Introduction
Hydrogen gas (H2), as a type of
endogenous gas, has been demonstrated to not only serve as a
crucial energy source, but also to exert crucial roles in
physiological regulation (1).
Hydrogen molecules can enter tissues and exert anti-inflammatory,
antioxidant and anti-apoptotic roles (2). Notably, H2 shows high safety
and efficacy to patients in a clinical setting (3). Intravenous administration of 500 ml
H2 significantly improved the erythema and associated
symptoms in 4 patients with acute erythematous skin diseases, with
no changes in the physiological parameters, such as body
temperature, blood pressure and pulse rate, or deterioration of
liver and kidney function. In addition, two volunteers (one for
intravenous H2 administration and the other for
H2 inhalation) demonstrated no discomfort (3). Recently, H2 has been also
applied in a clinical setting to improve the hearing loss induced
by radiotherapy in patients with nasopharyngeal cancer (ClinicalTrials.gov identifier, NCT03818347) (4). In that study, H2 (67%
hydrogen and 33% oxygen) was generated using the hydrogen-oxygen
nebulizer through the electrolysis of water and the results
demonstrated that hydrogen-oxygen therapy could markedly improve
binaural hearing (4). Another Phase
1 trial (ClinicalTrials.gov identifier,
NCT04046211) aims to explore the safety of H2 on healthy
adult volunteers. In the study, 8 volunteers were included, 2 of
which were exposed to 2.4% H2 in medical air for 24 h, 2
who were exposed to the same gas for 48 h and the other 4 patients
exposed to the same gas for 72 h (https://clinicaltrials.gov/ct2/show/NCT04046211). In
addition, Dole et al (5)
proposed in 1975, for the first time, that H2 has
anti-cancer potential. Similarly, our research group previously
found that H2 administration significantly represses
cell growth, migration and invasion capacities and induced cell
apoptosis in lung cancer A549 and H1975 cells (6). However, the molecular mechanisms remain
largely unknown.
Autophagy and apoptosis are indispensable biological
processes and serve crucial roles in individual development
(7). Autophagy, also known as
macroautophagy, is a ‘self-eating’ process, which engulfs
cytoplasmic proteins, complexes or organelles into the
autophagosome (a cytoplasmic double membrane structure), leading to
degradation and recycling (8).
Autophagy is modulated by autophagy-related genes (ATGs) through
two evolutionarily conserved ubiquitin-like conjugation systems,
known as the ATG12-ATG5 and the ATG8 light chain 3
(LC3)-phosphatidylethanolamine (PE) systems. Microtubule-associated
protein 1A/1B LC3BI is conjugated with PE to become LC3BII, which
associates with the outer and inner membranes of the autophagosome
(9). It is known that autophagy
maintains cellular homeostasis and protects against various
diseases, including cancer (10–12).
However, its role in carcinogenesis is controversial. Accumulating
evidence has found that autophagy to be a ‘double-edge sword’ in
the progression of cancer (13,14).
Signal transducer and activator of transcription
(STAT) 3 is a latent transcription factor which modulates
extracellular signals by interacting with polypeptide receptors
(15). STAT3 protein becomes
transcriptionally activated primarily through tyrosine
phosphorylation, which then dimerizes, translocates to the nucleus
and binds to sequence-specific DNA elements, leading to the
transcription of target genes (16).
STAT3 has been found to serve as an oncogene and frequently
hyper-activated in a number of types of cancer, including lung
cancer, contributing to cancer cell survival, proliferation,
metastasis and angiogenesis (17,18). In
addition, STAT3 serves a vital role in the process of autophagy
(19). For example, Guo et al
(20) reported that repression of
STAT3 via its inhibitor, isocryptotanshinone enhances autophagy in
the A549 lung cancer cell line.
The present study aimed to investigate the effects
of H2 on autophagy and apoptosis in lung cancer cells,
and the role of STAT3 in this process. The results demonstrated
that H2 could promote lung cancer cell apoptosis and
autophagy by inhibiting the activation of the STAT3/Bcl2 signaling
pathway and suppression of autophagy could enhance H2
roles in promoting lung cancer cell apoptosis.
Materials and methods
Cell lines and culture
The human A549 and H1975 lung cancer cell lines,
were purchased from American Type Culture Collection. The A549 cell
line was cultured in F-12K medium, supplemented with 10% fetal
bovine serum (FBS), while the H1975 cell line was cultured in
RPMI-1640 medium containing 10% FBS (all from Gibco; Thermo Fisher
Scientific, Inc.). The two cell lines were maintained in an
incubator at 37°C with 5% CO2.
