The Wnt/β-catenin signaling pathway serves important
roles in the tumorigenesis of colorectal cancer (CRC). In colon
cancer cell lines, the disruption of the adenomatous polyposis coli
protein, caused by loss of heterozygosity or mutation, leads to
deregulation of the β-catenin protein (1). Subsequently, it is transported into the
nucleus and activates its target genes by recruiting cofactors of
the transcription factor/lymphoid enhancer binding factor family
(2). It is crucial to identify
β-catenin target genes, since they are involved in cellular
processes that contribute to proliferation and migration in
colorectal carcinoma (3–8). Based on a literature review, the Nusse
group (9) revealed 28 β-catenin
target genes in human colon cancer, which are presented on their
website (https://web.stanford.edu/group/nusselab/cgi-bin/wnt/target_genes).
In an experimental approach, RNA interference (RNAi) of β-catenin
was implemented by treating DLD1 and SW480 cells with small
interfering RNA (siRNA) (10). By
comparison with controls, in which samples were treated with mock
siRNA, the identified differentially expressed genes are considered
to be potential β-catenin target genes (10). The study then incorporated results
from another study with a similar design, in which LS174T cells
were treated with short hairpin RNA against β-catenin (11), and identified a total of 335 target
genes. Recently, a multi-omics approach was used by Ewing et
al (12) to decipher the
oncogenic β-catenin network in HCT116 cells by comparing
transcriptome, expression proteome and interactome data of wild
type and β-catenin mutated samples. The results were subsequently
integrated into a functional molecular network. However, target
genes identified by these studies (10–12) lack
functional information. Additionally, expression levels
occasionally fail to reflect molecular dysfunction where
post-translational modifications are involved (13).
Therefore, the present study aimed to identify
β-catenin target genes by leveraging large-scale RNAi and
CRISPR-CRISPR associated protein 9 (Cas9) genetic perturbation
datasets, since they may provide an opportunity to derive gene-gene
functional associations. Initially, in yeast, the gene-gene
associations were revealed by epistatic analysis, where the
phenotypic readout of a genetic perturbation depends on the status
of a second gene (14). Based on
this, genetic interaction networks could be constructed to reveal
functionally associated genes (15).
For human cell lines, the utility of this approach is limited due
to the exponential increase in the combinatorial space (16,17).
However, genome-wide single gene perturbation screens are more
advanced and can be applied in a large collection of cell lines
(18,19). For these screens, highly variable
genetic dependencies for cellular fitness are observed in cancer
cell lines, which may reflect diverse alterations during
tumorigenesis (19). Upon
perturbation of each gene, the genomic state of each cancer cell
exhibits a unique overall fitness response. Notably, Project
Acheilles achieved promising results by systematically elucidating
genetic vulnerabilities across 501 cancer cell lines using RNAi
(18) and in 342 cancer cell lines
using CRISPR-Cas9 (19). The data
could provide an opportunity to identify potential β-catenin target
genes by deriving gene-gene functional associations.
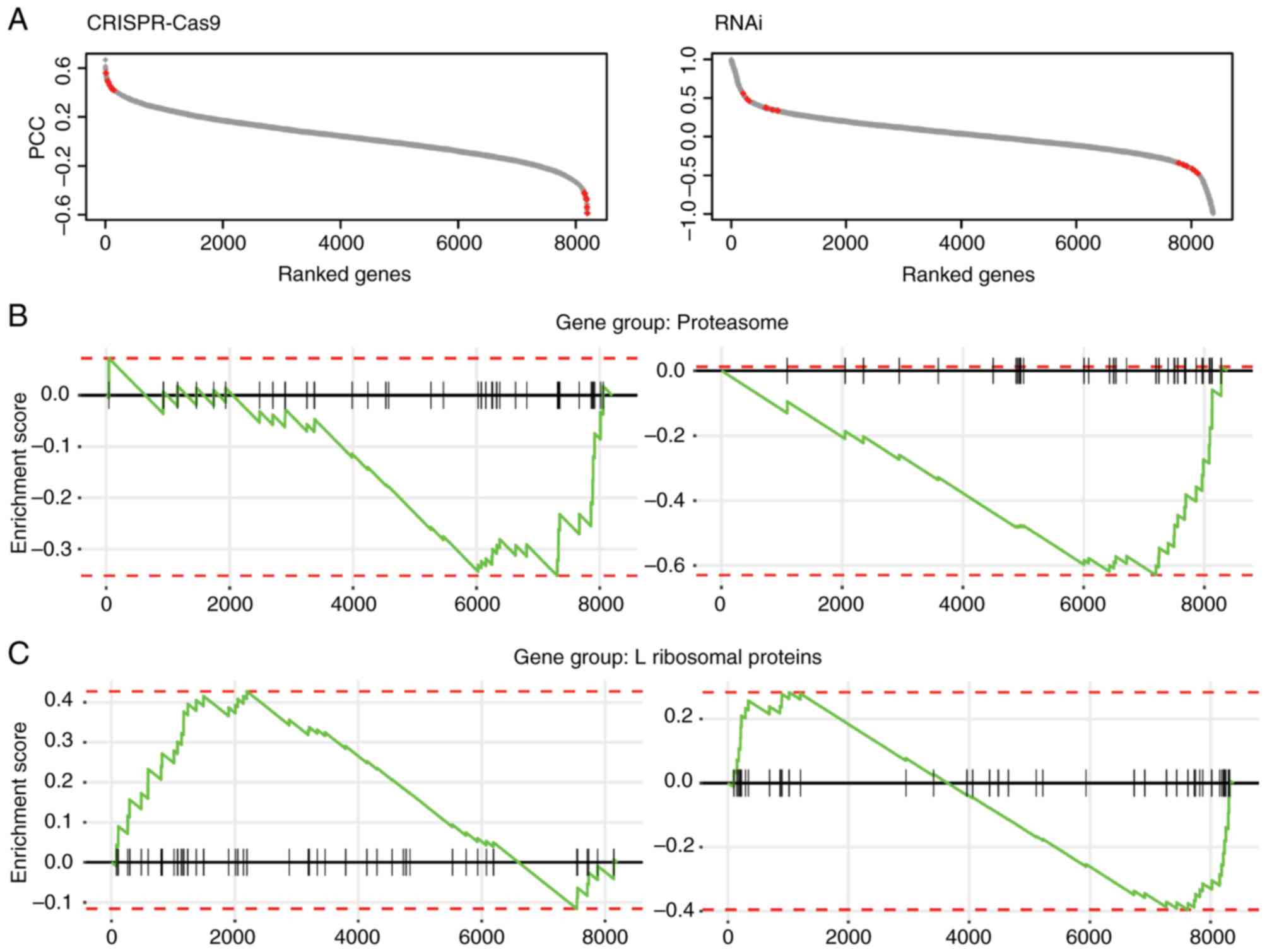
Following filtering of the fitness data, the Pearson
correlation coefficient (PCC) of fitness profiles of all other
genes with β-catenin was computed to generate the fitness profile
correlation landscape. The genes were ranked based on the PCC
scores (between-1 and 1; Fig. 1A).
