Introduction
Tumor microenvironments (TMEs) are a complex ecology
of cells, comprising cancer-associated fibroblasts and various
infiltrating immune cells that provide support to tumor cells
during their transition to malignancy, such as tumor-associated
macrophages (TAMs) (1). TAMs are a
major constituent among the various innate and adaptive immune
cells involved with the TME (2).
TAMs are closely associated with tumor proliferation and metastasis
and have been demonstrated to promote tumor angiogenesis, cancer
cell infiltration into the circulation and suppression of antitumor
immune mechanisms (3). Due to this
involvement, TAMs are considered to be potential therapy targets in
cancer treatment (4). However, the
regulatory mechanisms underlying TAMs are yet to be fully
elucidated and requires further research.
TAMs express cytokines and chemokines that suppress
antitumor immunity (5). During the
transition from benign growth to invasive tumor, colony stimulating
factor-1 (CSF1) has been reported to be one of the key cytokines
that regulates cancer-initiated inflammatory responses (6). CSF1 is a major lineage regulator of
numerous macrophage populations, exerting its effect by controlling
their production, differentiation and function (7). High CSF1 concentrations in tumors have
been reported to be associated with poor prognosis (8). Furthermore, CSF1 and its receptor
(CSF1R) have been reported to be central to the promotion of
migration, survival and proliferation of monocytes (9).
CSF1R belongs to the platelet-derived growth factor
receptor family and is a type III protein tyrosine kinase receptor
(10). In addition to CSF1, CSF1R
may also be recognized by other ligands, such as interleukin 34,
leading to the full activation of the receptor (11). CSF1R-expressing macrophages have been
reported to be associated with poor survival in various types of
tumors, such as chronic lymphocytic leukemia and gastric cancer
(12,13). Macrophages in tumor-infiltrating
areas can selectively ignore the presence of tumor cells by highly
expressing CSF1R (14).
Additionally, a group of CSF1R-targeting small molecules and
monoclonal antibodies have been revealed to be effective in mono-
or combination-therapy (15).
In hepatocellular carcinoma (HCC), CSF1/CSF1R
blockade has been reported to serve a critical role in the
immunosuppressive nature of TMEs (16). CSF1R expression in macrophages serves
a pivotal role in the interaction between macrophages and hepatoma
cells (17). However, CSF1R
expression and its potential regulatory mechanism in HCC requires
further investigation.
The effect of methylation factors has been an area
of interest in tumor research, and DNA methylation is reported to
be a key regulatory mechanism in HCC (18). Previous epigenetics studies have
revealed an association between CSF1R methylation and tumor
proliferation or migration (19,20). In
common malignant testicular germ cell tumors, CSF1R hypomethylation
has been reported to be associated with poorer prognoses (21). Additionally, CSF1R expression was
significantly elevated when demethylated, resulting in tumor
metastasis promotion in melanoma (22). The results of the aforementioned
studies have indicated that the CSF1R-mediated methylation
regulatory mechanism served an important role in tumor
development.
Therefore, in the present study, 160 adjacent
non-cancerous tissue (ANT) and paired HCC tissue samples were
collected, and methylation genotyping of the CSF1R promoter region
was performed. The aim was to identify the expression and
methylation alterations between HCC and normal tissues. As an
important microenvironmental factor, the correlation between the
methylation of the CSF1R promoter region and the
clinicopathological features of the patients were analysed.
Finally, the significance of methylation of the CSF1R promoter
region in ANTs was explored for the early detection and treatment
of HCC.
Materials and methods
Patients and tissue samples
The current study was approved by the Ethics
Committee of the Department of Hepatobiliary Surgery at Sir Run Run
Shaw Hospital and Zhejiang Hospital. All patients with HCC provided
written informed consent. A total of 160 HCC samples and paired ANT
samples (3 cm from the tumor) were collected from patients who
underwent surgical liver tumor resection between July 2008 and
February 2014. The samples were immediately frozen in liquid
nitrogen and then stored at −80°C for DNA/RNA extraction.
Pathological diagnosis was based on the morphological and
immunohistochemical criteria provided by the World Health
Organization (23). The tumor stages
were classified according to the AJCC tumor-node-metastasis staging
system (24).
Of the 160 patients with HCC, 111 were treated at
Zhejiang Cancer Hospital and 49 at Sir Run Run Shaw Hospital. Their
ages ranged between 31 and 76 years (mean age ± SD, 52.6±9.9
years), and the male-to-female ratio was 131:29. A total of 136
patients were positive for hepatitis B (HBV) and 53 patients were
≥TII according to the tumor-node-metastasis staging system.
Detailed clinical information is summarized in Table I.
 | Table I.Clinicopathological characteristics
of 160 patients with hepatocellular carcinoma and their mean CSF1R
methylation levels in adjacent non-cancerous tissues. |
Table I.
Clinicopathological characteristics
of 160 patients with hepatocellular carcinoma and their mean CSF1R
methylation levels in adjacent non-cancerous tissues.