Cell transfection and treatments
Three short interfering (si)RNAs used to silence
Beclin1 (si-Beclin1-1/2-3; cat. no. SR322490) in the A549 and H1975
cells, and the negative control vector (si-NC, cat. no. SR322490)
were purchased from OriGene Technologies, Inc.. The overexpressing
plasmid STAT3 (OE-STAT3, cat. no. SC124165) and OE-NC (cat. no.
SC124165) were also obtained from OriGene Technologies, Inc.. Cell
transfection was performed using the transfection reagent
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols with 2
µg OE-STAT3/OE-NC or 0.3 µg si-Beclin1/si-NC for each well of a
6-well plate. Following 48 h of transfection, cells were harvested
for analysis. The siRNA sequences were as follow: si-NC, Forward:
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse:
5′-ACGUGACACGUUCGGAGAATT-3′; si-Beclin1-1, forward:
5′-CUGGACACGAGUUUCAAGATT-3′ and reverse:
5′-UCUUGAAACUCGUGUCCAGTT-3′; si-Beclin1-2, forward:
5′-GUGGAAUGGAAUGAGAUUATT-3′ and reverse:
5′-UAAUCUCAUUCCAUUCCACTT-3′; si-Beclin1-3, forward:
5′-GCUGCCGUUAUACUGUUCUTT-3′ and reverse:
5′-AGAACAGUAUAACGGCAGCTT-3′.
For H2 treatment, A549 and H1975 cells
were cultured in different concentrations of H2 (20, 40
and 60%) with the assistance of Shanghai Nanobubble Technology Co.,
Ltd. (http://www.nanobubble.cn/p-about.html?app=mb) for
different time points (12, 24, 36, 48 or 72 h), and 5%
CO2 served as the negative control.
A549 and H1975 cells were treated with 2 mM
3-methyladenine (3-MA) or 100 µM rapamycin (RAPA) for 48 h to
repress and induce autophagy, respectively.
Western blot analysis
Total protein was extracted from the cells using the
RIPA lysis buffer (Sangon Biotech Co., Ltd.), containing protease
inhibitor (Beyotime Institute of Biotechnology) and according to
the manufacturer's instructions. After quantification with a
bicinchoninic acid protein kit (Bio-Rad Laboratories, Inc.), 30 µg
protein from each sample was loaded per lane and separated using
10% SDS-PAGE. Then, the proteins were transferred onto
polyvinylidene difluoride membranes (EMD Millipore), and blocked
with 5% skimmed milk at room temperature for 1 h, following which
the membranes were incubated with the following primary antibodies,
overnight at 4°C: Cleaved-caspase 3 (1:1,000 dilution; cat. no.
ab2302; Abcam), cleaved poly ADP-ribose polymerase (PARP; 1:1,000
dilution; cat. no. ab32064; Abcam), Beclin1 (1:2,000 dilution; cat.
no. ab207612; Abcam), p62 (1:1,000 dilution; cat. no. ab56416;
Abcam), LC3B (1:3,000 dilution; cat. no. ab51520; Abcam), Bcl2
(1:1,000 dilution; cat. no. 15071; Cell Signaling Technology,
Inc.), STAT3 (1:1,000 dilution; cat. no. 9139; Cell Signaling
Technology, Inc.), phosphorylated (p)-STAT3 (1:1,000 dilution; cat.
no. 9145; Cell Signaling Technology, Inc.) and β-actin (1:4,000
dilution; cat. no. 3700; Cell Signaling Technology, Inc.). After
the incubation with the corresponding secondary antibodies (Santa
Cruz Biotechnology, Inc.), the protein expression levels were
detected using a western blot imaging and quantitative system
(Bio-Rad Laboratories, Inc.). Protein quantification was performed
using the ImageJ software (version 1.48; National Institutes of
Health) after background subtraction, with β-actin as an internal
reference.
Cell counting Kit-8 assay (CCK-8)
A CCK-8 assay was used to determine cell
proliferation ability according to the manufacturer's instructions.
In brief, A549 and H1975 cells were cultured in 96-well plates at a
density of 2×103 cells/well, overnight at 37°C. Then,
the cells were treated with different concentrations of
H2 for 12, 24, 36, 48 or 72 h. Following which, the cell
culture medium was replaced with 10 µl CCK-8 reagent (Beyotime
Institute of Biotechnology) and 90 µl fresh medium, and incubated
at 37°C for another 4 h. The absorbance at 450 nm was determined
using a plate reader (model 680; Bio-Rad Laboratories, Inc.).
Flow cytometry assay
Cell apoptosis was determined using flow cytometry
and an Annexin V-FITC/propidium iodide (PI) kit (Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's protocols.
