Introduction
Gastric cancer is a common malignant tumor (1). According to the American College of
Surgeons, the local or regional recurrence rate following attempted
radical resection is ~40%, and the systemic recurrence rate is ~60%
(2). Patients with gastric cancer
develop recurrence and metastasis following surgical treatment
(3).
Enhancer of zeste homolog 2 (EZH2) is an important
target in the study of the inhibition of gastric cancer invasion
and metastasis. Studies have reported that abnormal expression of
epigenetic regulation factor polycomb group (PcG) protein is
closely associated with the occurrence and development of tumors
(4,5). In mammals, two main Polycomb group
complexes exist-Polycomb repressive complex 1 (PRC1) and 2 (PRC2)
(6). PRC2 contributes to chromatin
compaction and catalyzes the methylation of histone H3 at lysine 27
(6). The core PRC2 complex, which is
conserved from Drosophila to mammals, comprises four
components: EZH1/2, SUZ12, EED and RbAp46/48 (6). EZH2 is a key member of PRC2 function
that acts as a histone methyltransferase targeting lysine-27 of
histone H3 (H3K27) and occurs in various malignancies, including
prostate cancer, breast cancer, and glioblastoma multiforme
(7). Based on its ability to
modulate transcription of key genes involved in cell cycle control,
DNA repair, and cell differentiation, EZH2 is believed to serve a
crucial role in tissue-specific stem cell maintenance and tumor
development (7). In addition, EZH2
expression can influence the expression of tumor suppressor genes,
leading to malignant signalling pathways being activated, including
activation of the PI3K/Akt and Wnt/β-catenin pathways (8,9).
3-deazaneplanocin A (DZNep) is an
S-adenosylhomocysteine hydrolase inhibitor and a pharmacological
inhibitor of histone methylation; the effect of DZNep on cancer
cells is relatively specific for EZH2 (10) Studies on various malignant types of
tumor have demonstrated that DZNep may serve an antitumor role via
the inhibition of EZH2 (11–13); however, the underlying mechanisms
remain unclear.
The microenvironment of tumor cells during growth
provides stimulating signals of EZH2 expression. For example,
hypoxia inducible factor 1α (Hif-1α) of the solid tumor
microenvironment can induce EZH2 expression (14). During the process of tumor evolution,
Hif-1α may promote tumor cell proliferation and angiogenesis, and
may cause tumor recurrence and metastasis (15). There is interactive regulation among
Hif-1α and Wnt/β-catenin signalling molecules (16) and EZH2 in glioblastoma (17,18). In
the present study, DZNep was used to observe the effect of DZNep on
the proliferation, apoptosis, invasion and metastasis of BGC-823
gastric cancer cells, and to investigate the function of DZNep in
the regulation of Hif-1α and Wnt/β-catenin signalling molecules via
the inhibition of EZH2. The results of the present study provided a
theoretical basis for the functions and mechanisms of inhibition of
BGC-823 gastric cancer cells by DZNep.
Materials and methods
Cell culture
The poorly differentiated human gastric
adenocarcinoma cell line BGC-823 was purchased from Shanghai Xin Yu
Biotech Co., Ltd. BGC-823 cells were cultured in a sterile
thermostatic incubator at 37°C with 5% CO2 in RPMI-1640
culture medium (HyClone; GE Healthcare Life Sciences) containing
10% FBS (Hangzhou Sijiqing Biological Engineering Materials Co.,
Ltd.) supplemented with 100 U/ml penicillin and 100 mg/ml
streptomycin (HyClone; GE Healthcare Life Sciences). The culture
medium was replaced every other day, and cell passage was performed
every 3–4 days. Cells were cultured to the logarithmic growth phase
and subsequently plated in 60 mm culture plates. BGC-823 cells were
observed under an inverted microscope at ×200 and ×400
magnification.
Experimental groups
The cells in the experiment were divided into the
DZNep group and the control group. DZNep was purchased from
Sigma-Aldrich (Merck KGaA), dissolved in DMSO solution and stored
at −20°C. The DZNep stock solution (10 mM) was dissolved in
phosphate-buffered saline (PBS) for cell culture. The DZNep group
was incubated at 37°C with culture medium containing 5 µM DZNep
with 0.167 µl DMSO for cell colony, CCK8, wound healing, Transwell
and flow cytometry assays. The control group was incubated with
culture medium containing an equal volume of DMSO without
DZNep.
Cell colony assay
A total of 500 cells were seeded into a 60 mm
culture plate. The culture medium was replaced when the cells had
stably attached. Culture medium containing DZNep or DMSO was added
into the culture plate for continuous culture at 37°C for 5, 7 or
10 days. Cells were fixed in 4% paraformaldehyde for 10 min and
stained with 1% crystal violet for 20 min at room temperature. The
number of visible cell colonies was counted for statistical
analyses.
CCK-8 proliferation assay
CCK-8 is a kit developed by Tongren Chemical
Research Institute (Dojindo Molecular Technologies, Inc.) for the
detection of toxicity/proliferation of cells, which uses a
water-soluble tetrazolium salt assay. The cell suspension density
was adjusted to 2.5×104 cells/ml using culture medium
containing either DZNep or DMSO. Cells (200 µl/well) were plated
into six 96-well plates. Each group had eight replicate wells.
Culture medium containing 10% FBS was used as the blank control in
this experiment. After 0, 6, 12, 24, 36 and 48 h, one 96-well plate
was removed from the incubator, and 10 µl CCK-8 solution was added
into each well. Cells were continuously cultured for another 4 h,
and the absorbance (optical density) values of each well were
detected using a microplate reader at 450 nm. The lowest and
highest values in the experimental results were excluded, and the
mean values were calculated.
Wound healing assay
A pen was used to evenly draw a line on the back of
a 6-well plate, so that three lines crossed each well. Cells
(~3×105) were added into each well. When the cells had
stably attached and grown to ~100% confluence, a scratch
perpendicular to the lines on the back of the 6-well plate was made
using a pipette tip and a ruler. Cells were washed with PBS three
times to remove the scratched cells. Serum-free RPMI-1640 medium
containing DZNep or DMSO was added; the samples were incubated at
37°C and images were captured at 0, 20, 25 and 44 h using a CKX41
light microscope (magnification, ×100; Olympus Corporation).
