Introduction
Breast cancer is a lethal malignancy, which has
become the second leading cause of mortality in females in the
United States (1,2). A previous study has reported that the
new cases of breast cancer have an increased incidence of ~30%
compared with that noted in other types of cancer in women
(3). Despite recent advancements in
surgery, chemotherapy, radiation and hormone therapy, the 5-year
survival rate of patients with breast cancer is 80–85% (4–6). This is
mainly attributed to the lack of molecular mechanism analysis that
can explain in detail the pathways responsible for breast cancer
formation. Thus, detailed investigation of the pathogenesis of
breast cancer is of considerable significance. It has been
demonstrated that the prevention of metastasis is a vital factor
for the effective reduction of breast cancer (7,8).
Estrogen plays a crucial role, not only in the proliferation and
initiation of breast cancer, but also in migration (9). Thus, it is essential to unravel the
associated molecular mechanisms of breast cancer formation to
improve the treatment and prognosis of breast carcinomas.
17β-estradiol (E2) is one of the three main
self-producing estrogens and is considered an important hormone in
women (10). E2 has a crucial
influence on the growth and function of the female reproductive
system, and of the mammary gland under physiological conditions
(11–13). E2 is involved in the induction of
malignant transformation of several types of cancer cells via
regulation of the associated biological processes, such as
proliferation, invasion and migration (14,15).
Most of the biological effects of estrogens are mediated via
binding and activation of the classic estrogen receptors (ERs)
(16). The G protein -coupled
estrogen receptor (GPER) is also denoted as G protein receptor 30
(GPR30) and has been demonstrated to bind to E2 (17). GPER is responsible for interacting
with multiple genomic signaling pathways in different types of
cells, including breast, endometrial, ovarian and thyroid cancers
(18–24). Overexpression of GPER in breast
cancer is positively associated with a metastatic phenotype
(25). However, the molecular
mechanism of proliferation, invasion and migration of breast cancer
resulting from E2 binding to GPER remains largely unknown.
MicroRNAs (miRNAs/miRs) are small non-coding RNA
molecules that serve as endogenous regulators of gene expression
(26) and are involved in a variety
of vital biological processes (27–29).
Previous studies have demonstrated that most miRNAs inhibit the
metastasis of breast cancer and that their expression levels can be
used as metastatic predictors. For example, miR-124 is considered
an expression-rich miRNA (30–33),
which is usually expressed at low levels and is implicated in
hematological malignancies and solid tumors, including breast
cancer (34–36). A previous study has reported that
miR-124 expression is notably downregulated following treatment
with E2 in ER positive breast cancer cells (37). CD151 is a member of tetraspanins that
is characterized by 4 hydrophobic domains, and mediates signal
transduction events associated with cell proliferation, activation
and motility (38). It has been
demonstrated that the CD151 protein can accelerate the progression
of breast cancer (39). To the best
of our knowledge, miR-124 is a negative regulator of breast cancer
and it was hypothesized that E2 may regulate miR-124 and CD151
expression levels by GPER, which in turn will affect the
proliferation, invasion and migration of breast cancer cells.
In the present study, the potential internal
molecular mechanism was investigated with regards to the
proliferation, invasion and migration of breast cancer cells. The
experiments aimed to investigate the interaction between E2-GPER
signaling and alteration in the expression levels of miR-124 and
CD151, and the progression of breast cancer cell metastasis. The
results of the present study may provide a potential novel approach
to the treatment and prognosis of breast carcinomas.
Materials and methods
Cell culture and treatment
The human ER-positive breast cancer cell line,
MCF-7, was purchased from Type Culture Collection of the Chinese
Academy of Sciences (Shanghai, China) and stored in liquid
nitrogen. Cells were maintained in Dulbecco's modified Eagle's
medium (DMEM, Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with high glucose, 10% fetal bovine serum (FBS;
HyClone; Cytiva) and 1% penicillin-streptomycin (Invitrogen; Thermo
Fisher Scientific, Inc.), at 37°C with 5% CO2 in a humid
environment.
The E2 (5, 10 or 100 nM, Sigma-Aldrich; Merck
KGaA)-treated cells were maintained in complete medium with phenol
red (Sigma-Aldrich; Merck KGaA) until they reached 60% confluence
and subsequently replaced with 2.5% CS-FBS (HyClone; Cytiva) in
DMEM containing phenol red-free for 24 h. The dose of E2 was in
accordance with previous studies (40,41).
Cells were cultured for 24 h at 37°C in serum-free and phenol-red
free DMEM. Subsequently, a certain dose of E2 (5, 10 or 100 nM) was
added into the medium. G15 was used as a well-established
antagonist of GPER in this experiment (42). To determine whether E2-induced
miR-124 downregulation was directly regulated by GPER, MCF-7 cells
were pretreated with G15 (100 nM) for 6 h prior to addition of
E2.