| Parameter | No. of
patients | Mean methylation
level of CSF1R Mean levels (%) in ANTs |
P-valueb |
|---|
| Sex |
|
|
|
|
Female | 29 (18.1%) | 68.0±7.0 | 0.0756 |
|
Male | 131 (81.9%) | 65.0±7.6 |
|
| Age (years) |
|
|
|
|
≥55 | 79 (49.4%) | 64.6±7.0 | 0.1652 |
|
<55 | 81 (50.6%) | 66.4±7.9 |
|
| Alcohol
habita |
|
|
|
|
Yes | 55 (34.4%) | 65.0±8.2 | 0.4850 |
| No | 105 (65.6%) | 65.9±7.2 |
|
| Hypertension |
|
|
|
|
Yes | 37 (23.1%) | 63.8±6.7 | 0.1115 |
| No | 123 (76.9%) | 66.1±7.7 |
|
| Diabetes |
|
|
|
|
Yes | 20 (12.5%) | 61.3±5.4 | 0.0062 |
| No | 140 (87.5%) | 66.2±7.6 |
|
| Hyperlipemia |
|
|
|
|
Yes | 37 (23.1%) | 63.8±6.6 | 0.1234 |
| No | 121 (75.6%) | 66.0±7.8 |
|
| AFP (µg/l) |
|
|
|
|
<20 | 53 (33.1%) | 64.7±6.4 | 0.2658 |
|
≥20 | 104 (65.0%) | 66.1±8.0 |
|
| CEA (µg/l) |
|
|
|
| ≤5 | 132 (82.5%) | 66.1±7.6 | 0.0639 |
|
>5 | 24 (15%) | 63.1±6.5 |
|
| HBV |
|
|
|
|
Yes | 136 (85%) | 65.9±7.7 | 0.3296 |
| No | 22 (13.8%) | 64.2±6.1 |
|
| No. of tumors |
|
|
|
|
<2 | 145 (90.6%) | 65.9±7.2 | 0.0332 |
| ≥2 | 15 (9.4%) | 61.4±9.9 |
|
| Tumor diameter
(cm) |
|
|
|
| ≤3 | 38 (23.8%) | 64.2±6.8 | 0.0101 |
|
3-5 | 48 (30%) | 64.7±7.8 |
|
|
5-10 | 60 (37.5%) | 68.0±7.0 |
|
|
>10 | 14 (8.8%) | 61.3±8.4 |
|
| Capsule
invasion |
|
|
|
|
Yes | 108 (67.5%) | 65.8±8.3 | 0.4167 |
| No | 52 (32.5%) | 64.9±5.8 |
|
| Tumor necrosis |
|
|
|
|
Yes | 29 (18.1%) | 67.5±7.0 | 0.1227 |
| No | 131 (81.9%) | 65.1±7.6 |
|
| Liver
cirrhosis |
|
|
|
|
Yes | 142 (88.8%) | 65.3±7.8 | 0.1480 |
| No | 13 (8.1%) | 66.4±5.3 |
|
| Microvascular
invasion |
|
|
|
|
Yes | 106 (66.3%) | 66.2±7.8 | 0.1540 |
| No | 54 (33.8%) | 64.3±6.9 |
|
| TNM stage |
|
|
|
| I | 106 (66.3%) | 66.6±7.0 | 0.0244 |
|
≥II | 53 (33.1%) | 63.6±8.2 |
|
DNA extraction and bisulfite
conversion
Total DNA was extracted from 25 mg tissue (both ANTs
and tumor tissues) using a QIAamp® DNA mini kit (Qiagen
GmbH), according to the manufacturer's protocol. The DNA
concentration was determined using a NanoDrop 2000 (Thermo Fisher
Scientific, Inc.). Subsequently, a total of 500 ng DNA from each
sample was modified by sodium bisulfite using the EpiTect Fast DNA
Bisulfite kit (Qiagen GmbH) according to the manufacturer's
protocol.
Gene bioinformatics and Sequenom
analysis
CSF1R sequences were obtained from the human
reference genome (GRch37/hg19; http://genome.ucsc.edu/) and were utilized to design
the methylation genotyping primers using the online EpiDesigner
software (www.epidesigner.com). For
amplification of CSF1R from the DNA extracted from ANTs and tumor
tissues, the following PCR primer pair was used: Forward,
5′AGGAAGAGAGTTTAGAGAGAGTAAGGGAGGGGTTA-3′ and reverse,
5′-CAGTAATACGACTCACTATAGGGAGAAGGCTTCATAATCAAACCCCAAATAAAAAA-3′. PCR
was performed using the GeneAmp 9700 system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) in a 10-µl reaction containing 2 µl
bisulfite-converted DNA (~10 ng/µl), 0.08 µl PCR enzyme (5 U/µl), 2
µl of each primer (1 µM), 1 µl PCR buffer (10×; Sequenom) and 0.08
µl deoxyribonucleotide triphosphates mix (25 mM each). The
amplification process began with an initial 4-min denaturation at
94°C, followed by 45 cycles of 20 sec at 94°C, 30 sec at 56°C and 1
min at 72°C, and finally an extension step at 72°C for 3 min.
Subsequently, the PCR products were treated with three standard
procedures of the MassARRAY EpiTYPER (Sequenom), according to the
manufacturer's protocol: Shrimp Alkaline Phosphatase cleanup, T
cleavage and clean resin. Finally, the treated DNA was transferred
to a MassARRAY Analyzer 4 (Sequenom) to analyze CSF1R promoter
methylation, according to the manufacturer's protocol.
An in-house RNA sequencing (RNA-seq) dataset, which
included data from tumor and ANTs of 11 patients with HCC, was
established as previously described and was used to investigate the
alteration of CSF1R expression between HCC tissues and ANTs
(25). Additionally, the methylation
and expression levels of CSF1R in 50 paired samples of patients
with HCC were downloaded from The Cancer Genome Atlas (TCGA)
database (https://portal.gdc.cancer.gov/) for validation. A
total of 17 CpG probes in the CSF1R gene included in the Illumina
HumanMethylation450 array from the TCGA database were analyzed, and
14 probes were successfully genotyped, but the methylation data of
the remaining 3 probes were not available.
Immunohistochemical analysis
16 pairs (10% of patients with HCC) of ANTs and
tumor tissues were randomly selected from the 160 patients with HCC
to examine CSF1R protein expression via staining. Tissues used for
immunohistochemistry were fixed in 10% neutral formalin at room
temperature, embedded in paraffin and cut into 3-µm-thick sections.
For staining, the sections were deparaffinized in xylene and
rehydrated in a descending ethanol series (100, 95, 90, 80 and 70%,
sequentially), and washed in water at room temperature. Antigen
retrieval was performed using 0.01 M citrate buffer (pH 6.0) at
high temperature in a pressure cooker for 5 min. Subsequently, the
sections were treated with 3% hydrogen peroxide (diluted with
methanol) for 10 min at 20°C and then incubated with 10% bovine
serum albumin (cat. no. A8010; Beijing Solarbio Science &
Technology Co., Ltd.) at room temperature for 30 min to block
non-specific antibody binding. The slides were further incubated
overnight with CSF1R antibody (1:100; cat no. ab52864; Abcam) at
4°C and then incubated with biotinylated secondary antibodies
(ready to use; cat. no. GK600710/100; Gene Tech Biotechnology Co.,
Ltd.) for 30 min at room temperature. After light counterstaining
with hematoxylin (cat. no. MB9897; Dailan Meilun Biology Technology
Co., Ltd.) for 2 min at room temperature, the slides were
dehydrated in an ascending ethanol series (70, 80, 90, 95 and 100%,
sequentially), mounted with a coverslip and observed under a light
microscope (magnification, ×100 and ×400; Nikon Eclipse 80i; Nikon
Corporation).
Immunostaining was scored according to the German
immunoreactive score (26). This
13-point method is used to determine the percentage of positive
cells by assigning them 0–4 points: 0, no positive cells; 1,
<10% positive cells; 2, 10–50% positive cells; 3, 50–80%
positive cells; and 4, >80% positive cells. Staining intensity
was graded as follows: 0, negative; 1, weak; 2, moderate; and 3,
strong. The final score was calculated as the multiplication of
these two indicators and ranged between 0 and 12. The bioinformatic
analysis software TBtools (v0.67361) was used to generate the
heat-maps (27).