Briefly, A549 and H1975 cells were harvested through centrifugation
at 110 × g for 5 min at 4°C and washed once with PBS following 48 h
of H2 treatment and/or cell transfection. Then, the
cells were incubated with Annexin V-FITC and PI solution for 15 min
in the dark, at room temperature. The fluorescent signals were
evaluated using flow cytometry (BD Biosciences) within 1 h of
staining using Flowjo version 7.6 software (FlowJo LLC). Cells in
the FITC−/PI− quadrant represented live
cells, while FITC+/PI− represented early
apoptotic cells and FITC+/PI+ represented
late apoptotic cells.
Statistical analysis
Each experiment was performed in triplicate. Data
are presented the mean ± standard deviation. Statistically
significant comparisons between two groups and multiple groups were
performed using unpaired Student's t-test and one-way ANOVA,
followed by Dunnett's or Tukey's post hoc test. Data analysis was
performed using the SPSS software (version 23.0, IBM Corp.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
H2 treatment induces
significant increases in cell apoptosis and autophagy
The present study first investigated the effects of
H2 treatment on the apoptosis and autophagy of lung
cancer cells. Compared with that in the control group, cell
viability was reduced following treatment with H2, in a
time-dependent manner, in both A549 and H1975 cells (Fig. 1A), along with increased apoptosis
rates (Fig. 1B and C). The
expression levels of pro-apoptotic proteins, including
cleaved-caspase 3 and cleaved-PARP were significantly increased
followed H2 treatment compared with that in the control
group (Fig. 1D and E), in a
dose-dependent manner, in both cell lines. In addition, Beclin1
expression and the ratio of LC3BII/I were also significantly
increased following H2 treatment, while the expression
of p62 was significantly reduced (Fig.
1D and E), also in a dose-dependent manner. These findings
demonstrated that H2 could induce cell apoptosis and
autophagy in lung cancer cell lines.
Autophagy weakens the role of
H2 in promoting cell apoptosis in lung cancer cells
The autophagy roles in H2-induced cell
apoptosis in lung cancer cells were subsequently investigated. The
protein expression levels of Beclin1 and LC3BII/I were
significantly increased, while that of p62 was reduced when A549
and H1975 cells were treated with RAPA, an autophagy inducer
(Fig. 2A). As 60% H2
resulted in significant changes in cell viability and apoptosis,
60% H2 was selected for the subsequent experiments.
Following RAPA treatment, cell apoptosis rates (Fig. 2B) and the protein expression levels
of cleaved-caspase 3 and cleaved-PARP (Fig. 2C and D) were significantly decreased
compared with that in the H2 group. By contrast, the
protein expression levels of Beclin1 and LC3BII/I were
significantly decreased and p62 expression was increased following
cell treatment with 3-MA, an autophagy repressor (Fig. 2E). In addition, 3-MA treatment
enhanced H2-mediated increases in cell apoptosis rates
(Fig. 2F) and the protein expression
levels of cleaved-caspase 3 and cleaved-PARP as compared with the
H2 group (Fig. 2G and H).
These results revealed that autophagy weakened H2 roles
in inducing cell apoptosis in lung cancer cells.
 | Figure 2.Evaluation of autophagy effects on
H2-induced lung cancer cell apoptosis. (A) The protein
expression levels of p62, Beclin1, LC3BII and LC3BI, following
treatment of A549 and H1975 cells with RAPA were detected using
western blot analysis. (B) Cell apoptosis was investigated using a
flow cytometry assay. Western blot analysis of the expression of
cleaved-caspase 3 and cleaved-PARP in (C) A549 and (D) H1975 cells
treated with H2 or H2+RAPA. (E) The protein
expression levels of p62, Beclin1, LC3BII and LC3BI were detected
using western blot analysis following treatment of A549 and H1975
cells with 3-MA. (F) Cell apoptosis was investigated using flow
cytometry. Western blot analysis of the protein expression levels
of cleaved-caspase 3 and cleaved-PARP following treatment of (G)
A549 and (H) H1975 cells with H2 or H2+3-MA.
