Introduction
Retinoblastoma (RB) is the most prevalent type of
primary inherited intraocular malignant tumor in infancy and
children, affecting their life, vision, facial appearance and
psychological development (1,2). The
disease is prone to intracranial and distant metastasis, often
endangering the life of the child, particularly in children aged
<5 years (3,4). The predominant treatment for patients
includes chemotherapy, enucleation, laser therapy or cryotherapy,
the most common treatment protocol includes chemotherapy combined
with immunotherapy, which is cytotoxic to RB cells (1). Therefore, early detection, early
diagnosis and early treatment are the key to improving the cure
rate and reducing the mortality rate.
MiRNAs, a family of small and noncoding RNA
molecules consisting of 21–25 nucleotides, regulate the expression
of their target genes through suppressing the translation or
promoting the RNA degradation (5,6). They
are identified as tumor suppressors or oncogenes in many types of
tumors and thus regulate tumor progression and metastasis. Mounting
evidence has indicated that miRNAs were identified in RB and
regulated cell migration, invasion, proliferation and apoptosis
(7–11). Sun et al (12) showed that suppression of miR-492
inhibited RB cell viability and invasion by targeting LATS2.
miR-183 may be involved in RB progression by targeting LRP6
(13). Also, miR-34a exhibited
inhibitory effect on RB cell growth and promoting effect on cell
apoptosis (14). Moreover, miR-494
(miR-494-3p) was identified to be highly expressed in RB cells and
associated with its tumorigenesis (15). However, the mechanism regulating RB
progression by miR-494 expression is yet to be fully
elucidated.
Phosphatase and tensin homolog (PTEN) has been
reported to regulate the PI3K/AKT signaling pathway negatively
(16). Studies have confirmed that
PTEN was disordered in multiple cancers, and was considered to be
one of the potential factors in tumorigenesis (17,18).
Previous studies have summarized the role of miRNAs in regulating
PTEN in cancers and have determined that the miRNA/PTEN pathway is
involved in tumor cell growth, invasion and migration (19–21). For
instance, miR-296/PTEN axis facilitated gastric cancer
tumorigenesis in vitro and in vivo (22). miR-548 promoted NSCLC cell invasion
by regulating PTEN (23). Moreover,
miR-182 promoted breast cancer cell viability and invasion via
targeting PTEN (24). A previous
study has shown that miR-494-3p can promote the progression of
endometrial cancer by regulating the PTEN/PI3K/AKT pathway
(25). However, whether PTEN acted
as the target of miR-494 in regulating RB progression is
unclear.
Herei, it is suggested that miR-494 expression was
upregulated in RB tissues, and cells and miR-494 enhanced RB cell
viability, migration and invasion. PTEN was verified as the target
of miR-494 in RB cells. I was shown that miR-494 facilitated RB
cell progression by targeting PTEN through PI3K/AKT signaling
pathway.
Patients and methods
Tissue samples
Ten normal retina tissues and RB tissues were
provided by Jinan Zhangqiu District Hospital of TCM (Jinan, China).
None of the patients in this study received preoperative
radiotherapy or chemotherapy, and all patients were diagnosed by
three pathologists. The patients consisted of 6 males and 4 females
aged 1.2 to 9.9 years (mean, 5.4). According to ICRB, the RB
patients were divided into phase I (3 cases), phase II (5 cases)
and phase III (2 cases). There was no significant difference in
age, sex and clinical stages of the 10 patients. The adjacent
healthy tissues were set as the control. The fresh samples were
stored in −80°C refrigerator for further analysis. Parents of the
patients provided a written informed consent prior to surgical
excision. The Ethics Committee of Jinan Zhangqiu District Hospital
of TCM approved this study.
Cell culture
Two RB cell lines (Y79, SO-RB50) and normal retina
epithelium cell line (APRE-19) were obtained from ATCC. All the
cells were cultured in RPMI-1640 medium with 10% FBS. Cells were
then left to grow in a humidified incubator containing 5%
CO2 at 37°C.
qRT-PCR
Total RNA was extracted with the help of TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
concentration and purity of RNA were tested via NanoDrop 2000
instrument (Thermo Fisher Scientific, Inc.). To perform the
quantitative detection of miR-18a or mRNA expression, cDNAs were
synthesized through the PrimeScript RT reagent (Takara
Biotechnology Co.). GAPDH was used as the control for the
normalization of expression levels of genes. The snRNA U6 was used
as the control of miRNA. The primers were as follows: miR-494-F,
5′-GATACTCGAAGGAGAGGTTGTC-3′ and miR-494-R,
5′-GAGGTTTCCCGTGTATGTTTCAT-3′; PTEN-F, 5′-CGGCAGCATCAAATGTTTCAG-3′
and PTEN-R, 5′-AACTGGCAGGTAGAAGGCAACTC-3′; GAPDH-F,
5′-CTCTGCTCCTCCTGTTCGAC-3′ and GAPDH-R, 5′-CGACCAAATCCGTTGACTCC-3′;
U6-F, 5′-ATTGGAACGATACAGAGAAGATT-3′ and U6-R,
5′-GGAACGCTTCACGAATTTG-3′. The data were expressed as
2−ΔΔCt, indicating the target mRNA relative level.
