors since the
occurrence is significantly higher in females than males (27). Therefore, most patient samples from
the tissue microarray were female patients. PTC generally has small
size and slow progression, but early metastasis may be identified
(28). The underlying molecular
mechanism and associated molecular markers of metastasis are not
clear. Upregulation and downregulation of FKBP51 is observed in a
number of human tumors. It is notable that FKBP51 is associated
with the development of a variety of hormone-associated tumors, and
previous studies have demonstrated that FKBP51 is overexpressed in
androgen-dependent prostate cancer (8,10), and
decreased in estrogen- and progesterone-associated breast cancer
(29). In the present study, the
expression of FKBP51 in PTC and its clinical significance were
analyzed. It was identified that FKBP51 is expressed in both PTC
and adjacent tissues, however its expression was significantly
higher in cancer than in adjacent normal tissues. Furthermore, the
FKBP51 expression level was associated with clinical TNM stage.
Thus, it is hypothesized that FKBP51 may be associated with the
proliferation and metastasis of thyroid cancer.
Accumulating data have demonstrated that FKBP51
serves important functions in tumor cell growth, apoptosis, and
sensitivity to radiotherapy and chemotherapy (30,31).
Studies have demonstrated that FKBP51 may serve a positive function
in tumor progression by activating the NF-κB (32–34)
pathway, or a negative function by promoting the Akt pathway
through the dephosphorylation of Akt Ser473 to inhibit cell
proliferation (9,35). The mechanism of FKBP51 in promoting
cancer cell migration and invasion is not clear. Romano et
al (17) and D'Angelillo et
al (18) identified that FKBP51
may promote the migration and invasion of melanoma by activating
EMT-associated genes. TGF-β1 as the key inducer of EMT not only
promotes tumor cell metastasis but also mediates liver cirrhosis
(36) and renal tubulointerstitial
fibrosis (37). Further, FKBP51
reportedly not only promotes the activation of EMT genes but also
induces certain melanoma stem cell genes (38). Srivastava et al (39) identified that FKBP51 promotes the
proliferation and migration of melanoma by mediating interleukin
IL-8. Certain researchers have demonstrated that FKBP51 promotes
tubulin cytoskeleton formation through the ras homolog family
member A pathway to promote cell invasion (40). In the present study, K1 and TPC-1 cell
lines were selected, which are commonly used in study [the K1 cell
line is a GLAG-66 derivative (41)]
to overexpress and knockdown FKBP51 respectively. The results
demonstrated that FKBP51 did not affect cell proliferation, but
significantly promoted the migration and invasion of PTC cells. To
clarify the underlying mechanism of the latter observation, NF-κB
pathway- and EMT-associated proteins were evaluated. The results
revealed that both the basic and the TNF-α-simulated activity of
the NF-κB pathway were increased in FKBP51-overexpressing than in
control cells. Similarly, EMT-associated proteins were altered in
FKBP51-overexpressing K1 and FKBP51-knockdown TPC-1 cells.
Subsequently, the present study aimed to evaluate whether there is
an association between EMT and the NF-κB pathway. When the NF-κB
inhibitor PDTC was added to FKBP51-overexpressing K1 cells for 24
h, TGF-β1, N-cadherin, and β-catenin and MMP9 expression were
decreased. These results revealed that FKBP51 promotes migration
and invasion through NF-κB pathway-dependent EMT. This result is
consistent with previous findings, for example Ying et al
(42) demonstrated that induced EMT
was accompanied by nuclear translocation of NF-κB in lung cancer.
Lv et al (43) revealed that
twist1 regulates EMT via the NF-κB pathway in PTC. The present
study also assessed whether FKBP51 is able to promote tubulin
formation by using immunofluorescence, however no evidence of such
an effect was identified. It was also assessed whether FKBP51 can
promote tubulin formation by using immunofluorescence, however,
once again no evidence of such an effect was observed.
To conclude, the present study demonstrates that
FKBP51 promotes migration and invasion of PTC through the NF-κB
pathway and activation of EMT-associated genes, indicating its
diagnostic and therapeutic value.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 31370897), and all
the results come from the joint efforts of all authors.