It was hypothesized that potential targets could be among those
genes exhibiting a positive or negative correlation with the
fitness profile of β-catenin.
All microarray data used in the present study were
retrieved from the National Center for Biotechnology Information
(NCBI) Gene Expression Omnibus (GEO) database (ncbi.nlm.nih.gov/geo/) (21) using GEO accession numbers. The siRNA
β-catenin and control treatment microarray dataset accession
numbers were GSE44097 (10) for the
DLD1 and SW480 cell lines, and GSE18560 (12) for Ls174T cells. The CRC dataset
accession numbers were GSE68468 (22), GSE14333 (23), GSE17536 (24), GSE17537 (25), GSE24549 (26), GSE24550 (27), GSE31595 (28), GSE37892 (29) and GSE39582 (30). For the GSE24549 and GSE24550
datasets, gene expression profiles were generated using an
Affymetrix Human Exon 1.0 ST array, and GSE68468 gene expression
profiles were generated using an Affymetrix Human Genome U133A
array. The remaining data were generated using an Affymetrix Human
Genome U133 Plus 2.0 array. The disease-free survival outcome
information was retrieved from the Prediction of Clinical Outcomes
for Genomics database (https://precog.stanford.edu/index.php) (31) using the aforementioned NCBI GEO
accession numbers.
Raw intensity files (*.CEL) were downloaded from the
GEO database and then processed using an in-house bioinformatics
pipeline. Briefly, the raw files were loaded into R version 3.5.2
(32) environment using the
Bioconductor package oligo (version 1.46.0) (33). The rma algorithms from the oligo
package were applied for background correction and normalization.
Gene annotation was processed based on the custom chip definition
files (version 22.0.0) (34)
downloaded from the BrainArray website (http://brainarray.mbni.med.umich.edu/Brainarray/Database/CustomCDF/genomic_curated_CDF.asp).
RNA-seq data were retrieved from the NCBI Sequence
Read Archive database using the following accession numbers:
SRP029880 (35) for CRC samples and
SRP101345 (36) for HCT116 cell
lines with either mutant catenin b1 (CTNNB1)/β-catenin allele
disrupted or wild-type CTNNB1/β-catenin allele disrupted. The raw
reads files (*.fastq) were downloaded and processed using Salmon
software (version 0.13.0) (37) to
quantify the expression levels of transcripts. Reference
transcriptome data (GRCh38 release 94) were downloaded from the
Ensembl database (http://www.ensembl.org/)
RNA-seq expression profiles of colon cancer (n=521)
were retrieved from the TCGA-COAD dataset deposited in the NCI
Genomic Data Commons Data Portal (https://portal.gdc.cancer.gov) using the Bioconductor
package, TCGAbiolinks (version 2.10.5) (38). The raw fragments per kilobase million
values were converted to TPM (transcripts per million) values for
downstream analysis. The differences between the expression levels
between primary solid tumors and normal solid tissues were tested
using a Wilcoxon rank sum test.
The Cox proportional hazards modeling of CRC
disease-free survival and log-rank tests was conducted using the R
package, survival (version 2.44–1.1; CRAN.R-project.org/package=survival). Using this
statistical model, gene expression values were summarized into the
unified scores. The median value of these scores was chosen as a
threshold, samples with scores above which were defined as
high-risk groups and low-risk groups were determined. The
Kaplan-Meier plot was generated using the R package, survminer
(version 0.4.4; CRAN.R-project.org/package=survminer).
