Introduction
Gliomas account for 80% of all malignant primary
brain tumors, with an annual incidence of approximately 6 per
100,000 in the USA (1). Despite
great advances in therapeutic strategies in the past decade,
including surgical resection, radiotherapy and chemotherapy, the
prognosis of patients with glioma remains unsatisfactory due to
high metastasis and recurrence, with a median survival of only
10–14 months (2). Therefore, it is
important to understand the molecular mechanisms underlying
tumorigenesis and progression of glioma in order to identify novel
therapeutic targets.
The ERK signaling pathway is one of the most
frequently activated pathways in various human tumors, including
glioma. The ERK pathway is involved in regulation of multiple
biological functions, including cell proliferation,
differentiation, apoptosis, migration and invasion (3–6). Tumor
metastasis requires tumor cell-derived proteases, especially matrix
metalloproteinases (MMPs), to degrade the extracellular matrix
(ECM) (7,8). MMPs have been demonstrated to be
crucial for the invasive and metastatic potential of a variety of
malignant tumors, such as breast, lung and pancreatic cancer
(9). Among the MMPs, the present
study focused on MMP2 and MMP9, which were shown to promote glioma
cell motility (10). However, the
underlying mechanism of MMP2 and MMP9 in glioma remains
unclear.
Rap proteins are a subgroup of the Ras family; a
group of small guanosine 5′ triphosphate (GTP) binding proteins
with five different isoforms: Rap1A, Rap1B, Rap2A, Rap2B and Rap2C
(11). Rap2B, a member of the Ras
oncogene family, is highly expressed in diverse types of tumors,
including colorectal cancer (12),
hepatocellular carcinoma (13) and
lung cancer (14). Similar to other
Ras proteins, Rap2B can be activated and inhibited by a variety of
external and internal inducers, circulating between its GTP-bound
active state and GDP-bound inactive state (15). Several studies have reported that
Rap2B has a critical regulatory role in a series of cellular
biological processes, such as cell adhesion, proliferation,
migration and invasion, as well as signal transduction in human
cells (16,17). The association between Rap2B and
tumorigenesis has been previously confirmed, for example Rap2B
promotes the tumor growth in vivo in renal cell carcinoma
(18) and, in prostate cancer, Rap2B
regulates tumor growth and metastasis through the focal adhesion
kinase pathway (19). Additionally,
Rap2B is a direct target gene of p53 and functions in a
p53-dependent manner in the regulation of apoptosis and migration
(20). Zhang et al (21) reported that miR-194 suppresses cell
proliferation and invasion of bladder cancer by targeting Rap2B
expression. Miao et al (22)
showed that Rap2B-knockdown inhibits the malignant behaviors of
glioma cells through the NF-κB pathway. However, the molecular
mechanism of Rap2B in glioma has not yet been fully elucidated.
The present study investigated the protein and mRNA
expression levels of Rap2B in glioma cells and tissues, as well as
the role of Rap2B in regulating the biological activities of glioma
cells via using bi-directional regulation of Rap2B. Additionally,
the correlation between Rap2B expression and disease
characteristics or patient survival was further analyzed in this
study.
Materials and methods
Cell culture
Normal human astrocytes (NHAs) were purchased from a
typical cell culture collection Committee of the Chinese Academy of
Sciences Library and cultured in RPMI-1640 culture medium (Gibco;
Thermo Fisher Scientific, Inc.). Human glioma cell lines U87 (a
glioblastoma of unknown origin) and U251 were purchased from The
Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences and cultured in DMEM/high glucose (HyClone; Cytiva). These
media were supplemented with 10% FBS (HyClone; Cytiva) and 1%
penicillin-streptomycin. All cells were placed in an incubator with
5% CO2 at 37°C.
Cell transfection
Untransfected glioma cells were used as the control
group. Small interfering RNA against Rap2B (si-Rap2B) and
non-targeting negative control (si-NC) were purchased from
Guangzhou RiboBio Co., Ltd. The following sequences were used to
knockdown Rap2B: siRNA, 5′-CGACCAUCGAAGACUUUUATT-3′ (Sense) and
5′-UAAAAGUCUUCGAUGGUCGTT-3′ (antisense). Glioma cells were
transfected with 50 nM si-Rap2B or 50 nM si-NC until cell density
reached ~50% confluence by riboFECT CP reagent (Guangzhou RiboBio
Co., Ltd.) based on the manufacturer's indication. The
overexpression plasmids of pcDNA3.1-Rap2B (oe-Rap2B) and
pcDNA3.1-vector (oe-NC) were designed and synthesized by Shanghai
GeneChem Co., Ltd. When cell confluence reached ~90%, cells were
transfected with 5 µg overexpression plasmids using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) based on the manufacturer's instructions. The
medium was changed to serum-free medium without penicillin and
streptomycin prior to the transfection. Then, cells were subjected
to subsequent experimentation 48 h following transfection.
Cell Counting Kit (CCK)-8 assay
Cell proliferation was measured using CCK-8 (Dojindo
Molecular Technologies, Inc.) according to the manufacturer's
instruction. Briefly, transfected cells were planted into 96-well
plates at a density of 2,500 cells/well, and cultured for 24–72 h.
Then, 10 µl of CCK-8 solution was added to each well and the plates
were further incubated for 2 h at 37°C. The absorbance was measured
at 450 nm using a scanning microplate absorbance reader (Thermo
Fisher Scientific, Inc.).
5-Ethynyl-2′-deoxyuridine (EdU)
assay
After transfection, U87 and U251 cells were seeded
into 96-well plates and cultured in serum-free medium for 24 h
incubation at 37°C. When cell confluence reached 60%, each well was
incubated with 100 µl diluted EdU solution for 2 h at 37°C. The EdU
kit (cat. no. C10310-1) was obtained from Guangzhou RiboBio Co.,
Ltd. Subsequently, the nuclei could be found using an inverted
fluorescence microscope (Nikon Corporation). Then, we counted each
well at 5 different microscope fields under a fluorescence
microscope (magnification, ×200). The quantitative data were
presented as the percentage of EdU-positive nuclei relative to
total number of nuclei counted using ImageJ (version 1.52q;
National Institutes of Health). The experiment was performed in
triplicate.
Cell migration assay
Cell migration was carried out using two-chamber
plates with pore size of 8 µm. After transfection, 2×104
glioma cells were placed into the upper chambers with serum-free
medium, and 500 µl of DMEM (HyClone; Cytiva) containing 20% FBS
(HyClone; Cytiva) was added to the lower chambers. After incubation
at 37°C for 24 h, the cells remaining in the upper chambers were
removed with cotton swabs. The migrated cells in the lower chambers
were fixed with 100% methanol for 20 min at room temperature and
stained with 0.1% crystal violet for 30 min at room temperature. In
total, eight microscopy fields on the lower surface of each chamber
were randomly chosen, and images were captured using an inverted
fluorescence microscope (Nikon Corporation).
Enzyme-linked immunosorbent assay
(ELISA)
The supernatant of transfected U87 and U251 cells
were collected from each group after 24 h. The Human Matrix
MetalloProteinase (MMP)2 ELISA (cat. no. ab100606; Abcam) and MMP9
ELISA kits (cat. no. ab100610; Abcam) were used to detect the
concentrations of MMP2 and MMP9 based on the manufacturer's
guidelines. The absorbance at 450 nm wavelength was recorded using
the microplate reader (Thermo Fisher Scientific, Inc.).