Statistical analysis
SPSS software (version 25.0; IBM Corp.) was utilized
in the present study for statistical analysis. The continuous type
of clinical characteristics and methylation levels of each CpG
sites were presented as the mean ± SD, and the categorical type of
clinical characteristics were presented as number and percentage.
Statistical differences in CSF1R methylation, RNA expression and
HCC protein levels between tumor and paired ANT samples were
analyzed using a paired t-test. Correlation between methylation and
expression was analyzed by Pearson's correlation coefficient
analysis. Step-wise linear regression was performed to investigate
the association between clinicopathological characteristics and
methylation levels in ANTs. Unpaired t-test and one-way ANOVA with
the Scheffe post hoc test were used to evaluate methylation
difference between binary variables (such as diabetes status,
number of primary tumors and tumor stages) or multiple group
variables (tumor size), respectively. Receiver operating
characteristic (ROC) curves and the area under the curve (AUC) were
utilized to evaluate if CSF1R methylation can be used as a
predictor of biomarkers. P<0.05 was considered to indicate a
statistically significant difference.
Results
Clinicopathological
characteristics
Clinicopathological characteristics of 160 patients
with HCC are detailed in Table I. Of
those, 131 were male, 55 had a drinking habit (>50 g/day), 20
had diabetes and 37 had hyperlipidemia. Additionally, there were
104 patients with α fetoprotein (AFP) scores of >20 and 136 had
HBV. A total of 142 patients exhibited cirrhosis.
CSF1R methylation level analysis
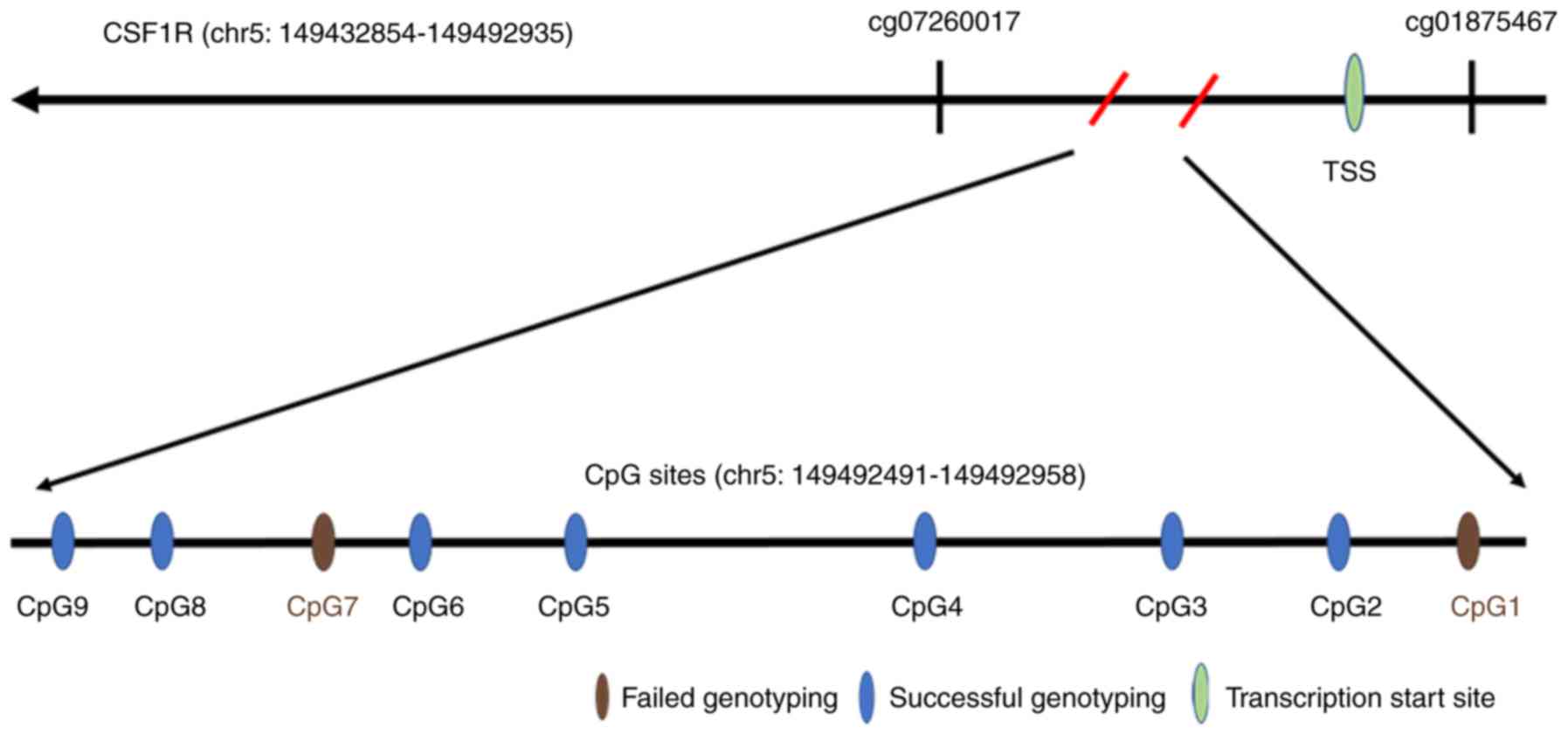
The CSF1R gene is located on
chr5:149432854-149492935, and the target amplicon for methylation
analysis was located on chr5:149492491-149492958. Therefore, the
468 bp amplicon ranged from −23 to 445 bp of the CSF1R gene, and
only included the transcription start sites. A total of 9 CpG sites
(sequentially named CpG1-9) were included in this amplicon region,
but 2 sites (CpG1 and CpG7) failed to be detected due to
limitations of the MassARRAY EpiTYPER technology (Fig. 1, Table
SI). The methylation levels of 7 successfully genotyped CpG
sites (CpG2, 3, 4, 5, 6, 8 and 9) demonstrated significant
correlation with each other in both HCC and ANTs (Tables SII and SIII).
Subsequently, paired t-tests were used to analyze
the methylation levels of each CpG site in HCC and normal tissues.
The results demonstrated that CSF1R methylation levels in all 7
successfully analyzed CpG sites were significantly decreased in HCC
compared with their paired ANT samples (Table II; Fig.