Transwell invasion assay
Cell density was adjusted to 1×105
cells/ml and the top chamber of the Transwell inserts was coated
with Matrigel prior to the assay. Cell suspension (200 µl) was
added to the top chamber, and 500 µl culture medium containing
DZNep or DMSO was added to the bottom chamber of the 24-well plate.
Serum-free medium was added to the upper chamber. Serum was added
to the medium in the lower chamber. Cells were cultured for 24 h at
37°C and the culture medium in the top chamber was aspirated. The
cells in the top chamber were wiped off using a cotton swab. The
invasive cells in the lower chamber were stained with 0.1% crystal
violet for 15 min at 37°C. An Olympus DP73 optical microscope
(Olympus Corporation) was used to observe migratory cells
(magnification, ×200). Cells in 5–10 fields were counted, and mean
values were calculated.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was performed to detect and quantify the
mRNA expression in the DZNep treatment groups. Total RNA in BGC-823
cells at different time points was extracted using
TRIzol® (cat. no. 15596026; Thermo Fisher Scientific,
Inc.), and cDNA was generated from the total RNA using a reverse
transcription reagent kit (cat. no. 639505; Takara Biotechnology
Co., Ltd.) for DNA amplification under the following thermocycling
conditions: 3 cycles at 37°C for 15 min, followed by 85°C for 5 sec
and hold at 4°C. The primers used were as follows: Hif-1α forward,
5′-GAAAGCGCAAGTCTTCAAAG-3′ and reverse, 5′-TGGGTAGGAGATGGAGATGC-3′
(amplified fragment, 167 bp; Sangon Biotech Co., Ltd.); β-actin
forward, 5′-ACACTGTGCCCATCTACG-3′ and reverse,
5′-TGTCACGCACGATTTCC-3′ (amplified fragment, 152 bp). qPCR was
performed using a SYBR® Green PCR kit (cat. no. KM4101;
Kapa Biosystems; Roche Diagnostics). The reactions were incubated
in a 96-well optical plate at 95°C for 3 min, 95°C for 5 sec, 56°C
for 10 sec, 72°C for 25 sec, and 39 cycles of 65°C for 5 sec and
95°C for 50 sec. The 2−ΔΔCq method was used and
expression values were normalized to β-actin (19).
Western blotting
BGC-823 cells were cultured for 2, 8, 12 and 24 h
with 3, 5 or 10 µM DZNep. Total protein was extracted using a whole
protein extraction reagent kit (Nanjing KeyGen Biotech Co., Ltd.).
The protein concentration was determined using a bicinchoninic acid
protein content detection reagent kit (Nanjing Keygen Biotech Co.,
Ltd.). An equal amount of protein (150 µg/lane) was separated by
SDS-PAGE (8–10%). Proteins were transferred onto a nitrocellulose
membrane and blocked in 10% skimmed milk at room temperature for
1–1.5 h. The membrane was incubated with the following primary
antibodies: Hif-1α rabbit polyclonal antibody (cat. no. 20960-1-AP;
ProteinTech Group, Inc.), EZH2 (D2C9 XP®) rabbit
monoclonal antibody (mAb; cat. no. 5246; Cell Signaling Technology,
Inc.), β-catenin (D10A8) XP® rabbit mAb (cat. no. 8480;
Cell Signaling Technology, Inc.), non-phosphorylated (active)
β-catenin (Ser45) (D2U8Y) XP® rabbit mAb (N-P-β-catenin;
cat. no. 19807; Cell Signaling Technology, Inc.), β-actin (8H10D10)
mouse mAb (cat. no. 3700; Cell Signaling Technology, Inc.) and
phosphorylated-β-catenin (P-β-catenin; cat. no. sc-16743-R; Santa
Cruz Biotechnology, Inc.); diluted at 1:1,000 in antibody dilution
buffer at 4°C overnight. The membrane was incubated with goat
anti-mouse immunoglobulin G (IgG; 1:5,000; cat. no. TA130005;
OriGene Technologies, Inc.) or goat anti-rabbit IgG (1:5,000; cat.
no. TA130024; OriGene Technologies, Inc.) secondary antibody at
room temperature for 0.5–1 h. The bands were developed using the
ECL chemiluminescence reagent (ECL kit; cat. no. KGP1121; Nanjing
KeyGen Biotech Co., Ltd.) for 1 min and visualized with an Amersham
Imager 600 instrument (GE Healthcare Life Sciences). Densitometric
analysis was performed using Image-Pro Plus version 6.0 (Media
Cybernetics, Inc.).
Flow cytometry
A total of 6×105 cells were added into
each well of a 6-well plate. Following stable attachment of the
cells, the culture medium was replaced with culture medium
containing either DZNep or DMSO for 24 h. Subsequently, cells were
digested, centrifuged in 800 × g at 4°C for 5 min and transferred
to a new centrifuge tube. A total of 400 µl 1X binding buffer was
added to each tube and mixed evenly. A total of 5 µl Annexin V-FITC
was added, gently mixed and incubated at 4°C in the dark for 15
min. Finally, 10 µl propidium iodide was added, gently mixed and
incubated at 4°C in the dark for 5 min. Cells were detected within
30 min using a FACScalibur flow cytometer and CellQuest software
(BD Biosciences).
Statistical analysis
Experiments were performed in triplicate. The
results are presented as the mean ± standard deviation. Differences
between two groups were compared using an unpaired Student's
t-test. For multiple groups, statistical significance was analysed
using one-way analysis of variance followed by a Bonferroni
comparison post hoc test. Analysis was performed using SPSS version
17.0 software (SPSS, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
DZNep reduces proliferation and colony
formation in gastric cancer cells
Following treatment with 5 µM DZNep, BGC-823 cells
were observed under an inverted microscope. The results
demonstrated that cells exhibited morphological changes after 24 h.
In the control group, cells were plump, had round and smooth edges,
and had an orderly arrangement. In the DZNep group, cells became
thin, long and thread-shaped with a small circular terminal, and
cells were arranged loosely (Fig.
1A). CCK-8 proliferation experiment results demonstrated that
after 6, 12, 36, and 48 h of DZNep-treatment, the proliferation
activity of BGC-823 cells was significantly lower compared with
that in the control group (Fig. 1B).
Cell colony formation results illustrated that the number of cell
colonies in the DZNep group was significantly lower compared with
that in the control group, and the differences on days 5 and 7 were
statistically significant (Fig. 1C).