Western blot analysis
Total protein was extracted with RIPA lysis buffer.
The protein concentration was determined using the BCA method
(Beyotime Institute of Biotechnology). The assay was performed as
follow: 50 µg of total proteins were loaded onto the gel and
electrophoresed. After transferring to the NC membranes, the
membranes were blocked with 5–10% skim milk. Then, the membranes
were incubated with the primary antibodies at 4°C overnight.
Subsequently, the second antibodies were added for incubating at
room temperature for 2 h. Finally, ECL agents were applied for
observing the protein bands. ImageJ software (version 1.48;
National Institutes of Health) was used for densitometry. GAPDH was
employed as an internal control.
MTT assay
RB cells (5×103) were placed onto 96-well
plates. When the cells were cultured for 0, 1, 2, 3 and 4 days, MTT
solution (20 µl) was added and cultured for 4 h at 37°C. A Bio-Rad
microplate reader (Bio-Rad Laboratories, Inc.) was used to read the
absorbance at 490 nm.
Overexpression of miR-494 or silence
miR-494 and overexpression of PTEN
miR-494 mimic or inhibitor was purchased from
RiboBio Co., Ltd. SiRNA PTEN was obtained from Genechem Co., Ltd.
The transfection was performed with the help of Lipofectamine 2000
for 48 h.
Transwell assay
The Transwell assay was applied to perform cell
migration and invasion using Transwell inserts (8 µm pores; BD
Biosciences). The differences between cell migration and invasion
were whether the top chamber was pre-coated with or without
Matrigel. Firstly, the cells were placed into the top chamber to
incubate for 24 h. The lower chamber was added with complete medium
with 10% FBS. The cells in the lower chamber that migrated or
invaded from the top chamber were then fixed with 4%
paraformaldehyde and stained with 0.5% crystal violet. Finally, the
number of migrated or invaded cell was counted using a microscope
(magnification, ×200).
Luciferase assay
The wild-type (wt) and mutant (mut) 3′UTR of PTEN
was first inserted into the pGL3 reporter vector (Promega
Corporation). Then, the PTEN 3′UTR-pGL3 reporter vector and miR-494
mimic were co-transfected into Y79 cells and plated in 24-well
plates for 48 h, the cells were harvested and lysed, and the
luciferase activity was detected by a Dual-Luciferase Reporter
System (Promega) according to the protocol of the manufacturer.
Relative firefly luciferase activity was normalized to
Renilla luciferase activity.
Statistical analysis
Results are represented as the mean ± SD of at least
triplicates. SPSS 22.0 Software (IBM Corp.) was applied for
performing statistical analyses. Student's t-test was carried out
for determining the statistical significance of differences in two
groups and one-way analysis of variance with Tukey's post hoc test
was applied in more than two groups. P<0.05 indicates
statistical significance.
Results
Increase of miR-494 in RB tissues
The average expression level of miR-494 in RB
tissues was analyzed by qRT-PCR and the findings indicated that it
was obviously increased in RB tissue samples compared with normal
tissues (Fig. 1A). Then miR-494
expression level was evaluated in 5 clinical RB specimens versus 5
normal tissues. Results showed that its expression was obviously
increased in RB tissues (Fig. 1B).
Moreover, the correlation between miR-494 expression and the
clinicopathological features of 66 RB patients was analysed. The
median value of miR-494 expression was used as the cut-off point to
divide miR-494 expression into high and low expression of miR-494.
Results demonstrated that the high miR-494 expression was
associated with N classification and differentiation in patients
with RB (Table I). Collectively,
these results indicated that miR-494 might play important roles in
RB progression.
 | Table I.Association study, and correlation
between miR-494 expression and clinicopathological
characteristics. |
Table I.
Association study, and correlation
between miR-494 expression and clinicopathological
characteristics.