Cell transfection
MCF-7 cells were incubated for 24 h prior to
transfection. miR-124 negative control (mimic-NC; forward,
5′-UUCUCCGAACGUGUCACGUTT−3′ and reverse,
5′-ACGUGACAGGUUCGGAGAATT−3′; 50 nM), miR-124 mimic (forward,
5′-UAAGGCACGCGGUGAAUGCCAA-3′ and reverse,
5′-CAUUCACCGCGUGCCUUAUU-3′; 50 nM), CD151-small interfering (si)RNA
(50 nM) and negative control (siRNA-NC; 50 nM) sequences were
synthesized by Shanghai GenePharma, Co., Ltd. Cells were seeded
into 6-well plates and cultured until they reached 60–70%
confluence. Transfection was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 24 h, according to the manufacturer's
instructions. Subsequent experiments were performed 24 h
post-transfection.
Cell proliferation assay
Cell proliferation was assessed via the Cell
Counting Kit-8 (CCK-8) assay (Dojindo Molecular Technologies,
Inc.). The cell suspension was seeded into a 96-well plate and
cultured for 6 h at 37°C. Subsequently, cells were incubated with
10 µl CCK-8 solution for 1 h and cell proliferation was measured at
a wavelength of 450 nm (A450), using a microplate
reader.
Colony formation assay
MCF-7 cells were seeded into 6-well plates at a
density of 500 cells/well and cultured at 37°C in the presence of
5% CO2 for 7–12 days. Cells were subsequently fixed with
4% paraformaldehyde for 10 min at room temperature and stained with
0.2% crystal violet for 5 min at room temperature. The number of
cell colonies were manually counted using ImageJ software (version
1.52r; National Institutes of Health).
Invasion assay
The Transwell assay was performed to assess cell
invasion. A total of 1×106 MCF-7 cells were plated in
the upper chambers of Transwell plates in serum-free medium (DMEM,
Invitrogen; Thermo Fisher Scientific, Inc.). The Transwell
membranes were pre-coated with 100 µl cell suspension and Matrigel
(BD Biosciences) overnight at 4°C. DMEM supplemented with 10% FBS
was plated in the lower chambers. Following incubation at 37°C with
5% CO2 for 24 h, cells that did not pass through the
polycarbonate membrane were removed. The invasive cells were fixed
with 4% paraformaldehyde for 30 min at room temperature, stained
with 1% Giemsa for 15 min at room temperature, rinsed using
phosphate-buffered saline (PBS) and air dried. All experiments were
performed in triplicate. Stained cells were counted in five
randomly selected fields using an inverted light microscope
(Olympus Corporation; magnification, ×100) and the results were
analyzed using ImageJ software (version 1.52r; National Institutes
of Health).
Cell migration assay
The cell migratory ability was assessed via the
wound healing assay. Briefly, cells were seeded into a 6-well plate
(4×105 cells/well) and incubated at 37°C until they
reached 80% confluence. Subsequently, the cell monolayers were
scratched using a plastic scriber. Cells were washed with PBS to
elute the debris and the media were replaced with serum-free
medium. The average distance that the cells migrated into the wound
surface was detected under an inverted light microscope (Olympus
Corporation; magnification, ×100) after 48 h.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from MCF-7 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions.
Single-stranded cDNA was synthesized at 42°C for 30 min using the
one-step cDNA synthesis kit (cat. no. 210210; Qiagen, Inc.). The
following thermocycling conditions were used: 40 cycles of pre-
denaturation at 95°C for 10 min, denaturation at 95°C for 30 sec,
annealing at 60°C for 20 sec and extension at 72°C for 35 sec. The
following primer sequences were used: miR-124 forward,
5′-ACGTTGTGTAGCTTATCAGACTG-3′ and reverse,
5′-AATGGTTGTTCTCCACACTCTC-3′; CD151 forward,
5′-ACAGCCTACATCCTGGTGGT-3′ and reverse, 5′-TTCTCCTTGAGCTCCGTGTT-3′;
U6 forward, 5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GGAACGCTTCACGAATTTG-3′; and GAPDH forward,
5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse,
5′-GCCATCACGCCACAGTTTC-3′. Relative expression levels were
calculated using the 2−ΔΔCq method (43).
Western blotting
Anti-GPER rabbit polyclonal antibody (cat. no.