*P<0.05. **P<0.01. LC3, light chain 3; RAPA, rapamycin; PARP,
poly ADP-ribose polymerase; PI, propidium iodide; 3-MA,
3-methyladenine; H2, hydrogen gas. |
Knockdown of Beclin1 enhances the
roles of H2 in promoting cell apoptosis in lung cancer
cell lines
To further clarify the role of autophagy in
H2-mediated lung cancer cell apoptosis, knockdown
experiments were performed. The expression of Beclin1 was
significantly decreased in A549 and H1975 cells transfected with
the siRNAs targeting the human Beclin1 gene. As si-Beclin1-3
significantly reduced the protein expression level of Beclin1 among
the 3 siRNAs (Fig. 3A), this was
chosen for the subsequent experiments. Following cell transfection
with si-Beclin1-3, the protein expression level of p62 was
increased and the expression ratio of LC3BII:LC3BI was decreased
(Fig. 3B and C). Notably, cell
apoptosis induced by H2 treatment was significantly
increased when Beclin1 was silenced in both A549 and H1975 cells as
compared with the H2 group (Fig. 3D). In addition, knockdown of Beclin1
increased the protein expression levels of cleaved-caspase 3 and
cleaved-PARP in A549 and H1975 cells following H2
treatment compared with cells treated with H2 only
(Fig. 3E and F). These findings
further confirmed that autophagy inhibition could enhance the
effects of H2 on inducing cell apoptosis in lung cancer
cell lines.
H2 treatment induces lung
cancer cell apoptosis and autophagy by repressing the STAT3/Bcl2
signaling pathway
Next, the roles of the STAT3/Bcl2 signaling pathway
in H2-mediated cell apoptosis and autophagy in lung
cancer cell lines were investigated. The protein expression levels
of p-STAT3 and Bcl2 were significantly decreased following
H2 treatment in H1975 and A549 cells in a dose-dependent
manner (Fig. 4A and B), whereas
STAT3-overexpression abolished this effect (Fig. 4C-E). In addition, overexpression of
STAT3 in H2-treated cells inhibited lung cancer cell
apoptosis and neutralized H2-mediated increases in
apoptosis (Fig. 4F and G), along
with the decreased protein expression levels of cleaved-caspase 3,
cleaved-PARP, Beclin1, LC3BII/I and the increased expression of p62
(all vs. H2 group; Fig. 4H
and I). The aforementioned findings suggested that
H2 treatment promoted lung cancer cell apoptosis and
autophagy by repressing the activation of the STAT3/Bcl2 signaling
pathway.
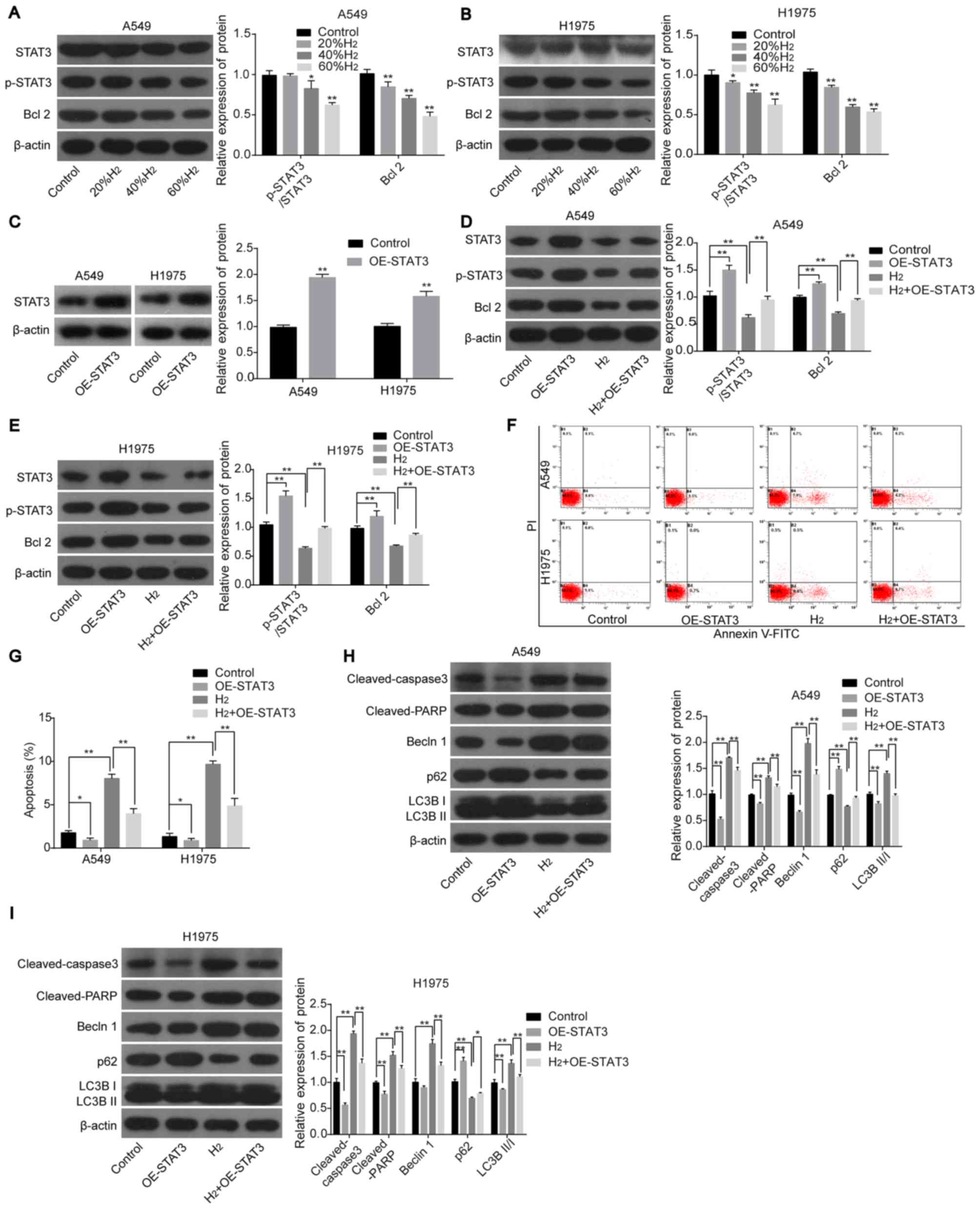 | Figure 4.H2 treatment increases
lung cancer cell apoptosis and autophagy by repressing the
activation of the STAT3/Bcl2 signaling pathway. The protein
expression levels of STAT3, p-STAT3 and Bcl2 were detected using
western blot analysis in (A) A549 and (B) H1975 cells following
treatment with different concentrations of H2. (C) STAT3
expression was detected using western blot analysis following
transfection with OE-STAT3 or OE-NC. The protein expression levels
of STAT3, p-STAT3 and Bcl2 were detected using western blot
analysis in (D) A549 and (E) H1975 cells following treatment with
H2 and/or transfected with OE-STAT3. Cell apoptosis was
investigated using (F) flow cytometry and the results were
subsequently (G) quantified. The protein expression levels of