Availability of data and materials
The datasets generated/analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG, RZ, EE, HY, SL, LS and YZ designed the study.
YG, JD and MW performed the experiments. YG wrote the manuscript
and analyzed the data.
Ethics approval and consent to
participate
All patients provided written informed consent prior
to their inclusion. The present study was approved by the Ethical
Committee of Shandong Provincial Hospital.
Patient consent for publication
All patients provided written informed consent for
the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dornan J, Taylor P and Walkinshaw MD:
Structures of immunophilins and their ligand complexes. Curr Top
Med Chem. 3:13922003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheung-Flynn J, Roberts PJ, Riggs DL and
Smith DF: C-terminal sequences outside the tetratricopeptide repeat
domain of FKBP51 and FKBP52 cause differential binding to Hsp90. J
Biol Chem. 278:17388–17394. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheung J and Smith DF: Molecular chaperone
interactions with steroid receptors: An update. Mol Endocrinol.
14:939–946. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Riggs DL, Roberts PJ, Chirillo SC,
Cheung-Flynn J, Prapapanich V, Ratajczak T, Gaber R, Picard D and
Smith DF: The Hsp90-binding peptidylprolyl isomerase FKBP52
potentiates glucocorticoid signaling in vivo. Embo J. 22:1158–1167.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tai PK, Maeda Y, Nakao K, Wakim NG,
Duhring JL and Faber LE: A 59-kilodalton protein associated with
progestin, estrogen, androgen, and glucocorticoid receptors.
Biochemistry. 25:5269–5275. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gallo LI, Ghini AA, Pilipuk Piwien G and
Galigniana MD: Differential recruitment of tetratricorpeptide
repeat domain immunophilins to the mineralocorticoid receptor
influences both heat-shock protein 90-dependent retrotransport and
hormone-dependent transcriptional activity. Biochemistry.
46:14044–14057. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brucker SY, Eisenbeis S, König J, Lamy M,
Salker MS, Zeng N, Seeger H, Henes M, Schöller D, Schönfisch B, et
al: Decidualization is impaired in endometrial stromal cells from
uterine rudiments in Mayer-Rokitansky-Küster-Hauser syndrome. Cell
Physiol Biochem. 41:1083–1097. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leach DA, Trotta AP, Need EF, Risbridger
GP, Taylor RA and Buchanan G: The prognostic value of stromal
FK506-binding protein 1 and androgen receptor in prostate cancer
outcome. Prostate. 77:185–195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hou J and Wang L: FKBP5 as a selection
biomarker for gemcitabine and Akt inhibitors in treatment of
pancreatic cancer. PLoS One. 7:e362522012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ni L, Yang CS, Gioeli D, Frierson H, Toft
DO and Paschal BM: FKBP51 promotes assembly of the Hsp90 chaperone
complex and regulates androgen receptor signaling in prostate
cancer cells. Mol Cell Biol. 30:1243–1253. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang W, Cazacu S, Xiang C, Zenklusen JC,
Fine HA, Berens M, Armstrong B, Brodie C and Mikkelsen T: FK506
binding protein mediates glioma cell growth and sensitivity to
rapamycin treatment by regulating NF-kappaB signaling pathway.
Neoplasia. 10:235–243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Solassol J, Mange A and Maudelonde T: FKBP
family proteins as promising new biomarkers for cancer. Curr Opin
Pharmacol. 11:320–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Storer CL, Dickey CA, Galigniana MD, Rein
T and Cox MB: FKBP51 and FKBP52 in signaling and disease. Trends
Endocrinol Metab. 22:481–490. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pyo JS, Kang G, Kim DH, Chae SW, Park C,
Kim K, Do Si, Lee HJ, Kim JH and Sohn JH: Activation of nuclear
factor-κB contributes to growth and aggressiveness of papillary
thyroid carcinoma. Pathol Res Pract. 209:228–232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei L, Hui M, Sun D, Li W, Wang D, Zhang G
and Tan J: The relationship between BRAFV600E, NF-κB and TgAb
expression in papillary thyroid carcinoma. Pathol Res Pract.