|
|
| miR-494 |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | N=66 | Low expression | High
expression | P-value |
|---|
| Age (years) |
|
|
| 0.569 |
| ≤5 | 52 | 18 | 34 |
|
|
>5 | 14 | 6 | 8 |
|
| Sex |
|
|
| 0.930 |
|
Male | 39 | 14 | 25 |
|
|
Female | 27 | 10 | 17 |
|
| Clinical stage |
|
|
| 0.478 |
|
I–II | 20 | 6 | 14 |
|
|
III–IV | 46 | 18 | 28 |
|
| N
classification |
|
|
| 0.019a |
| N0 | 29 | 6 | 23 |
|
|
N1+2 | 37 | 18 | 19 |
|
|
Differentiation |
|
|
| 0.047a |
| Well
and moderate | 24 | 5 | 19 |
|
|
Poorly | 42 | 19 | 23 |
|
| Largest tumor
(mm) |
|
|
| 0.070 |
|
≤15 | 40 | 18 | 22 |
|
|
>15 | 26 | 6 | 20 |
|
miR-494 enhances RB cell viability,
invasion and migration
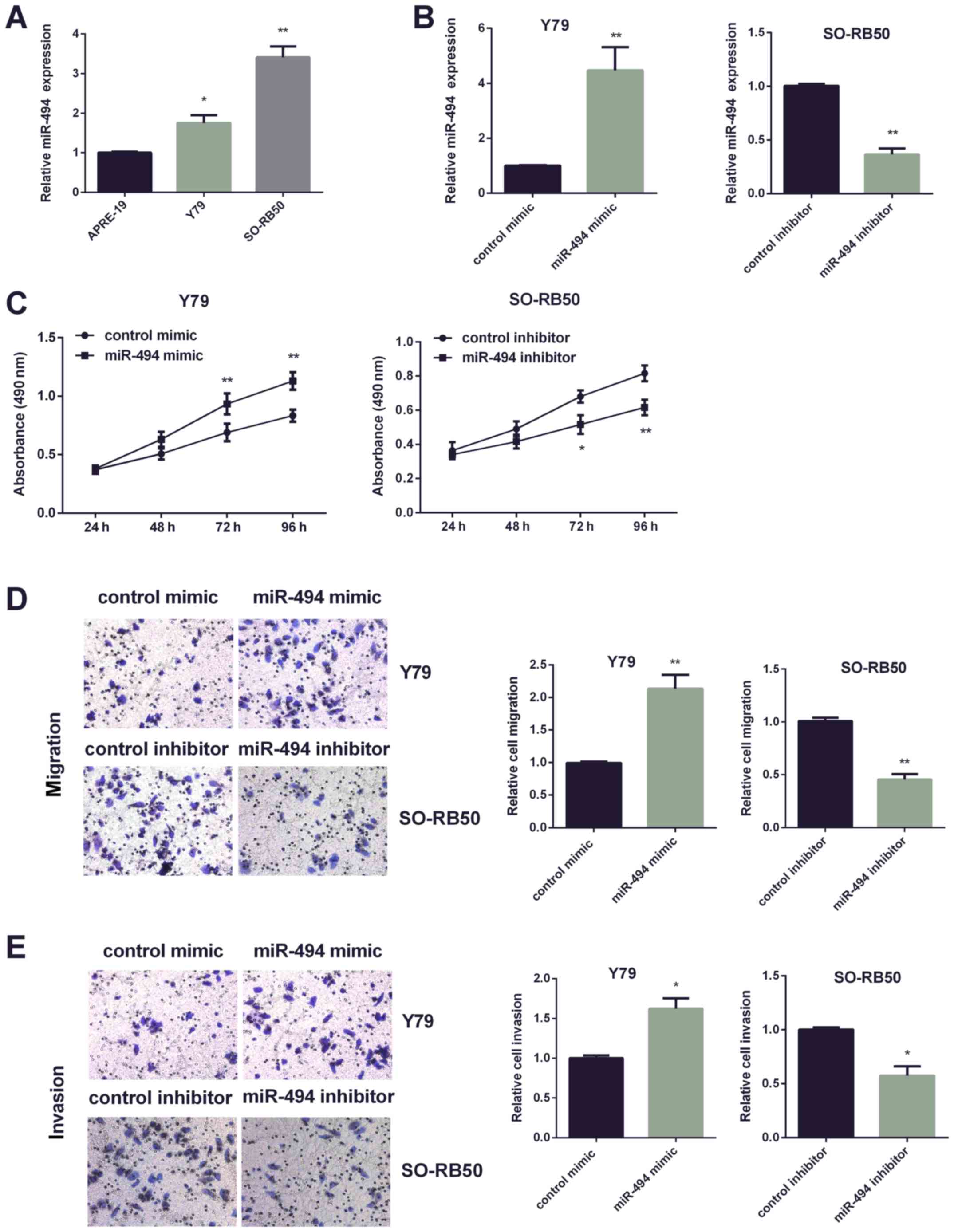
Two RB cell lines (Y79, SO-RB50) were applied to
investigate miR-494 function in RB progression. Due to the higher
expression of miR-494 in Y79 cells, miR-494 expression was
increased in Y79 cells, and miR-494 expression in SO-RB50 cells was
suppressed (Fig. 2A). qRT-PCR was
applied for confirming efficiency of miR-494 transfection in the
two cell lines respectively (Fig.
2B). Then, MTT assay was performed to detect cell viability
affected by altered miR-494. It was found that increasing miR-494
enhanced Y79 cell paoliferation, while decreasing miR-494 inhibited
SO-RB50 cells viability (Fig. 2C).
Furthermore, Transwell assay was used to investigate miR-494 effect
on cell invasion and migration. Data showed dramatically increased
mobility and invasiveness of Y79 cells by miR-494 mimic, whereas it
was markedly reduced by miR-494 inhibitor in SO-RB50 cells
(Fig. 2D and E).
PTEN is the miR-494 target gene in RB
cells
We used the algorithm provided by TargetScan Human
to predict the candidate targets of miR-494 and to determine the
mechanisms responsible for the effects of miR-494 in RB cells. PTEN
was temporarily selected for further validation among the predicted
targets, because it has a potential role in the regulation of RB
malignancies (Fig. 3A). To further
verify whether PTEN was the direct target of miR-494,
Luciferase-based assay was carried out to perform this experiment.