cleaved-caspase 3, cleaved-PARP, Beclin1, p62, LC3BII and LC3BI in
(H) A549 and (I) H1975 cells were detected using western blot
analysis following transfection with OE-STAT3 and/or treated with
H2. *P<0.05 and **P<0.01. STAT, signal transducer
and activator of transcription; p-, phosphorylated; PARP, poly
ADP-ribose polymerase; LC3, light chain 3; H2, hydrogen
gas; OE, overexpression; NC, negative control |
Discussion
As a colorless, odorless, tasteless, non-toxic and
highly combustible gas, H2 exerts multiple roles,
including anti-reactive oxygen species, anti-inflammation and
antitumor (21). Increasing evidence
has revealed that H2 not only improved the side effects
induced by chemotherapeutics, but also inhibited cancer growth
(22–24). In addition, Chen et al
(25) reported that H2
treatment reduced the metastases size in the abdominal cavity,
improved anemia and hypoalbuminemia and gradually returned the
lymphocyte and tumor marker levels, including CA19-9, α fetoprotein
and carcinoembryonic antigen to normal. In our previous study, it
was found that H2 treatment effectively inhibited lung
cancer cell growth and induced cell apoptosis by targeting the
structural maintenance of chromosomes 3 gene in a dose-dependent
manner, with no side effects on the non-tumor burdened mice
(6). The current study demonstrated
that H2 treatment repressed cell growth and increased
cell apoptosis by inhibiting the STAT3/Bcl2 signaling pathway in
A549 and H1975 lung cancer cell lines.
Autophagy and apoptosis, which lead to the
degradation of proteins and organelles or cell death upon cellular
stress, serve vital roles in the progression of lung cancer
(26). Previous studies indicated
that autophagy serves as a ‘double-edged sword’ in lung cancer cell
apoptosis (26). Typically,
autophagy repression decreases the cell's ability to overcome
stress and maintain homeostasis (27). However, several studies have reported
that autophagy induced cell apoptosis (28–30).
Autophagy exerts an oncogenic or a tumor suppressive role in
carcinogenesis (31). In lung
cancer, inhibition of autophagy enhances the anti-angiogenic
property of anlotinib (32),
suggesting an angiogenic role of autophagy in lung cancer. Yin
et al (33) reported that
inhibition of autophagy induced by mucolipin-1 (which is a member
of the transient receptor potential cation channel family)
downregulation repressed the progression of lung cancer. Li et
al (34), demonstrated that
autophagy enhancement caused by apurinic endonuclease 1
significantly increased the resistance of lung cancer cells to
cisplatin. These findings suggested that autophagy may exert an
oncogenic role in lung cancer progression. By contrast, Fan et
al (35) reported that Bruceine
D, a quassinoid compound, which can be extracted from the seeds of
Brucea javanica, inhibits lung cancer progression by
inducing cell apoptosis and autophagy, suggesting a suppressive
role of autophagy in lung cancer progression. These findings
indicate that autophagy serves an important role in lung cancer
progression, therefore autophagy plays a role in
H2-induced cell apoptosis in lung cancer cell lines.