D161727; 1:350), goat anti-rabbit IgG antibody (cat. no. D111018;
1:8,000), anti-matrix metalloproteinase (MMP)2 (cat. no. D161447;
1:500), anti-MMP9 (cat. no. D162000; 1:500) and anti-GAPDH (cat.
no. D110016; 1:500) antibodies were purchased from Sangon Biotech
Co., Ltd. Cells were washed with PBS and total proteins were lysed
from the cells using SDS lysis buffer (Beyotime Institute of
Biotechnology) supplemented with proteinase inhibitor cocktail
(Sigma-Aldrich, Merck KGaA). Total protein was quantified using the
bicinchoninic acid kit (Beyotime Institute of Biotechnology) and 40
µg protein/lane was fractionated with 8% SDS-PAGE. The separated
proteins were subsequently transferred onto polyvinylidene fluoride
membranes (EMD Millipore) and subsequently incubated in PBS 0.2%
Tween-20 containing 5% skimmed milk powder for 60 min at room
temperature to block non-specific binding. Following washing, the
blots were incubated overnight at 4°C with anti-GPER rabbit
polyclonal antibody and GAPDH (cat. no. D110016; 1:500; Sangon
Biotech Co., Ltd.) was used as the reference protein. Subsequently,
the membranes were washed with PBS Tween-20 and incubated with goat
anti-rabbit IgG antibody for 2 h at room temperature. Protein bands
were visualized using an enhanced chemiluminescence system ChemiDoc
MP (Bio-Rad Laboratories, Inc.) and quantified using ImageJ
software (version 1.46; National Institutes of Health).
Dual-luciferase reporter assay
The target gene of miR-124 was predicted using the
TargetScan database (http://www.targetscan.org/vert_71). A dual-luciferase
reporter assay system (Promega Corporation) was used to detect
luciferase activities, according to the manufacturer's
instructions, using the Envision Multilabel Plate Reader
(PerkinElmer, Inc.). The CD151 3′-untranslated region (UTR) was
amplified from human cDNA by Shanghai GenePharma Co., Ltd., cloned
into a pGL3 luciferase vector (Promega Corporation). The following
sequences were used: i) Wild-type (WT) of CD151 3′-UTR forward,
5′-TCTAGAACCCAACTACTGAGCTGAGA-3′ and reverse,
5′-TCTAGAGTACAGCAGTGAACAAAACCA−3′; and ii) mutant type (MUT) of
CD151 3′-UTR forward,5′-CTTCTTCCGAGTTTTGCTGCGCACCAATGC-3′ and
reverse, 5′-AGCAAAACTCGGAAGAAGCTGCCTCTGAGGT-3′. Following seeding
into 24-well plates and incubation overnight, cells were
co-transfected with miR-124 mimic or mimic-NC for 48 h at 37°C,
using Lipofectamine® 2000. The Renilla luciferase
activities were normalized to the firefly luciferase activities to
set the control samples for the transfection efficiency
experiments. All experiments were performed in triplicate.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 6.0 software. All experiments were performed in triplicate
and data are presented as the mean ± standard deviation. Unpaired
student's t-test was used to compare differences between two
groups, while one way ANOVA and Tukey's post hoc test were used to
compare differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
E2 induces miR-124 expression in MCF-7
cells
MCF-7 cells were treated with E2 at different time
points and concentrations and miR-124 expression was detected. The
results demonstrated that miR-124 expression significantly
decreased following treatment with E2 compared with the control
group (P<0.01, P<0.001; Fig.
1). The concentration of E2 was selected based on the lowest
level of miR-124 expression. The dose-dependent experiments
revealed that miR-124 expression considerably decreased in MCF-7
cells following treatment with 10 nM E2 for 8 h compared with the
control group. Thus, 10 nM E2 was selected for subsequent
experiments, which was in accordance with a previous study
(44).
Induction of GPER by E2
MCF-7 cells were treated with 10 nM E2 at different
time points (0, 6, 8 and 12 h) and GPER expression was detected.
The results demonstrated that GPER expression was upregulated in
MCF-7 cells following treatment with E2 for 6, 8 and 12 h, and
maximum GPER expression was attained following treatment for 8 h
(P<0.001; Fig. 2). Thus, cells
treated with E2 for 8 h was selected for subsequent
experiments.
GPER mediates the E2-induced decrease
in miR-124 expression
The effects of GPER on miR-124 expression were
investigated, which were induced by E2 in MCF-7 cells. G15 was used
as a well-established antagonist of GPER (41) in this experiment. To determine
whether E2-induced miR-124 downregulation was directly regulated by
GPER, MCF-7 cells were pretreated with G15 (100 nM) for 6 h prior
to addition of E2. The results demonstrated that miR-124 expression
increased in the E2 + G15 group compared with the E2 group
(P<0.05; Fig. 3A), suggesting
that GPER mediates the E2-induced decrease of miR-124
expression.
GPER mediates the E2-induced
proliferation, invasion and migration of MCF-7 cells
Cell proliferation was assessed via the CCK-8 assay.
The results indicated that the proliferation of MCF-7 cells in the
E2 + G15 group was significantly inhibited compared with the E2
group (P<0.01, P<0.001; Fig. 3B
and C). In addition, the results of the Transwell assays
demonstrated that the number of invasive cells in the E2 group was
significantly higher compared with the control group, the effects
of which were reversed following treatment with G15 (P<0.01;
Fig. 3D and E). The wound healing
assay was performed to assess cell migration. As presented in
Fig. 4A and B, the ability of cell
migration in the E2 group was notably enhanced compared with the
control group, whereas G15 intervention attenuated the effect of E2
on cell migration. The assessment of the expression of
migration-associated proteins, including matrix metalloproteinase
(MMP)2 and MMP9 exhibited similar results to the cell invasion
assay (Fig. 4A-C). Taken together,
these results suggest that GPER mediates the E2-induced
proliferation, invasion and migration of MCF-7 cells.