The correlated genes were filtered using stringent
criteria (P≤0.03; Pearson correlation test), and 210 genes in the
CRISPR-Cas9 dataset and 754 genes in RNAi dataset were identified
as potential β-catenin targets, since their fitness profiles were
significantly correlated with β-catenin in CRC cell lines. Since
the resulting fitness profile correlations exhibited similar trends
in terms of gene set enrichment analysis, the overlapping
significantly correlated genes from both datasets were selected,
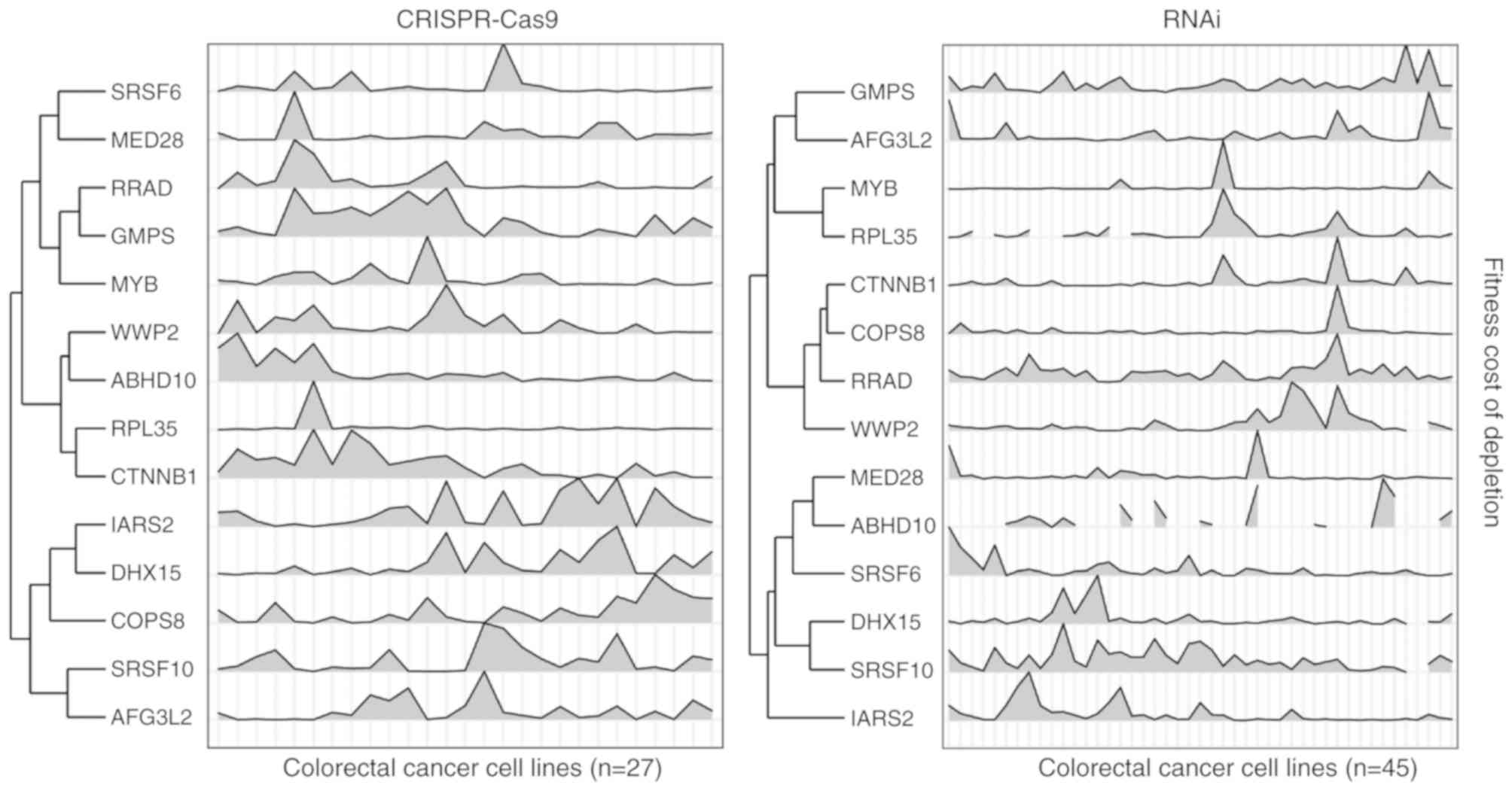
and 13 genes were identified as high confidence β-catenin targets
(Fig. 2; Table I). Notably, these genes seldom
exhibited significant expression alterations in colon cancer cell
lines that were treated with siRNA that inhibited β-catenin, or
harbored β-catenin mutations (Fig.
3).
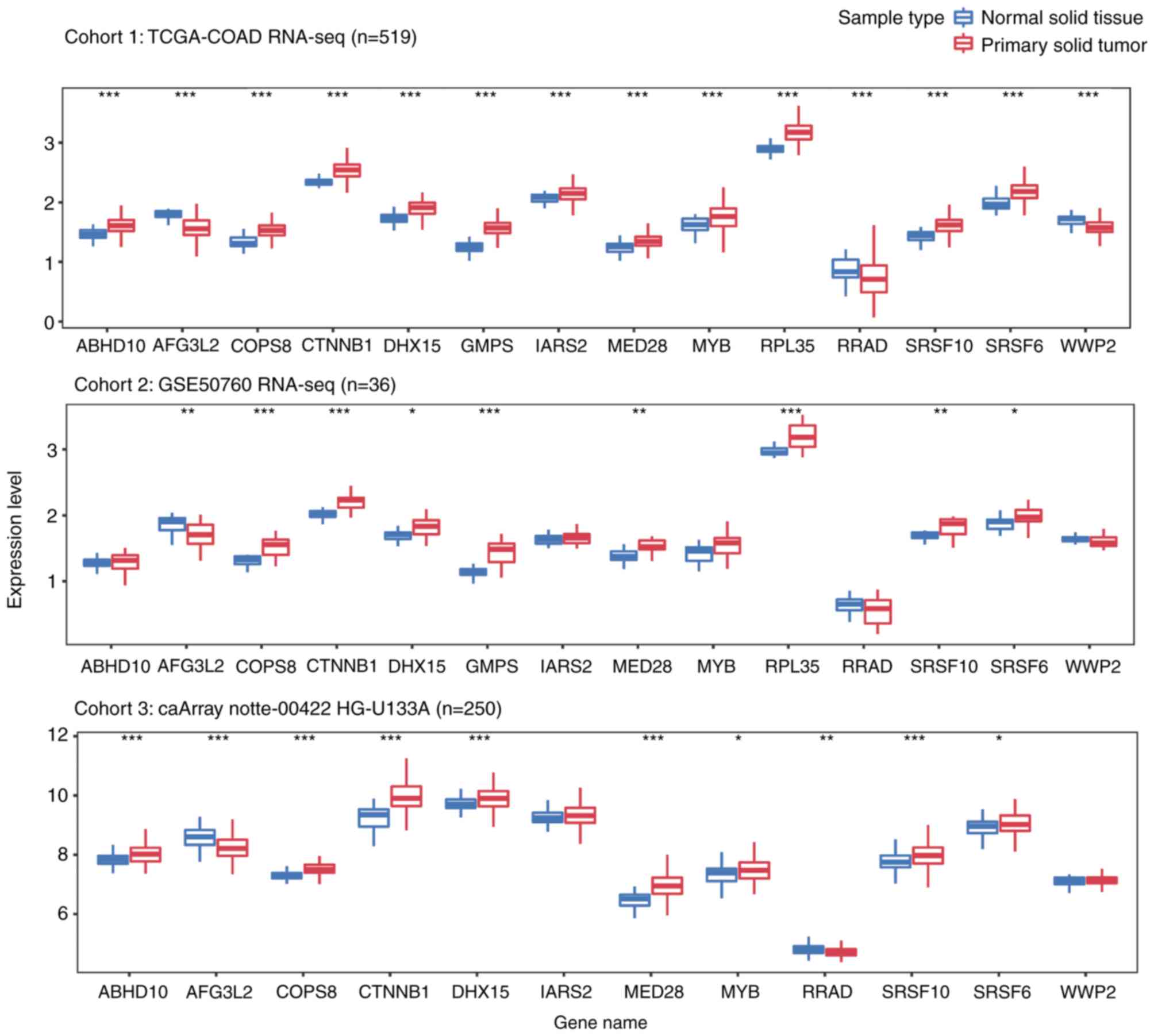
Additionally, the present study explored the