Overall, these results suggest that DZNep inhibits the
proliferation and colony formation abilities of BGC-823 cells.
 | Figure 1.DZNep reduces proliferation and
colony formation of BGC-823 cells. (A) Compared with cells in the
control group, cells in the DZNep group became thin and long, and
exhibited a thread-like shape with a small circular terminal, and
the arrangement of the cells became loose (magnification, ×200 and
×400). (B) In the CCK-8 assay, the absorbance of the DZNep group
was significantly lower compared with that of the control group at
6 and 48 h. *P<0.05, **P<0.01 vs. respective DZNep group. (C)
The number of colonies in the DZNep group was significantly lower
compared with that in the control group at 5 and 7 days.
*P<0.05, **P<0.01 vs. respective control group. CCK-8, Cell
Counting kit-8; DZNep, 3-deazaneplanocin A; OD, optical
density. |
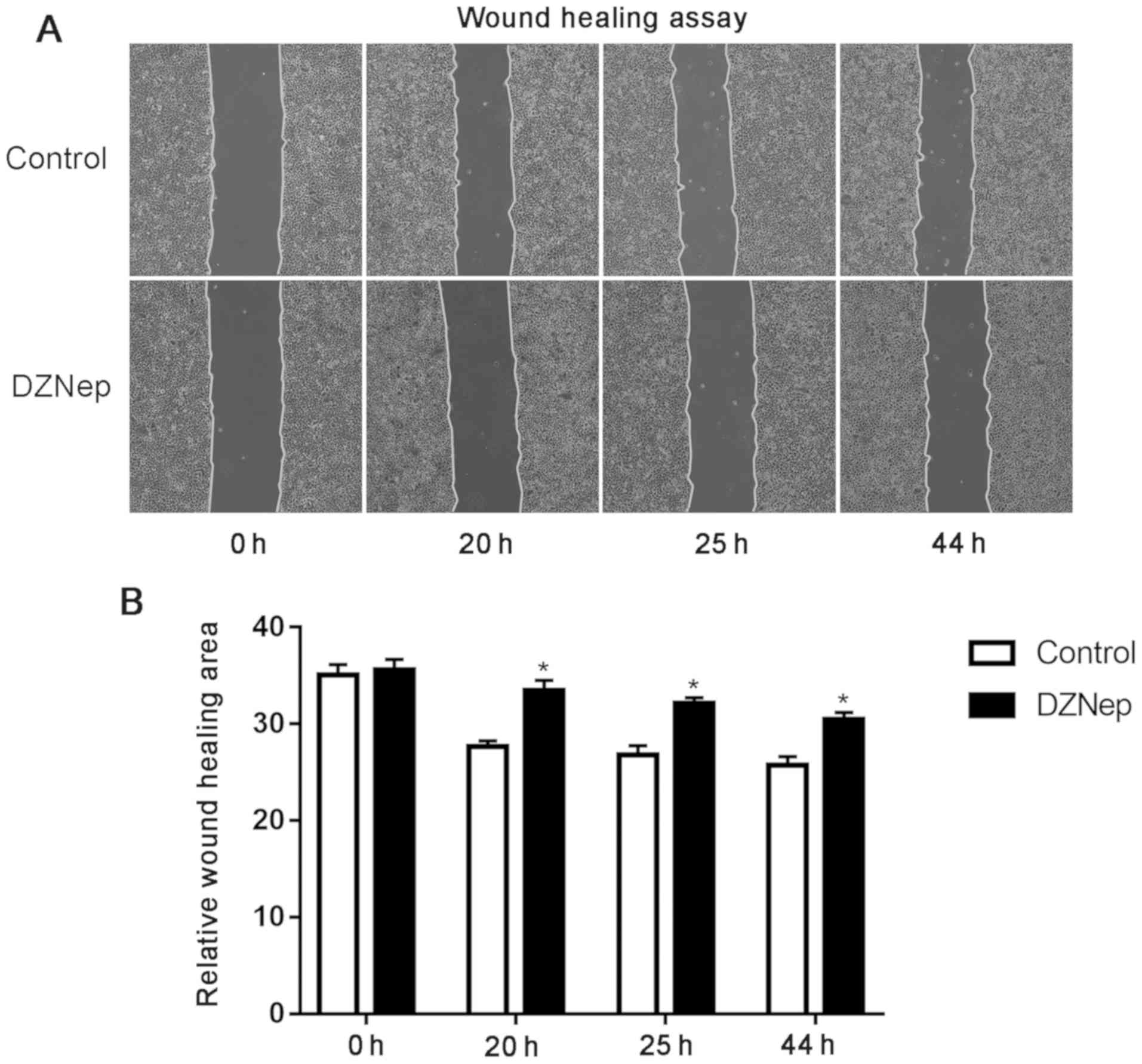
Wound healing is reduced by DZNep
treatment
Scratch treatment in the DZNep (5 µM) and control
groups was performed at the same time. The width of the cell
scratch at the same location was measured after 0, 20, 25 and 44 h.
Over time, cells gradually spread from the scratch edge and grew to
the edge of the other side, thus gradually reducing the distance
between the residual scratches. The results demonstrated that the
distance between residual scratches in the DZNep intervention group
was significantly wider compared with that in the control group
(Fig. 2), which suggests that DZNep
reduced the migration ability of the cells.
DZNep reduces the invasion ability of
gastric cancer cells
Following 2, 6, 12 and 24 h of DZNep treatment (5
µM) of BGC-823 cells, numbers of cells that passed through the
Transwell membranes were observed. The results demonstrated that
there was no obvious movement of cells through the membrane after 2
h in the control and DZNep groups. At 6 h, a small number of cells
in the DZNep and control groups passed through the membrane. At 12
and 24 h, the number of cells that passed through the membrane in
the control group was significantly higher compared with that in
the DZNep group (Fig. 3). The
results suggest that DZNep reduced the invasive ability of the
cells.
