Introduction
Hepatocellular carcinoma (HCC) is the most common
primary liver cancer with a high recurrence rate and a poor
prognosis. Annually, among the newly diagnosed HCC cases, those in
China account for approximately >50% of all global cases
(1). Although great advances have
been made in traditional hepatectomy and other comprehensive
therapeutic strategies for HCC, the results remain unsatisfactory
(2). Early recurrence and metastasis
are the leading causes of poor patient outcome; therefore, in order
to improve HCC outcome, the investigation of novel molecular
mechanisms underlying HCC recurrence and metastasis is urgently
required.
Insulin-like growth factor II mRNA-binding protein 3
(IGF2BP3/IMP3), a member of the IGF2BP family, has been shown to
exhibit low or undetectable protein levels in normal adult tissues,
whereas higher expression levels are observed in malignant tumors
(3–5). Moreover, a number of studies have found
that the overexpression of IGF2BP3 is associated with poor outcome
in various types of cancer, such as breast cancer, cervical cancer,
renal cell carcinoma, neuroblastoma and HCC (6–13).
Pre-clinical studies have demonstrated that IGF2BP3 can promote
cancer cell motility, invasiveness and migration (14–17).
Functional studies have also demonstrated that IGF2BP3, an RNA
binding protein, plays a critical physiological role in regulating
RNA splicing, stability, localization, modification and translation
(18,19). Studies of multiple cancer types have
demonstrated that a set of cancer-related pathways, particularly
those involved in invasion, may be mediated by IGF2BP3 (13,16). The
results of a mechanical study also demonstrated that IGF2BP3
facilitated partial invasion-promoted gene expression and decreased
the expression of certain tumor-suppressing genes (20,21). For
example, IGF2BP3 has been shown to function as a cytoplasmic
‘safehouse’ and to prevent the mRNA-directed decay of oncogene
Hmga2 during tumorigenesis (22).
However, IGF2BP3 can also influence the mRNA degradation of
tumor-suppressor genes by enhancing micro(mi)RNA-mRNA interactions
(23). To date, the detailed
molecular pathways underlying the effects of IGF2BP3 on HCC cell
invasion ability remain unknown.
Zonula occludens-1 (ZO-1) is a member of the zona
occludens protein family, which is associated with the function of
tight and adherens junctions, and may serve critical roles in HCC
progression and metastasis by promoting cellular migration and
invasion (24,25). Previous studies have demonstrated
that ZO-1 expression is associated with the prognosis of lung
cancer and HCC (26,27). In a previous in vitro study,
increasing ZO-1 expression significantly decreased cellular
invasiveness; by contrast, the knockdown of ZO-1 markedly increased
the invasive capacity of HCC cell lines. The result of IGF2BP3 RIP
(RNP immunoprecipitation) sequencing data from pancreatic ductal
adenocarcinoma cells also demonstrated that IGF2BP3 could bind ZO-1
mRNA (23). However, whether IGF2BP3
can bind and mediate ZO-1 expression in HCC invasion remains to be
elucidated.
In the present study, the potential association
between IGF2BP3 expression and poor prognosis in patients with HCC
was determined by analyzing data from the Gene Expression Omnibus
(GEO), the European Genome-phenome Archive (EGA) and The Cancer
Genome Atlas (TCGA). In in vitro experiments, the results
demonstrate that the knockdown of IGF2BP3 deceases cell
invasiveness by increasing ZO-1 expression; conversely, the
overexpression of IGF2BP3 increased invasion capacity by decreasing
ZO-1 expression in HCC cell lines. Mechanistic analyses revealed
that IGF2BP3 suppressed ZO-1 expression by enhancing
miR191-5p-induced ZO-1 mRNA silencing. Taken together, the findings
of the present study suggest that directly targeting IGF2BP3 or
miR191-5p may improve ZO-1 expression and suppress cell
invasiveness.
Materials and methods
Bioinformatics data
Liver cancer RNA-seq data (http://kmplot.com/analysis/index.php?p=background)
were extracted from the GEO, the EGA and TCGA databases. IGF2BP3
expression data associated with overall survival (OS) and
relapse-free survival (RFS) were analyzed using the Kaplan-Meier
plotter online tool (http://kmplot.com).
Cell culture
The Huh-7 and HA22T liver cancer cell lines (both
adult hepatocellular carcinoma) were purchased from the American
Type Culture Collection (28–30).
Cells were maintained in Dulbecco's modified Eagle's medium
(Invitrogen; Thermo Fisher Scientific Inc.) supplemented with 10%
fetal bovine serum (Thermo Fisher Scientific, Inc.), 1% glutamine
and 1% penicillin/streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) (31,32), and cultured at 37°C in a humidified
incubator [5% (v/v) CO2]. PCR detection for mycoplasma
contamination yielded negative results.
Reagents and materials
Mouse anti-ZO-1 (1:1,000; sc-33725), mouse
anti-GAPDH (1:1,000; sc-47724) and mouse anti-IGF2BP3 (1:1,000;
sc-365641) antibodies were purchased from Santa Cruz Biotechnology,
Inc. Rabbit polyclonal argonaute-2 antibody (1:100; ab32381) was
purchased from Abcam. Anti-rabbit/mouse secondary antibodies
(1:5,000; A10547 and A10668, respectively) were purchased from
Invitrogen; Thermo Fisher Scientific, Inc. Rabbit IgG (1:100;
sc-69786) was also obtained from Santa Cruz Biotechnology, Inc. The
miRNA-191-5p inhibitor was purchased from Shanghai GenePharma Co.,
Ltd.