expression levels of these genes in primary CRC and normal tissue
samples from three independent cohorts, including a total of 805
samples. As presented in Fig. 4, all
13 genes were differentially expressed in solid tumor tissues from
cohort 1. Among the genes, ten were upregulated and three were
downregulated in CRC tumor samples. Although similar trends were
observed in the other cohorts, certain genes (ABHD10, IARS2, MYB,
RRAD, WWP2 in cohort2; IARS2 and WWP2 in cohor3), were not
significantly differentially expressed between tumor and normal
samples.
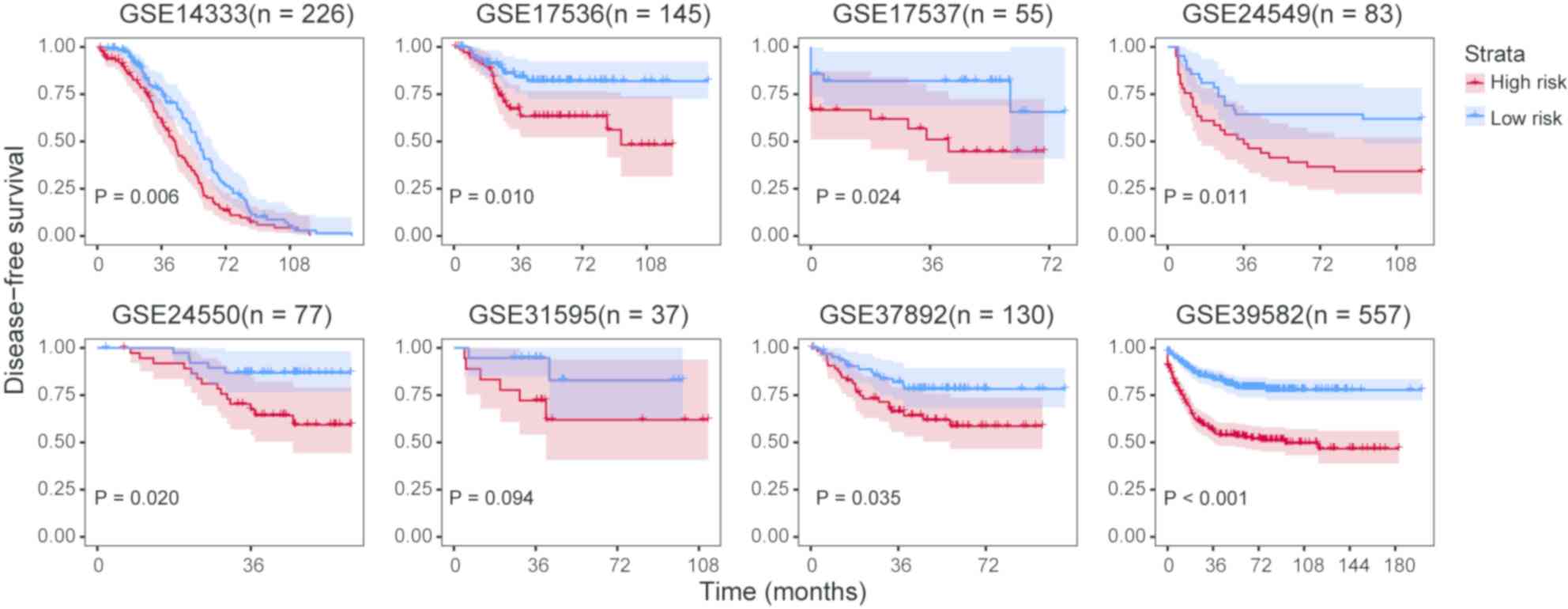
There is increasing interest to discover CRC
prognostic biomarkers as patients could benefit from earlier
diagnosis and a personalized treatment strategy. For this purpose,
the present study aimed to establish gene signature prognostic
models that could predict disease-free survival outcomes in CRC.