Reverse transcription-quantitative
(RT-q)PCR analysis
Total RNA was extracted from transfected U87 and
U251 cells using TRIzol® (Invitrogen, Thermo Fisher
Scientific, Inc.). cDNA was synthesized using the Reverse
Transcription kit (Takara Bio, Inc.) according to the
manufacturer's protocol. qPCR was performed using SYBR®
Premix Ex Taq™ II kit (Takara Bio, Inc.) on Light Cycler 480
instrument (Roche Diagnostics). The following thermocycling
conditions were used: Initial denaturation at 95°C for 30 sec, 45
cycles of 95°C for 5 sec, 60°C for 30 sec, melting curve at 95°C
for 5 sec, 60°C for 60 sec, 95°C for 5 sec, and cooling at 50°C for
30 sec. The primers were as follows: Rap2B, Forward:
5′-TTACCGCAAGGAGATTGAG-3′, reverse: 5′-GGCTGTAGACCAGGATGAAG-3′;
GAPDH, forward: 5′-GAAGGTGAAGGTCGGAGT-3′, reverse:
5′-GAAGATGGTGATGGGATTTC-3′. The 2−ΔΔCq method was used
to calculate gene expression (23),
and GAPDH was selected as the internal reference.
Western blotting
Total proteins from cultured glioma cells were
extracted using RIPA lysis buffer (Beyotime Institute of
Biotechnology) and their concentrations were determined using a
bicinchoninic acidprotein assay reagent kit (cat. no. P0010S;
Beyotime Institute of Biotechnology). A total of 30 µg proteins
were loaded per lane and separated via 10% SDS-PAGE, then
transferred to a PVDF membrane (Bio-Rad Laboratories, Inc.). After
blocking with 5% fat-free milk for 2 h at room temperature, the
membranes were incubated with primary antibodies overnight at 4°C.
After washing with TBS-Tween-20 (0.5% Tween), the membranes were
incubated with secondary antibody for 1 h at 37°C. Image Lab
software version 3.0 (Bio-Rad Laboratories, Inc.) was used to
detect the protein bands, and proteins were visualized using
enhanced chemiluminescent substrate (Thermo Fisher Scientific,
Inc.). The antibodies used were: Rap2B (cat. no. ab101369;
1:1,000), MMP-2 (cat. no. ab92536; 1:1,000), MMP-9 (cat. no.
ab76003; 1:1,000), ERK (cat. no. ab184699; 1:10,000) and
phosphorylated (p)-ERK (cat. no. ab76299; 1:5,000) (all Abcam).
Secondary antibody (cat. no. SA00001-2; 1:2,000) and GAPDH (cat.
no. 10494-1-AP; 1:10,000) were purchased from ProteinTech Group,
Inc.
Acquisition of clinical parameters and
RNA-sequencing data
The clinical and Rap2B expression data of 156 cases
of patients with glioblastoma multiforme (GBM) and 511 cases of
low-grade glioma (LGG) were acquired from the dataset (Project ID:
TCGA-GBM and TCGA-LGG) in The Cancer Genome Atlas (TCGA) database
(https://portal.gdc.cancer.gov/). Then,
the normalized mRNA count (‘count’), which represented Rap2B gene
expression, was used. Exclusion criteria were those samples with
histological diagnosis excluding GBM or LGG and incomplete
RNA-sequencing data. Finally, Rap2B expression files containing GBM
(152 GBM tissues; five matched normal tissues) and 509 LGG (248
grade 2; 261 grade 3) were downloaded from TCGA project. The
grading system was based on the World Health Organization
classification (24).
Statistical analysis
All experiments were independently repeated at least
three times. Data were analyzed with SPSS 19.0 software (IBM Corp.)
and GraphPad Prism 6.01 software (GraphPad Software, Inc.) and
presented as mean ± standard deviations. Comparisons between two
groups were executed using Student's t-test. Comparisons among ≥3
groups were performed using one-way ANOVA followed by Tukey's test.
χ2 or Fisher's exact tests were performed to assess the
relationship between Rap2B expression and clinicopathological
characteristics. Then, the survival curves of Rap2B were plotted
using Kaplan-Meier analysis based on the log-rank test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Rap2B upregulation in human