2A) and that the mean methylation difference between HCC and
ANTs ranged between 4.9 and 11.0% in the 7 CpG sites (Table II). The mean methylation level in
the CSF1R promoter was 57.3±14.4% in HCC tissues and 65.3±7.5% in
ANTs, respectively (P<0.0001; Table
II). Additionally, the present results were supported by data
from TCGA database. The methylation data of CSF1R in 50 HCC tissues
and their paired ANTs from TCGA (https://portal.gdc.cancer.gov/) database were
assessed, and 12 CpG probes of the Illumina HumanMethylation450
array in the CSF1R gene were hypomethylated in HCC tissues
(Fig. 2B).
| Table II.CSF1R promoter methylation in
patients with HCC. |
Table II.
CSF1R promoter methylation in
patients with HCC.
|
|
| HCC |
|---|
|
|
|
|
|---|
| CpG | Group | Mean (%) | ΔMean
(%)b | P-value |
|---|
| CpG 2 | ANT | 68.2±12.1 | 10.3 | <0.0001 |
|
| HCC | 57.9±20.2 |
|
|
| CpG 3 | ANT | 70.8±9.4 | 7.4 | <0.0001 |
|
| HCC | 63.4±15.9 |
|
|
| CpG 4 | ANT | 46.7±13.2 | 4.9 | 0.0070 |
|
| HCC | 41.8±19.5 |
|
|
| CpG 5 | ANT | 57.9±12.6 | 11.0 | <0.0001 |
|
| HCC | 46.9±18.3 |
|
|
| CpG 6 | ANT | 70.8±9.4 | 7.4 | <0.0001 |
|
| HCC | 63.4±15.9 |
|
|
| CpG 8 | ANT | 66.8±10.1 | 8.3 | <0.0001 |
|
| HCC | 58.5±16.5 |
|
|
| CpG 9 | ANT | 77.0±8.4 | 7.9 | <0.0001 |
|
| HCC | 69.1±15.5 |
|
|
| Meana | ANT | 65.3±7.5 | 8.1 | <0.0001 |
|
| HCC | 57.3±14.4 |
|
|
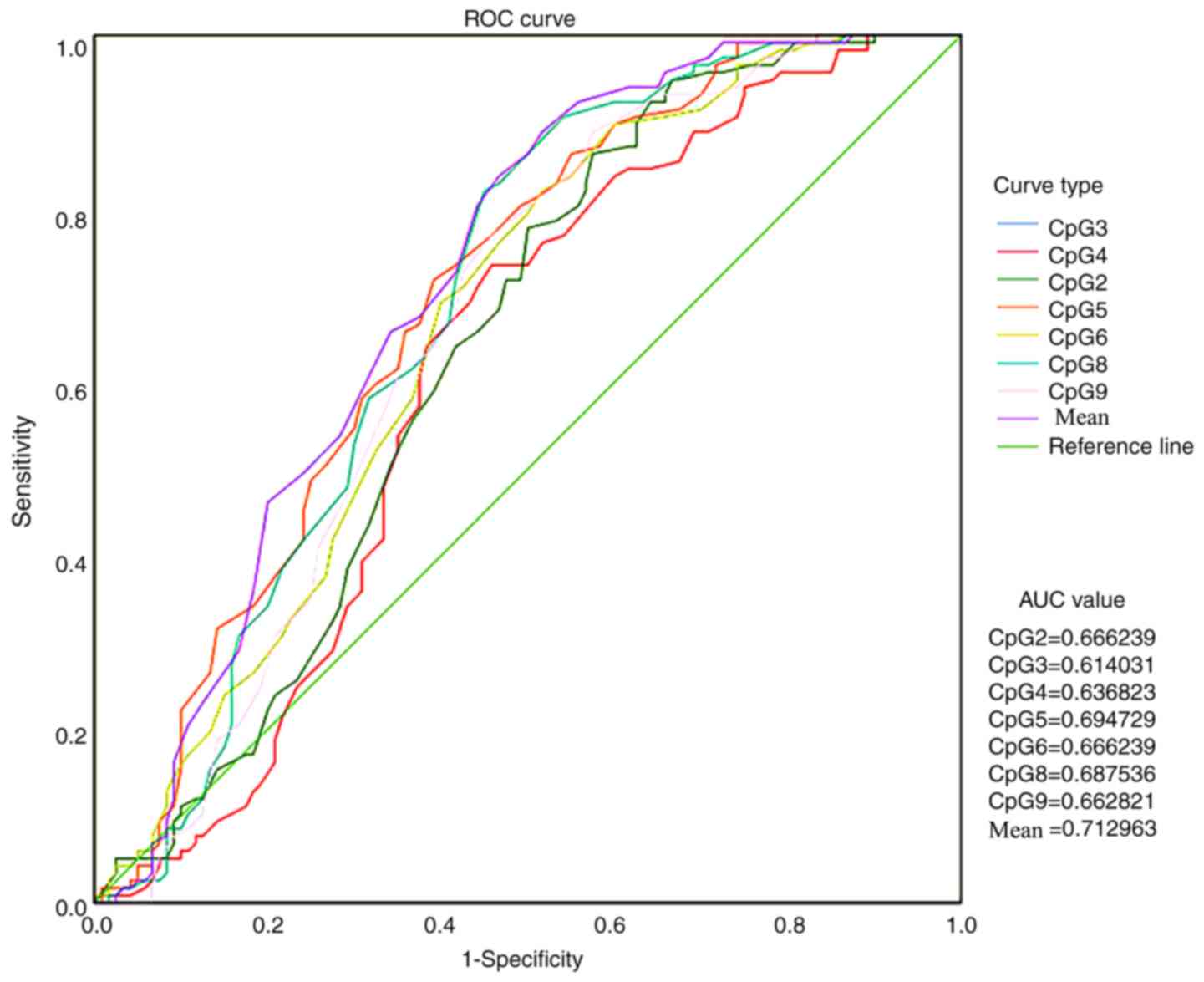
ROC curve analysis was conducted to compare CSF1R
methylation levels in ANTs and HCC tissues and to verify whether
the methylation level of the CSF1R promoter region could be used as
a biomarker for HCC diagnosis and treatment (Fig. 3). All AUC values were >0.5 and the
mean value of all methylation sites was 0.713.
To analyze the alteration of CSF1R expression in
HCC, an in-house RNA-seq dataset containing 11 HCC tissues and
their paired ANTs was used. In this dataset, a significantly higher
CSF1R expression was detected in ANTs compared with HCC tissues
[normal tissues vs. cancer tissues; reads per kilobase of exon
model per million mapped reads (FPKM), 23.94 vs. 11.43; P=0.0026;
Fig. 4A]. In addition, CSF1R
expression was then examined from TCGA dataset, from which 50
paired HCC RNA-seq data were downloaded and analyzed using paired
t-tests. The results from TCGA database revealed that CSF1R
expression was significantly decreased in HCC tissues compared with
that in ANTs (ANTs vs. HCC tissues; FPKM, 10.21 vs. 9.19;
P<0.0001; Fig. 4B).