The expression profiles of the 13 genes were fitted into a Cox
proportional hazards model and a unified score was computed for
each sample. For each cohort, the median value of the
aforementioned unified score was selected to divide high- and
low-risk groups. Log-rank tests indicated that the high- and
low-risk groups exhibited significant differences in disease-free
survival prognosis except for GSE31595 (Fig. 5).
The present study explored large-scale RNAi and
CRISPR-Cas9 screening data and identified β-catenin target genes
based on fitness profile similarities generated from a large-scale
study, CCLE (20) and the DepMap
(18,19) project. As illustrated in a previous
study, this fitness correlation strategy can recapitulate gene
functional modularity by rewiring human protein complexes and
establishing a human functional similarity network (43). A total of 79 CRC cell lines were
considered in the present study. This set the CRC context, in which
β-catenin regulates target genes. For both RNAi and CRISPR-cas9
datasets, the genes where the fitness data were highly correlated
with β-catenin were significantly enriched in the proteasome and
ribosomal families. Lack of proteasomal degradation of β-catenin
facilitates its entry to the nucleus and targeting genes, including
cyclin D1 (4) and AKT1 (44). This eventually leads to the
proliferation and differentiation of cells. Furthermore,
disturbances in ribosomal proteins have been observed in a variety
of cancerous tissues, including glioblastoma (45), breast (46), esophagus (47), liver (48) and cervix tissues (49). For CRC, similar trends have been
observed. For example, the expression levels of several ribosomal
proteins are abnormally regulated in primary (50) and metastatic (51) CRC. It is worth noting that, in the
previous strategy, the identified β-catenin targets from different
cell lines merely overlapped (10).
This limitation was overcome by computing gene fitness correlation
and thus yielded reliable results.
Notably, among the 13 identified targets, the MYB
proto-oncogene, transcription factor (MYB), has previously been
reported as a known β-catenin target (52). Upregulated MYB levels and activated
β-catenin may induce robust upregulation of MYC promoter activity
in CRC (53). Previous studies have
suggested that the dysregulation of MYB is associated with several
rare types of cancer, including adenoid cystic cancer (54–60),
spiradenocarcinoma (61) and
cutaneous cylindroma (62). For the
mediator complex subunit 28 gene, the same trend in expression
alterations has been observed in epithelial-derived types of cancer
(63). Although, to the best of our
knowledge, there are no reports indicating the involvement of COP9
signalosome subunit 8 (COPS8) in CRC, a proteomic study revealed
that COPS8 is upregulated in prostate cancer (64). The serine and arginine rich splicing
factor 6 and serine and arginine rich splicing factor 10 genes are
members of the serine-arginine family, which regulates RNA
splicing. As a result, their roles in regulating alternative
splicing may promote cancer pathogenesis (65–67). WW
domain containing E3 ubiquitin protein ligase 2 modulates
transforming growth factor b-dependent transcription and
epithelial-mesenchymal transition (68,69). It
should be noted that some of the identified targets seldom
exhibited altered expressions in the siRNA β-catenin approaches.
This suggests that the method used in the present study may serve
as a complementary alternative for the identification of β-catenin
targets, which may be potentially missed by other strategies.
The present study further revealed the associations
of the 13 identified targets with CRC disease free survival
outcomes in eight independent cohorts encompassing 1,310
individuals. Indeed, the prognostic potential of these genes has
been revealed in previous studies (70,71). For
instance, loss of the AFG3 like matrix AAA peptidase subunit 2
gene, located in the 18p11.32–21 region, have been associated with
a significantly longer progression-free survival in patients with
CRC (70). Additionally, MYB is
associated with metastasis in pancreatic tumors (71).
However, it worth mentioning that the present study
used a stringent criterion to filter the data and reported
β-catenin targets with high confidence. While there are two
datasets available, CRISPR-Cas9 and RNAi, the present study
selected the overlap of both datasets for downstream analysis. This
may lead to the misinterpretation of the remaining potential
targets. For example, survivin, cyclin-D1 and axin-2 are well
established β-catenin targets. The present study could not
recapture them, since they exhibited less significant correlations
with β-catenin comparing the identified 13 targets in this study.
It was anticipated that a more robust strategy, such as machine
learning, should be employed to explore the data. Additionally, the
prognostic model requires improvements by statistical modeling, so
that these findings can be applied to clinical practice using less
complicated assays. Finally, experimental studies are required to
verify the findings. The identified β-catenin target genes are of
high confidence and the pathways are associated with CRC
pathogenesis, which provides resources for the research
community.
Not applicable.
No funding was received.
The datasets generated and/or analyzed during the
present study are available in the figshare repository, doi.org/10.6084/m9.figshare.8872769.