Lentiviral expression plasmids and
virus infection
The lentivirus system and standard calcium chloride
transfection method were applied to generate the virus. The
pWPI–IGF2BP3, pLKO.1 pLKO.1-shIGF2BP3#1 or pLKO.1-shIGF2BP3#2,
pWPI-ZO-1 and pLKO.1-oemiR191-5p/pLKO.1/pLKO.1-oemiR429, or the
corresponding empty control plasmids (EMD Millipore) were
co-transfected into 293 cells with the pMD2G envelope plasmid and
psPAX2 packaging plasmid (12259 and 12260, respectively; both
Addgene, Inc.), using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.). After 8 h, the medium was replaced
with fresh warm medium. The cells were then cultured in a virus
room incubator for virus generation as previously decreased
(33). After 48 h, the
virus-containing supernatants were harvested, used immediately or
stored at −80°C for later use. Huh-7 or HA22T cells were seeded
into a 6-well plate (~1×106/well) and infected with
virus (MOI=2). Green fluorescence protein was used to monitor gene
overexpression, and 1 µg/ml puromycin was to select the cells with
gene knockdown or miR-191-5p overexpression. In order to
downregulate miR-191-5p expression, ~1×106 cells/well
were seeded into a 6-well plate, and transfected with 50 pmol
miR-191-5p inhibitor or NC inhibitor using
Lipofectamine® 3000, and incubated for 24 h. The
expression level of miR-191-5p was monitored by reverse
transcription-quantitative (RT-q) PCR to determine whether
upregulation or downregulation was successful. The shRNA sequences
were as follows: shIGF2BP3#1 targeting sequence,
5′-GCAGGAATTGACGCTGTATAA-3′; shIGF2BP3#2 targeting sequence,
5′-TCTGCGGCTTGTAAGTCTAT-3′; miR-191-5p inhibitor,
5′-CAGCUGCUUUUGGGAUUCCGUUG-3′; and NC inhibitor,
5′-CAGUACUUUUGUGUAGUACAA-3′.
RNA extraction and RT-qPCR
analysis
Total RNA was extracted from Huh-7 and HA22T cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA (1 µg) was reverse transcribed into
cDNA using Superscript III transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.) at 42°C for 60 min and 95°C for 5 min. qPCR was
conducted using a Bio-Rad CFX96 system (Applied Biosystems) with
SYBR-Green to detect the mRNA expression level of a gene of
interest using the following thermocycling conditions: 95°C for 3
min, followed by 35 cycles at 95°C for 15 sec, 60°C for 30 sec and
68°C for 1 min. Expression levels were normalized to the GAPDH
levels using the 2−ΔΔCq method (34). The primer sequences were as follows:
miR-191-5p forward, 5′-CAACGGAATCCCAAAAGCAGCTG-3′ and reverse,
5′-TGTCGTGGAGTCGGC-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′; IGF2BP3 forward,
5′-ACTGCACGGGAAACCCATAG-3′ and reverse, 5′-ACTATCCAGCACCTCCCACT-3′;
ZO-1 forward, 5′-GTGGGTAACGCCATCCTCTG-3′ and reverse,
5′-TCCGGGATTTCACCAGTGTG-3′; and GAPDH forward,
5′-TGCACCACCAACTGCTTAGC-3′ and reverse,
5′-GGCATGGACTGTGGTCATGAG-3′. All primers were purchased from
Integrated DNA Technologies, Inc.
Invasion assay
As previously described, a Transwell assay was used
to determine the invasive capacity of HCC cells. Following
transfection, 5×104 cells/well were seeded with
serum-free medium into the upper chambers of the Transwell inserts,
which had been pre-coated with diluted Matrigel (Corning Inc.) at
37°C for 4 h. Subsequently, 750 µl media supplemented with 10% FBS
was pipetted into the lower chambers, and the cells were incubated
at 37°C [5% (v/v) CO2] for 18 h. After being fixed with
methanol at room temperature for 20 min, the invasive cells were
stained with 0.1% (w/v) crystal violet at room temperature for 15
min. The average number of cells from three randomly selected
microscopic areas using an inverted light microscope
(magnification, ×100; Olympus Corporation) were counted and for
quantification, and the number of invasive cells was calculated
using ImageJ software 1.8 (National Institutes of Health).
Western blot analysis
Cells were lysed to extract total proteins using
RIPA buffer (Wuhan Boster Biological Technology, Ltd.,). The
protein concentration was measured using a BCA Protein assay kit
(ab102536; Abcam) and ultraviolet spectrophotometry. Proteins (30
µg/well) were separated using 6–10% SDS/PAGE gels and then
transferred onto PVDF membranes (EMD Millipore). After blocking
with 5% milk at room temperature for 1.5 h, the membranes were
incubated with primary antibodies at 4°C overnight, and
subsequently with HRP-conjugated secondary antibodies (detail in
the Reagents and materials part) at room temperature for 2 h. An
enhanced chemiluminescent substrate (35055; Thermo Fisher
Scientific, Inc.) was then used to detect and visualize the
signals. The relative expression levels were quantified using the
Image Lab soft 4.1 (Bio-Rad Laboratories, Inc.) with GAPDH as the
internal reference.