CD151 is a direct target of miR-124 in
breast cancer cells
The TargetScan database was used to predict the
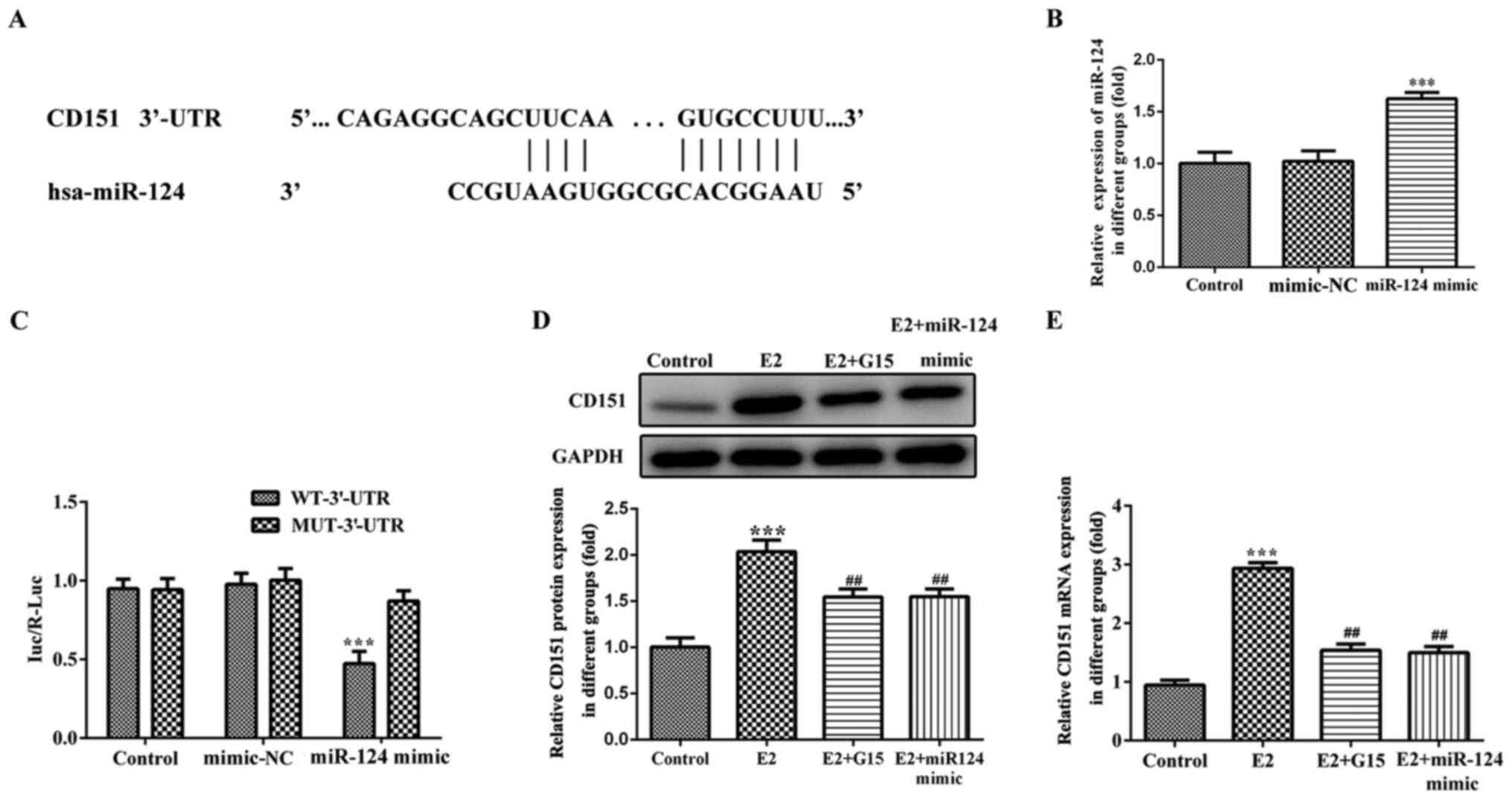
target of miR-124. miR-124 was bound to the 3′-UTR of CD151
(Fig. 5A). Cells were transfected
with miR-124 mimic to overexpress miR-124 (Fig. 5B). To validate whether CD151 was
regulated by miR-124 in MCF-7 cells, reporter plasmids were
generated (WT-CD151 3′-UTR or MUT-CD151 3′-UTR). The results
demonstrated that the luciferase activity of CD151 decreased
following co-transfection of miR-124 mimic with WT-CD151 3′-UTR in
MCF-7 cells compared with the control mimic group. Notably, no
significant differences were observed in the luciferase activity
following co-transfection of miR-124 mimic with MUT-CD151 3′-UTR in
MCF-7 cells (Fig. 5C). To
investigate whether CD151 was regulated via the inhibition of
miR-124 following GPER induced expression by E2 in MCF-7 cells,
western blot and RT-qPCR analyses were performed to detect CD151
expression. In this experiment, MCF-7 cells were treated with 100
nM G15 for 2 h or pre-transfected with miR-124 mimic for 12 h prior
to addition of 10 nM E2 for 8 h. The results demonstrated that
CD151 protein and mRNA expression levels significantly increased in
the E2 group compared with the control group (Fig. 5D and E). However, the effects of E2
on CD151 expression were blocked following pretreatment with
miR-124 mimic or G15. Taken together, these results suggest that
CD151 is a direct target gene of miR-124, which is consistent with
previous findings (26).