HZ and BS conceived of and designed the study. HZ
and LH performed the gene fitness correlation, gene set enrichment,
differential gene experssion and prognostic modelling analysis. DY
collected and re-formatted the data. HZ and BS wrote the
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Novellasdemunt L, Antas P and Li VS:
Targeting Wnt signaling in colorectal cancer. A review in the
theme: Cell signaling: Proteins, pathways and mechanisms. Am J
Physiol Cell Physiol. 309:C511–C521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barker N and Clevers H: Mining the Wnt
pathway for cancer therapeutics. Nat Rev Drug Discov. 5:997–1014.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tetsu O and McCormick F: Beta-Catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shtutman M, Zhurinsky J, Simcha I,
Albanese C, D'Amico M, Pestell R and Ben-Ze'ev A: The cyclin D1
gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad
Sci USA. 96:5522–5527. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He TC, Chan TA, Vogelstein B and Kinzler
KW: PPARδ is an APC-regulated target of nonsteroidal
anti-inflammatory drugs. Cell. 99:335–345. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crawford HC, Fingleton BM, Rudolph-Owen
LA, Goss KJ, Rubinfeld B, Polakis P and Matrisian LM: The
metalloproteinase matrilysin is a target of beta-catenin
transactivation in intestinal tumors. Oncogene. 18:2883–2891. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brabletz T, Jung A, Dag S, Hlubek F and
Kirchner T: Beta-Catenin regulates the expression of the Matrix
Metalloproteinase-7 in human colorectal cancer. Am J Pathol.
155:1033–1038. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hlubek F, Spaderna S, Jung A, Kirchner T
and Brabletz T: Beta-Catenin activates a coordinated expression of
the proinvasive factors laminin-5 gamma2 chain and MT1-MMP in
colorectal carcinomas. Int J Cancer. 108:321–326. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Herbst A, Jurinovic V, Krebs S, Thieme SE,
Blum H, Göke B and Kolligs FT: Comprehensive analysis of β-catenin
target genes in colorectal carcinoma cell lines with deregulated
Wnt/β-catenin signaling. BMC Genomics. 15:742014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mokry M, Hatzis P, Schuijers J, Lansu N,
Ruzius FP, Clevers H and Cuppen E: Integrated genome-wide analysis
of transcription factor occupancy, RNA polymerase II binding and
steady-state RNA levels identify differentially regulated
functional gene classes. Nucleic Acids Res. 40:148–158. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ewing RM, Song J, Gokulrangan G, Bai S,
Bowler EH, Bolton R, Skipp P, Wang Y and Wang Z: Multiproteomic and
transcriptomic analysis of oncogenic β-Catenin molecular networks.
J Proteome Res. 17:2216–2225. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Y, Beyer A and Aebersold R: On the
dependency of cellular protein levels on mRNA abundance. Cell.
165:535–550. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baryshnikova A, Costanzo M, Myers CL,
Andrews B and Boone C: Genetic interaction networks: Toward an
understanding of heritability. Annu Rev Genomics Hum Genet.
14:111–133. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Costanzo M, VanderSluis B, Koch EN,
Baryshnikova A, Pons C, Tan G, Wang W, Usaj M, Hanchard J, Lee SD,
et al: A global genetic interaction network maps a wiring diagram
of cellular function. Science. 353:aaf14202016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boettcher M, Tian R, Blau JA, Markegard E,
Wagner RT, Wu D, Mo X, Biton A, Zaitlen N, Fu H, et al: Dual gene
activation and knockout screen reveals directional dependencies in
genetic networks. Nat Biotechnol. 36:170–178. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen JP, Zhao D, Sasik R, Luebeck J,
Birmingham A, Bojorquez-Gomez A, Licon K, Klepper K, Pekin D,
Beckett AN, et al: Combinatorial CRISPR-Cas9 screens for de novo
mapping of genetic interactions. Nat Methods. 14:573–576. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tsherniak A, Vazquez F, Montgomery PG,
Weir BA, Kryukov G, Cowley GS, Gill S, Harrington WF, Pantel S,
Krill-Burger JM, et al: Defining a cancer dependency map. Cell.
170:564–576.e16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meyers RM, Bryan JG, McFarland JM, Weir
BA, Sizemore AE, Xu H, Dharia NV, Montgomery PG, Cowley GS, Pantel
S, et al: Computational correction of copy number effect improves
specificity of CRISPRCas9 essentiality screens in cancer cells. Nat
Genet. 49:1779–1784. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barretina J, Caponigro G, Stransky N,
Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehár J, Kryukov GV,
Sonkin D, et al: The cancer cell line encyclopedia enables
predictive modelling of anticancer drug sensitivity. Nature.
483:603–607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41(Database Issue):
D991–D995. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsafrir D, Bacolod M, Selvanayagam Z,
Tsafrir I, Shia J, Zeng Z, Liu H, Krier C, Stengel RF, Barany F, et
al: Relationship of gene expression and chromosomal abnormalities
in colorectal cancer. Cancer Res. 66:2129–2137. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jorissen RN, Gibbs P, Christie M, Prakash
S, Lipton L, Desai J, Kerr D, Aaltonen LA, Arango D, Kruhøffer M,
et al: Metastasis-Associated gene expression changes predict poor
outcomes in patients with dukes Stage B and C colorectal cancer.
Clin Cancer Res. 15:7642–7651. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Smith JJ, Deane NG, Wu F, Merchant NB,
Zhang B, Jiang A, Lu P, Johnson JC, Schmidt C, Bailey CE, et al:
Experimentally derived metastasis gene expression profile predicts
recurrence and death in patients with colon cancer.
Gastroenterology. 138:958–968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Freeman TJ, Smith JJ, Chen X, Washington
MK, Roland JT, Means AL, Eschrich SA, Yeatman TJ, Deane NG and
Beauchamp RD: Smad4-mediated signaling inhibits intestinal
neoplasia by inhibiting expression of β-catenin. Gastroenterology.