RNA immunoprecipitation (RIP)
Following transfection, cells were lysed in ice-cold
lysis buffer supplemented with RNase inhibitor. Following
centrifugation, 500 µl of the supernatant was clarified using
protein A/G beads for 1 h and incubated with an argonaute-2
antibody at 4°C overnight. RNA was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol and subjected to
RT-qPCR analysis.
Luciferase reporter assay
A 500 bp fragment of the ZO-1 3′ untranslated region
(UTR), with wild-type or mutant IGF2BP3 or miRNA-responsive
elements, was cloned into the psiCHECK-2 vector (Promega
Corporation) downstream of the Renilla luciferase open reading
frame. HA22T and Huh-7 cells were seeded into 24-well plates, and
the cDNA was transfected with Lipofectamine® 3000
transfection reagent according to the manufacturer's instructions.
PRL-TK was used as an internal control that served as the baseline
control response. Luciferase activity was measured at 48 h
post-transfection using a dual-luciferase reporter assay (Promega
Corporation) according to the manufacturer's protocol. The relative
luciferase activity was normalized to Renilla luciferase
activity.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 23.0 (IBM Inc.). The Mann-Whitney U test was
conducted to compare two groups of continuous data. The
Kruskal-Wallis test with the Dunn-Bonferroni post hoc test was
applied to multiple groups of continuous data. All cellular
experiments were conducted in three technical replicates.
Quantitation Results are presented as the mean ± SD. Statistical
significance for cell experiments was determined using the
independent-sample t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Higher IGF2BP3 expression may indicate
a lower survival rate in patients with HCC
The datasets, which included gene expression
associated with OS and RFS, were retrieved from the GEO, EGA and
TCGA, and were analyzed online (http://kmplot.com). The results revealed that patients
with a high IGF2BP3 expression level (50% of the top IGF2BP3
expression samples) exhibited lower OS and RFS rates than those
with low IGF2BP3 expression (50% of the lower IGF2BP3 expression
samples) (Fig. 1A and B), suggesting
that IGF2BP3 may be involved in HCC progression. Furthermore,
IGF2BP3 expression was determined in patients at different grades
or stages of HCC using data from TCGA database. The results
demonstrated that patients with higher grades or stages of the
disease possessed higher IGF2BP3 expression levels (Fig. 1C and D). Taken together, these data
are consistent with those previously published, which suggest that
IGF2BP3 promotes HCC progression (13).
IGF2BP3 may promote HCC cell
invasiveness by suppressing ZO-1 expression
To further demonstrate the function of IGF2BP3 in
HCC progression, its effects on cell invasiveness were assessed by