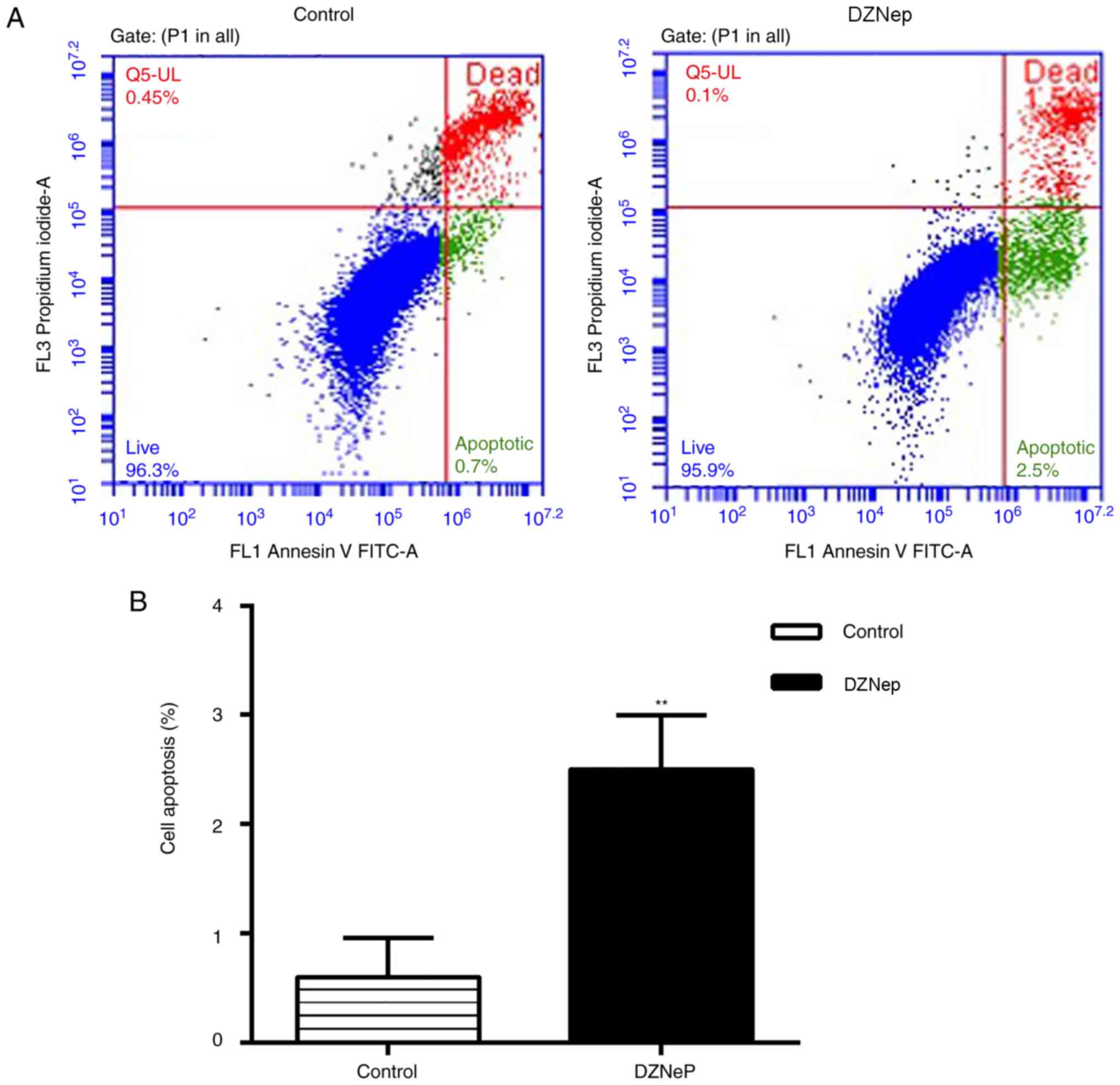
DZNep promotes BGC-823 cell
apoptosis
Following 24 h of 5 µM DZNep treatment of BGC-823
cells, flow cytometry analysis revealed that the apoptotic rate was
2.5% in the DZNep group and 0.7% in the control group (Fig. 4), suggesting that DZNep promoted
apoptosis.
DZNep inhibits mRNA and protein
expression of Hif-1α
Detection of Hif-1α mRNA expression in BGC-823 cells
after 4, 8 and 24 h following 1 and 5 µM DZNep treatment was
performed by RT-qPCR. The internal control used was β-actin. The
results demonstrated that following 4 h of treatment with 1 and 5
µM DZNep, and after 24 h with 5 µM DZNep, Hif-1α was significantly
decreased by DZNep treatment. At 8 h, 1 µM DZNep slightly increased
Hif-1α expression compared with the control group, but the change
was non-significant; this suggested that the increase in Hif-1α
reactivity may be transient (Fig.
5A).