213:183–188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rotoli D, Morales M, Del Carmen Maeso M,
Del Pino García M, Morales A, Ávila J and Martín-Vasallo P:
Expression and localization of the immunophilin FKBP51 in
colorectal carcinomas and primary metastases, and alterations
following oxaliplatin-based chemotherapy. Oncol Lett. 12:1315–1322.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Romano S, D'Angelillo A, D'Arrigo P,
Staibano S, Greco A, Brunetti A, Scalvenzi M, Bisogni R, Scala I
and Romano MF: FKBP51 increases the tumour-promoter potential of
TGF-beta. Clin Transl Med. 3:12014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
D'Angelillo A, Staibano S, Russo M, Romano
MF and Romano S: Molecular aspects of FKBP51 that enable melanoma
dissemination. Curr Mol Pharmacol. 9:141–147. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song IH, Kim KR, Lim S, Kim SH and Sung
CO: Expression and prognostic significance of
epithelial-mesenchymal transition-related markers and phenotype in
serous ovarian cancer. Pathol Res Pract. 2018. View Article : Google Scholar
|
|
20
|
Ceresini G, Corcione L, Michiara M, Sgargi
P, Teresi G, Gilli A, Usberti E, Silini E and Ceda GP: Thyroid
cancer incidence by histological type and related variants in a
mildly iodine-deficient area of Northern Italy, 1998 to 2009.
Cancer. 118:5473–5480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jonklaas J, Murthy S, Liu D,
Klubo-Gwiezdzinska J, Krishnan J, Burman KD, Boyle L, Carrol N,
Felger E and Loh YP: Novel biomarker SYT12 may contribute to
predicting papillary thyroid cancer outcomes. Future Sci OA.
4:FSO2492017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Derwahl M and Nicula D: Estrogen and its
role in thyroid cancer. Endocr Relat Cancer. 21:T273–T283. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu J, Chen G, Meng XY, Liu ZH and Dong S:
Serum levels of sex hormones and expression of their receptors in
thyroid tissue in female patients with various types of thyroid
neoplasms. Pathol Res Pract. 210:830–835. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Amin MB, Greene FL, Edge SB, Compton CC,
Gershenwald JE, Brookland RK, Meyer L, Gress DM, Byrd DR and
Winchester DP: The Eighth Edition AJCC Cancer Staging Manual:
Continuing to build a bridge from a population-based to a more
“personalized” approach to cancer staging. CA Cancer J Clin.
67:93–99. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Haixia Guan, Zhongyan Shan, Xiaoyi Mi, et
al: 11 years of pathological analysis of thyroid cancer before and
after common salt iodization. J China Med Univer. 35:284–285.
2006.
|
|
26
|
Zhang Q, Liu SZ, Zhang Q, Guan YX, Chen QJ
and Zhu QY: Meta-analyses of association between BRAF(V600E)
mutation and clinicopathological features of papillary thyroid
carcinoma. Cell Physiol Biochem. 38:763–776. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen GG, Vlantis AC, Zeng Q and van
Hasselt CA: Regulation of cell growth by estrogen signaling and
potential targets in thyroid cancer. Curr Cancer Drug Targets.
8:367–377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi MH, Moon JY, Cho SH, Chung BC and Lee
EJ: Metabolic alteration of urinary steroids in pre- and
post-menopausal women, and men with papillary thyroid carcinoma.
BMC Cancer. 11:3422011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Le Bihan S, Marsaud V, Mercier-Bodard C,
Baulieu EE, Mader S, White JH and Renoir JM: Calcium/calmodulin
kinase inhibitors and immunosuppressant macrolides rapamycin and
FK506 inhibit progestin-and glucocorticosteroid receptor-mediated
transcription in human breast cancer T47D cells. Mol Endocrinol.