IGF2BP3 shRNA knockdown or overexpression with IGF2BP3-cDNA in
HA22T and Huh-7 cells, respectively. As shown in Fig. 2A and B, IGF2BP3 knockdown with
IGF2BP3-shRNA#1 markedly decreased the invasive ability of HA22T
cells. To avoid the off-target effect of IGF2BP3-shRNA, a second
shIGF2BP3#2 plasmid was constructed to repeat the experiment, and
the results were consistent with those obtained with
IGF2BP3-shRNA#1 (Fig. S1A and B).
By contrast, the overexpression of IGF2BP3 markedly increased the
invasive capacity of Huh-7 cells (Fig.
S1C and D).
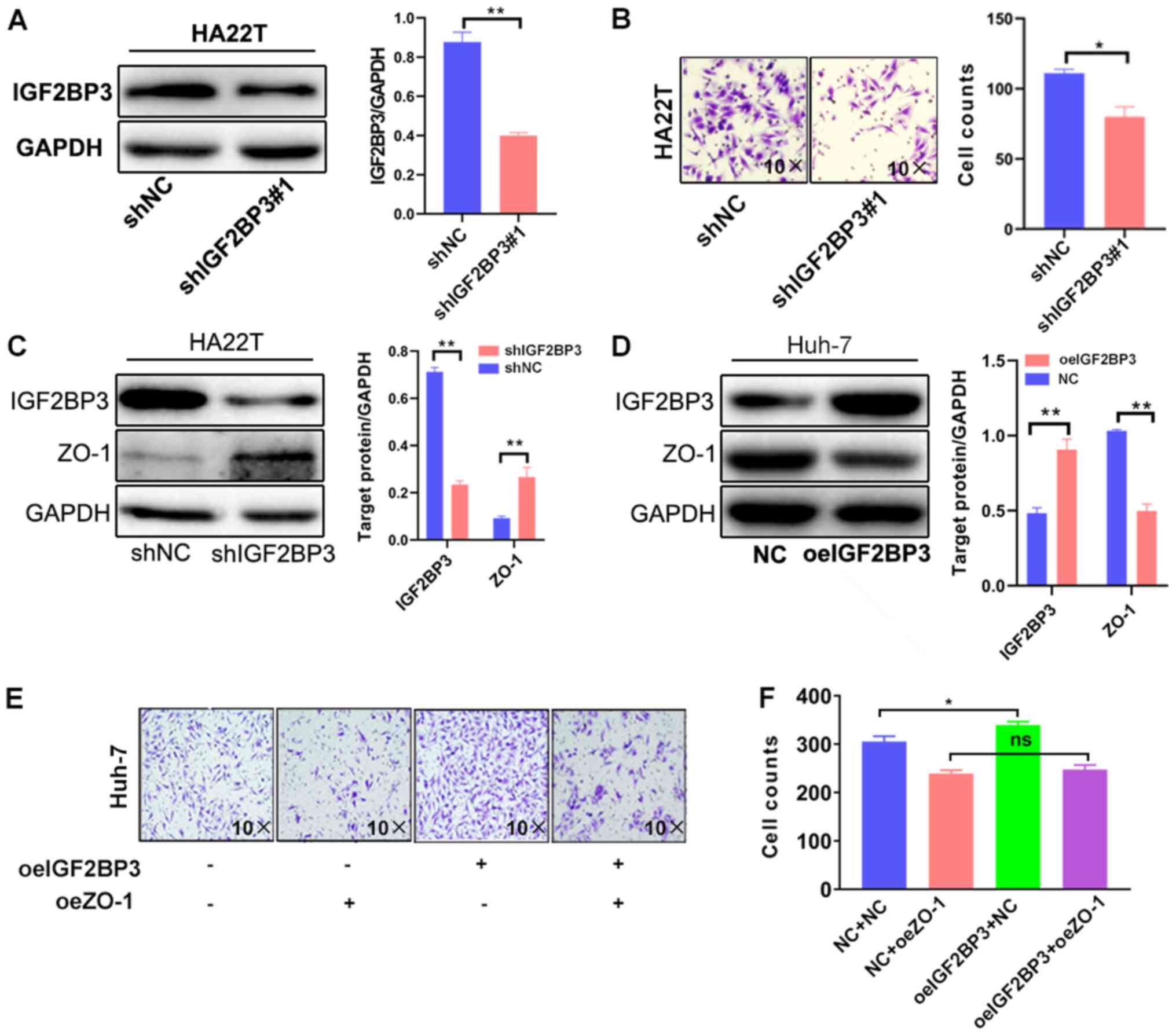 | Figure 2.IGF2BP3 promotes HCC cell
invasiveness by suppressing ZO-1 expression. (A) Western blot
analysis of IGF2BP3 knockdown in HA22T cells. (B) Transwell
invasion assays were performed using HA22T cells transfected with
pLKO and IGF2BP3-shRNA. The invasive cells were counted and
averaged from 3 randomly selected microscopic fields
(magnification, ×10). Each sample was run in triplicate. Western
blot analysis was performed on (C) HA22T cells transfected with
pLKO and IGF2BP3-shRNA and (D) Huh-7 cells transfected with pWPI
and IGF2BP3-cDNA. (E) Transwell invasion assays were performed
using Huh-7 cells transfected with pWPI+pWPI, pWPI+oeZO-1,
oeIGF2BP3+pWPI or oeIGF2BP3+oeZO-1. (F) All quantifications are
shown, and are presented as the mean ± SD. *P≤0.05 and **P≤0.01.
ns, not significant; IGF2BP3, insulin-like growth factor II
mRNA-binding protein 3; HCC, hepatocellular carcinoma; ZO-1, zonula
occludens-1; sh, short hairpin (RNA); oe, overexpression; NC,
negative control. |
To elucidate the mechanisms responsible for the
effects of IGF2BP3 on HCC cell invasiveness, the expression of ZO-1
was examined, a protein which is associated with cellular
invasiveness in a variety of cancers, such as breast, pancreatic
and lung cancer (27,35,36).
IGF2BP3 knockdown in HA22T cells resulted in increased expression
of ZO-1 (Fig. 2C). Conversely, the
overexpression of IGF2BP3 in Huh-7 cells decreased the expression
of ZO-1 (Fig. 2D). To verify whether
increased levels of ZO-1 could reduce cellular invasion capacity, a
lentivirus system was used to overexpress ZO-1 and western blot
analysis was used confirm expression in Huh-7 cells (Fig. S1E). The results of Transwell assay
revealed that ZO-1 overexpression markedly decreased Huh-7 cell
invasiveness (Fig. 2E). These
results suggest that IGF2BP3 affects HCC cell invasiveness by
altering ZO-1 expression.
A reverse assay was then employed to examine the
roles of ZO-1 in the IGF2BP3-induced increase in HCC cell
invasiveness. The results revealed that the ZO-1 overexpression
partially blocked the IGF2BP3-induced increase in HCC cell
invasiveness (Fig. 2E-F).
Collectively, the results shown in Fig.
2A-F demonstrate that IGF2BP3 may increase HCC cell
invasiveness by suppressing ZO-1 expression.