glioma
Compared with NHA, the results of western blotting
showed that the protein level of Rap2B in U87 and U251 cells was
upregulated (Fig. 1A; P<0.001).
To confirm these findings, Rap2B expression was further analyzed
using TCGA database. The results revealed that Rap2B expression was
significantly upregulated in 152 GBM tissues compared with that in
five matched normal tissues (Fig.
1B; P=0.0372). In 509 patients with LGG, Rap2B expression in
patients with tumor grade 3 was significantly higher compared with
patients with tumor grade 2 (Fig.
1C; P<0.0001). Among histological types of LGG, Rap2B from
astrocytoma was more highly expressed compared with
oligodendroglioma based on TCGA-LGG dataset (Fig. 1D; P=0.001). The results demonstrated
that Rap2B was upregulated and associated with tumor grade and
histological type in human glioma.
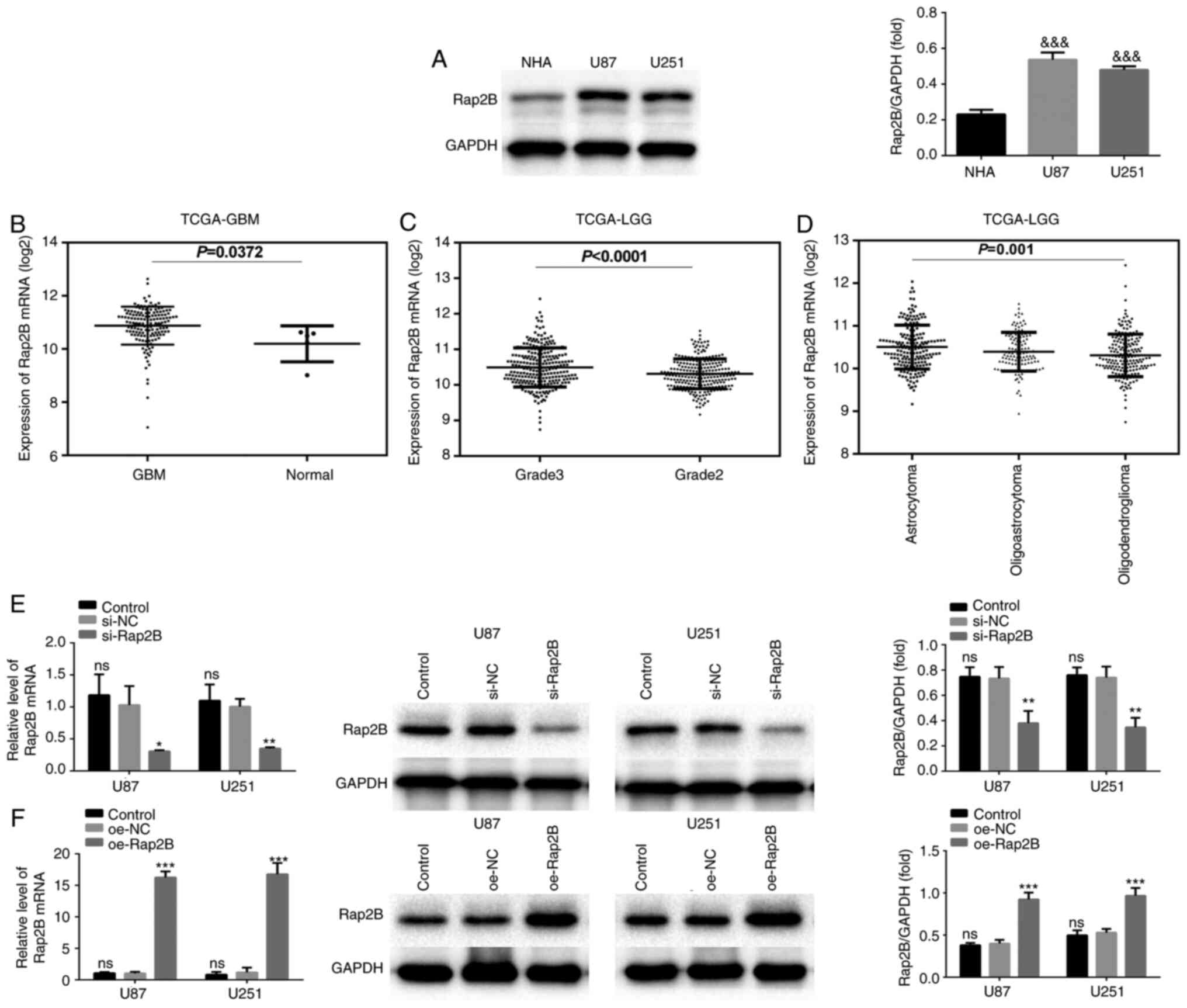 | Figure 1.Rap2B expression in glioma. (A)
Expression of Rap2B analyzed using western blot analysis in NHA,
U87 and U251 cells. (B) Rap2B expression was elevated in 152 GBM
samples compared with that in five adjacent normal samples in the
TCGA-GBM dataset. (C) In the TCGA-LGG dataset, Rap2B expression in
patients with tumor grade 3 (n=261) was notably higher compared
with patients with tumor grade 2 (n=248). (D) Rap2B in astrocytoma
(n=192) was upregulated relative to oligodendroglioma (n=190) based
on TCGA-LGG dataset. (E) Detection of Rap2B-silencing in U87 and
U251 cells. (F) Detection of Rap2B overexpression in U87 and U251
cells. &&&P<0.001 vs. NHA. *P<0.05,
**P<0.01 and ***P<0.001 vs. the corresponding NC group. TCGA,
The Cancer Genome Atlas; GBM, glioblastoma multiforme; LGG,
low-grade glioma; NHA, normal human astrocyte; NC, negative
control; ns, no statistical significance; si, short interfering;
oe, overexpressed. |
Then, Rap2B was simultaneously silenced or
overexpressed in U87 and U251 cells. No significant difference was
observed in the levels of Rap2B mRNA and protein between the
control group and the respective NC group (Fig. 1E and F). Relative to the respective
NC group, mRNA and protein expression of Rap2B in both U87 and U251
cells was decreased in the silenced Rap2B group (Fig. 1E; P<0.05) and increased in the
overexpressed Rap2B group (Fig. 1F;
P<0.001).
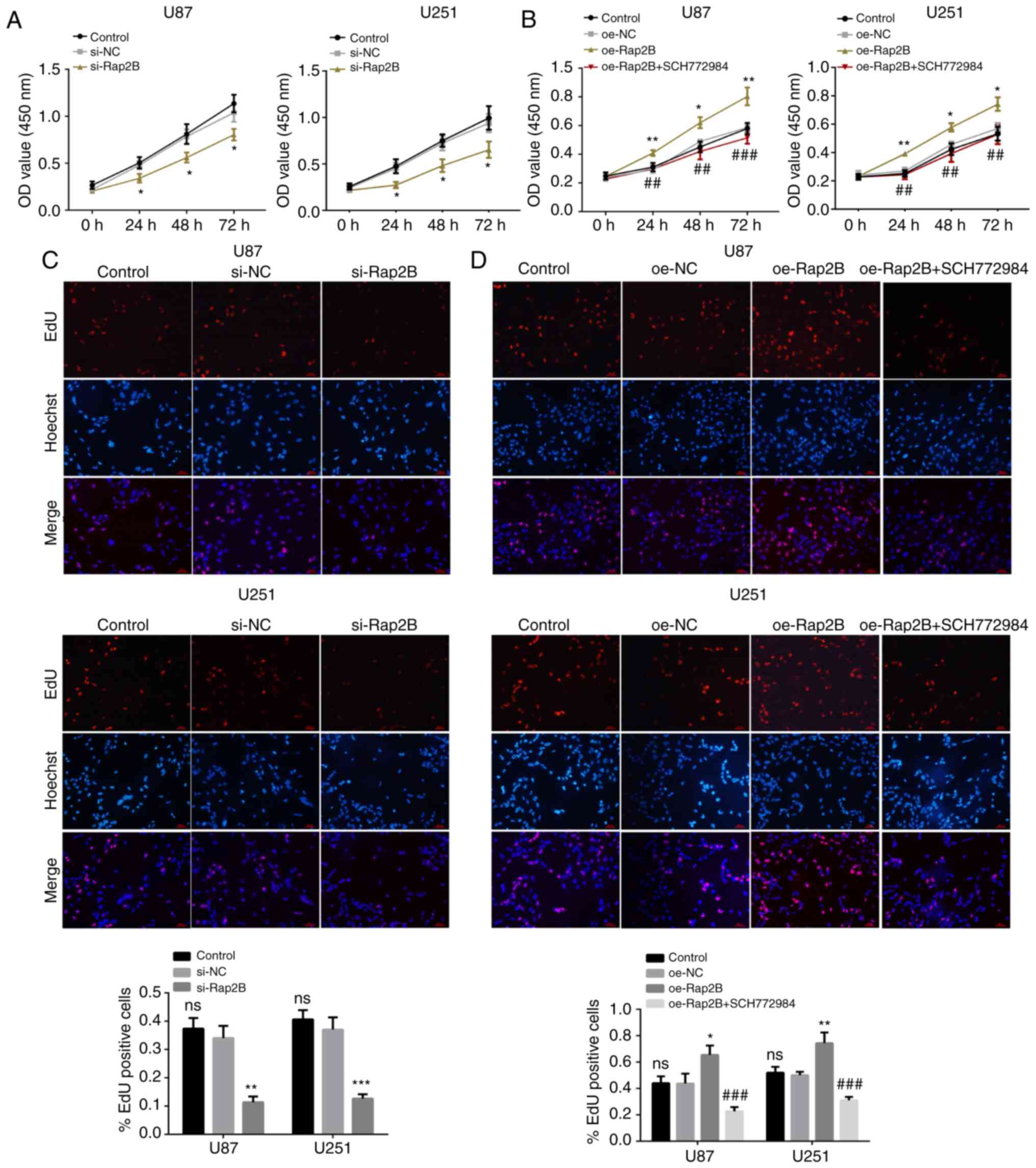
Silenced or overexpressed Rap2B
influences proliferation of glioma cells
To investigate whether Rap2B affects the
proliferation of glioma cells, a CCK-8 assay was performed. It was
observed that there was no significant difference in cell
proliferation between the respective NC and control groups. The
CCK-8 assay revealed that proliferation rates of U87 and U251 cells
were significantly reduced in the si-Rap2B group (Fig. 2A; P<0.05), but significantly
enhanced in the Rap2B-overexpression group compared with the
corresponding NC group (Fig. 2B;
P<0.05). To further verify the effect of Rap2B on glioma cell
proliferation, an EdU assay was conducted. Similar results were
obtained, showing a significant decrease in the number of
EdU-positive glioma cells with Rap2B-knockdown (Fig. 2C; P<0.05), while the number of
EdU-positive glioma cells transfected with Rap2B overexpression
plasmids was significantly elevated relative to the corresponding
NC group (Fig. 2D; P<0.05). The
inhibitor SCH772984 weakened this promotive effect of proliferation
elicited by overexpressed Rap2B in both U87 and U251 cells
(P<0.05; Fig. 2B and D),
suggesting the involvement of ERK pathway in regulating
Rap2B-mediated proliferation. The aforementioned results
demonstrated that the alterations of Rap2B were closely associated
with the proliferation of glioma cells.