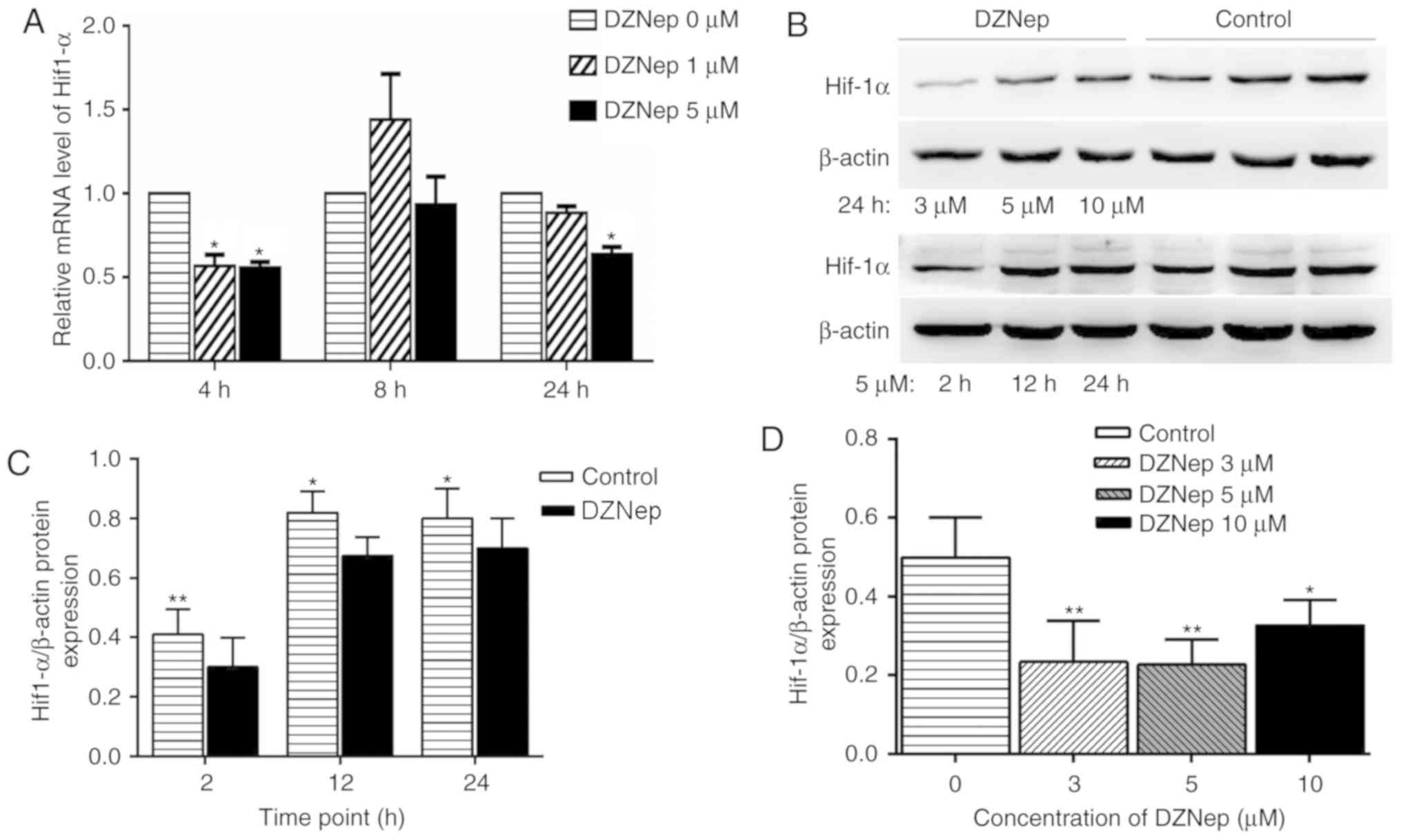 | Figure 5.DZNep-treatment reduces Hif-1α gene
and protein expression. (A) Hif-1α relative mRNA expression levels
in BGC-823 cells after 4, 8 and 24 h treatment with 1 and 5 µM
DZNep were detected by reverse transcription-quantitative PCR.
Following 4 h of treatment with 1 and 5 µM DZNep, and 24 h with 5
µM DZNep, Hif-1α expression was significantly reduced. *P<0.05
vs. respective DZNep 0 µM group. (B) Effects of 3, 5 and 10 µM
DZNep treatment on Hif-1α protein expression levels after 24 h were
detected using western blotting. The control group was treated with
DMSO. The internal control was β-actin. (C) Hif-1α protein
expression levels after 2, 12 and 24 h treatment with 5 µM DZNep
decreased at each time point, and the differences were
statistically significant. *P<0.05, **P<0.01 vs. control. (D)
DZNep treatment (3, 5 and 10 µM) significantly decreased the
protein expression levels of Hif-1α, and the differences were
statistically significant. *P<0.05 and **P<0.01 vs. control.
DZNep, 3-deazaneplanocin A; Hif-1α, hypoxia-inducible factor
1α. |
Detection of Hif-1α protein expression was performed
by western blot analysis. The internal control was β-actin.
Following treatment with 3, 5 and 10 µM DZNep for 24 h, western
blotting demonstrated that 3, 5 and 10 µM DZNep significantly
reduced the protein expression level of Hif-1α. The alterations in
the protein expression levels of Hif-1α after 2, 12 and 24 h of 5
µM DZNep treatment indicated that the Hif-1α protein expression
levels at each time point were significantly reduced (Fig. 5B-D).
Inhibitory function of DZNep on EZH2
and β-catenin proteins
The inhibitory function of 3, 5 and 10 µM DZNep
treatment for 2, 8, 12 and 24 h on the protein expression levels of
EZH2, P-β-catenin and N-P-β-catenin in BGC-823 cells was detected
using western blot analysis. Compared with the control group, EZH2
protein expression significantly decreased at 8, 12 and 24 h in the
5 µM DZNep group, and P-β-catenin significantly decreased at 2, 8,
12 and 24 h in the 5 µM DZNep group. In addition, N-P-β-catenin
expression significantly increased after 2, 8 and 24 h in the 5 µM
DZNep group compared with the control group. Treatment with 3, 5
and 10 µM DZNep for 24 h significantly decreased the EZH2 protein
expression level. In addition, 5 and 10 µM DZNep treatment
decreased the total β-catenin protein level (Fig. 6). These results suggested that DZNep
inhibited EZH2 protein expression and further inhibited the
phosphorylation of β-catenin.
 | Figure 6.DZNep treatment reduces EZH2 protein
expression, decreases P-β-catenin protein expression and increases
N-P-β-catenin protein expression. (A) Protein expression levels of
EZH2, P-β-catenin and N-P-β-catenin following 5 µM DZNep treatment
for 2, 8, 12 and 24 h, and the protein expression levels of EZH2
and total-β-catenin following 3, 5 and 10 µM DZNep treatment for 24
h were detected using western blot analysis. The internal control
was β-actin. (B) EZH2 expression significantly increased after 8,
12 and 24 h DZNep treatment, and the differences were statistically
significant. *P<0.05 and **P<0.01 vs. respective control.
Inhibition of EZH2 protein expression by DZNep was time-dependent.
(C) Following 5 µM DZNep treatment in BGC-823 cells for 2, 8, 12
and 24 h, P-β-catenin significantly decreased at all time points,
and the differences were statistically significant. **P<0.01 vs.
respective control. (D) N-P-β-catenin significantly increased after
2, 8 and 24 h of DZNep treatment, and the differences were
statistically significant. *P<0.05 and **P<0.01 vs. control.
(E) Following 3, 5 and 10 µM DZNep treatment for 24 h, the EZH2
protein expression levels decreased in a dose-dependent manner.