Mechanistic analysis of the mechanisms
through which IGF2BP3 suppresses ZO-1 expression: By synergizing
with miR191-5p through the RNA-induced silencing complex
(RISC)
The present study then aimed to reveal the
mechanisms responsible for the suppressive effects of IGF2BP3 on
ZO-1 expression. It has been previously reported that IGF2BP3 acts
as an RNA-binding protein by recognizing specific motifs to
determine targeted RNA fate (37),
and that IGF2BP3 can modulate invasion-associated gene transcripts
by promoting Ago2-mRNA interactions (23). Therefore, the potential regulation of
ZO-1 by Ago2-mRNA interactions was investigated by detecting ZO-1
mRNA in the Ago2 complex using an RIP assay. ZO-1 mRNA
co-precipitation with Ago2 was decreased in the IGF2BP3-depleted
cells (Fig. 3A). Subsequently, to
determine which miRNAs mediated ZO-1 expression through the
IGF2BP3-Ago2 complex, online miRNA software (http://www.targetscan.org) was used to screen six
potential candidates and detect miRNA expression from the
Ago2-antibody pull-down complex between the negative control (NC)
cells and IGF2BP3-cDNA cells. The results revealed that the
miR-191-5p and miR429 levels were significantly higher in the
IGF2BP3-overexpressing cells than in the NC cells in the Ago2
complex (Fig. 3B). The roles of the
two miRNA candidates in ZO-1 reduction were then investigated; an
increase in miR191-5p alone decreased ZO-1 expression (Fig. S2A and B). However, overexpressing
miR429 in HA22T cells (Fig. S2C)
had no impact on ZO-1 protein expression (Fig. S2D). It was thus hypothesized that
the miR191-5p regulatory effects were associated with the 3′UTR
overlap of the IGF2BP3 binding sites, and that IGF2BP3 synergized
miR191-5p via the RISC. To further investigate this hypothesis, a
rescue experiment was performed to determine the role of miR191-5p
in the IGF2BP3-induced increase in HCC cell invasiveness. The
findings revealed that miR191-5p knockdown with an miR191-5p
inhibitor (Fig. S2E) partially
reversed the IGF2BP3-induced increase in HCC cell invasion and
decreased ZO-1 expression (Fig.
3C-D). Taken together, the data shown in Fig. 3A-D suggest that IGF2BP3 decreases
ZO-1 expression by cooperating with the miR191-5p-Ago2 complex via
the RISC.
 | Figure 3.IGF2BP3 decreases ZO-1 expression by
synergizing with miR191-5p via the RISC. (A) ZO-1 mRNA expression
was detected in the argonaute 2 complex using an RIP assay in HA22T
cells transfected with pLKO and shIGF2BP3. (B) Reverse
transcription-quantitative PCR was used to screen 6 potential
miRNAs which may be able to regulate ZO-1 from the argonaute 2
complex in Huh-7 cells transfected with pWPI and IGF2BP3-cDNA. (C)
Transwell invasion assays were performed using Huh-7 cells
transfected with pWPI+pWPI, pWPI+oeIGF2BP3, oemiR-191-5p+pWPI or
oemiR-191-5p+oeIGF2BP3. (D) Western blotting was performed to
detect ZO-1 expression in Huh-7 cells transfected with pWPI+pWPI,
pWPI+oeIGF2BP3, oemiR-191-5p+ pWPI or oemiR-191-5p+oeIGF2BP3. For C
and D, the quantifications are shown on the right, and are
presented as the mean ± SD. *P≤0.05 and **P≤0.01. ns, not
significant; IGF2BP3, insulin-like growth factor II mRNA-binding
protein 3; ZO-1, zonula occludens-1; miRNA/miR, microRNA; RISC,
RNA-induced silencing complex; RIP, RNA interaction-precipitation;
sh, short hairpin (RNA); oe, overexpression; NC, negative
control. |
Molecular mechanistic analysis of the
mechanisms by which IGF2BP3 enhances RISC function: By promoting
Ago2-mRNA interactions
To further determine the mechanisms of the
regulatory effects of the miR191-5p-IGF2BP3-Ago2 complex on ZO-1
mRNA degradation. The 3′UTRs of ZO-1 mRNA were first analyzed with
a consensus sequence of ‘AAUGU’ and IGF2BP3-RNA was found to
crosslink the overlapping miR191-5p target sites in the of ZO-1
mRNA 3′UTRs (23). A luciferase
assay was then applied to determine whether IGF2BP3-miR191-5p
directly targeted the overlap of ZO-1 mRNA 3′UTR to downregulate
its expression. The wild-type and mutant miRNA-target sites were
first cloned into the psicheck2 vector to construct the reporter
plasmids (Fig. 4A). As expected,
increasing miR191-5p resulted in decreased luciferase activity in
HA22T cells transfected with the wild-type, but not the mutant ZO-1
3′UTR (Fig. 4B). Subsequently, other
reporter plasmids were constructed with psicheck2 vectors carrying
the wild-type and mutant IGF2BP3-target sites (Fig. 4C). The luciferase assay revealed that
increasing IGF2BP3 expression resulted in decreased luciferase
activity in Huh-7 cells transfected with the wild-type ZO-1 3′UTR,
but not in the mutant 3′UTR. In addition, attenuating IGF2BP3
significantly increased the luciferase activity in HA22T cells
transfected with the wild-type, but not in the mutant ZO-1 3′UTR
(Fig. 4D). Taken together, the
results presented in Fig. 4A-D
suggest that IGF2BP3 can promote the function of miR191-5p by
binding the overlap target sites and enhancing Ago2-mRNA
interactions.