Furthermore, immunohistochemistry was performed to
detect CSF1R protein expression in 16 paired HCC and ANTs, and to
verify expression differences at the protein level. The results
indicated that CSF1R protein levels in ANTs were markedly higher
compared with HCC tissues (Fig. 4C),
which was consistent with the data that was sequenced and obtained
from TCGA database. Using the 13-point method, it was also
suggested that CSF1R protein expression in ANTs was significantly
higher compared with that in tumor tissues (P=0.0025; Fig. 4D and E).
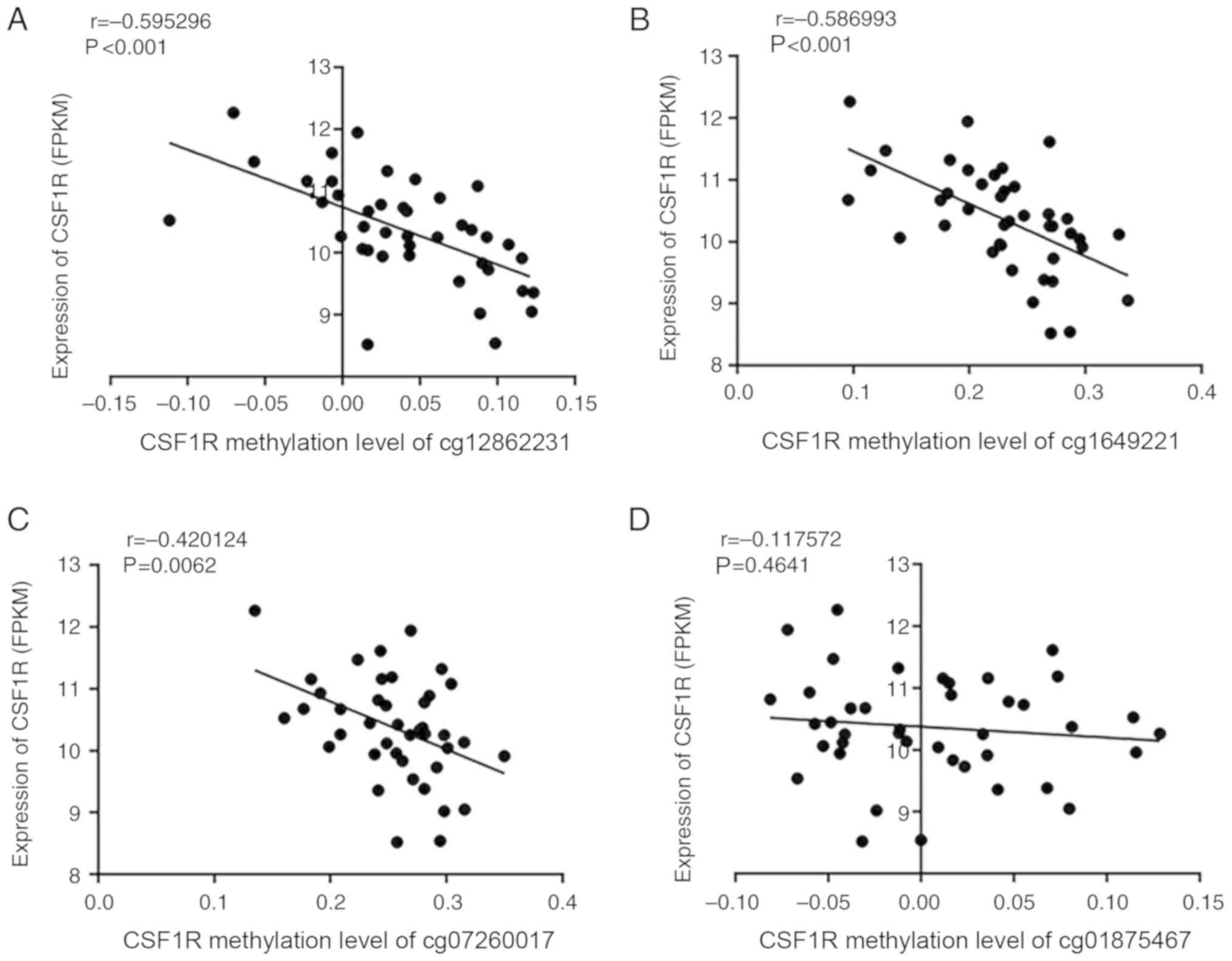
CSF1R expression and methylation data were
downloaded from TCGA datasets to identify the correlation between
the expression and methylation status. A significantly negative
correlation was identified between methylation and expression of
CSF1R in ANTs (Fig. 5A and B),
particularly in sites cg12862231 and cg16492211 (P<0.001). Sites
cg07260017 and cg01875467, which were adjacent to the selected CpG
islands according to TCGA data, were also analyzed and revealed to
follow the same trend (Fig. 5C and
D). However, when combining the ANT and paired HCC data
together, there was no correlation between CSF1R methylation and
expression (Fig. SI).