*P<0.05 and **P<0.01 vs. control. (F) Following 3, 5 and 10
µM DZNep treatment for 24 h, the total-β-catenin protein expression
levels decreased in a dose-dependent manner; following 5 and 10 µM
DZNep treatment in BGC-823 cells, total-β-catenin protein
expression was significantly decreased. *P<0.05 and **P<0.01
vs. control. DZNep, 3-deazaneplanocin A; EZH2, enhancer of zeste
homolog 2; N-P-β-catenin, non-phosphorylated β-catenin;
P-β-catenin, phosphorylated β-catenin. |
Discussion
The abnormal expression of the epigenetic regulatory
factors PcG proteins is closely associated with the development and
progression of tumors. Based on their different functions, PcG
proteins are divided into two protein complexes, PRC1 and PRC2.
PRC2 is a highly conserved histone methyltransferase that functions
on the K27 lysine site of histone H3 (H3K27me3). The core
component, EZH2, has histone methyltransferase activity and may
catalyse the methylation of the H3K27me3 histone to inhibit target
gene expression, thus resulting in tumor development (4,20,21). In
gastric cancer cells, EZH2 is involved in the regulation of cell
cycle-associated proteins to promote the proliferation and
metastasis of gastric cancer cells (22). A previous study reported that
clinical trials evaluating EZH2-targeting agents, including DZNep,
should consider stratifying patients with gastric cancer by their
TP53 genomic status (23). EZH2 is
an important target in the study of gastric cancer invasion and
metastasis. DZNep may reduce EZH2 protein expression levels, reduce
H3K27me3 levels, activate PRC2 target genes and may specifically
induce tumor cell apoptosis (24).
Abnormal proliferation is a common feature of
malignant tumors. DZNep has been confirmed to be able to inhibit
the proliferation of Eca109 oesophageal squamous cell carcinoma
cells and HTC116 colorectal cancer cells (25,26). The
present study used the poorly differentiated human BGC-823 gastric
adenocarcinoma cell line. The CCK-8 experiment results demonstrated
that DZNep inhibited BGC-823 cell proliferation. The colony
formation assay results demonstrated that DZNep reduced the number
of colonies formed in BGC-823 cells. The flow cytometry results
demonstrated that DZNep promoted BGC-823 cell apoptosis. Notably,
BGC-823 cells exhibited morphological alterations following DZNep
treatment. The cell edges became sharp, and the margins exhibited
thread-like and circular structures. These morphological
alterations may allow cells to better attach onto the culture
plate. Therefore, it was speculated that DZNep may attenuate the
invasion and metastasis abilities of cells. Subsequently,
wound-healing experiments and Transwell invasion experiments were
performed to investigate this. The results of these assays
demonstrated that DZNep attenuated the wound healing activity of
BGC-823 cells and reduced the number of cells that passed through
the membrane, suggesting that DZNep inhibited the invasion and
metastasis of human BGC-823 gastric cancer cells.
To further study the mechanisms, it was considered
that during the process of tumor progression, the blood vessel
network cannot be rapidly established due to the rapid
proliferation of tumor cells, thus causing a reduction in the
oxygen content in the microenvironment, a lack of nutritional
substances and an accumulation of acidic substances (27,28). In
a hypoxic microenvironment, tumor cells are prone to develop
invasion and metastasis abilities and activate drug-resistance
genes to increase their adaptive ability (29,30).
Hif-1 is a nuclear protein with transcriptional activity. Under the
hypoxic growth environment of tumors, Hif-1 can promote tumor cell
proliferation and angiogenesis to cause tumor recurrence and
metastasis at the late stages and attenuate the sensitivity of
tumors to chemotherapy and radiotherapy (31). Hif-1 is a heterodimeric transcription
factor activated by hypoxia composed of two different subunits,
HIF-1α and ARNT (aryl receptor nuclear translocator) (32). When activated, HIF-1 mediates
differential expression of genes (32). The present study is the first, to the
best of our knowledge, to detect the alterations in the mRNA and
protein expression levels of Hif-1α following DZNep-treatment. The
results demonstrated that the relative mRNA expression level of
Hif-1α significantly decreased following 1 and 5 µM DZNep-treatment
for 4 h, and 5 µM DZNep-treatment for 24 h. The protein expression
level of Hif-1α significantly decreased following 3, 5 and 10 µM
DZNep-treatment for 2, 12 and 24 h. These results suggest that
DZNep inhibited the Hif-1α protein expression level in a dose- and
time-dependent manner.
Hif-1α in the microenvironment of solid tumors may
induce high expression of EZH2. Chang et al (33) reported that in breast tumor
initiating cells (BTICs), the transactivation function mediated by
Hif-1α significantly upregulates EZH2 expression, whereas high EZH2
expression causes BTIC proliferation and promotes their
progression. The conserved HRE sequence that interacts with Hif-1α
was identified in the promoter region of EZH2, which confirmed that
hypoxia induces Hif-1α to bind to this HRE region to activate EZH2
transcription, and to promote breast cancer growth (34). DZNep was the first discovered
inhibitor of the enzyme activity of EZH2. Additionally, DZNep
reduced the protein expression levels of EZH2 (35). The present study demonstrated that
treatment with 3, 5 and 10 µM DZNep significantly reduced the
protein expression level of EZH2. In addition, 5 µM DZNep
significantly inhibited EZH2 expression after 8, 12 and 24 h. These
results suggest that DZNep inhibited the protein expression of EZH2
in a dose- and time-dependent manner.
It has been demonstrated that EZH2 may silence a
number of differentiation-associated factors, including the Gate,
Sox and Pax transcription factor families, in addition to the
components of the Wnt/β-catenin and transforming growth factor β
signalling pathways, to cause tumor development (20,21,36).
Zhang et al (37) revealed
that β-catenin signalling and EZH2 have interactive regulation in
glioblastoma. The growth inhibitory effect of EZH2 small
interference RNA was reversed by dominant active β-catenin,
indicating that EZH2 activates Wnt signalling to promote gastric
carcinogenesis. EZH2-promoted cell growth and activation of Wnt
signalling were attenuated by ectopic CXXC finger protein 4 (CXXC4)
expression. EZH2 promotes the activation of Wnt signalling in
gastric carcinogenesis via the downregulation of CXXC4 expression.