 | Figure 4.IGF2BP3 improves miR191-5p-induced
silencing complex function by binding the 3′-UTR of ZO-1 mRNA and
promoting Ago2-mRNA interactions. (A) Sequence alignment of the
ZO-1 3′UTR with wild-type vs. mutant potential miR191-5p targeting
sites. The left red sequence represents the potential IGF2BP3
binding site of the ZO-1 mRNA 3′UTR. The right red sequence
represents the potential miR191-5p binding site of the ZO-1 mRNA
3′UTR, and the crossed sequence represents deletion in the mutant
ZO-1 3′UTR. (B) Luciferase reporter activity after transfection of
wild-type and mutant ZO-1 3′UTR reporter constructs in HA22T cells
with/without miR191-5p. (C) Sequence alignment of the ZO-1 3′UTR
with wild-type vs. mutant potential IGF2BP3 targeting sites. The
left red sequence represents the potential IGF2BP3 binding site of
ZO-1 mRNA 3′UTR, and the crossed sequence represents deletion in
the mutant ZO-1 3′UTR. The right red sequence represents the
potential miR191-5p binding site of ZO-1 mRNA 3′UTR. (D) Luciferase
reporter activity following transfection with wild-type or mutant
ZO-1 3′UTR reporter constructs in Huh-7 cells with/without
IGF2BP3-cDNA, and HA22T cells treated with/without IGF2BP3-shRNA,
compared with the control cells. All quantifications are presented
as the mean ± SD. *P≤0.05 and **P≤0.01. ns, not significant;
IGF2BP3/IMP3, insulin-like growth factor II mRNA-binding protein 3;
ZO-1, zonula occludens-1; UTR, untranslated region; miR, microRNA;
sh, short hairpin (RNA); oe, overexpression; NC, negative
control. |
Discussion
IGF2BP3 is an oncofetal protein that is detected in
a number of malignant tumors (5).
Clinical research has indicated that IGF2BP3 can be used as a
biomarker to distinguish tumors from normal tissue. In addition,
according to previous studies, IGF2BP3 can also be used as an
independent prognostic indicator (11,38,39).
Notably, using data from online databases, the findings of the
present study indicate that higher IGF2BP3 expression levels may be
associated with the poor survival rates of patients with HCC, which
is in agreement with the findings of a previous study demonstrating
that IGF2BP3 may be involved in tumorigenesis, and that its
expression was significantly upregulated in HCC tissues (13). Thus, increased IGF2BP3 expression may
contribute to HCC progression. By contrast, decreasing the
expression of IGF2BP3 may result in decreased HCC cell
invasiveness.
miRNAs maturing from hairpin miRNA precursors act as
critical post-transcriptional regulators involved in tumorigenesis,
tumor progression and metastasis (40–44). The
RISC is the primary component of post-transcriptional regulation.
miRNAs recruit Ago2, a member of the argonaute protein family, to
guide the targeting of mRNA cleavage or translation inhibition
through the RISC (45). A number of
proteins have also been reported to enhance Ago2 activation to
improve RISC formation (46). One of
these proteins is IGF2BP3, which can influence the expression of
invasion- and migration-associated genes by strengthening the
function of Ago2 (23).
The present study demonstrated that IGF2BP3 promotes
cell invasiveness by downregulating ZO-1 expression, which seems to
oppose the suggestion that IGF2BP3 facilitates mRNA translation by
binding to and maintaining mRNA stability (37,47).
Thus, from the investigation of the mechanisms responsible for the
regulatory effects of IGF2BP3 on the invasion of HCC, and analysis
of related gene expression, the previous study identified that
IGF2BP3 promoted RISC formation (23). In the present study, the results of
Ago2 antibody pulldown assays confirm that IGF2BP3 enhances the
function of Ago2 (Fig. 3A), but also
suggest that miRNAs were targeting ZO-1 mRNA.
The present study demonstrated that miR191-5p
increases cell invasiveness. However, decreasing IGF2BP3 expression
partly reversed this miR191-5p-induced invasiveness. The
combination of other published and those of the present study
suggests that targeting IGF2BP3 blocks cell invasiveness via the
miR191-5p-Ago2 complex.
ZO-1 is a member of the membrane-associated
guanylate kinase homolog family. It interacts with transmembrane
proteins and links tight junction components to the cortical actin
cytoskeleton to maintain epithelial tight junction integrity
(48,49). Increasing evidence indicates that the
downregulation of ZO-1 expression is associated with the
enhancement of cancer dissemination and metastasis (27,50). The
present study revealed that miR191-5p and IGF2BP3 increases
cellular invasiveness by binding to the overlapping targeting sites
of ZO-1 mRNA. The overexpression of miR-191-5p or IGF2BP3 decreased
the ZO-1 protein expression level and promoted HCC cell
invasiveness. Conversely, the inhibition of miR191-5p or the
knockdown of IGF2BP3 restored ZO-1 expression and decreased
invasiveness.