The findings showed that miR-494 mimic dramatically inhibited PTEN
3′UTR luciferase activity in Y79 cells (Fig. 3B). Consistently, miR-494 inhibitor
dramatically enhanced PTEN 3′UTR luciferase activity in SO-RB50
cells (Fig. 3C). Apart from this,
western blot analysis and qRT-PCR were performed to test PTEN
expression. Results indicated that PTEN protein (Fig. 3D) and mRNA (Fig. 3E) levels were remarkably inhibited by
miR-494 overexpression in Y79 cells, and increased by miR-494
suppression in SO-RB50 cells. Fig.
3F shows the negative relationship between miR-494 and PTEN
expression by Spearman's Rank correlation.
miR-494 promotes RB cell progression
via PTEN
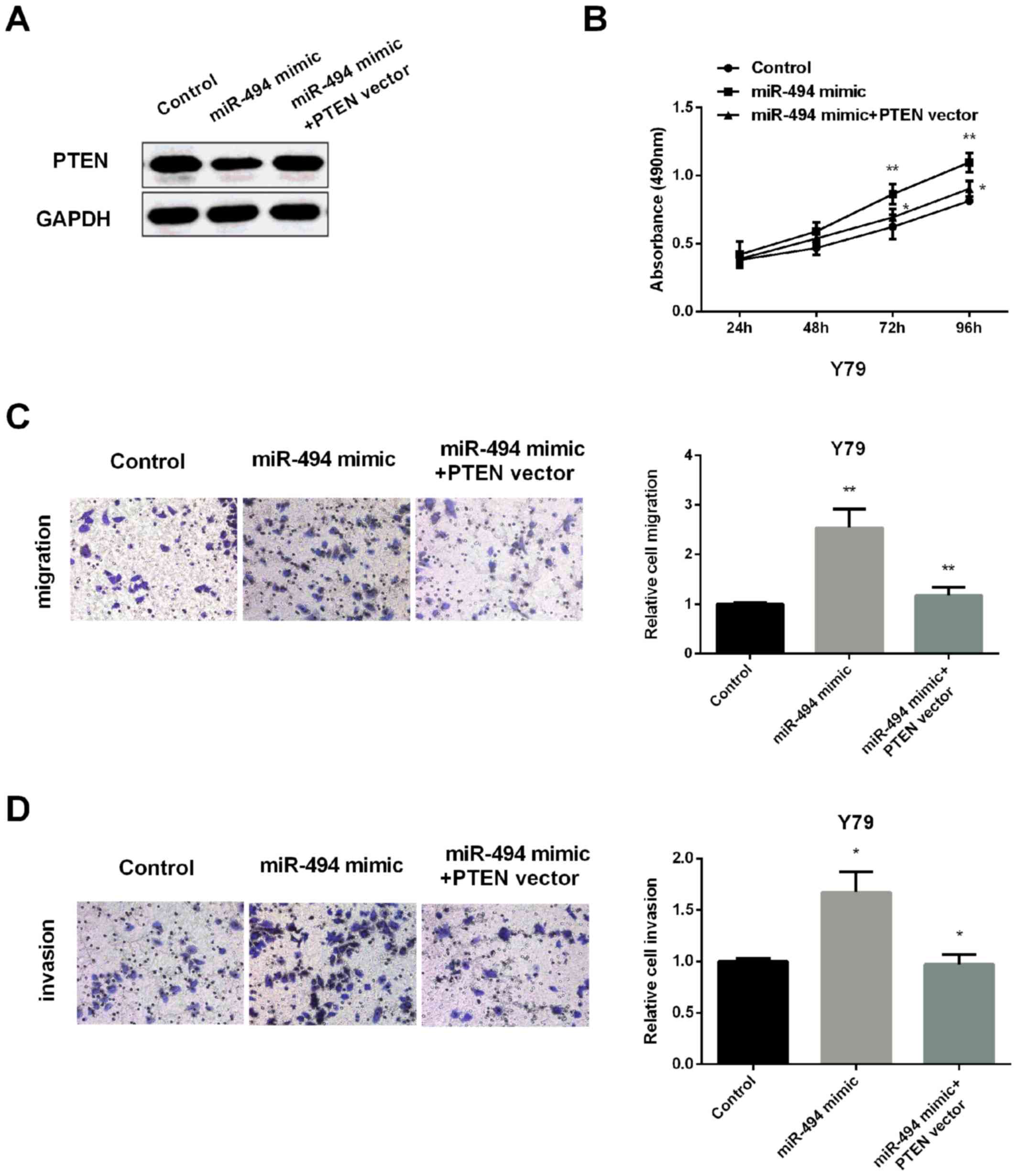
To further detect whether PTEN regulates miR-494
effect on RB cell viability, invasion and metastasis, PTEN level
was restored in Y79 cells inhibited by miR-494 overexpression
(Fig. 4A). qRT-PCR was applied for
confirming efficiency of PTEN transfection in Y79 cells. Then, MTT
assay was performed to test cell viability. As expected, miR-494
mimic increased the cell proliferation, but its effect was
overturned by overexpression of PTEN (Fig. 4B). Similarly, increasing the
expression of PTEN also reversed the increased migration (Fig. 4C) and invasion (Fig. 4D) regulated by miR-494 mimic. All the
results demonstrated that miR-494 promoted RB cell proliferation,
invasion and migration via inhibiting PTEN expression.