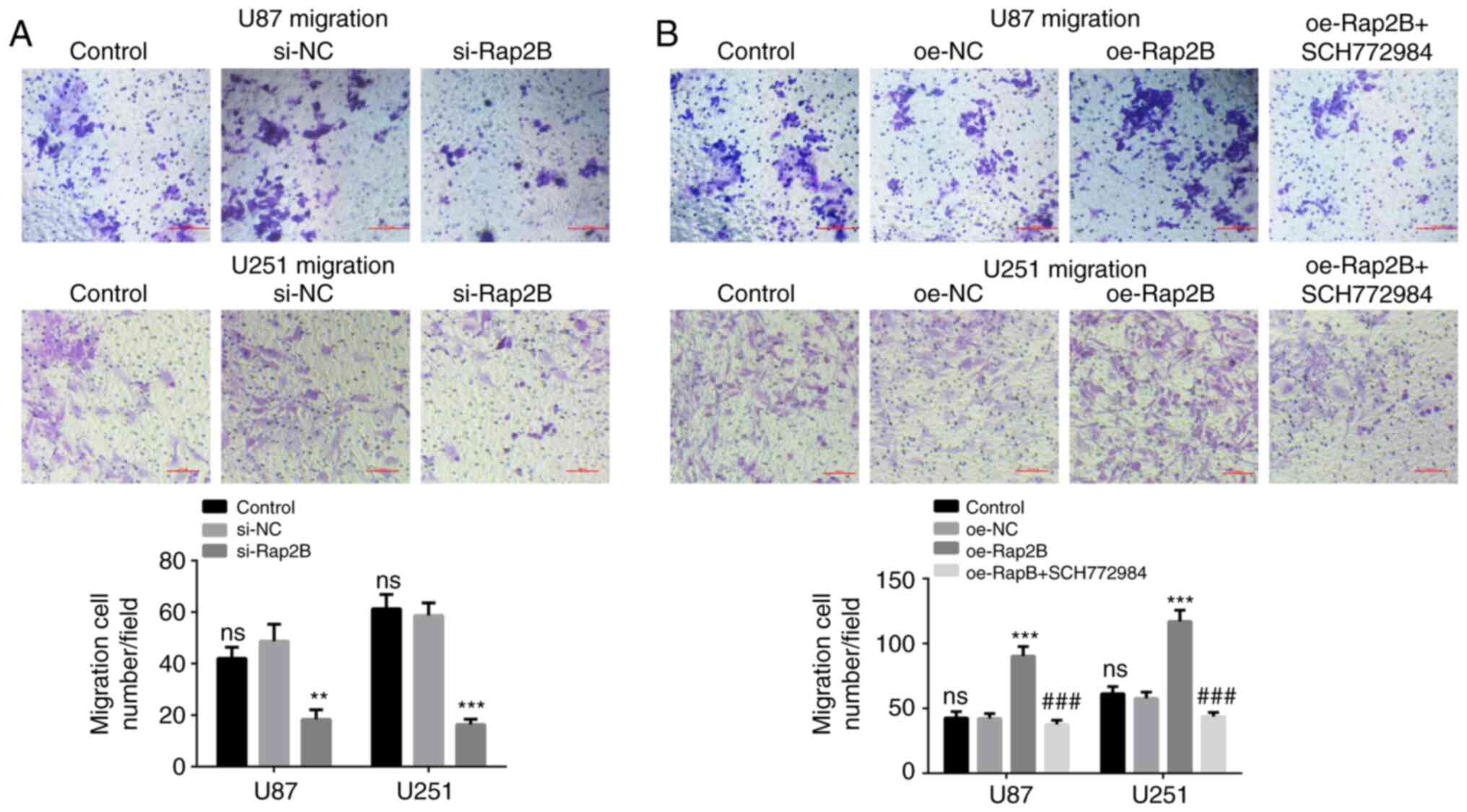
Silenced or overexpressed Rap2B
influences migration of glioma cells
The effect of Rap2B on the migration ability of
glioma cells was evaluated using a cell migration assay. Compared
with the NC group, Rap2B-knockdown significantly reduced the
migration ability of U87 and U251 cells (Fig. 3A; P<0.05), whereas Rap2B
overexpression led to a significantly enhanced migration ability
(Fig. 3B; P<0.001). No
significant difference could be observed in the migration of U87
and U251 cells between the control group and relative NC group.
These results indicated that Rap2B could promote the migration
ability of glioma cells.
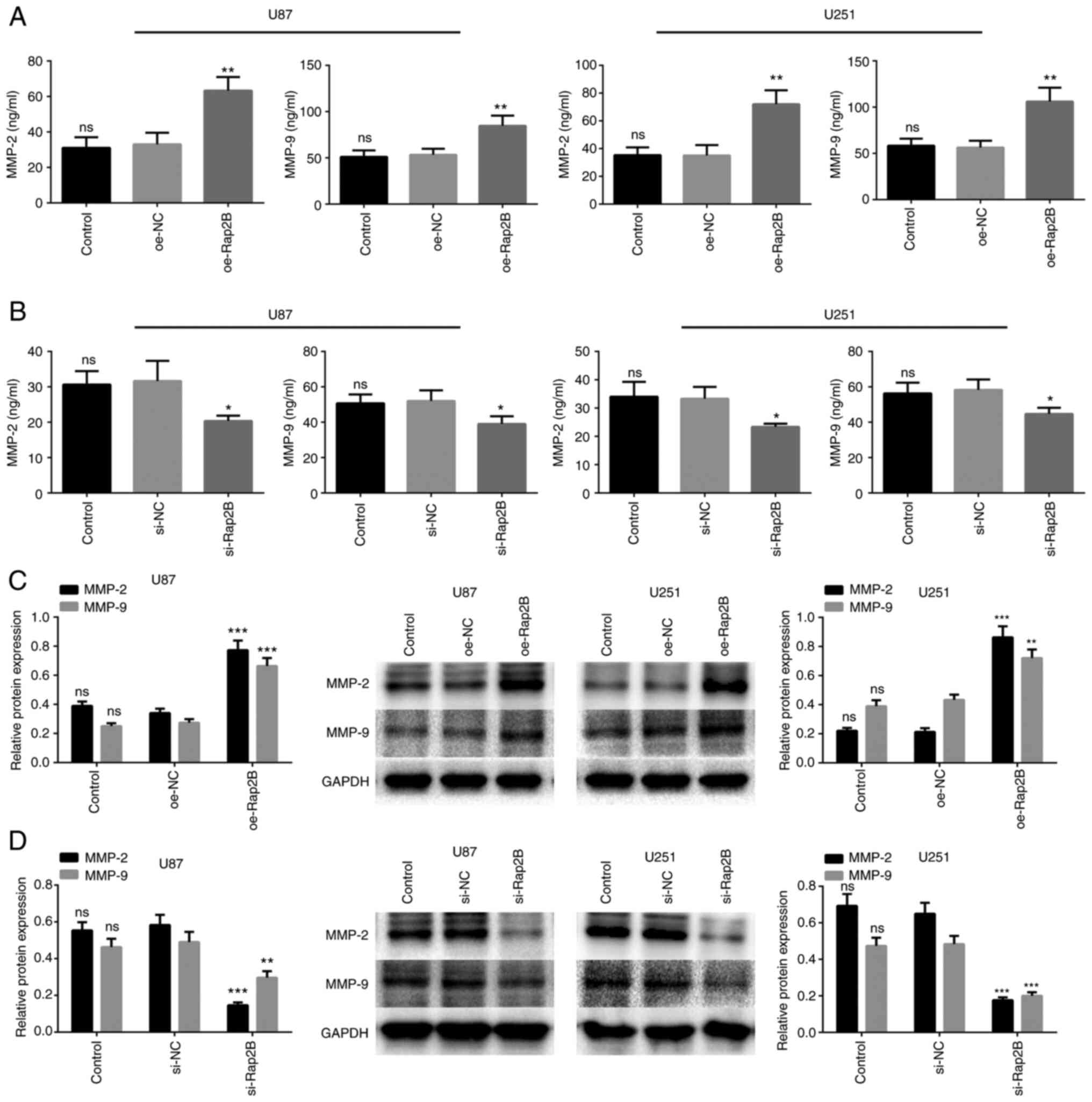
Rap2B enhances MMP2 and MMP9
expression in glioma cells
To investigate whether Rap2B could increase MMP
expression, an ELISA assay and western blotting were performed to
examine the expression of MMP2 and MMP9. As shown in Fig. 4A, Rap2B-overexpressing U87 and U251
cells presented significant increases in protein levels of MMP2 and
MMP9 in the supernatant of glioma cells (P<0.01). Rap2B
expression in U87 and U251 cells was silenced by Rap2B siRNA to
determine the effect on protein expression of MMP2 and MMP9. The
results suggested that Rap2B-knockdown attenuated the secretion of
MMP2 and MMP9 compared with the NC group in U87 and U251 cells (all
P<0.05; Fig. 4B). Moreover, this
was corroborated by western blot analysis, showing the increased
levels of MMP2 and MMP9 in the overexpressed Rap2B group and
decreased MMP2 and MMP9 expression in Rap2B-silenced U87 and U251
cells compared with the NC group (all P<0.01 or P<0.001;
Fig. 4C and D). Collectively, these
results demonstrated that Rap2B facilitated the expression of MMP2