12:986–1001. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rotoli D, Morales M, Ávila J, Maeso MDC,
García MDP, Mobasheri A and Martín-Vasallo P: Commitment of
scaffold proteins in the onco-biology of human colorectal cancer
and liver metastases after oxaliplatin-based chemotherapy. Int J
Mol Sci. 18:E8912017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo K, Li Y, Yin Y, Li L, Wu C, Chen Y,
Nowsheen S, Hu Q, Zhang L, Lou Z and Yuan J: USP49 negatively
regulates tumorigenesis and chemoresistance through FKBP51-AKT
signaling. EMBO J. 36:1434–1446. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Loercher A, Lee TL, Ricker JL, Howard A,
Geoghegen J, Chen Z, Sunwoo JB, Sitcheran R, Chuang EY, Mitchell
JB, et al: Nuclear factor-kappaB is an important modulator of the
altered gene expression profile and malignant phenotype in squamous
cell carcinoma. Cancer Res. 64:6511–6523. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Romano S, Xiao Y, Nakaya M, D'Angelillo A,
Chang M, Jin J, Hausch F, Masullo M, Feng X, Romano MF and Sun SC:
FKBP51 employs both scaffold and isomerase functions to promote
NF-κB activation in melanoma. Nucleic Acids Res. 43:6983–6993.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li P, Zhang X, Wang L, Du L, Yang Y, Liu
T, Li C and Wang C: lncRNA HOTAIR contributes to 5FU resistance
through suppressing miR-218 and activating NF-κB/TS signaling in
colorectal cancer. Mol Ther Nucleic Acids. 8:356–369. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stechschulte LA, Hinds TD Jr, Ghanem SS,
Shou W, Najjar SM and Sanchez ER: FKBP51 reciprocally regulates GRα
and PPARγ activation via the Akt-p38 pathway. Mol Endocrinol.
28:1254–1264. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim JY, An HJ, Kim WH, Gwon MG, Gu H, Park
YY and Park KK: Anti-fibrotic effects of synthetic
oligodeoxynucleotide for TGF-β1 and smad in an animal model of
liver cirrhosis. Mol Ther Nucleic Acids. 8:250–263. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu J, Zhu Q, Li PL, Wang W, Yi F and Li N:
Stem cell conditioned culture media attenuated albumin-induced
epithelial-mesenchymal transition in renal tubular cells. Cell
Physiol Biochem. 35:1719–1728. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Romano S, Staibano S, Greco A, Brunetti A,
Nappo G, Ilardi G, Martinelli R, Sorrentino A, Di Pace A, Mascolo
M, et al: FK506 binding protein 51 positively regulates melanoma
stemness and metastatic potential. Cell Death Dis. 4:e5782013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Srivastava SK, Bhardwaj A, Arora S, Tyagi
N, Singh AP, Carter JE, Scammell JG, Fodstad Ø and Singh S:
Interleukin-8 is a key mediator of FKBP51-induced melanoma growth,
angiogenesis and metastasis. Br J Cancer. 112:1772–1781. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Takaoka M, Ito S, Miki Y and Nakanishi A:
FKBP51 regulates cell motility and invasion via RhoA signaling.
Cancer Sci. 108:380–389. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ribeiro FR, Meireles AM, Rocha AS and
Teixeira MR: Conventional and molecular cytogenetics of human
non-medullary thyroid carcinoma: Characterization of eight cell
line models and review of the literature on clinical samples. BMC
Cancer. 8:3712008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ying Y, Qingwu L, Mingming X, Zhenju S,
Chaoyang T and Zhengang T: Emodin: One main ingredient of shufeng
jiedu capsule reverses chemoresistance of lung cancer cells through
inhibition of EMT. Cell Physiol Biochem. 42:1063–1072. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lv N, Shan Z, Gao Y, Guan H, Fan C, Wang H
and Teng W: Twist1 regulates the epithelial-mesenchymal transition
via the NF-κB pathway in papillary thyroid carcinoma. Endocrine.
51:469–477. 2016. View Article : Google Scholar : PubMed/NCBI
|