142:562–571.e2. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sveen A, Agesen TH, Nesbakken A, Rognum
TO, Lothe RA and Skotheim RI: Transcriptome instability in
colorectal cancer identified by exon microarray analyses:
Associations with splicing factor expression levels and patient
survival. Genome Med. 3:322011. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Agesen TH, Sveen A, Merok MA, Lind GE,
Nesbakken A, Skotheim RI and Lothe RA: ColoGuideEx: A robust gene
classifier specific for stage II colorectal cancer prognosis. Gut.
61:1560–1567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thorsteinsson M, Kirkeby LT, Hansen R,
Lund LR, Sørensen LT, Gerds TA, Jess P and Olsen J: Gene expression
profiles in stages II and III colon cancers: Application of a
128-gene signature. Int J Colorectal Dis. 27:1579–1586. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Laibe S, Lagarde A, Ferrari A, Monges G,
Birnbaum D and Olschwang S; COL2 Project, : A seven-gene signature
aggregates a subgroup of stage II colon cancers with stage III.
OMICS. 16:560–565. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Marisa L, de Reyniès A, Duval A, Selves J,
Gaub MP, Vescovo L, Etienne-Grimaldi MC, Schiappa R, Guenot D,
Ayadi M, et al: Gene expression classification of colon cancer into
molecular subtypes: Characterization, validation, and prognostic
value. PLoS Med. 10:e10014532013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gentles AJ, Newman AM, Liu CL, Bratman SV,
Feng W, Kim D, Nair VS, Xu Y, Khuong A, Hoang CD, et al: The
prognostic landscape of genes and infiltrating immune cells across
human cancers. Nat Med. 21:938–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
RCoreTeam: R, . A language and environment
for statistical computing. 2018.
|
|
33
|
Carvalho BS and Irizarry RA: A framework
for oligonucleotide microarray preprocessing. Bioinformatics.
26:2363–2367. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dai M, Wang P, Boyd AD, Kostov G, Athey B,
Jones EG, Bunney WE, Myers RM, Speed TP, Akil H, et al: Evolving
gene/transcript definitions significantly alter the interpretation
of GeneChip data. Nucleic Acids Res. 33:e1752005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim SK, Kim SY, Kim JH, Roh SA, Cho DH,
Kim YS and Kim JC: A nineteen gene-based risk score classifier
predicts prognosis of colorectal cancer patients. Mol Oncol.
8:1653–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu D, Skomorovska Y, Song J, Bowler E,
Harris R, Ravasz M, Bai S, Ayati M, Tamai K, Koyuturk M, et al:
ELF3 is an antagonist of oncogenic-signalling-induced expression of
EMT-TF ZEB1. Cancer Biol Ther. 20:90–100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Patro R, Duggal G, Love MI, Irizarry RA
and Kingsford C: Salmon provides fast and bias-aware quantification
of transcript expression. Nat Methods. 14:417–419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Colaprico A, Silva TC, Olsen C, Garofano
L, Cava C, Garolini D, Sabedot TS, Malta TM, Pagnotta SM,
Castiglioni I, et al: TCGAbiolinks: An R/Bioconductor package for
integrative analysis of TCGA data. Nucleic Acids Res. 44:e712016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Series B (Methodological).
57:289–300. 1995. View Article : Google Scholar
|
|
41
|
Sergushichev A: An algorithm for fast
preranked gene set enrichment analysis using cumulative statistic
calculation. Jun 20–2016.doi: https://doi.org/10.1101/060012.
View Article : Google Scholar
|
|
42
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pan J, Meyers RM, Michel BC, Mashtalir N,
Sizemore AE, Wells JN, Cassel SH, Vazquez F, Weir BA, Hahn WC, et
al: Interrogation of mammalian protein complex structure, function,
and membership using genome-scale fitness screens. Cell Syst.
6:555–568.e7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dihlmann S, Kloor M, Fallsehr C and von
Knebel Doeberitz M: Regulation of AKT1 expression by
beta-catenin/Tcf/Lef signaling in colorectal cancer cells.
Carcinogenesis. 26:1503–1512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lopez CD, Martinovsky G and Naumovski L:
Inhibition of cell death by ribosomal protein L35a. Cancer Lett.
180:195–202. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Henry JL, Coggin DL and King CR:
High-level expression of the ribosomal protein L19 in human breast
tumors that overexpress erbB-2. Cancer Res. 53:1403–1408.
1993.PubMed/NCBI
|
|
47
|
Wang Q, Yang C, Zhou J, Wang X, Wu M and
Liu Z: Cloning and characterization of full-length human ribosomal
protein L15 cDNA which was overexpressed in esophageal cancer.
Gene. 263:205–209. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim JH, You KR, Kim IH, Cho BH, Kim CY and
Kim DG: Over-expression of the ribosomal protein L36a gene is
associated with cellular proliferation in hepatocellular carcinoma.
Hepatology. 39:129–138. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cheng Q, Lau WM, Chew SH, Ho TH, Tay SK
and Hui KM: Identification of molecular markers for the early
detection of human squamous cell carcinoma of the uterine cervix.
Br J Cancer. 86:274–281. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kitahara O, Furukawa Y, Tanaka T, Kihara
C, Ono K, Yanagawa R, Nita ME, Takagi T, Nakamura Y and Tsunoda T:
Alterations of gene expression during colorectal carcinogenesis
revealed by cDNA microarrays after laser-capture microdissection of
tumor tissues and normal epithelia. Cancer Res. 61:3544–3549.
2001.PubMed/NCBI
|
|
51
|
Bertucci F, Salas S, Eysteries S, Nasser
V, Finetti P, Ginestier C, Charafe-Jauffret E, Loriod B, Bachelart
L, Montfort J, et al: Gene expression profiling of colon cancer by
DNA microarrays and correlation with histoclinical parameters.