When autophagy is induced, Beclin1 and LC3 are
distributed to the autophagosome membrane to trigger the formation
of autophagosome, whereas p62 expression is decreased (36,37). The
transformation of LC3I to LC3BII is the most common autophagosome
marker, as the amount of LC3BII reflects the number of
autophagosomes and autophagy-related structures (38). The present study found that
H2 treatment significantly increased the protein
expression level of Beclin1, and the conversion of LC3BI to LC3BII,
and decreased the expression level of p62, indicating that
H2 treatment could trigger lung cancer cell autophagy.
Several studies have investigated the role of H2
treatment in autophagy. In detail, Guan et al (39) reported that H2 treatment
induced autophagy and protected Sprague-Dawley rats against chronic
intermittent hypoxia-induced renal dysfunction. In addition,
inhaling 2% H2 resulted in an increase in the expression
of LC3BII and attenuated sepsis-induced liver injury (40). However, in a myocardial ischemia
reperfusion model of rats, H2 inhalation significantly
reduced the ischemic size and serum troponin I level by repressing
autophagy (41). The effects might
be tissue specific.
In addition, the present study also investigated the
role of autophagy in H2-mediated cell apoptosis. The
results demonstrated that autophagy enhancement by RAPA stimulation
significantly weakened H2-mediated cell apoptosis in
both A549 and H1975 lung cancer cell lines, whereas autophagy
inhibition by 3-MA treatment or siRNA-Beclin1 transfection
significantly enhanced the roles of H2 in promoting lung
cancer cell apoptosis, indicating that autophagy inhibition could
enhance the antitumor role of H2 in lung cancer.
As the STAT3/Bcl2 signaling pathway serves an
important role in cell autophagy and apoptosis (42,43), the
present study also investigated the effects of the STAT3/Bcl2
signaling pathway on H2-mediated lung cancer cell
autophagy and apoptosis. It was found that the phosphorylation
protein expression levels of STAT3 and Bcl2 were significantly
inhibited when A549 and H1975 cells were treated with
H2, suggesting that H2 could repress the
activation of the STAT3/Bcl2 signaling pathway. Consistent with
this result, Bai et al (44)
found that hydrogen-rich saline treatment increased LC3B and
Beclin1 protein expression levels and decreased the protein
phosphorylation level of STAT3 in hypoxic-ischemic brain damaged
mice. To further investigate the roles of STAT3/Bcl2 in
H2-mediated cell apoptosis and autophagy in lung cancer,
rescue experiments were also performed in the present study. The
results demonstrated that overexpression of STAT3 neutralized the
H2 roles in increasing cell autophagy and apoptosis,
suggesting that H2 induced lung cancer apoptosis and
autophagy by inhibiting the STAT3/Bcl2 signaling pathway.
In summary, the present study revealed that
H2 could promote lung cancer cell apoptosis and
autophagy by inhibiting the STAT3/Bcl2 signaling pathway and that
suppression of autophagy could enhance the roles of H2
in promoting lung cancer cell apoptosis. The current study might
provide a novel application of H2 in the treatment of
lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Excellent
Personnel Training Program (Studying of the Molecular Mechanism and
Clinical Transformation of Hydrogen in the Treatment of Lung
Cancer; grant no. zh2018006).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
GC conceived and designed the project. LL performed
the majority of the experiments and wrote the paper. ZY analyzed
the data. YW and JM performed parts of the experiments. All authors
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhao P, Jin Z, Chen Q, Yang T, Chen D,
Meng J, Lu X, Gu Z and He Q: Local generation of hydrogen for
enhanced photothermal therapy. Nat Commun. 9:42412018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohsawa I, Ishikawa M, Takahashi K,
Watanabe M, Nishimaki K, Yamagata K, Katsura K, Katayama Y, Asoh S
and Ohta S: Hydrogen acts as a therapeutic antioxidant by
selectively reducing cytotoxic oxygen radicals. Nat Med.
13:688–694. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ono H, Nishijima Y, Adachi N, Sakamoto M,
Kudo Y, Nakazawa J, Kaneko K and Nakao A: Hydrogen(H2) treatment
for acute erythymatous skin diseases. A report of 4 patients with
safety data and a non-controlled feasibility study with H2
concentration measurement on two volunteers. Med Gas Res. 2:142012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen J, Kong X, Mu F, Lu T, Du D and Xu K:
Hydrogen-oxygen therapy can alleviate radiotherapy-induced hearing
loss in patients with nasopharyngeal cancer. Ann Palliat Med.