Collectively, the findings of the present study
demonstrate that suppressing IGF2BP3 expression may inhibit HCC
cell invasiveness by altering IGF2BP3/miR191-5P/ZO-1 signaling
(Fig. 5), and that targeting this
signaling molecule with small molecules may aid in the development
of novel therapeutic strategies with which to better retard HCC
progression.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81672469).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG and XQ conceived and designed the study. TL, XO,
CZ and JZ made substantial contributions to the design of the
study, acquisition of data, interpretation of data and revising the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nielsen J, Christiansen J, Lykke-Andersen
J, Johnsen AH, Wewer UM and Nielsen FC: A family of insulin-like
growth factor II mRNA-binding proteins represses translation in
late development. Mol Cell Biol. 19:1262–1270. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yaniv K and Yisraeli JK: The involvement
of a conserved family of RNA binding proteins in embryonic
development and carcinogenesis. Gene. 287:49–54. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mueller F, Bommer M, Lacher U, Ruhland C,
Stagge V, Adler G, Gress TM and Seufferlein T: KOC is a novel
molecular indicator of malignancy. Br J Cancer. 88:699–701. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohashi R, Sangen M, Namimatsu S, Takei H
and Naito Z: IMP3 contributes to poor prognosis of patients with
metaplastic breast carcinoma: A clinicopathological study. Ann
Diagn Pathol. 31:30–35. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Samanta S, Sun H, Goel HL, Pursell B,
Chang C, Khan A, Greiner DL, Cao S, Lim E, Shultz LD and Mercurio
AM: IMP3 promotes stem-like properties in triple-negative breast
cancer by regulating SLUG. Oncogene. 35:1111–1121. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu D, Yang X, Jiang NY, Woda BA, Liu Q,
Dresser K, Mercurio AM, Rock KL and Jiang Z: IMP3, a new biomarker
to predict progression of cervical intraepithelial neoplasia into
invasive cancer. Am J Surg Pathol. 35:1638–1645. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li C, Rock KL, Woda BA, Jiang Z, Fraire AE
and Dresser K: IMP3 is a novel biomarker for adenocarcinoma in situ
of the uterine cervix: An immunohistochemical study in comparison
with p16(INK4a) expression. Mod Pathol. 20:242–247. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tschirdewahn S, Panic A, Püllen L, Harke
NN, Hadaschik B, Riesz P, Horváth A, Szalontai J, Nyirády P, Baba
HA, et al: Circulating and tissue IMP3 levels are correlated with
poor survival in renal cell carcinoma. Int J Cancer. 145:531–539.
2019.PubMed/NCBI
|
|
11
|
Jiang Z, Chu PG, Woda BA, Rock KL, Liu Q,
Hsieh CC, Li C, Chen W, Duan HO, McDougal S and Wu CL: Analysis of
RNA-binding protein IMP3 to predict metastasis and prognosis of
renal-cell carcinoma: A retrospective study. Lancet Oncol.
7:556–564. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen ST, Jeng YM, Chang CC, Chang HH,
Huang MC, Juan HF, Hsu CH, Lee H, Liao YF, Lee YL, et al:
Insulin-like growth factor II mRNA-binding protein 3 expression
predicts unfavorable prognosis in patients with neuroblastoma.
Cancer Sci. 102:2191–2198. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jeng YM, Chang CC, Hu FC, Chou HY, Kao HL,
Wang TH and Hsu HC: RNA-binding protein insulin-like growth factor
II mRNA-binding protein 3 expression promotes tumor invasion and
predicts early recurrence and poor prognosis in hepatocellular
carcinoma. Hepatology. 48:1118–1127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan J, Wei Q, Jian W, Qiu B, Wen J, Liu J,
Fu B, Zhou X and Zhao T: IMP3 predicts invasion and prognosis in
human lung adenocarcinoma. Lung. 194:137–146. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li W, Liu D, Chang W, Lu X, Wang YL and
Wang H, Zhu C, Lin HY, Zhang Y, Zhou J and Wang H: Role of IGF2BP3
in trophoblast cell invasion and migration. Cell Death Dis.
5:e10252014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Taniuchi K, Furihata M, Hanazaki K, Saito
M and Saibara T: IGF2BP3-mediated translation in cell protrusions
promotes cell invasiveness and metastasis of pancreatic cancer.
Oncotarget. 5:6832–6845. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pasiliao CC, Chang CW, Sutherland BW,
Valdez SM, Schaeffer D, Yapp DT and Ng SS: The involvement of
insulin-like growth factor 2 binding protein 3 (IMP3) in pancreatic
cancer cell migration, invasion, and adhesion. BMC Cancer.
15:2662015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jens M and Rajewsky N: Competition between
target sites of regulators shapes post-transcriptional gene
regulation. Nat Rev Genet. 16:113–126. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bell JL, Wächter K, Mühleck B, Pazaitis N,
Köhn M, Lederer M and Hüttelmaier S: Insulin-like growth factor 2
mRNA-binding proteins (IGF2BPs): Post-transcriptional drivers of
cancer progression? Cell Mol Life Sci. 70:2657–2675. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang W, Cheng X, Wang T, Song X, Zheng Y
and Wang L: LINC00467 promotes cell proliferation and metastasis by
binding with IGF2BP3 to enhance the mRNA stability of TRAF5 in
hepatocellular carcinoma. J Gene Med. 22:e31342020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mizutani R, Imamachi N, Suzuki Y, Yoshida
H, Tochigi N, Oonishi T, Suzuki Y and Akimitsu N: Oncofetal protein
IGF2BP3 facilitates the activity of proto-oncogene protein eIF4E
through the destabilization of EIF4E-BP2 mRNA. Oncogene.
35:3495–3502. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jønson L, Christiansen J, Hansen TVO,
Vikeså J, Yamamoto Y and Nielsen FC: IMP3 RNP safe houses prevent
miRNA-directed HMGA2 mRNA decay in cancer and development. Cell
Rep. 7:539–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ennajdaoui H, Howard JM, Sterne-Weiler T,
Jahanbani F, Coyne DJ, Uren PJ, Dargyte M, Katzman S, Draper JM,
Wallace A, et al: IGF2BP3 modulates the interaction of
invasion-associated transcripts with RISC. Cell Rep. 15:1876–1883.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
González-Mariscal L, Betanzos A and
Avila-Flores A: MAGUK proteins: Structure and role in the tight
junction. Semin Cell Dev Biol. 11:315–324. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagai T, Arao T, Nishio K, Matsumoto K,
Hagiwara S, Sakurai T, Minami Y, Ida H, Ueshima K, Nishida N, et
al: Impact of tight junction protein ZO-1 and twist expression on
postoperative survival of patients with hepatocellular carcinoma.