Correlation between CSF1R promoter
methylation status and patient clinicopathological
characteristics
To determine the potential effects of CSF1R
methylation and expression in HCC, the correlation between the
methylation status of CSF1R and comprehensive clinicopathological
features was analyzed. According to the high correlation among the
methylation levels of the CpG sites (Tables SII and SIII), the mean methylation levels of the 7
CpG sites were used to evaluate their association with
clinicopathological characteristics. The results revealed that
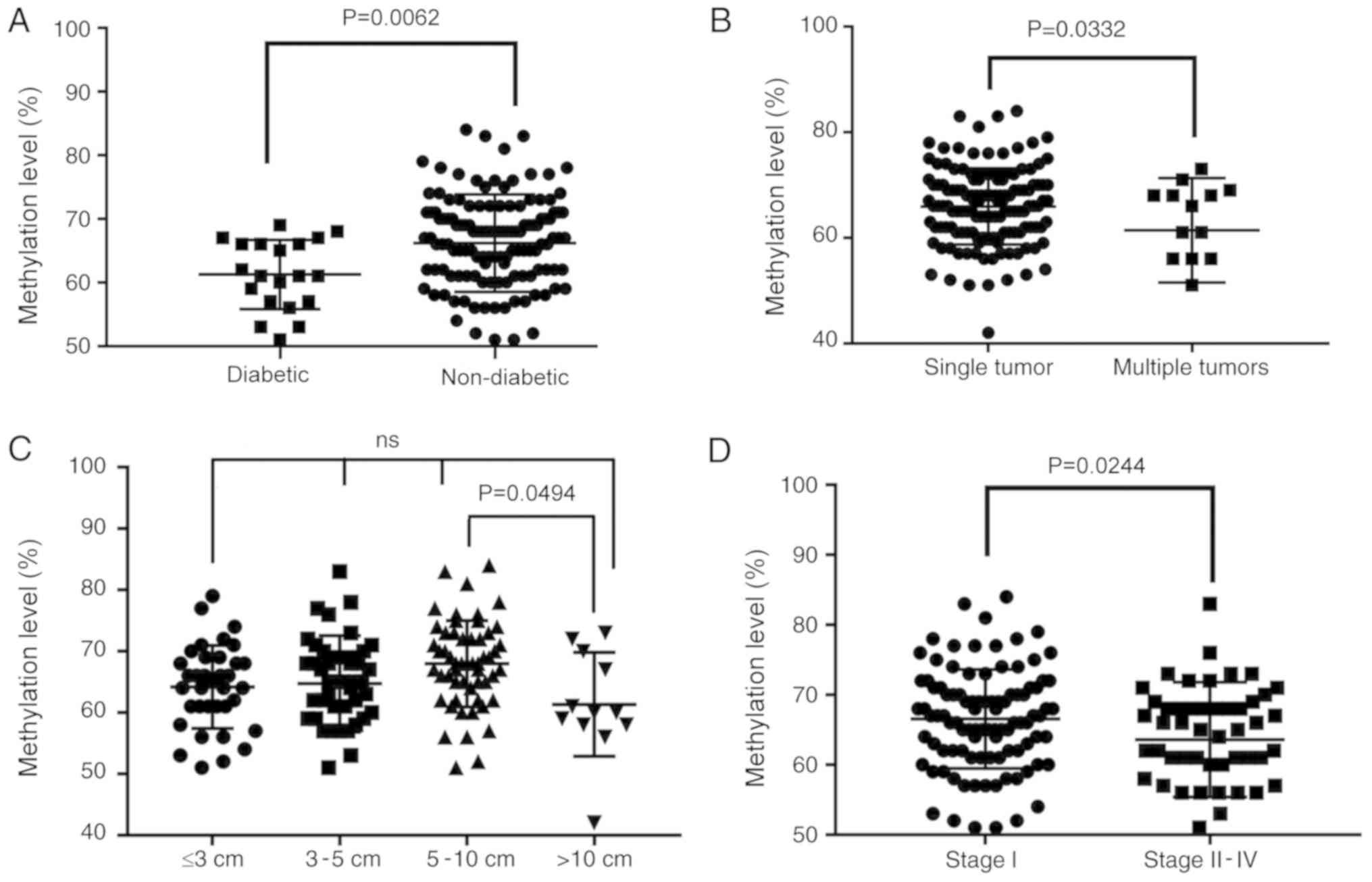
diabetes status was associated with methylation levels of CSF1R.
The mean value of the methylation site in the ANTs of patients with
diabetes was significantly different from that in tissues from
patients without diabetes. Patients with HCC that concurrently
presented with diabetes exhibited low ANT methylation (66.2% in 140
non-diabetic vs. 61.3% in 20 diabetic patients; P=0.0062; Fig. 6A and Table
I).
Furthermore, the results demonstrated that patients
with HCC who presented with multiple tumors exhibited an average
methylation level that was significantly decreased compared with
patients with single tumors (65.9 vs. 61.4%; P=0.0332; Fig. 6B and Table
I). Additionally, there was a significant difference between
tumor diameters of >10 cm and 5–10 cm (P=0.0494; Fig. 6C and Table
I) as demonstrated by ANOVA analysis followed by the Scheffe
post hoc test. However, a linear decrease with an increase in tumor
diameter was not reported. This could be due to the current sample
size being too small. Concurrently, the results revealed that the
average ANT methylation levels of patients with high-stage HCC also
exhibited a significant decrease (66.5 vs. 63.6%; P=0.0237;
Fig. 6D and Table I).
Discussion
The present study investigated whether CSF1R
methylation levels in ANTs from patients with HCC had a regulatory
effect on HCC progression and whether CSF1R methylation levels in
ANTs could be used as clinical biomarkers for HCC. The target
region of the CSF1R promoter includes 9 CpG sites, but 2 sites
failed to be genotyped in the present study. The methylation levels
of the remaining 7 CpG sites exhibited significant correlation
among each other, and the mean methylation levels of these 7 CpG
sites were used to analyze the association between CSF1R
methylation and clinicopathological characteristics. Therefore,
although 2 CpG sites in the CSF1R promoter were excluded from the
present analysis, it is hypothesized that they would not have had a
significant impact on the results of the present study.
By genotyping CSF1R methylation in ANTs and tumor
tissues of patients with HCC, the level of methylation in ANTs was
determined to be significantly higher compared with tumor tissues.
CSF1R expression in ANTs was also revealed to be significantly
higher than that of tumor tissues. However, methylation and
expression of CSF1R in ANTs demonstrated a significant negative
correlation as evidenced by HCC data from TCGA database. These
results appear to be contradictory. In fact, the methylation levels
of CSF1R in tumor tissues had a more discrete distribution than
those in ANTs (Fig. 2). When
combining the ANT and HCC data together, there was no correlation
between CSF1R methylation and expression (Fig. SI). A potential reason may be the
different percentages of immune cells between ANTs and HCC tissues.
CSF1R promoter region methylation in ANTs may have a special
regulatory pattern for CSF1R expression, and therefore further
studies are required to confirm these results.
ROC curve analysis of ANT and tumor methylation data
was conducted and revealed that CSF1R methylation had a potential
role in differentiating between cancer and normal tissues. This
indicated that the CSF1R methylation site in ANTs may be a possible
biomarker for HCC diagnosis (28).
In addition, several studies have indicated that TMEs are crucial
in tumor progression and cancer treatment (29–31).
CSF1R has been reported to serve an important regulatory role in
TMEs (32–34). As the receptor for CSF1, CSF1R is
activated after CSF1 binding and can regulate macrophage
differentiation (35). It has been
reported that high CSF1R expression in TMEs may cause the
progression of TAMs into the M2 type, which results in the loss of
macrophage immunity (36,37). Additionally, M2 type TAMs can promote
malignant tumor progression (2). A
previous study has reported that CSF1R inhibitors could be
developed as novel potential anticancer compounds (9). Therefore, CSF1R seems to have a
comprehensive clinical application value in HCC and should be
further investigated in future studies.