and MMP9.
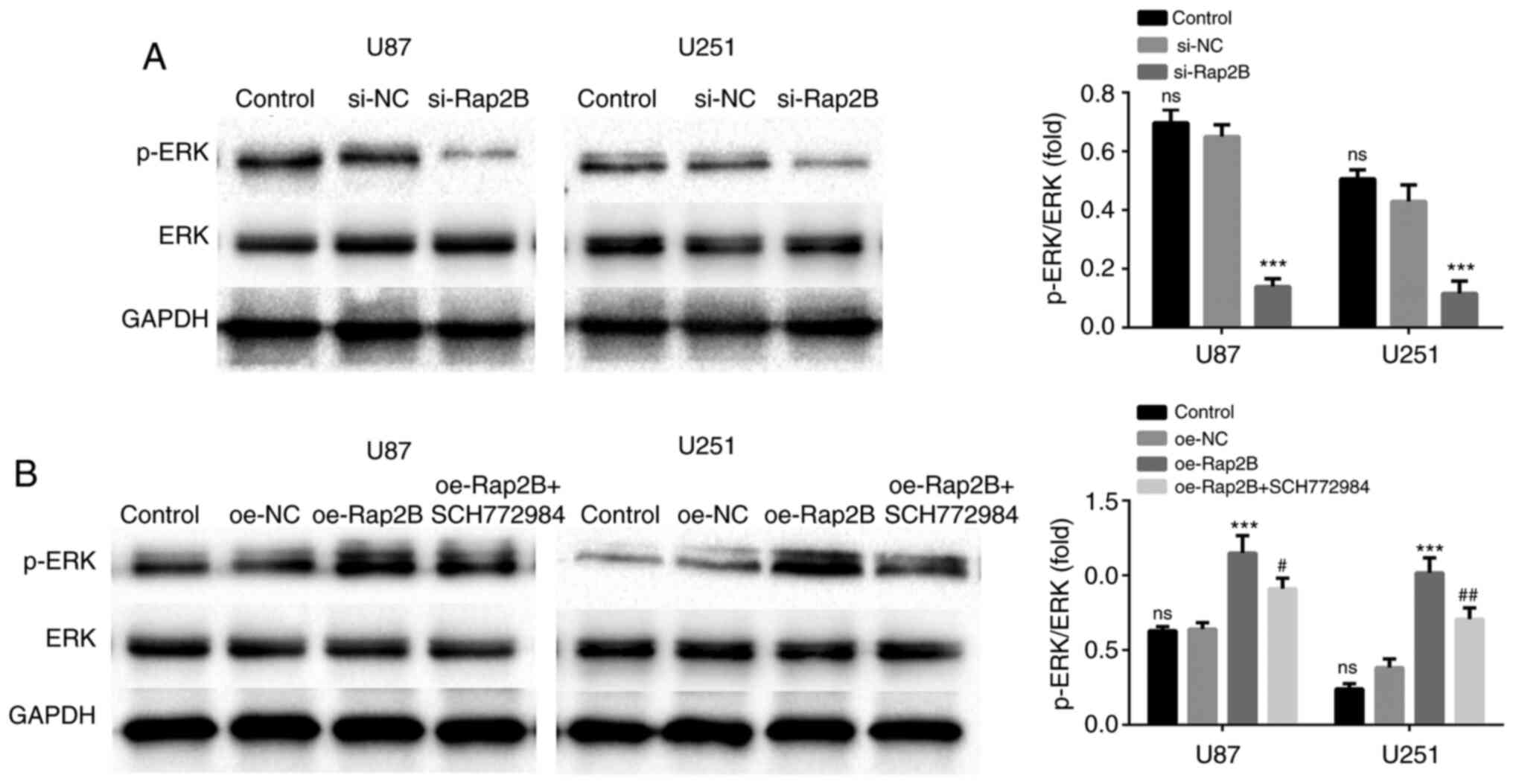
Rap2B is associated with the
activation of the ERK pathway
Rap2B could potentially activate multiple pathways,
such as MAPK/ERK, PI3K/AKT and NF-κB (22,25,26). To
analyze whether Rap2B could lead to ERK activation, glioma cells
were transfected with Rap2B-overexpression plasmids. Western blot
analysis revealed that the phosphorylation of ERK was significantly
enhanced in Rap2B-overexpressing U87 and U251 cells compared with
that in the NC group (Fig. 5B;
P<0.001). However, SCH772984 (an ERK pathway inhibitor) reversed
the promotion in the phosphorylation of ERK mediated by
Rap2B-overexpression in U87 and U251 cells (P<0.05). In
contrast, the level of ERK phosphorylation was attenuated in
Rap2B-silenced U87 and U251 cells relative to the NC group
(Fig. 5A; P<0.001). These
findings demonstrated that Rap2B significantly promoted ERK
activation in glioma cells.
Effects of ERK signaling on
Rap2B-induced cell proliferation, migration and expression of MMP2
and MMP9
To further confirm the involvement of ERK pathway in
Rap2B-stimulated secretion of MMP2 and MMP9, glioma cells were
treated with SCH772984. As presented in Figs. 2B-D and 3B, it was observed that SCH772984
significantly weakened Rap2B-induced promotion of proliferation and
migration of glioma cells (P<0.05), indicating that ERK
signaling was required for proliferation and migration induced by
Rap2B. Moreover, the results of the ELISA assays and western
blotting showed that MMP2 and MMP9 expression was significantly
reduced following co-treatment with Rap2B and SCH772984 compared
with that in the overexpressed Rap2B group (all P<0.05,
P<0.01 or P<0.001; Fig. 6A-D).