CXXC4 is a novel potential tumor suppressor directly regulated by
EZH2. Additionally, CXXC4 is a negative regulator of Wnt/β-catenin
signalling (38). Inhibition of the
EZH2 activity using RNA interference or small molecule inhibitors
downregulates β-catenin expression. An in vivo study has
confirmed that EZH2-knockout downregulates β-catenin expression to
inhibit tumor growth (37). The
mechanism is as follows: When the canonical Wnt signalling pathway
is activated, secreted extracellular Wnt interacts with the
transmembrane receptors LDL receptor related protein (LRP)5/6 and
Frizzled to form a complex to cause the phosphorylation of the
intracellular fragment of LRP5/6 and formation of the
Wnt-Axin-adenomatosis polyposis coli-β-catenin complex to block
β-catenin phosphorylation/degradation; free β-catenin enters into
the cell nucleus and interacts with TCF/LEF to initiate the
transcription of downstream genes, achieve sustained cell
proliferation and induce tumors (39). Therefore, the present study used
western blot analysis to detect β-catenin, P-β-catenin and
N-P-β-catenin in BGC-823 cells following 3, 5 and 10 µM
DZNep-treatment for 24 h. The results demonstrated that when EZH2
was inhibited, β-catenin expression also decreased. Additionally,
cells were treated with DZNep for 2, 8, 12 and 24 h and expression
levels of P-β-catenin and N-P-β-catenin were detected. The results
demonstrated that P-β-catenin expression levels significantly
decreased and NP-β-catenin expression levels significantly
increased following DZNep treatment. It was speculated that DZNep
may inhibit the Wnt signalling pathway to some extent to cause the
degradation of accumulated P-β-catenin and an increase in
N-P-β-catenin. Therefore, through the reduction of Hif-1α levels,
DZNep may inhibit BGC-823 cell proliferation, promote apoptosis,
reduce colony formation, and attenuate the invasion and metastasis
behaviours of BGC-823 cells. The underlying action mechanism is
likely to involve the inhibition of EZH2 to further block β-catenin
phosphorylation.
In conclusion, the data presented in the current
study indicated a positive relationship between EZH2 and Hif-1α and
between EZH2 and Wnt/β-catenin signalling molecules. DZNep may
inhibit the proliferation and invasion of gastric cancer cells by
inhibiting EZH2 and Hif-1α expression and Wnt/β-catenin signalling.
These results provide a theoretical basis for the application of
DZNep in clinical trials.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 81460433) and the
Natural Science Foundation of Ningxia Province in China (grant nos.
NZ14282 and NZ14080).
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
RH performed the experiments and data analysis, and
contributed to the writing of the manuscript. XJ performed the
RT-qPCR experiments and revised the manuscript. HY performed the
RT-qPCR experiments. FW performed the western blotting experiments
and prepared manuscript content. YG and XC performed the data
analysis and were involved in the technical aspects of the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wanebo HJ, Kennedy BJ, Chmiel J, Steele G
Jr, Winchester D and Osteen R: Cancer of the stomach. A patient
care study by the American College of Surgeons. Ann Surg.
218:583–592. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Koeda K, Nishizuka S and Wakabayashi G:
Minimally invasive surgery for gastric cancer: The future standard
of care. World J Surg. 35:1469–1477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Piunti A and Pasini D: Epigenetic factors
in cancer development: Polycomb group proteins. Future Oncol.
7:57–75. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gergely JE, Dorsey AE, Dimri GP and Dimri
M: Timosaponin A-III inhibits oncogenic phenotype via regulation of
PcG protein BMI1 in breast cancer cells. Mol Carcinog. 57:831–841.
2018. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Margueron R and Reinberg D: The polycomb
complex PRC2 and its mark in life. Nature. 469:343–349. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suvà ML, Riggi N, Janiszewska M,
Radovanovic I, Provero P, Stehle JC, Baumer K, Le Bitoux MA, Marino
D, Cironi L, et al: EZH2 is essential for glioblastoma cancer stem
cell maintenance. Cancer Res. 69:9211–9218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Riquelme E, Behrens C, Lin HY, Simon G,
Papadimitrakopoulou V, Izzo J, Moran C, Kalhor N, Lee JJ, Minna JD
and Wistuba II: Modulation of EZH2 expression by MEK-ERK or
PI3K-AKT signaling in lung cancer is dictated by different KRAS
oncogene mutations. Cancer Res. 76:675–685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jung HY, Jun S, Lee M, Kim HC, Wang X, Ji
H, McCrea PD and Park JI: PAF and EZH2 induce Wnt/β-catenin
signaling hyperactivation. Mol Cell. 52:193–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miranda TB, Cortez CC, Yoo CB, Liang G,
Abe M, Kelly TK, Marquez VE and Jones PA: DZNep is a global histone
methylation inhibitor that reactivates developmental genes not
silenced by DNA methylation. Mol Cancer Ther. 8:1579–1588. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mochizuki D, Misawa Y, Kawasaki H, Imai A,
Endo S, Mima M, Yamada S, Nakagawa T, Kanazawa T and Misawa K:
Aberrant epigenetic regulation in head and neck cancer due to
distinct EZH2 overexpression and dna hypermethylation. Int J Mol
Sci. 19(pii): E37072018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yao Y, Hu H, Yang Y, Zhou G, Shang Z, Yang
X, Sun K, Zhan S, Yu Z, Li P, et al: Downregulation of enhancer of
zeste homolog 2 (EZH2) is essential for the induction of autophagy
and apoptosis in colorectal cancer cells. Genes (Basel). 7:832016.
View Article : Google Scholar
|
|
13
|
Wei FZ, Cao Z, Wang X, Wang H, Cai MY, Li
T, Hattori N, Wang D, Du Y, Song B, et al: Epigenetic regulation of
autophagy by the methyltransferase EZH2 through an MTOR-dependent
pathway. Autophagy. 11:2309–2322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hebert M, Potin S, Sebbagh M, Bertoglio J,
Bréard J and Hamelin J: Rho-ROCK-dependent ezrin-radixin-moesin
phosphorylation regulates Fas-mediated apoptosis in Jurkat cells. J
Immunol. 181:5963–5973. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Manalo DJ, Rowan A, Lavoie T, Natarajan L,
Kelly BD, Ye SQ, Garcia JG and Semenza GL: Transcriptional
regulation of vascular endothelial cell responses to hypoxia by
HIF-1. Blood. 105:659–669. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qi C, Zhang J, Chen X, Wan J, Wang J,
Zhang P and Liu Y: Hypoxia stimulates neural stem cell
proliferation by increasing HIF-1α expression and activating
Wnt/β-catenin signaling. Cell Mol Biol (Noisy-le-grand). 63:12–19.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pang B, Zheng XR, Tian JX, Gao TH, Gu GY,
Zhang R, Fu YB, Pang Q, Li XG and Liu Q: EZH2 promotes metabolic
reprogramming in glioblastomas through epigenetic repression of
EAF2-HIF1α signaling. Oncotarget. 7:45134–45143. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Q, Cai J, Wang Q, Wang Y, Liu M, Yang
J, Zhou J, Kang C, Li M and Jiang C: Long noncoding RNA NEAT1,
regulated by the EGFR pathway, contributes to glioblastoma
progression through the WNT/β-catenin pathway by scaffolding EZH2.