Effects of CD151 on the proliferation,
invasion and migration of MCF-7 cells following treatment with
E2
To investigate the proliferation, invasion and
migration of E2-treated MCF-7 cells, cells were transfected with
CD151-siRNA. RT-qPCR analysis was performed to verify CD151
knockdown in MCF-7 cells and the results demonstrated that CD151
expression significantly decreased following transfection with
CD151-siRNA (Fig. 6A). The effect of
CD151 knockdown on cell proliferation was assessed via the CCK-8
assay. The results demonstrated that cell proliferation
significantly decreased in the CD151-siRNA group compared with the
control group (Fig. 6B and C). The
invasive (Fig. 6D and E) and
migratory (Fig. 7A and B) abilities
of MCF-7 cells following CD151 interference were assessed via the
Transwell and wound healing assays, respectively. The results
demonstrated that cell invasion and migration significantly
decreased following transfection with CD151-siRNA. Collectively,
these results suggest that the proliferation, invasion and
migration of E2-induced MCF-7 cells are inhibited following CD151
interference.
Discussion
Breast cancer is one of the most common malignancies
worldwide that is responsible for a high number of fatalities
(45). The number of new breast
cancer cases is estimated at 272,400 in China, annually (46). The identification of novel and
efficacious methods for early diagnosis and treatment of this
disease remains a major challenge. In the present study, the
molecular mechanism of the proliferation, invasion and migration of
breast cancer cells was investigated.
Estrogens play an important role in breast cancer
metastasis (47–49). E2 is one of the typical estrogens
that is responsible for the occurrence of 70–80% of human breast
cancer and has been generally investigated for its contribution in
the development of the human mammary gland (50). It is considered that GPER can mediate
estrogenic signals in breast cancer (51). Previous studies have demonstrated
that GPER is bound with E2, owing to its high affinity (20,52,53).
Thus, it is important to investigate the effects of E2-GPER
signaling on the proliferation, invasion and migration of breast
cancer cells. A previous study reported that GPER expression is
upregulated in patients with breast cancer (54). Another study indicated that
E2-treated cancer-associated fibroblasts exhibit a positive
feedback behavior, which involves GPER/EGFR/ERK signaling and E2
production, and contributes to the progression of breast cancer
(55). Breast cancer progression is
associated with overexpression of GPER and PM localization, whereas
the lack of GPER is associated with the long-term prognosis of
primary breast cancer in PM (56).
However, the molecular mechanism underlying the proliferation,
invasion and migration of breast cancer cells via E2-GPER signaling
remains largely unknown. The results of the present study
demonstrated that GPER-E2 signaling decreased miR-124 expression,
suggesting that inhibition of this target may be used as a novel
treatment in breast carcinoma.
miRNAs are important post-transcriptional regulators
of gene expression that play a major role in carcinogenesis
(28,29). It has been demonstrated that miR-124
acts as a tumor suppressor that can inhibit the proliferation,
invasion and migration of breast cancer cells (37). A previous study reported that miR-124
targets Slug to regulate epithelial-to-mesenchymal transition and
metastasis of breast cancer (57).
Feng et al (58) discovered
that downregulation of miR-124 results in breast cancer progression
via regulation of long non-coding RNA-MALAT1 and activation of
CDK4/E2F1 signaling. In addition, another
study demonstrated that downregulation of miR-124-3p promotes
breast cancer progression by targeting Beclin-1 (59). Thus, it is crucial to investigate the
expression levels and regulation of miR-124 in breast cancer. Taken
together, the results of the present study demonstrated that E2
inhibited miR-124 expression via the receptor, GPER. Furthermore,
E2-GPER signaling promoted CD151 expression by suppressing miR-124
expression.