In the present study, patients with HCC who also had
diabetes exhibited significant ANT hypomethylation levels.
Furthermore, immunohistochemical staining of tissue sections from
patients with HCC demonstrated that CSF1R expression in ANTs was
significantly higher compared with HCC tissues. The correlation
between CSF1R methylation and expression in the TCGA database
identified a significant negative correlation. These results
indicated that there was an association between the methylation
level of CSF1R in the ANTs of patients with HCC and diabetes,
thereby regulating CSF1R expression. A previous study has
demonstrated that diabetes is more likely to trigger tumor
macrophages to promote colorectal cancer formation (38). Additionally, diabetes has been
reported to be an important factor in the induction of liver cancer
(39). Furthermore, numerous studies
have revealed that diabetes is associated with the development of
multiple types of cancer, such as liver and endometrial cancer
(40,41). Therefore, the detection of CSF1R
methylation levels may be a possible predictor of HCC in patients
with diabetes. However, the specific regulatory relationship
requires further experimental research.
Correlation analysis between ANT methylation data
and clinicopathological characteristics revealed that the level of
methylation in ANT sites was significantly reduced in patients with
HCC exhibiting multiple tumors. When the tumor diameter was >10
cm, a significant decrease was observed in the average methylation
level of the ANTs. No linear decline was observed in methylation
level according to diameter growth. This may have been due to the
insufficient experimental sample size. These results further
demonstrated that lower methylation resulted in high CSF1R
expression, which may be causing a decrease in the
immunosuppressive function of TAMs and may be leading to tumor
development and metastasis. Additionally, the level of CSF1R
methylation in ANTs was significantly reduced in patients with
advanced stage HCC. Lower methylation was accompanied by higher
expression, indicating that there is a regulatory relationship
between CSF1R hypomethylation in ANTs and tumor progression.
Methylation levels of CSF1R in ANTs may therefore be utilized to
predict tumor progression in patients with HCC.
In conclusion, the current study demonstrated that
the methylation level of CSF1R in the ANTs from patients with HCC
regulated TMEs, which serve a role in the regulation of metastasis.
Methylation was a key regulatory mechanism of CSF1R expression, and
CSF1R hypomethylation in ANTs was associated with poor
clinicopathological characteristics of patients with HCC.
Furthermore, CSF1R may be a potential immunological therapeutic
target for HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Young Elite
Scientist Sponsorship Program of the China Association for Science
and Technology (grant no. YESS 20150026), the National Key Research
and Development Program (grant no. 2016YFC0906400), the National
Natural Science Foundation of China (grant nos. 81872297 and
81874059), the Zhejiang Province Analysis and Test Technology
Project (grant no. 2018C37062) and the Fundamental Research Fund
for the Central Universities (grant no. 2016XZZX002-05).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request. Detailed pathological features and methylation data of
patients are not available to the public.
Authors' contributions
BC and XF contributed to the data analysis and
writing of the manuscript. DL and HL designed the experiments and
supervised the process. DZ, LH and YL contributed to the collection
of tissue samples, DNA extraction, bisulfite conversion and
methylation analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The pressent study was approved by the Ethics
Committee of the Department of Hepatobiliary Surgery at Sir Run Run
Shaw Hospital and Zhejiang Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Noy R and Pollard JW: Tumor-associated
macrophages: From mechanisms to therapy. Immunity. 41:49–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Choi J, Gyamfi J, Jang H and Koo JS: The
role of tumor-associated macrophage in breast cancer biology.
Histol Histopathol. 33:133–145. 2018.PubMed/NCBI
|
|
3
|
Vilgelm AE and Richmond A: Chemokines
modulate immune surveillance in tumorigenesis, metastasis, and
response to immunotherapy. Front Immunol. 10:3332019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rhee I: Diverse macrophages polarization
in tumor microenvironment. Arch Pharm Res. 39:1588–1596. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang L and Zhang Y: Tumor-associated
macrophages, potential targets for cancer treatment. Biomark Res.
5:252017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Konno T, Kasanuki K, Ikeuchi T, Dickson DW
and Wszolek ZK: CSF1R-related leukoencephalopathy: A major player
in primary microgliopathies. Neurology. 91:1092–1104. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Arcuri F, Buchwalder L, Toti P, Cintorino
M, Tosi P, Lockwood CJ, Rybalov B and Schatz F: Differential
regulation of colony stimulating factor 1 and macrophage migration
inhibitory factor expression by inflammatory cytokines in term
human decidua: Implications for macrophage trafficking at the
fetal-maternal interface. Biol Reprod. 76:433–439. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Richardsen E, Uglehus RD, Johnsen SH and
Busund LT: Macrophage-colony stimulating factor (CSF1) predicts
breast cancer progression and mortality. Anticancer Res.
35:865–874. 2015.PubMed/NCBI
|
|
9
|
Cannarile MA, Weisser M, Jacob W, Jegg AM,
Ries CH and Rüttinger D: Colony-stimulating factor 1 receptor
(CSF1R) inhibitors in cancer therapy. J Immunother Cancer.
5:532017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Williams H, Brenner S and Venkatesh B:
Identification and analysis of additional copies of the
platelet-derived growth factor receptor and colony stimulating
factor 1 receptor genes in fugu. Gene. 295:255–264. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Noda Y, Kawaguchi T, Korenaga M, Yoshio S,
Komukai S, Nakano M, Niizeki T, Koga H, Kawaguchi A, Kanto T and
Torimura T: High serum interleukin-34 level is a predictor of poor
prognosis in patients with non-viral hepatocellular carcinoma.
Hepatol Res. 49:1046–1053. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Galletti G, Caligaris-Cappio F and
Bertilaccio MT: B cells and macrophages pursue a common path toward
the development and progression of chronic lymphocytic leukemia.