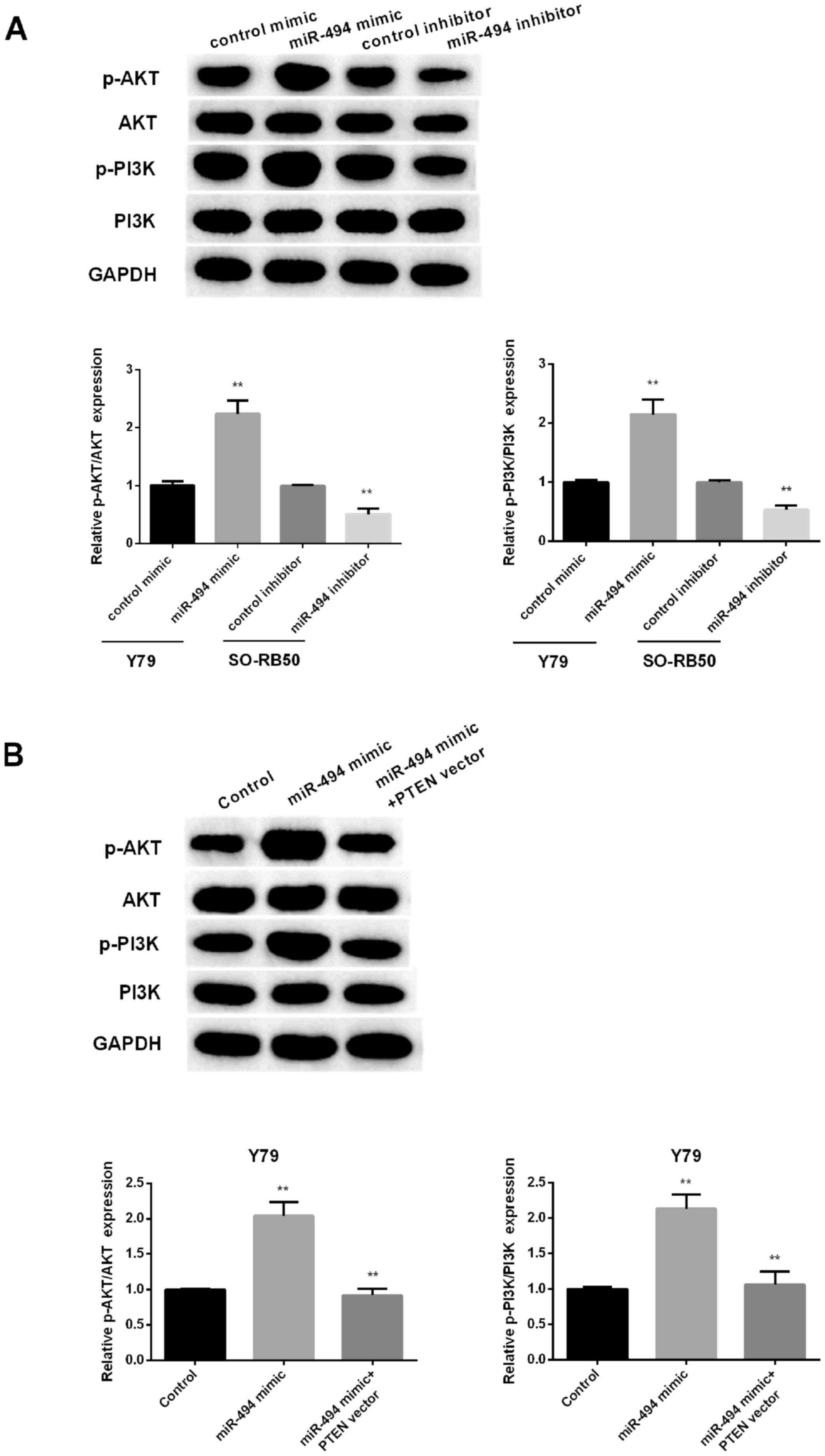
miR-494 regulates PI3K/AKT signaling
pathway via PTEN
PTEN has been reported to negatively regulate
PI3K/AKT signaling pathway.
To investigate whether a change in miR-665
expression affects the PI3K/AKT pathway in RB cells, the expression
levels of several important molecules in the pathway, including
p-AKT, AKT, p-PI3K and PI3K, were measured in RB cells following
upregulation or downregulation of miR-494. As presented in Fig. 5A, miR-494 overexpression remarkably
promoted the phosphorylation of PI3K and AKT in Y79 cells. However,
silencing miR-494 obviously inhibited the phosphorylation of PI3K
and AKT in SO-RB50 cells. Restoration of PTEN reversed effect of
miR-494 mimic on PI3K/AKT pathway (Fig.