Oncogene. 23:1377–1391. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
van de Wetering M, Sancho E, Verweij C, de
Lau W, Oving I, Hurlstone A, van der Horn K, Batlle E, Coudreuse D,
Haramis AP, et al: The beta-catenin/TCF-4 complex imposes a crypt
progenitor phenotype on colorectal cancer cells. Cell. 111:241–250.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ciznadija D, Tothill R, Waterman ML, Zhao
L, Huynh D, Yu RM, Ernst M, Ishii S, Mantamadiotis T, Gonda TJ, et
al: Intestinal adenoma formation and MYC activation are regulated
by cooperation between MYB and Wnt signaling. Cell Death Differ.
16:1530–1538. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gao R, Cao C, Zhang M, Lopez MC, Yan Y,
Chen Z, Mitani Y, Zhang L, Zajac-Kaye M, Liu B, et al: A unifying
gene signature for adenoid cystic cancer identifies parallel
MYB-dependent and MYB-independent therapeutic targets. Oncotarget.
5:12528–12542. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rettig EM, Tan M, Ling S, Yonescu R,
Bishop JA, Fakhry C and Ha PK: MYB rearrangement and
clinicopathologic characteristics in head and neck adenoid cystic
carcinoma. Laryngoscope. 125:E292–E299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
North JP, McCalmont TH, Fehr A, van Zante
A, Stenman G and LeBoit PE: Detection of MYB alterations and other
immunohistochemical markers in primary cutaneous adenoid cystic
carcinoma. Am J Surg Pathol. 39:1347–1356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bishop JA, Yonescu R, Epstein JI and
Westra WH: A subset of prostatic basal cell carcinomas harbor the
MYB rearrangement of adenoid cystic carcinoma. Hum Pathol.
46:1204–1208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Argyris PP, Wetzel SL, Greipp P, Wehrs RN,
Knutson DL, Kloft-Nelson SM, García JJ and Koutlas IG: Clinical
utility of myb rearrangement detection and p63/p40
immunophenotyping in the diagnosis of adenoid cystic carcinoma of
minor salivary glands: A pilot study. Oral Surg Oral Med Oral
Pathol Oral Radiol. 121:282–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Drier Y, Cotton MJ, Williamson KE,
Gillespie SM, Ryan RJ, Kluk MJ, Carey CD, Rodig SJ, Sholl LM,
Afrogheh AH, et al: An oncogenic MYB feedback loop drives alternate
cell fates in adenoid cystic carcinoma. Nat Genet. 48:265–272.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen TY, Keeney MG, Chintakuntlawar AV,
Knutson DL, Kloft-Nelson S, Greipp PT, Garrity JA, Salomao DR and
Garcia JJ: Adenoid cystic carcinoma of the lacrimal gland is
frequently characterized by MYB rearrangement. Eye (Lond).
31:720–725. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
van der Horst MP, Marusic Z, Hornick JL,
Luzar B and Brenn T: Morphologically low-grade spiradenocarcinoma:
A clinicopathologic study of 19 cases with emphasis on outcome and
MYB expression. Mod Pathol. 28:944–953. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Rajan N, Andersson MK, Sinclair N, Fehr A,
Hodgson K, Lord CJ, Kazakov DV, Vanecek T, Ashworth A and Stenman
G: Overexpression of MYB drives proliferation of CYLD-defective
cylindroma cells. J Pathol. 239:197–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang L, Maul RS, Rao J, Apple S, Seligson
D, Sartippour M, Rubio R and Brooks MN: Expression pattern of the
novel gene EG-1 in cancer. Clin Cancer Res. 10:3504–3508. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee EK, Cho H and Kim CW: Proteomic
analysis of cancer stem cells in human prostate cancer cells.
Biochem Biophys Res Commun. 412:279–285. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wei N, Cheng Y, Wang Z, Liu Y, Luo C, Liu
L, Chen L, Xie Z, Lu Y and Feng Y: SRSF10 plays a role in myoblast
differentiation and glucose production via regulation of
alternative splicing. Cell Rep. 13:1647–1657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhou X, Li X, Cheng Y, Wu W, Xie Z, Xi Q,
Han J, Wu G, Fang J and Feng Y: BCLAF1 and its splicing regulator
SRSF10 regulate the tumorigenic potential of colon cancer cells.
Nat Commun. 5:45812014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li H, Cheng Y, Wu W, Liu Y, Wei N, Feng X,
Xie Z and Feng Y: SRSF10 regulates alternative splicing and is
required for adipocyte differentiation. Mol Cell Biol.
34:2198–2207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Soond SM, Smith PG, Wahl L, Swingler TE,
Clark IM, Hemmings AM and Chantry A: Novel WWP2 ubiquitin ligase
isoforms as potential prognostic markers and molecular targets in
cancer. Biochim Biophys Acta 1832. 2127–2135. 2013.
|
|
69
|
Soond SM and Chantry A: Selective
targeting of activating and inhibitory Smads by distinct WWP2
ubiquitin ligase isoforms differentially modulates TGFβ signalling
and EMT. Oncogene. 30:2451–2462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Haan JC, Labots M, Rausch C, Koopman M,
Tol J, Mekenkamp LJ, van de Wiel MA, Israeli D, van Essen HF, van
Grieken NC, et al: Genomic landscape of metastatic colorectal
cancer. Nat Commun. 5:54572014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Srivastava SK, Bhardwaj A, Arora S, Singh
S, Azim S, Tyagi N, Carter JE, Wang B and Singh AP: MYB is a novel
regulator of pancreatic tumour growth and metastasis. Br J Cancer.
113:1694–1703. 2015. View Article : Google Scholar : PubMed/NCBI
|