Dig Dis. 34:702–707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Wang L, Zhang H, Tu F, Qiang Y
and Nie C: Decreased expression of ZO-1 is associated with tumor
metastases in liver cancer. Oncol Lett. 17:1859–1864.
2019.PubMed/NCBI
|
|
27
|
Hsu YL, Hung JY, Chang WA, Lin YS, Pan YC,
Tsai PH, Wu CY and Kuo PL: Hypoxic lung cancer-secreted exosomal
miR-23a increased angiogenesis and vascular permeability by
targeting prolyl hydroxylase and tight junction protein ZO-1.
Oncogene. 36:4929–4942. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aden DP, Fogel A, Plotkin S, Damjanov I
and Knowles BB: Controlled synthesis of HBsAg in a differentiated
human liver carcinoma-derived cell line. Nature. 282:615–616. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Knowles BB, Howe CC and Aden DP: Human
hepatocellular carcinoma cell lines secrete the major plasma
proteins and hepatitis B surface antigen. Science. 209:497–499.
1980. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
31
|
Chang C, Lin Y, O-Lee TW, Chou CK, Lee TS,
Liu TJ, P'eng FK, Chen TY and Hu CP: Induction of plasma protein
secretion in a newly established human hepatoma cell line. Mol Cell
Biol. 3:1133–1137. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nakabayashi H, Taketa K, Yamane T,
Miyazaki M, Miyano K and Sato J: Phenotypical stability of a human
hepatoma cell line, HuH-7, in long-term culture with chemically
defined medium. Gan. 75:151–158. 1984.PubMed/NCBI
|
|
33
|
Stepanenko AA and Dmitrenko VV: HEK293 in
cell biology and cancer research: Phenotype, karyotype,
tumorigenicity, and stress-induced genome-phenotype evolution.
Gene. 569:182–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen X, Zhao M, Huang J, Li Y, Wang S,
Harrington CA, Qian DZ, Sun XX and Dai MS: MicroRNA-130a suppresses
breast cancer cell migration and invasion by targeting FOSL1 and
upregulating ZO-1. J Cell Biochem. 119:4945–4956. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu M, Yang J, Zhang Y, Zhou Z, Cui X,
Zhang L, Fung KM, Zheng W, Allard FD, Yee EU, et al: ZIP4 promotes
pancreatic cancer progression by repressing ZO-1 and claudin-1
through a ZEB1-dependent transcriptional mechanism. Clin Cancer
Res. 24:3186–3196. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lederer M, Bley N, Schleifer C and
Hüttelmaier S: The role of the oncofetal IGF2 mRNA-binding protein
3 (IGF2BP3) in cancer. Semin Cancer Biol. 29:3–12. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li D, Yan D, Tang H, Zhou C, Fan J, Li S,
Wang X, Xia J, Huang F, Qiu G and Peng Z: IMP3 is a novel
prognostic marker that correlates with colon cancer progression and
pathogenesis. Ann Surg Oncol. 16:3499–3506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li S, Cha J, Kim J, Kim KY, Kim HJ, Nam W
and Cha IH: Insulin-like growth factor II mRNA-binding protein 3: A
novel prognostic biomarker for oral squamous cell carcinoma. Head
Neck. 33:368–374. 2011.PubMed/NCBI
|
|
40
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. MicroRNA cancer regulation. Schmitz
U, Wolkenhauer O and Vera J: Dordrecht Springer Netherlands. 1–20.
2013. View Article : Google Scholar
|
|
43
|
Di Leva G and Croce CM: Roles of small
RNAs in tumor formation. Trends Mol Med. 16:257–267. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Meister G, Landthaler M, Patkaniowska A,
Dorsett Y, Teng G and Tuschl T: Human Argonaute2 mediates RNA
cleavage targeted by miRNAs and siRNAs. Mol Cell. 15:185–197. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jing Q, Huang S, Guth S, Zarubin T,
Motoyama A, Chen J, Di Padova F, Lin SC, Gram H and Han J:
Involvement of microRNA in AU-rich element-mediated mRNA
instability. Cell. 120:623–634. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Glisovic T, Bachorik JL, Yong J and
Dreyfuss G: RNA-binding proteins and post-transcriptional gene
regulation. FEBS Lett. 582:1977–1986. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Anderson JM: Cell signalling: MAGUK magic.
Curr Biol. 6:382–384. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gottardi CJ, Arpin M, Fanning AS and
Louvard D: The junction-associated protein, zonula occludens-1,
localizes to the nucleus before the maturation and during the
remodeling of cell-cell contacts. Proc Natl Acad Sci USA.
93:10779–10784. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hoover KB, Liao SY and Bryant PJ: Loss of
the tight junction MAGUK ZO-1 in breast cancer: Relationship to
glandular differentiation and loss of heterozygosity. Am J Pathol.
153:1767–1773. 1998. View Article : Google Scholar : PubMed/NCBI
|
