5B). The results suggested that miR-494 promoted RB cell
progression via PTEN/PI3K/AKT signaling pathway.
Discussion
Herein, we displayed that miR-494 was increased in
RB tissue samples and cells, and it was related to the
clinicopathological features of RB patients. Moreover, increasing
miR-494 facilitated, while inhibiting miR-494, repressed RB cell
viability, invasion and migration. Furthermore, PTEN was verified
as the target of miR-494 in RB. In addition, it was demonstrated
that miR-494 regulated PI3K/AKT signaling pathway by suppressing
PTEN in RB cells. Collectively, our findings suggested that
overexpression of miR-494 promoted RB cell progression through
PTEN/PI3K/AKT axis.
In previous studies, miR-494 has been implicated in
the development and progression of various types of tumors. Zhang
et al (26) stated that
miR-494 played an oncomiR role in lung cancer progression. Zhu
et al (25) proved that
miR-494 overexpression obviously promoted endometrial cancer cell
viability, invasion and migration. Besides, miR-494 mimic
significantly enhanced nasopharyngeal carcinoma cell proliferation,
invasion and migration (27). In our
study, it was shown that miR-494 functioned as an oncogene in RB
progression by enhancing cell viability, invasion and migration. In
general, miR-494 played oncomiR roles in multiple cancers via
targeting different mRNA genes. For example, miR-494 took part in
gastric cancer cell survival by suppressing BAG-1 (28). Cheng et al (29) found that miR-494 repressed cervical
cancer cell proliferation, invasion via promoting SOCS6. Zhang
et al (30) stated that
miR-494 was higher in colorectal cancer and promoted cell
progression by targeting APC. In this study we first showed that
miR-494 regulated RB development by targeting PTEN, which was the
novelty of this research.
PTEN is essential for the maintenance of normal
cells and acts as a tumor suppressor in human cancers (31). A large number of studies have
reported that PTEN played important roles in tumor development and
progression, including lung cancer, ovarian cancer, and prostate
cancer (23,32,33).
Furthermore, in the RB development, PTEN showed an inhibitory
effect and it served as the target of multiple miRNAs. For
instance, it was the directly target of miR-93, and miR-198 in
promoting human RB progression, and the decreased PTEN expression
was also found, which was consists with our findings (34,35). In
the present study, it was shown that miR-494 targeted PTEN in
regulating RB cell progression, and PTEN overexpression can reverse
the effect of miR-494 upregulation in RB cells.
The PI3K/Akt axis is well known to participate in
the cell growth and survival of various types of cancers, including
RB (36–38). A previous study reported that the
ethanol extracts of R. japonica radix (ERJR) could play an
inhibitory effect on hepatocellular carcinoma metastasis via
PI3K/Akt signaling pathways (39).
PTEN is reported to act as an antagonist of PI3K action, it
regulates PI3K/Akt signaling negatively (40). For instance, microRNA-155-5p promoted
hepatocellular carcinoma progression by suppressing PTEN through
the PI3K/Akt pathway (41). Our
findings demonstrated that overexpression of miR-494 promoted
PI3K/Akt signaling pathways by suppressing PTEN, and PTEN
overexpression can reverse the effect of miR-494 upregulation in
PI3K/Akt signaling pathways.
Potential limitation of this study is that though
miR-494 promoted proliferation of retinoblastoma in our study, this
was only verified in RB cell lines, and mouse model might be more
convincing. For the experiments using cells transfected with
miR-494, using an untransfected control cell line to show that
transfection alone did not affect the behavior would be more
convincing. Because miR-494 is upregulated and promoted cell
viability, invasion and migration in RB, it might be a possible
biomarker for predicting progression. In summary, our study
indicated that miR-494 acted as an oncogenic microRNA in RB cells
by suppressing the tumor suppressor PTEN. miR-494 regulated
PI3K/AKT signaling pathway via PTEN.
The present study revealed the important role of
miR-494 in promoting the progression of retinoblastoma, although
further studies are required to confirm this. Taken together, our
research stated that miR-494 overexpression enhanced RB cell
progression through PTEN/PI3K/AKT signaling pathway.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FX conceived the study and drafted the manuscript.
FX, GL and LW were responsible for cell culture, qRT-PCR and
western blot analysis. XW, XJ and WB performed MTT assay and
luciferase assay. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Jinan Zhangqiu District Hospital of TCM (Jinan, China). Patients
who participated in this research had complete clinical data.