Hence, inhibition of ERK pathway using SCH772984 treatment could
restrain Rap2B-mediated production of MMP2 and MMP9 (Fig. 6A-D), demonstrating the involvement of
ERK signaling in modulating Rap2B-stimulated MMP2 and MMP9
production.
Association of Rap2B expression with
clinical parameters in 152 GBM and 509 LGG samples
As Rap2B expression promoted the proliferation and
migration of glioma cells, the association between Rap2B expression
and clinical features in GBM and LGG was further analyzed using
TCGA database. As presented in Table
I, the relationship between Rap2B expression and clinical
characteristics of patients with GBM were detailed. It was revealed
that Rap2B expression was not significantly associated with sex,
age and tumor recurrence or progression status in GBM. Kaplan-Meier
survival analysis indicated that patients with GBM with high
expression of Rap2B showed a shorter survival time compared with
those in the low expression group, but there was no significant
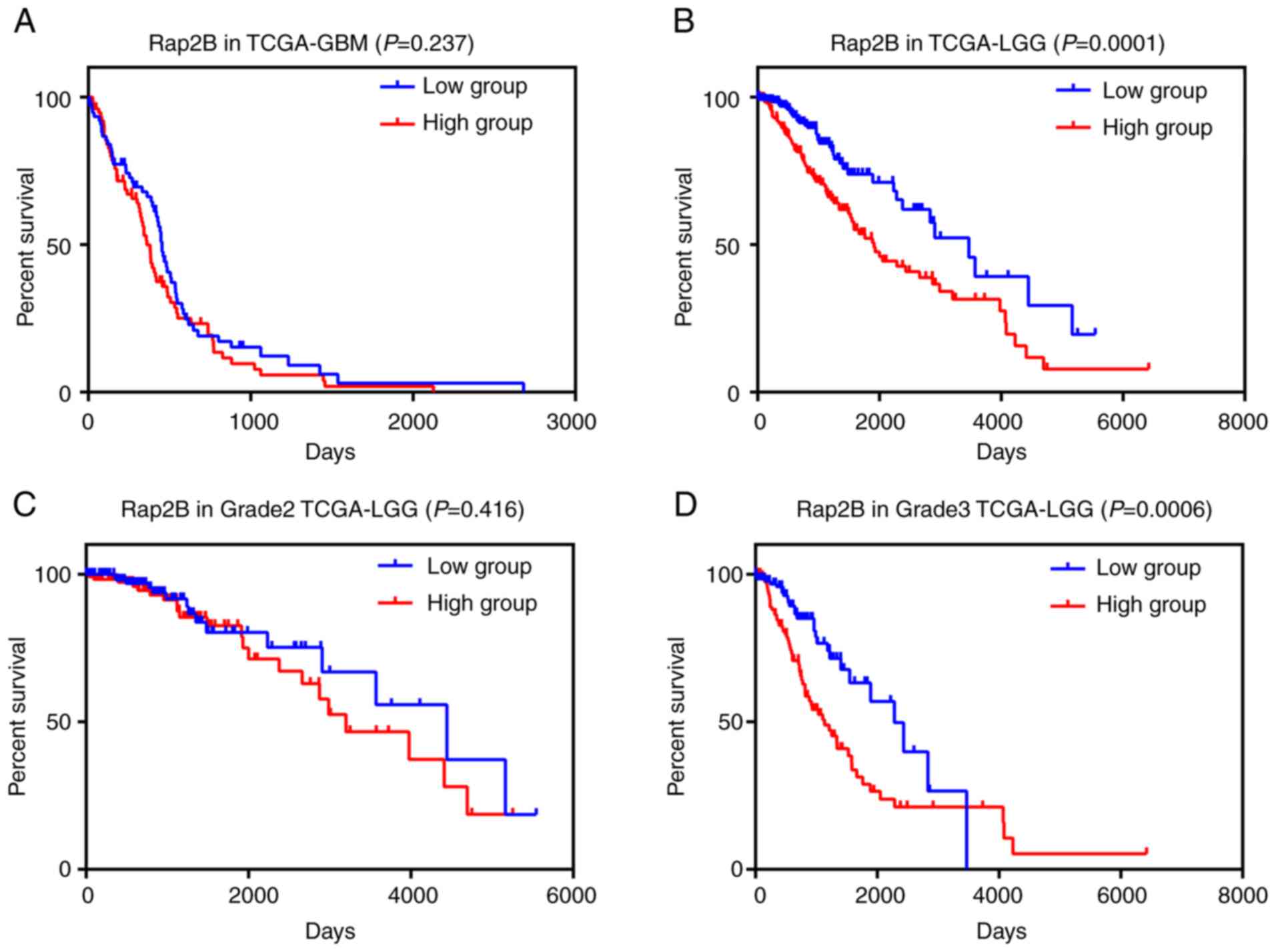
difference between two groups (Fig.
7A). In LGG samples, as presented in Table II, the results revealed that Rap2B
expression was significantly associated with tumor grade and
histological type (P<0.05), while no significant association was
observed between Rap2B expression and sex or age in this
population. Further survival analysis showed that patients with LGG
with higher expression of Rap2B had significantly unfavorable
survival compared with those in the lower expression group
(Fig. 7B; P=0.0001). Moreover, high
expression of Rap2B was found to be associated with poor prognosis
of patients with tumor grade 3 (Fig.
7D; P=0.0006), while there was no significant difference
between high and low expression groups of patients with grade 2 LGG
(Fig. 7C). These results indicated
that Rap2B expression was closely associated with certain clinical
characteristics and higher Rap2B expression was associated with
poorer survival of patients with LGG.
 | Table I.Association of Rap2B expression with
clinical features in 152 patients with glioblastoma multiforme. |
Table I.
Association of Rap2B expression with
clinical features in 152 patients with glioblastoma multiforme.
|
|
| Rap2B
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Value, n | High group | Low group | P-value |
|---|
| Age, years |
|
>60 | 76 | 37 | 39 | 0.746 |
|
≤60 | 76 | 39 | 37 |
|
| Sex |
|
Female | 54 | 26 | 28 | 0.735 |
|
Male | 98 | 50 | 48 |
|
| Recurrence or
progression |
|
Yes | 84 | 44 | 40 | 0.514 |
| No | 68 | 32 | 36 |
|
| Table II.Expression of Rap2B associated with
clinical features in 509 patients with low-grade glioma. |
Table II.
Expression of Rap2B associated with
clinical features in 509 patients with low-grade glioma.
|
|
| Rap2B
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Value, n | High group | Low group | P-value |
|---|
| Age, years |
|
>43 | 222 | 107 | 115 | 0.499 |
|
≤43 | 287 | 147 | 140 |
|
| Sex |
|
Female | 228 | 112 | 116 | 0.752 |
|
Male | 281 | 142 | 139 |
|
| Histological
type |
|
Astrocytoma | 192 | 114 | 78 | 0.001 |
|
Oligoastrocytoma | 127 | 63 | 64 |
|
|
Oligodendroglioma | 190 | 77 | 113 |
|
| Grade |
| G2 | 248 | 109 | 139 | 0.009 |
| G3 | 261 | 145 | 116 |
|
Discussion
Rap2B, a member of the Ras family of the guanosine
triphosphate-binding proteins, has attracted considerable attention
since its identification in the early 1990s (27). A previous study reported that
mutations or overexpression of Ras genes are implicated in
tumorigenesis and predicted poor prognostic survival in numerous
types of tumors, such as thyroid, lung and colorectal cancer
(28). Rap2B is observed to be
predominantly upregulated in a variety of tumors and acts as an
oncogenic activator (29,30). In the present study, Rap2B was highly
expressed in both glioma cells and tissues. Additionally, it was
revealed that Rap2B could facilitate the proliferation and
migration of glioma cells. These findings suggested that Rap2B may
have a pivotal role in the development and progression of human
glioma.