In conclusion, the present study demonstrated that
activation of GPER was mediated by E2 and promoted the
proliferation, invasion and migration of breast cancer cells via
regulation of the miR-124/CD151 pathway. These findings may provide
a novel therapeutic strategy for the treatment and prognosis of
breast cancer. However, use of a single breast cancer cell line is
a major limitation of the present study. Thus, prospective studies
will focus on using different clinical types of breast cancer
cells.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and or/analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
HY and CW designed and performed the experiments. HL
drafted the initial manuscript. HY, HL and QW contributed to data
collection, analysis and interpretation. QW reviewed and edited the
manuscript. HY and QW confirmed the authenticity of all the raw
data. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Ma J, Goding Sauer A, Newman
LA and Jemal A: Breast cancer statistics, 2017, racial disparity in
mortality by state. CA Cancer J Clin. 67:439–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartelink H, Horiot JC, Poortmans P,
Struikmans H, Van den Bogaert W, Barillot I, Fourquet A, Borger J,
Jager J, Hoogenraad W, et al: Recurrence rates after treatment of
breast cancer with standard radiotherapy with or without additional
radiation. N Engl J Med. 345:1378–1387. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mattiuzzi C and Lippi G: Current cancer
epidemiology. J Epidemiol Glob Health. 9:217–222. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clayforth C, Fritschi L, McEvoy SP, Byrne
MJ, Ingram D, Sterrett G, Harvey JM, Joseph D and Jamrozik K:
Five-year survival from breast cancer in Western Australia over a
decade. Breast. 16:375–381. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan J, Xiao C, Lu H, Yu H, Hong H, Guo C
and Wu Z: miR-200b regulates breast cancer cell proliferation and
invasion by targeting radixin. Exp Ther Med. 19:2741–2750.
2020.PubMed/NCBI
|
|
8
|
Si CS, Yu Q and Yao YF: Effect of
miR-146a-5p on proliferation and metastasis of triple-negative
breast cancer via regulation of SOX5. Exp Ther Med. 15:4515–4521.
2018.PubMed/NCBI
|
|
9
|
Yager JD and Davidson NE: Estrogen
carcinogenesis in breast cancer. N Engl J Med. 354:270–282. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Holden MP and Hampson E: Endogenous
variation in estradiol in women affects the weighting of metric and
categorical information in spatial location memory. Horm Behav.
128:62021. View Article : Google Scholar
|
|
11
|
Mantri AV, Allaway HCM, Brezicha JE, Hogan
HA and Bloomfield SA: Oral estradiol impact on mitigating
unloading-induced bone loss in ovary-intact rats. Aerosp Med Hum
Perform. 92:65–74. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fang GZ, Huang GY, Ying GG, Qiu SQ, Shi
WJ, Xie L, Yang YY and Ma DD: Endocrine disrupting effects of
binary mixtures of 17β-estradiol and testosterone in adult female
western mosquitofish (Gambusia affinis). Ecotox Environ Safe.
208:1115662021. View Article : Google Scholar
|
|
13
|
Huang S, Liu M, Fu F, Liu H, He B, Xiao D
and Yang J: High serum estradiol reduces acute hepatotoxicity risk
induced by epirubicin plus cyclophosphamide chemotherapy in
premenopausal women with breast cancer. Front Pharmacol.
11:5724442021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shang D, Li Z, Zhu Z, Chen H, Zhao L, Wang
X and Chen Y: Baicalein suppresses 17-β-estradiol-induced
migration, adhesion and invasion of breast cancer cells via the G
protein -coupled receptor 30 signaling pathway. Oncol Rep.
33:2077–2085. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ignatov A, Ignatov T, Roessner A, Costa SD
and Kalinski T: Role of GPR30 in the mechanisms of tamoxifen
resistance in breast cancer MCF-7 cells. Breast Cancer Res Treat.
123:87–96. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Muhammad JS, Guimei M, Jayakumar MN,
Shafarin J, Janeeh AS, AbuJabal R, Eladl MA, Ranade AV, Ali A and
Hamad M: Estrogen-induced hypomethylation and overexpression of
YAP1 facilitate breast cancer cell growth and survival. Neoplasia.
23:68–79. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng G, Cai J, Huang Y, Zhu X, Gong B,
Yang Z, Yan C, Hu Z, Yang L and Wang Z: G-protein-coupled estrogen
receptor 1 promotes gender disparities in hepatocellular carcinoma
via modulation of SIN1 and mTOR complex 2 activity. Mol Cancer Res.
18:1863–1875. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maggiolini M, Vivacqua A, Fasanella G,
Recchia AG, Sisci D, Pezzi V, Montanaro D, Musti AM, Picard D and
Andò S: The G protein -coupled receptor GPR30 mediates c-fos
up-regulation by 17beta-estradiol and phytoestrogens in breast
cancer cells. J Biol Chem. 279:27008–27016. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vivacqua A, Bonofiglio D, Recchia AG,
Musti AM, Picard D, Andò S and Maggiolini M: The G protein -
coupled receptor GPR30 mediates the proliferative effects induced
by 17beta-estradiol and hydroxytamoxifen in endometrial cancer
cells. Mol Endocrinol. 20:631–646. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Revankar CM, Cimino DF, Sklar LA,
Arterburn JB and Prossnitz ER: A transmembrane intracellular
estrogen receptor mediates rapid cell signaling. Science.