8:746–751. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dole M, Wilson FR and Fife WP: Hyperbaric
hydrogen therapy: A possible treatment for cancer. Science.
190:152–154. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang D, Wang L, Zhang Y, Zhao Y and Chen
G: Hydrogen gas inhibits lung cancer progression through targeting
SMC3. Biomed Pharmacother. 104:788–797. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gordy C and He YW: The crosstalk between
autophagy and apoptosis: Where does this lead? Protein Cell.
3:17–27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sotthibundhu A, McDonagh K, von Kriegsheim
A, Garcia-Munoz A, Klawiter A, Thompson K, Chauhan KD, Krawczyk J,
McInerney V, Dockery P, et al: Rapamycin regulates autophagy and
cell adhesion in induced pluripotent stem cells. Stem Cell Res
Ther. 7:1662016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Boya P, Reggiori F and Codogno P: Emerging
regulation and functions of autophagy. Nat Cell Biol. 15:713–720.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Levine B: Cell biology: Autophagy and
cancer. Nature. 446:745–747. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Galluzzi L, Pietrocola F, Bravo-San Pedro
JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J,
Gewirtz DA, Karantza V, et al: Autophagy in malignant
transformation and cancer progression. EMBO J. 34:856–880. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wasik AM, Grabarek J, Pantovic A,
Cieślar-Pobuda A, Asgari HR, Bundgaard-Nielsen C, Rafat M, Dixon
IM, Ghavami S and Łos MJ: Reprogramming and
carcinogenesis-parallels and distinctions. Int Rev Cell Mol Biol.
308:167–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sridhar S, Botbol Y, Macian F and Cuervo
AM: Autophagy and disease: Always two sides to a problem. J Pathol.
226:255–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Levy DE and Darnell JE Jr: Stats:
Transcriptional control and biological impact. Nat Rev Mol Cell
Biol. 3:651–662. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Akira S, Nishio Y, Inoue M, Wang XJ, Wei
S, Matsusaka T, Yoshida K, Sudo T, Naruto M and Kishimoto T:
Molecular cloning of APRF, a novel IFN-stimulated gene factor 3
p91-related transcription factor involved in the gp130-mediated
signaling pathway. Cell. 77:63–71. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tong M, Wang J, Jiang N, Pan H and Li D:
Correlation between p-STAT3 overexpression and prognosis in lung
cancer: A systematic review and meta-analysis. PLoS One.
12:e01822822017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
You L, Wang Z, Li H, Shou J, Jing Z, Xie
J, Sui X, Pan H and Han W: The role of STAT3 in autophagy.
Autophagy. 11:729–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo S, Luo W, Liu L, Pang X, Zhu H, Liu A,
Lu J, Ma DL, Leung CH, Wang Y and Chen X: Isocryptotanshinone, a
STAT3 inhibitor, induces apoptosis and pro-death autophagy in A549
lung cancer cells. J Drug Target. 24:934–942. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li S, Liao R, Sheng X, Luo X, Zhang X, Wen
X, Zhou J and Peng K: Hydrogen gas in cancer treatment. Front
Oncol. 9:6962019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li FY, Zhu SX, Wang ZP, Wang H, Zhao Y and
Chen GP: Consumption of hydrogen-rich water protects against ferric
nitrilotriacetate-induced nephrotoxicity and early tumor
promotional events in rats. Food Chem Toxicol. 61:248–254. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou P, Lin B, Wang P, Pan T, Wang S, Chen
W, Cheng S and Liu S: The healing effect of hydrogen-rich water on
acute radiation-induced skin injury in rats. J Radiat Res.
60:17–22. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu Y, Yuan M, Song J, Chen X and Yang H:
Hydrogen gas from inflammation treatment to cancer therapy. ACS
Nano. 13:8505–8511. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen JB, Pan ZB, Du DM, Qian W, Ma YY, Mu
F and Xu KC: Hydrogen gas therapy induced shrinkage of metastatic
gallbladder cancer: A case report. World J Clin Cases. 7:2065–2074.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu G, Pei F, Yang F, Li L, Amin AD, Liu
S, Buchan JR and Cho WC: Role of autophagy and apoptosis in
non-small-cell lung cancer. Int J Mol Sci. 18:3672017. View Article : Google Scholar
|
|
27
|
Kroemer G, Mariño G and Levine B:
Autophagy and the integrated stress response. Mol Cell. 40:280–293.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dang S, Yu ZM, Zhang CY, Zheng J, Li KL,
Wu Y, Qian LL, Yang ZY, Li XR, Zhang Y and Wang RX: Autophagy
promotes apoptosis of mesenchymal stem cells under inflammatory
microenvironment. Stem Cell Res Ther. 6:2472015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang M, Su L, Xiao Z and Liu X and Liu X:
Methyl jasmonate induces apoptosis and pro-apoptotic autophagy via
the ROS pathway in human non-small cell lung cancer. Am J Cancer
Res. 6:187–199. 2016.PubMed/NCBI
|
|
30
|
Galluzzi L, Bravo-San Pedro JM, Vitale I,
Aaronson SA, Abrams JM, Adam D, Alnemri ES, Altucci L, Andrews D,
Annicchiarico-Petruzzelli M, et al: Essential versus accessory
aspects of cell death: Recommendations of the NCCD 2015. Cell Death
Differ. 22:58–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
White E: The role for autophagy in cancer.