Rap2B is reported to be involved in regulating
cellular processes including cytoskeletal organization and cell
proliferation (31). Increasing
focus on the Ras signaling pathway reveals critical effects of
Rap2B on tumorigenesis. Previous research reported that Rap2B
exhibits oncogenic status and functions as a tumor promoter in
human tumors (20,32). Evidence suggests that Rap2B promotes
cell proliferation, migration and invasion in cervical cancer
(33). Another study revealed that
increased Rap2B is associated with cell migration and invasion of
human suprarenal epithelioma (34).
Consistent with the results of aforementioned studies, the present
study demonstrated that Rap2B enhanced the proliferation and
migration of U87 and U251 cells, whereas silencing Rap2B restrained
this effect, further indicating the biological function of
Rap2B.
As a member of the Ras superfamily of proteins,
Rap2B facilitates the activation of multiple pathways such as
MAPK/ERK, PI3K/AKT and NF-κB, which are implicated in the
occurrence and progression of various tumors, including breast
cancer, hepatocellular carcinoma and glioma (18,26). ERK
is a key factor in cancer, and it is reported that the
Ras/Raf/MAPK/ERK pathway modulates a series of cellular biological
processes associated with tumorigenesis(35). Rap2B enhances cell
proliferation through the ERK pathway, and ERK activation has a
critical role in the development of cervical cancer, such as cell
proliferation, migration and invasion (33). A previous study reported that Rap2B
contributes to cell proliferation, migration and invasion in breast
cancer via the calcium-induced ERK pathway (25). A recent study showed that Rap2B
silencing inhibits the development of hepatocellular carcinoma via
inhibition of the PTEN/PI3K/Akt and ERK pathways (26). Concordantly, the present study
indicated that Rap2B activated the ERK signaling pathway, which
enhanced the proliferation and migration of glioma cells.
Although accumulating evidence has revealed the
crucial role of Rap2B in the development and progression of
multiple tumors (18,25,26), the
potential molecular mechanism by which Rap2B promotes the
proliferation and migration of glioma cells remains unknown. Tumor
metastasis is induced partially by MMPs (36–38).
MMP2 and MMP9 drive cell motility and tumor growth by digesting the
ECM, basal lamina and adhesion proteins (10,39). The
present study reported that Rap2B enhanced MMP2 and MMP9
expression, while the inhibition of the ERK signaling pathway
reversed Rap2B-mediated MMP2 and MMP9 expression, demonstrating
that ERK signaling is required for Rap2B-induced production of MMP2
and MMP9. Since MMP2 and MMP9 are crucial to tumor growth and
metastasis (40), Rap2B may enhance
the proliferation and migration of glioma cells through increasing
MMP2 and MMP9 expression.
To further investigate the association between Rap2B
expression and certain clinical parameters, TCGA, a publicly
available database, was used. Kaplan-Meier analysis showed that
high expression of Rap2B was associated with overall survival time
of patients with LGG, indicating a risk factor in LGG. In patients
with GBM, the same trend was observed, but no significant
difference was found between the high and low Rap2B expression
group. It is worth noting that patients with GBM are generally
treated with preoperative radiotherapy and concomitant temozolomide
(41). It was postulated that the
discrepancy in these statistical results may stem from these
preoperative therapies in patients with GBM, thereby affecting
Rap2B transcription. These findings suggested that higher Rap2B
expression predicts poorer prognosis in patients with glioma.
Therefore, Rap2B may be used as a novel prognostic biomarker and
therapeutic target for glioma.
Of note, there were limitations in the present
study. Due to the small number of adjacent normal brain tissues in
TCGA-GBM dataset, it may be not adequate to accurately compare
Rap2B expression between GBM tissues and matched normal tissues. So
immunohistochemistry data from patients with glioma should be
obtained. Furthermore, there was a lack of animal experiments in
the study, but this will be the focus of the authors' future
research. Xenograft experiments should be performed to confirm the
present findings in vivo. Further investigation on the
effect of Rap2B on glioma growth and the detailed mechanisms in
glioma is the direction of our future study.
In summary, the present study revealed that Rap2B
enhanced the proliferation and migration of human glioma cells via
activation of ERK pathway. Additionally, Rap2B expression was
upregulated in glioma and is associated with overall survival time
of patients with glioma, indicating that it might act as a
prognostic biomarker and therapeutic target for glioma.
Acknowledgements
Not applicable.
Funding
This research was supported by The Natural Science
Foundation of Liaoning Province (grant no. 2019-ZD-0772) and The
National Natural Science Foundation of China (grant nos. 81471809
and 81971639).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request. Additional datasets generated and/or analyzed during the
current study are available in The Cancer Genome Atlas repository,
(https://portal.gdc.cancer.gov/).
Authors' contributions
GHS and ZZ designed the experiments and analyzed
data. GHS contributed to performing the experiments, collecting
data, and data analysis and interpretation, as well as drafting the
manuscript. Both authors read and approved the final manuscript.
GHS and ZZ confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Wen PY and Reardon DA: Neuro-oncology in
2015: Progress in glioma diagnosis, classification and treatment.
Nat Rev Neurol. 12:69–70. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carlsson SK, Brothers SP and Wahlestedt C:
Emerging treatment strategies for glioblastoma multiforme. EMBO Mol
Med. 6:1359–1370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zeng Z, Leng T, Feng X, Sun H, Inoue K,
Zhu L and Xiong ZG: Silencing TRPM7 in mouse cortical astrocytes
impairs cell proliferation and migration via ERK and JNK signaling
pathways. PLoS One. 10:e01199122015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heigener DF, Gandara DR and Reck M:
Targeting of MEK in lung cancer therapeutics. Lancet Respir Med.
3:319–327. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
5. Liu Z, Dai H, Jia G, Li Y, Liu X and
Ren W: Insufficient radiofrequency ablation promotes human hepatoma
SMMC7721 cell proliferation by stimulating vascular endothelial
growth factor overexpression. Oncol Lett. 9:1893–1896. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dong F, Tian H, Yan S, Li L, Dong X, Wang
F, Li J, Li C, Cao Z, Liu X, et al: Dihydroartemisinin inhibits
endothelial cell proliferation through the suppression of the ERK
signaling pathway. Int J Mol Med. 35:1381–1387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Figueira RC, Gomes LR, Neto JS, Silva FC,
Silva ID and Sogayar MC: Correlation between MMPs and their
inhibitors in breast cancer tumor tissue specimens and in cell
lines with different metastatic potential. BMC Cancer. 9:202009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bachmeier BE, Nerlich AG, Lichtinghagen R
and Sommerhoff CP: Matrix metalloproteinases (MMPs) in breast
cancer cell lines of different tumorigenicity. Anticancer Res.