307:1625–1630. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Filardo EJ, Quinn JA, Bland KI and
Frackelton AR Jr: Estrogen-induced activation of Erk-1 and Erk-2
requires the G protein -coupled receptor homolog, GPR30, and occurs
via trans-activation of the epidermal growth factor receptor
through release of HB-EGF. Mol Endocrinol. 14:1649–1660. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Albanito L, Madeo A, Lappano R, Vivacqua
A, Rago V, Carpino A, Oprea TI, Prossnitz ER, Musti AM, Andò S and
Maggiolini M: G protein -coupled receptor 30 (GPR30) mediates gene
expression changes and growth response to 17beta- estradiol and
selective GPR30 ligand G-1 in ovarian cancer cells. Cancer Res.
67:1859–1866. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vivacqua A, Bonofiglio D, Albanito L,
Madeo A, Rago V, Carpino A, Musti AM, Picard D, Andò S and
Maggiolini M: 17beta-estradiol, genistein, and 4-hydroxytamoxifen
induce the proliferation of thyroid cancer cells through the G
protein -coupled receptor GPR30. Mol Pharmacol. 70:1414–1423. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thomas P, Pang Y, Filardo EJ and Dong J:
Identity of an estrogen membrane receptor coupled to a G protein in
human breast cancer cells. Endocrinology. 146:624–632. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Q, Li JG, Zheng XY, Jin F and Dong HT:
Expression of CD133, PAX2, ESA, and GPR30 in invasive ductal breast
carcinomas. Chin Med J (Engl). 122:2763–2769. 2009.PubMed/NCBI
|
|
26
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu S, Chen X, Zhang S, Wang X, Du X, Chen
J and Zhou G: miR-106b-5p targeting SIX1 inhibits TGF-β1-induced
pulmonary fibrosis and epithelial-mesenchymal transition in asthma
through regulation of E2F1. Int J Mol Med. 47:12021. View Article : Google Scholar
|
|
28
|
Wan JH and Liu SS: miR-613 inhibits the
proliferation of human ovarian granulosa cells by arresting cell
cycle progression via the targeting of IGF-1. Mol Med Rep.
23:12021.
|
|
29
|
Dong B, Li SY, Zhu SL, Yi M, Luo S and Wu
K: miRNA- mediated EMT and CSCs in cancer chemoresistance. Exp
Hematol Oncol. 10:122021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng LC, Pastrana E, Tavazoie M and
Doetsch F: miR-124 regulates adult neurogenesis in the
subventricular zone stem cell niche. Nat Neurosci. 12:399–408.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu JY, Chung KH, Deo M, Thompson RC and
Turner DL: MicroRNA miR-124 regulates neurite outgrowth during
neuronal differentiation. Exp Cell Res. 314:2618–2633. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Makeyev EV, Zhang J, Carrasco MA and
Maniatis T: The MicroRNA miR-124 promotes neuronal differentiation
by triggering brain-specific alternative pre-mRNA splicing. Mol
Cell. 27:435–448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mishima T, Mizuguchi Y, Kawahigashi Y and
Takizawa T and Takizawa T: RT-PCR-based analysis of microRNA (miR-1
and-124) expression in mouse CNS. Brain Res. 1131:37–43. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Silber J, Lim DA, Petritsch C, Persson AI,
Maunakea AK, Yu M, Vandenberg SR, Ginzinger DG, James CD, Costello
JF, et al: miR-124 and miR-137 inhibit proliferation of
glioblastoma multiforme cells and induce differentiation of brain
tumor stem cells. BMC Med. 6:142008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wong KY, So CC, Loong F, Chung LP, Lam WW,
Liang R, Li GK, Jin DY and Chim CS: Epigenetic inactivation of the
miR-124-1 in haematological malignancies. PLoS One. 6:e190272011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xia J, Wu Z, Yu C, He W, Zheng H, He Y,
Jian W, Chen L, Zhang L and Li W: miR-124 inhibits cell
proliferation in gastric cancer through down-regulation of SPHK1. J
Pathol. 227:470–480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang CF, Li DM, Shi ZM, Wang L, Liu MM,
Ge X, Liu X, Qian YC, Wen YY, Zhen LL, et al: Estrogen regulates
miRNA expression: Implication of estrogen receptor and miR-124/AKT2
in tumor growth and angiogenesis. Oncotarget. 7:36940–36955. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Lei W, Li G, Ma H, Guo H and Li S:
CD151 promotes proliferation and migration of SK-NEP-1 cells via
the GSK-3β/P21/cyclinD signaling pathway. Pathol Res Pract.
215:329–334. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Han ZB, Yang Z, Chi Y, Zhang L, Wang Y, Ji
Y, Wang J, Zhao H and Han ZC: MicroRNA-124 suppresses breast cancer
cell growth and motility by targeting CD151. Cell Physiol Biochem.