Leukemia. 30:2293–2301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dammeijer F, Lievense LA, Kaijen-Lambers
ME, van Nimwegen M, Bezemer M, Hegmans JP, van Hall T, Hendriks RW
and Aerts JG: Depletion of tumor-associated macrophages with a
csf-1r kinase inhibitor enhances antitumor immunity and survival
induced by DC immunotherapy. Cancer Immunol Res. 5:535–546. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stanley ER and Chitu V: CSF-1 receptor
signaling in myeloid cells. Cold Spring Harb Perspect Biol. 6:2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xun Q, Wang Z, Hu X, Ding K and Lu X:
Small-molecule CSF1R inhibitors as anticancer agents. Curr Med
Chem. 2019.PubMed/NCBI
|
|
16
|
Zhu Y, Yang J, Xu D, Gao XM, Zhang Z, Hsu
JL, Li CW, Lim SO, Sheng YY, Zhang Y, et al: Disruption of
tumour-associated macrophage trafficking by the osteopontin-induced
colony-stimulating factor-1 signalling sensitises hepatocellular
carcinoma to anti-PD-L1 blockade. Gut. 68:1653–1666. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Y, Wen H, Zhou C, Su Q, Lin Y, Xie Y,
Huang Y, Qiu Q, Lin J, Huang X, et al: TNF-α derived from M2
tumor-associated macrophages promotes epithelial-mesenchymal
transition and cancer stemness through the Wnt/β-catenin pathway in
SMMC-7721 hepatocellular carcinoma cells. Exp Cell Res. 378:41–50.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Villanueva A, Portela A, Sayols S,
Battiston C, Hoshida Y, Méndez-González J, Imbeaud S, Letouzé E,
Hernandez-Gea V, Cornella H, et al: DNA methylation-based prognosis
and epidrivers in hepatocellular carcinoma. Hepatology.
61:1945–1956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shui IM, Wong CJ, Zhao S, Kolb S, Ebot EM,
Geybels MS, Rubicz R, Wright JL, Lin DW, Klotzle B, et al: Prostate
tumor DNA methylation is associated with cigarette smoking and
adverse prostate cancer outcomes. Cancer. 122:2168–2177. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Johnson KC, Houseman EA, King JE and
Christensen BC: Normal breast tissue DNA methylation differences at
regulatory elements are associated with the cancer risk factor age.
Breast Cancer Res. 19:812017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bo H, Cao K, Tang R, Zhang H, Gong Z, Liu
Z, Liu J, Li J and Fan L: A network-based approach to identify DNA
methylation and its involved molecular pathways in testicular germ
cell tumors. J Cancer. 10:893–902. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Neubert NJ, Schmittnaegel M, Bordry N,
Nassiri S, Wald N, Martignier C, Tillé L, Homicsko K, Damsky W,
Maby-El Hajjami H, et al: T Cell-induced CSF1 promotes melanoma
resistance to PD1 blockade. Sci Transl Med. 10(pii): eaan33112018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Najib and Haboubi: Pathology and genetics:
Tumours of the digestive system. Stanley R Hamilton and Lauri A
Aaltonen: ISBN No.: 02-832-2410-8.
|
|
24
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang H, Ye J, Weng X, Liu F, He L, Zhou D
and Liu Y: Comparative transcriptome analysis reveals that the
extracellular matrix receptor interaction contributes to the venous
metastases of hepatocellular carcinoma. Cancer Genet. 208:482–491.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tang L, Tan YX, Jiang BG, Pan YF, Li SX,
Yang GZ, Wang M, Wang Q, Zhang J, Zhou WP, et al: The prognostic
significance and therapeutic potential of hedgehog signaling in
intrahepatic cholangiocellular carcinoma. Clin Cancer Res.
19:2014–2024. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen C, Chen H, He Y and Xia RJB: TBtools,
a toolkit for biologists integrating various biological data
handling tools with a user-friendly interface. 289660. 2018.
|
|
28
|
Ahsan M, Ek WE, Rask-Andersen M, Karlsson
T, Lind-Thomsen A, Enroth S, Gyllensten U and Johansson Å: The
relative contribution of DNA methylation and genetic variants on
protein biomarkers for human diseases. PLoS Genet. 13:e10070052017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cooper J and Giancotti FG: Integrin
signaling in cancer: Mechanotransduction, stemness, epithelial
plasticity, and therapeutic resistance. Cancer Cell. 35:347–367.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Puisieux A: Role of epithelial-mesenchymal
transition in tumor progression. Bull Acad Natl Med. 193:2017–2032;
discussion 2032–2014. 2009.(In French). PubMed/NCBI
|
|
31
|
Alvarez MA, Freitas JP, Mazher Hussain S
and Glazer ES: TGF-β inhibitors in metastatic pancreatic ductal
adenocarcinoma. J Gastrointest Cancer. 50:207–213. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Balkwill F and Mantovani A: Inflammation
and cancer: Back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zitvogel L and Kroemer G: CD103+ dendritic
cells producing interleukin-12 in anticancer immunosurveillance.
Cancer Cell. 26:591–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mantovani A, Ming WJ, Balotta C,
Abdeljalil B and Bottazzi B: Origin and regulation of
tumor-associated macrophages: The role of tumor-derived chemotactic
factor. Biochim Biophys Acta. 865:59–67. 1986.PubMed/NCBI
|
|
35
|
Papadopoulos KP, Gluck L, Martin LP,
Olszanski AJ, Tolcher AW, Ngarmchamnanrith G, Rasmussen E, Amore
BM, Nagorsen D, Hill JS and Stephenson J Jr: First-in-human study
of AMG 820, a monoclonal anti-colony-stimulating factor 1 receptor
antibody, in patients with advanced solid tumors. Clin Cancer Res.
23:5703–5710. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Paolino M and Penninger JM: The role of
TAM family receptors in immune cell function: Implications for
cancer therapy. Cancers (Basel). 8:2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Q, Lu Y, Li R, Jiang Y, Zheng Y, Qian
J, Bi E, Zheng C, Hou J, Wang S and Yi Q: Therapeutic effects of
CSF1R-blocking antibodies in multiple myeloma. Leukemia.
32:176–183. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ito K, Ishigamori R, Mutoh M, Ohta T, Imai
T and Takahashi M: Ay allele promotes azoxymethane-induced
colorectal carcinogenesis by macrophage migration in
hyperlipidemic/diabetic KK mice. Cancer Sci. 104:835–843. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rawla P, Sunkara T, Muralidharan P and Raj
JP: Update in global trends and aetiology of hepatocellular
carcinoma. Contemp Oncol (Pozn). 22:141–150. 2018.PubMed/NCBI
|
|
40
|
The Editors Of The Lancet Diabetes
Endocrinology, . Retraction and republication-Worldwide burden of
cancer attributable to diabetes and high body-mass index: A
comparative risk assessment. Lancet Diabetes Endocrinol. 6:4372018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pearson-Stuttard J, Zhou B, Kontis V,
Bentham J, Gunter MJ and Ezzati M: Worldwide burden of cancer
attributable to diabetes and high body-mass index: A comparative
risk assessment. Lancet Diabetes Endocrinol. 6:95–104. 2018.
View Article : Google Scholar : PubMed/NCBI
|