21A:3821–3828. 2001.
|
|
9
|
Hotary KB, Yana I, Sabeh F, Li XY,
Holmbeck K, Birkedal-Hansen H, Allen ED, Hiraoka N and Weiss SJ:
Matrix metalloproteinases (MMPs) regulate fibrin-invasive activity
via MT1-MMP-dependent and -independent processes. J Exp Med.
195:295–308. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Forsyth PA, Wong H, Laing TD, Rewcastle
NB, Morris DG, Muzik H, Leco KJ, Johnston RN, Brasher PM,
Sutherland G, et al: Gelatinase-A (MMP-2), gelatinase-B (MMP-9) and
membrane type matrix metalloproteinase-1 (MT1-MMP) are involved in
different aspects of the pathophysiology of malignant gliomas. Br J
Cancer. 79:1828–1835. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo XX, An S, Yang Y, Liu Y, Hao Q and Xu
TR: Rap-Interacting Proteins are Key Players in the Rap Symphony
Orchestra. Cell Physiol Biochem. 39:137–156. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yi L, Zhong X, Chen Z, Wang Q, Yan Y, Wang
J and Deng X: MicroRNA-147b Promotes Proliferation and Invasion of
Human Colorectal Cancer by Targeting RAS Oncogene Family (RAP2B).
Pathobiology. 86:173–181. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Duan HB and Yang YS: Knockdown of
Rap2B Inhibits the Proliferation and Invasion in Hepatocellular
Carcinoma Cells. Oncol Res. 25:19–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng YG, Zhang ZQ, Chen YB and Huang JA:
Rap2b promotes proliferation, migration, and invasion of lung
cancer cells. J Recept Signal Transduct Res. 36:459–464. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Raaijmakers JH and Bos JL: Specificity in
Ras and Rap signaling. J Biol Chem. 284:10995–10999. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Itoh M, Nelson CM, Myers CA and Bissell
MJ: Rap1 integrates tissue polarity, lumen formation, and
tumorigenic potential in human breast epithelial cells. Cancer Res.
67:4759–4766. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kooistra MR, Dubé N and Bos JL: Rap1: A
key regulator in cell-cell junction formation. J Cell Sci.
120:17–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Di J, Gao K, Qu D, Yang J and Zheng J:
Rap2B promotes angiogenesis via PI3K/AKT/VEGF signaling pathway in
human renal cell carcinoma. Tumour Biol. 39:10104283177016532017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Di J, Cao H, Tang J, Lu Z, Gao K, Zhu Z
and Zheng J: Rap2B promotes cell proliferation, migration and
invasion in prostate cancer. Med Oncol. 33:582016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, He Y, Lee KH, Dubois W, Li Z, Wu
X, Kovalchuk A, Zhang W and Huang J: Rap2b, a novel p53 target,
regulates p53-mediated pro-survival function. Cell Cycle.
12:1279–1291. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang M, Zhuang Q and Cui L: miR-194
inhibits cell proliferation and invasion via repression of RAP2B in
bladder cancer. Biomed Pharmacother. 80:268–275. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Miao F, Cui C, Zuo D, Zhang H, Mei P, Chen
H, Wei S, Yang F, Zheng J, Bai J, et al: Rap2B promotes cell
adhesion, proliferation, migration and invasion of human glioma. J
Neurooncol. 143:221–229. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Di J, Huang H, Qu D, Tang J, Cao W, Lu Z,
Cheng Q, Yang J, Bai J, Zhang Y, et al: Rap2B promotes
proliferation, migration, and invasion of human breast cancer
through calcium-related ERK1/2 signaling pathway. Sci Rep.
5:123632015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu L, Sun Y, Zhang S and Wang L: Rap2B
knockdown suppresses malignant progression of hepatocellular
carcinoma by inactivating the PTEN/PI3K/Akt and ERK1/2 pathways.
Mol Cell Biochem. 466:55–63. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Farrell FX, Ohmstede CA, Reep BR and
Lapetina EG: cDNA sequence of a new ras-related gene (rap2b)
isolated from human platelets with sequence homology to rap2.
Nucleic Acids Res. 18:42811990. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mascaux C, Iannino N, Martin B, Paesmans
M, Berghmans T, Dusart M, Haller A, Lothaire P, Meert AP, Noel S,
et al: The role of RAS oncogene in survival of patients with lung
cancer: A systematic review of the literature with meta-analysis.
Br J Cancer. 92:131–139. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lv GY, Miao J and Zhang XL: Long Noncoding
RNA XIST Promotes Osteosarcoma Progression by Targeting Ras-Related
Protein RAP2B via miR-320b. Oncol Res. 26:837–846. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Q, Duan L, Xu H, Yuan J, Wu W, Wu Y,
Gao Y and Cheng S: Effects of Rap2b gene on foci formation and
wound-healing of NIH3T3 cells. Wei Sheng Yan Jiu. 39:403–406.
2010.(In Chinese). PubMed/NCBI
|
|
31
|
Zhu Z, Di J, Lu Z, Gao K and Zheng J:
Rap2B GTPase: Structure, functions, and regulation. Tumour Biol.
37:7085–7093. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie X, Liu H, Wang M, Ding F, Xiao H, Hu
F, Hu R and Mei J: miR-342-3p targets RAP2B to suppress
proliferation and invasion of non-small cell lung cancer cells.
Tumour Biol. 36:5031–5038. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Li S and Huang L: Knockdown of
Rap2B, a Ras Superfamily Protein, Inhibits Proliferation,
Migration, and Invasion in Cervical Cancer Cells via Regulating the
ERK1/2 Signaling Pathway. Oncol Res. 26:123–130. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Di JH, Qu DB, Lu Z, Li LT, Cheng Q, Xin Y,
Zhang LZ, Zhang Y and Zheng JN: Rap2B promotes migration and
invasion of human suprarenal epithelioma. Tumour Biol.
35:9387–9394. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Martinelli E, Morgillo F, Troiani T and
Ciardiello F: Cancer resistance to therapies against the
EGFR-RAS-RAF pathway: The role of MEK. Cancer Treat Rev. 53:61–69.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Markovic DS, Vinnakota K, Chirasani S,
Synowitz M, Raguet H, Stock K, Sliwa M, Lehmann S, Kälin R, van
Rooijen N, et al: Gliomas induce and exploit microglial MT1-MMP
expression for tumor expansion. Proc Natl Acad Sci USA.
106:12530–12535. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mentlein R, Hattermann K and Held-Feindt
J: Lost in disruption: Role of proteases in glioma invasion and
progression. Biochim Biophys Acta. 1825:178–185. 2012.PubMed/NCBI
|
|
38
|
Zheng X, Chopp M, Lu Y, Buller B and Jiang
F: miR-15b and miR-152 reduce glioma cell invasion and angiogenesis
via NRP-2 and MMP-3. Cancer Lett. 329:146–154. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Alaseem A, Alhazzani K, Dondapati P,
Alobid S, Bishayee A and Rathinavelu A: Matrix Metalloproteinases:
A challenging paradigm of cancer management. Semin Cancer Biol.
56:100–115. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Velinov N, Poptodorov G, Gabrovski N and
Gabrovski S: The role of matrixmetalloproteinases in the tumor
growth and metastasis. Khirurgiia (Sofiia). 1:44–49. 2010.(In
Bulgarian).
|
|
41
|
Wang C, Zhao N, Zheng Q, Zhang D and Liu
Y: BHLHE41 promotes U87 and U251 cell proliferation via
ERK/cyclinD1 signaling pathway. Cancer Manag Res. 11:7657–7672.
2019. View Article : Google Scholar : PubMed/NCBI
|