31:823–832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li Q, Gao H, Yang H, Wei W and Jiang Y:
Estradiol promotes the progression of ER+ breast cancer through
methylation- mediated RSK4 inactivation. Onco Targets Ther.
12:5907–5916. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Khan S, Abdelrahim M, Samudio I and Safe
S: Estrogen receptor/Sp1 complexes are required for induction of
cad gene expression by 17beta-estradiol in breast cancer cells.
Endocrinology. 144:2325–2335. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dennis MK, Burai R, Ramesh C, Petrie WK,
Alcon SN, Nayak TK, Bologa CG, Leitao A, Brailoiu E, Deliu E, et
al: In vivo effects of a GPR30 antagonist. Nat Chem Biol.
5:421–427. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hayashi T, Hikichi M, Yukitake J, Harada N
and Utsumi T: Estradiol suppresses phosphorylation of ERα serine
167 through upregulation of PP2A in breast cancer cells. Oncol
Lett. 14:8060–8065. 2017.PubMed/NCBI
|
|
45
|
Emens LA: Breast cancer immunotherapy:
Facts and hopes. Clin Cancer Res. 24:511–520. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen WQ, Zheng RS, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yuan S, Huang C, Ji X, Ma M, Rao K and
Wang Z: Prediction of the combined effects of multiple estrogenic
chemicals on MCF-7 human breast cancer cells and a preliminary
molecular exploration of the estrogenic proliferative effects and
related gene expression. Ecotoxicol Environ Saf. 160:1–9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Parl FF, Crooke PS, Plummer WD Jr and
Dupont WD: Genomic-epidemiologic evidence that estrogens promote
breast cancer development. Cancer Epidemiol Biomarkers Prev.
27:899–907. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sampayo RG, Toscani AM, Rubashkin MG, Thi
K, Masullo LA, Violi IL, Lakins JN, Cáceres A, Hines WC, Coluccio
Leskow F, et al: Fibronectin rescues estrogen receptor a from
lysosomal degradation in breast cancer cells. J Cell Biol.
217:2777–2798. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Meneses-Morales I, Tecalco-Cruz AC,
Barrios-García T, Gómez-Romero V, Trujillo-González I,
Reyes-Carmona S, García-Zepeda E, Méndez-Enríquez E,
Cervantes-Roldán R, Pérez-Sánchez V, et al: SIP1/NHERF2 enhances
estrogen receptor alpha transactivation in breast cancer cells.
Nucleic Acids Res. 42:6885–6900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liang S, Chen Z, Jiang G, Zhou Y, Liu Q,
Su Q, Wei W, Du J and Wang H: Activation of GPER suppresses
migration and angiogenesis of triple negative breast cancer via
inhibition of NF-κB/IL-6 signals. Cancer Lett. 386:12–23. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Moriarty K, Kim KH and Bender JR:
Minireview: Estrogen receptor-mediated rapid signaling.
Endocrinology. 147:5557–5563. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Prossnitz ER, Arterburn JB and Sklar LA:
GPR30: A G protein -coupled receptor for estrogen. Mol Cell
Endocrinol. 265-266:138–142. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Girgert R, Emons G and Gründker C:
Inhibition of growth hormone receptor by Somavert reduces
expression of GPER and prevents growth stimulation of
triple-negative breast cancer by 17β-estradiol. Oncol Lett.
15:9559–9566. 2018.PubMed/NCBI
|
|
55
|
Luo H, Yang G, Yu T, Luo S, Wu C, Sun Y,
Liu M and Tu G: GPER-mediated proliferation and estradiol
production in breast cancer-associated fibroblasts. Endocr Relat
Cancer. 21:355–369. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yu T, Liu M, Luo H, Wu C, Tang X, Tang S,
Hu P, Yan Y, Wang Z and Tu G: GPER mediates enhanced cell viability
and motility via non-genomic signaling induced by 17β- estradiol in
triple-negative breast cancer cells. J Steroid Biochem Mol Biol.
143:392–403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liang YJ, Wang QY, Zhou CX, Yin QQ, He M,
Yu XT, Cao DX, Chen GQ, He JR and Zhao Q: miR-124 targets Slug to
regulate epithelial-mesenchymal transition and metastasis of breast
cancer. Carcinogenesis. 34:713–722. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Feng T, Shao F, Wu Q, Zhang X, Xu D, Qian
K, Xie Y, Wang S, Xu N, Wang Y and Qi C: miR-124 downregulation
leads to breast cancer progression via LncRNA-MALAT1 regulation and
CDK4/E2F1 signal activation. Oncotarget. 7:16205–16216. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang F, Wang B, Long H, Yu J, Li F, Hou H
and Yang Q: Decreased miR-124-3p expression prompted breast cancer
cell progression mainly by targeting Beclin-1. Clin Lab.
62:1139–1145. 2016. View Article : Google Scholar : PubMed/NCBI
|