Parents of the child patients signed the informed consent form.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dimaras H, Dimba EA and Gallie BL:
Challenging the global retinoblastoma survival disparity through a
collaborative research effort. Br J Ophthalmol. 94:1415–1416. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kivelä T: The epidemiological challenge of
the most frequent eye cancer: Retinoblastoma, an issue of birth and
death. Br J Ophthalmol. 93:1129–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Narang S, Mashayekhi A, Rudich D and
Shields CL: Predictors of long-term visual outcome after
chemoreduction for management of intraocular retinoblastoma. Clin
Exp Ophthalmol. 40:736–742. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shinohara ET, DeWees T and Perkins SM:
Subsequent malignancies and their effect on survival in patients
with retinoblastoma. Pediatr Blood Cancer. 61:116–119. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al European Organisation for Research and Treatment of Cancer
Brain Tumor and Radiotherapy Groups; National Cancer Institute of
Canada Clinical Trials Group, : Radiotherapy plus concomitant and
adjuvant temozolomide for glioblastoma. N Engl J Med. 352:987–996.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Holland EC: Gliomagenesis: Genetic
alterations and mouse models. Nat Rev Genet. 2:120–129. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu S, Hu C, Wang Y, Shi G, Li Y and Wu H:
miR-124 inhibits proliferation and invasion of human retinoblastoma
cells by targeting STAT3. Oncol Rep. 36:2398–2404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Lyu X, Ma Y, Wu F and Wang L:
MicroRNA 504 targets AEG 1 and inhibits cell proliferation and
invasion in retinoblastoma. Mol Med Rep. 19:2935–2942.
2019.PubMed/NCBI
|
|
9
|
Guo L, Bai Y, Ji S and Ma H: MicroRNA 98
suppresses cell growth and invasion of retinoblastoma via targeting
the IGF1R/k Ras/Raf/MEK/ERK signaling pathway. Int J Oncol.
54:807–820. 2019.PubMed/NCBI
|
|
10
|
Golabchi K, Soleimani-Jelodar R, Aghadoost
N, Momeni F, Moridikia A, Nahand JS, Masoudifar A, Razmjoo H and
Mirzaei H: MicroRNAs in retinoblastoma: Potential diagnostic and
therapeutic biomarkers. J Cell Physiol. 233:3016–3023. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu F, Pang G and Zhao G: ANRIL acts as
onco-lncRNA by regulation of microRNA-24/c-Myc, MEK/ERK and
Wnt/β-catenin pathway in retinoblastoma. Int J Biol Macromol.
128:583–592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun Z, Zhang A and Zhang L: Inhibition of
microRNA 492 attenuates cell proliferation and invasion in
retinoblastoma via directly targeting LATS2. Mol Med Rep.
19:1965–1971. 2019.PubMed/NCBI
|
|
13
|
Wang J, Wang X, Li Z, Liu H and Teng Y:
MicroRNA-183 suppresses retinoblastoma cell growth, invasion and
migration by targeting LRP6. FEBS J. 281:1355–1365. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dalgard CL, Gonzalez M, deNiro JE and
O'Brien JM: Differential microRNA-34a expression and tumor
suppressor function in retinoblastoma cells. Invest Ophthalmol Vis
Sci. 50:4542–4551. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao JJ, Yang J, Lin J, Yao N, Zhu Y,
Zheng J, Xu J, Cheng JQ, Lin JY and Ma X: Identification of miRNAs
associated with tumorigenesis of retinoblastoma by miRNA microarray
analysis. Childs Nerv Syst. 25:13–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Yen C, Liaw D, Podsypanina K, Bose
S, Wang SI, Puc J, Miliaresis C, Rodgers L, McCombie R, et al:
PTEN, a putative protein tyrosine phosphatase gene mutated in human
brain, breast, and prostate cancer. Science. 275:1943–1947. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tay Y, Kats L, Salmena L, Weiss D, Tan SM,
Ala U, Karreth F, Poliseno L, Provero P, Di Cunto F, et al:
Coding-independent regulation of the tumor suppressor PTEN by
competing endogenous mRNAs. Cell. 147:344–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu W, Yang Z, Zhou SF and Lu N:
Posttranslational regulation of phosphatase and tensin homolog
(PTEN) and its functional impact on cancer behaviors. Drug Des
Devel Ther. 8:1745–1751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu W, Yang J, Feng X, Wang H, Ye S, Yang
P, Tan W, Wei G and Zhou Y: MicroRNA-32 (miR-32) regulates
phosphatase and tensin homologue (PTEN) expression and promotes
growth, migration, and invasion in colorectal carcinoma cells. Mol
Cancer. 12:302013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang ZX, Lu BB, Wang H, Cheng ZX and Yin
YM: MicroRNA-21 modulates chemosensitivity of breast cancer cells
to doxorubicin by targeting PTEN. Arch Med Res. 42:281–290. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma F, Zhang J, Zhong L, Wang L, Liu Y,
Wang Y, Peng L and Guo B: Upregulated microRNA-301a in breast
cancer promotes tumor metastasis by targeting PTEN and activating
Wnt/β-catenin signaling. Gene. 535:191–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rong D, Lu C, Zhang B, Fu K, Zhao S, Tang
W and Cao H: CircPSMC3 suppresses the proliferation and metastasis
of gastric cancer by acting as a competitive endogenous RNA through
sponging miR-296-5p. Mol Cancer. 18:252019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Akgun S, Kucuksayan H, Ozes ON, Can O,
Alikanoglu AS, Yildiz M and Akca H: NF-κB-induced upregulation of
miR-548as-3p increases invasion of NSCLC by targeting PTEN.