Clin Cancer Res. 24:684–695. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Panousis D, Patsouris E, Lagoudianakis E,
Pappas A, Kyriakidou V, Voulgaris Z, Xepapadakis G, Manouras A,
Athanassiadou AM and Athanassiadou P: The value of TOP2A, EZH2 and
paxillin expression as markers of aggressive breast cancer:
relationship with other prognostic factors. Eur J Gynaecol Oncol.
32:156–159. 2011.PubMed/NCBI
|
|
21
|
Choi JH, Song YS, Yoon JS, Song KW and Lee
YY: Enhancer of zeste homolog 2 expression is associated with tumor
cell proliferation and metastasis in gastric cancer. Apmis.
118:196–202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng LL, Itahana Y, Lei ZD, Chia NY, Wu
Y, Yu Y, Zhang SL, Thike AA, Pandey A, Rozen S, et al: TP53 genomic
status regulates sensitivity of gastric cancer cells to the histone
methylation inhibitor 3-deazaneplanocin A (DZNep). Clin Cancer Res.
18:4201–4212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang CJ, Yang JY, Xia W, Chen CT, Xie X,
Chao CH, Woodward WA, Hsu JM, Hortobagyi GN and Hung MC: EZH2
promotes expansion of breast tumor initiating cells through
activation of RAF1-catenin signaling. Cancer Cell. 19:86–100. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Glazer RI, Hartman KD, Knode MC, Richard
MM, Chiang PK, Tseng CK and Marquez VE: 3-Deazaneplanocin: A new
and potent inhibitor of S-adenosylhomocysteine hydrolase and its
effects on human promyelocytic leukemia cell line HL-60. Biochem
Biophys Res Commun. 135:688–694. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu S, Liu F, Huang W, Gu L, Meng L, Ju Y,
Wu Y, Li J, Liu L and Sang M: MAGE-A11 is activated through
TFCP2/ZEB1 binding sites de-methylation as well as histone
modification and facilitates ESCC tumor growth. Oncotarget.
9:3365–3378. 2017.PubMed/NCBI
|
|
26
|
Sha M, Mao G, Wang G, Chen Y, Wu X and
Wang Z: DZNep inhibits the proliferation of colon cancer HCT116
cells by inducing senescence and apoptosis. Acta Pharm Sin B.
5:188–193. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang Y, Lin D and Taniguchi CM: Hypoxia
inducible factor (HIF) in the tumor microenvironment: Friend or
foe? Sci China Life Sci. 60:1114–1124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martin JD, Fukumura D, Duda DG, Boucher Y
and Jain RK: Reengineering the tumor microenvironment to alleviate
hypoxia and overcome cancer heterogeneity. Cold Spring Harb
Perspect Med. 6(pii): a0270942016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li DW, Dong P, Wang F, Chen XW, Xu CZ and
Zhou L: Hypoxia induced multidrug resistance of laryngeal cancer
cells via hypoxia-inducible factor-1α. Asian Pac J Cancer Prev.
14:4853–4858. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Borsi E, Terragna C, Brioli A, Tacchetti
P, Martello M and Cavo M: Therapeutic targeting of hypoxia and
hypoxia-inducible factor 1 alpha in multiple myeloma. Transl Res.
165:641–650. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lou JJ, Chua YL, Chew EH, Gao J, Bushell M
and Hagen T: Inhibition of hypoxia-inducible factor-1alpha protein
synthesis by DNA damage inducing agents. PLoS One. 5:e105222010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Michel G, Minet E, Ernest I, Durant F,
Remacle J and Michiels C: Molecular modeling of the
hypoxia-inducible factor-1 (HIF-1). Theor Chem Acc. 101:51–56.
1999. View Article : Google Scholar
|
|
33
|
Chang CJ, Yang JY, Xia W, Chen CT, Xie X,
Chao CH, Woodward WA, Hsu JM, Hortobagyi GN and Hung MC: EZH2
promotes expansion of breast tumor initiating cells through
activation of RAF1-β-catenin signaling. Cancer Cell. 19:86–100.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mahara S, Lee PL, Feng M, Tergaonkar V,
Chng WJ and Yu Q: HIFI-α activation underlies a functional switch
in the paradoxical role of Ezh2/PRC2 in breast cancer. Proc Natl
Acad Sci USA. 113:E3735–E3744. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan J, Yang X, Zhuang L, Jiang X, Chen W,
Lee PL, Karuturi RK, Tan PB, Liu ET and Yu Q: Pharmacologic
disruption of Polycomb-repressive complex 2-mediated gene
repression selectively induces apoptosis in cancer cells. Genes
Dev. 21:1050–1063. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsang DP and Cheng AS: Epigenetic
regulation of signaling pathways in cancer: Role of the histone
methyltransferase EZH2. J Gastroenterol Hepatol. 26:19–27. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Chen L, Han L, Shi Z, Zhang J, Pu
P and Kang C: EZH2 is a negative prognostic factor and exhibits
pro-oncogenic activity in glioblastoma. Cancer Lett. 356:929–936.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lu H, Sun J, Wang F, Feng L, Ma Y, Shen Q,
Jiang Z, Sun X, Wang X and Jin H: Enhancer of zeste homolog 2
activates Wnt signaling through downregulating CXXC finger protein
4. Cell Death Dis. 4:e7762013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Minde DP, Radli M, Forneris F, Maurice MM
and Rüdiger SG: Large extent of disorder in Adenomatous Polyposis
Coli offers a strategy to guard Wnt signalling against point
mutations. PLoS One. 8:e772572013. View Article : Google Scholar : PubMed/NCBI
|