J Clin Invest. 125:42–46. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liang L, Hui K, Hu C, Wen Y, Yang S, Zhu
P, Wang L, Xia Y, Qiao Y, Sun W, et al: Autophagy inhibition
potentiates the anti-angiogenic property of multikinase inhibitor
anlotinib through JAK2/STAT3/VEGFA signaling in non-small cell lung
cancer cells. J Exp Clin Cancer Res. 38:712019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yin C, Zhang H, Liu X, Zhang H, Zhang Y,
Bai X, Wang L, Li H, Li X, Zhang S, et al: Downregulated MCOLN1
attenuates the progression of non-small-cell lung cancer by
inhibiting lysosome-autophagy. Cancer Manag Res. 11:8607–8617.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li Z, Wang Y, Wu L, Dong Y, Zhang J, Chen
F, Xie W, Huang J and Lu N: Apurinic endonuclease 1 promotes the
cisplatin resistance of lung cancer cells by inducing
Parkin-mediated mitophagy. Oncol Rep. 42:2245–2254. 2019.PubMed/NCBI
|
|
35
|
Fan J, Ren D, Wang J, Liu X, Zhang H, Wu M
and Yang G: Bruceine D induces lung cancer cell apoptosis and
autophagy via the ROS/MAPK signaling pathway in vitro and in vivo.
Cell Death Dis. 11:1262020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
El-Khattouti A, Selimovic D, Haikel Y and
Hassan M: Crosstalk between apoptosis and autophagy: Molecular
mechanisms and therapeutic strategies in cancer. J Cell Death.
6:37–55. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thorburn A: Apoptosis and autophagy:
Regulatory connections between two supposedly different processes.
Apoptosis. 13:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gupta NA, Kolachala VL, Jiang R,
Abramowsky C, Shenoi A, Kosters A, Pavuluri H, Anania F and Kirk
AD: Mitigation of autophagy ameliorates hepatocellular damage
following ischemia-reperfusion injury in murine steatotic liver. Am
J Physiol Gastrointest Liver Physiol. 307:G1088–G1099. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Guan P, Sun ZM, Luo LF, Zhou J, Yang S,
Zhao YS, Yu FY, An JR, Wang N and Ji ES: Hydrogen protects against
chronic intermittent hypoxia induced renal dysfunction by promoting
autophagy and alleviating apoptosis. Life Sci. 225:46–54. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan M and Yu Y, Mao X, Feng J, Wang Y,
Chen H, Xie K and Yu Y: Hydrogen gas inhalation attenuates
sepsis-induced liver injury in a FUNDC1-dependent manner. Int
Immunopharmacol. 71:61–67. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gao Y, Yang H, Chi J, Xu Q, Zhao L and
Yang W, Liu W and Yang W: Hydrogen gas attenuates myocardial
ischemia reperfusion injury independent of postconditioning in rats
by attenuating endoplasmic reticulum stress-induced autophagy. Cell
Physiol Biochem. 43:1503–1514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu Y, Gong W, Yang ZY, Zhou XS, Gong C,
Zhang TR, Wei X, Ma D, Ye F and Gao QL: Quercetin induces
protective autophagy and apoptosis through ER stress via the
p-STAT3/Bcl-2 axis in ovarian cancer. Apoptosis. 22:544–557. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu K, Ren T, Huang Y, Sun K, Bao X, Wang
S, Zheng B and Guo W: Apatinib promotes autophagy and apoptosis
through VEGFR2/STAT3/BCL-2 signaling in osteosarcoma. Cell Death
Dis. 8:e30152017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bai X, Liu S, Yuan L, Xie Y, Li T, Wang L,
Wang X, Zhang T, Qin S, Song G, et al: Hydrogen-rich saline
mediates neuroprotection through the regulation of endoplasmic
reticulum stress and autophagy under hypoxia-ischemia neonatal
brain injury in mice. Brain Res. 1646:410–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
