Anticancer Agents Med Chem. 19:1058–1068. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao YS, Yang WC, Xin HW, Han JX and Ma
SG: MiR-182-5p knockdown targeting PTEN inhibits cell proliferation
and invasion of breast cancer cells. Yonsei Med J. 60:148–157.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu L, Wang X, Wang T, Zhu W and Zhou X:
miR-494-3p promotes the progression of endometrial cancer by
regulating the PTEN/PI3K/AKT pathway. Mol Med Rep. 19:581–588.
2019.PubMed/NCBI
|
|
26
|
Zhang Q, Li Y, Zhao M, Lin H, Wang W, Li
D, Cui W, Zhou C, Zhong J and Huang C: MiR-494 acts as a tumor
promoter by targeting CASP2 in non-small cell lung cancer. Sci Rep.
9:30082019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He H, Liao X, Yang Q, Liu Y, Peng Y, Zhong
H, Yang J, Zhang H, Yu Z, Zuo Y, et al: MicroRNA-494-3p promotes
cell growth, migration, and invasion of nasopharyngeal carcinoma by
targeting Sox7. Technol Cancer Res Treat. Jan 1–2018.(Epub ahead of
print). doi: 10.1177/1533033818809993. View Article : Google Scholar
|
|
28
|
Shen Z, Li Y, Zhao C, Wang F, Zhou R and
Chen G: miR-494 BAG 1 axis is involved in cinobufacini induced cell
proliferation and apoptosis in gastric cancer. Mol Med Rep.
17:7435–7441. 2018.PubMed/NCBI
|
|
29
|
Cheng L, Kong B, Zhao Y and Jiang J:
miR-494 inhibits cervical cancer cell proliferation through
upregulation of SOCS6 expression. Oncol Lett. 15:3075–3080.
2018.PubMed/NCBI
|
|
30
|
Zhang Y, Guo L, Li Y, Feng GH, Teng F, Li
W and Zhou Q: MicroRNA-494 promotes cancer progression and targets
adenomatous polyposis coli in colorectal cancer. Mol Cancer.
17:12018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee YR, Chen M and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor: New modes
and prospects. Nat Rev Mol Cell Biol. 19:547–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schroeder C, Grell J, Hube-Magg C, Kluth
M, Lang D, Simon R, Höflmayer D, Minner S, Burandt E, Clauditz TS,
et al: Aberrant expression of the microtubule-associated protein
tau is an independent prognostic feature in prostate cancer. BMC
Cancer. 19:1932019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim TH, Park JH and Woo JS: Resveratrol
induces cell death through ROS dependent downregulation of
Notch1/PTEN/Akt signaling in ovarian cancer cells. Mol Med Rep.
19:3353–3360. 2019.PubMed/NCBI
|
|
34
|
Cao Y, Xia F, Wang P and Gao M: MicroRNA
93-5p promotes the progression of human retinoblastoma by
regulating the PTEN/PI3K/AKT signaling pathway. Mol Med Rep.
18:5807–5814. 2018.PubMed/NCBI
|
|
35
|
Wei D, Miao Y, Yu L, Wang D and Wang Y:
Downregulation of microRNA-198 suppresses cell proliferation and
invasion in retinoblastoma by directly targeting PTEN. Mol Med Rep.
18:595–602. 2018.PubMed/NCBI
|
|
36
|
Lin A, Piao HL, Zhuang L, Sarbassov D, Ma
L and Gan B: FoxO transcription factors promote AKT Ser473
phosphorylation and renal tumor growth in response to pharmacologic
inhibition of the PI3K-AKT pathway. Cancer Res. 74:1682–1693. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Spangle JM, Roberts TM and Zhao JJ: The
emerging role of PI3K/AKT-mediated epigenetic regulation in cancer.
Biochim Biophys Acta Rev Cancer. 1868:123–131. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xie C, Lu H, Nomura A, Hanse EA, Forster
CL, Parker JB, Linden MA, Karasch C and Hallstrom TC: Co-deleting
Pten with Rb in retinal progenitor cells in mice results in fully
penetrant bilateral retinoblastomas. Mol Cancer. 14:932015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim BR, Ha J, Lee S, Park J and Cho S:
Anti-cancer effects of ethanol extract of Reynoutria
japonica Houtt. radix in human hepatocellular carcinoma cells
via inhibition of MAPK and PI3K/Akt signaling pathways. J
Ethnopharmacol. 245:1121792019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Haddadi N, Lin Y, Travis G, Simpson AM,
Nassif NT and McGowan EM: PTEN/PTENP1: ‘Regulating the regulator of
RTK-dependent PI3K/Akt signalling’, new targets for cancer therapy.
Mol Cancer. 17:372018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fu X, Wen H, Jing L, Yang Y, Wang W, Liang
X, Nan K, Yao Y and Tian T: MicroRNA-155-5p promotes hepatocellular
carcinoma progression by suppressing PTEN through the PI3K/Akt
pathway. Cancer Sci. 108:620–631. 2017. View Article : Google Scholar : PubMed/NCBI
|