Introduction
Cervical cancer (CC) is the second most common
malignant tumor diagnosed in women worldwide (1,2). The
recognition of human papillomavirus (HPV) as a primary cause of CC
led to the development of HPV testing. There has been a shift from
reflex HPV testing for mild cytological abnormalities, to
co-testing with cytology and HPV, and lately to primary HPV
screening (3). In the past decade,
the popularization of CC screening has effectively improved the
efficiency of diagnosis and treatment, while reducing the mortality
rate. However, the number of new CC cases exhibits an annual
increasing trend worldwide. In China, it is estimated that nearly
100,000 new diagnosed cases and >30,000 cases of CC-associated
mortality occur each year (4).
Advances in surgery, chemotherapy and postoperative radiotherapy
have contributed to a 5-year survival rate of ~80%. However, the
actual long-term outcome of clinical treatment is not satisfactory.
The high metastatic rate of CC reduces this result to <50%
(5,6). The clinical difficulties in CC
treatment highlight the importance of identifying novel key
biomarkers of CC, unraveling the pathogenetic mechanism of this
disease and improving the treatment strategies.
The Eph receptor A2 (EphA2) is a transmembrane
receptor tyrosine kinase, which has been shown to be elevated in
several types of cancer. For example, it is positively correlated
with the aggressive behavior of certain cancer cells, including
malignant melanomas, breast cancer, pulmonary cancer, prostate
cancer and esophageal cancer (6–11).
These previous studies have reported high expression levels of
EphA2 in both tissue and cell models of these diseases. This
elevated expression level was shown to be associated with high
histological grade, lymph node metastasis and poor prognosis in the
aforementioned tumors (6–11). Upregulation of EphA2 and moderate
or high expression of EphrinA-1 in squamous cervical carcinoma are
associated with shorter overall survival, suggesting that they
could be used as valuable prognostic markers (12). ALW-II-41-27 is a newly developed
ATP-competitive EphA2 inhibitor (13). ALW-II-41-27 and small interfering
RNA (siRNA)-based downregulation of EphA2 can effectively inhibit
conditioned medium of cancer-associated fibroblast-induced gastric
tumorigenesis (14). EphA2-siRNA
inhibits proliferation and induces cell cycle arrest in SBC-5 cells
by interfering with EphA2 gene expression (15). Similarly, ALW-II-41-27 and the
tyrosine kinase inhibitor dasatinib inhibit the proliferation of
EphA2-positive small cell lung cancer cells by downregulating EphA2
gene expression (15). Based on
this evidence, the present study aimed to explore the mechanism by
which ALW-II-41-27 may inhibit tumor progression in CC.
According to a recent study, EphA2 antagonism can
notably inhibit lipopolysaccharide-induced Ras homolog family
member A (RhoA)/Rho-associated protein kinase (ROCK) activation in
lung tissues, whereas antagonism of EphA2 can be used as a
therapeutic method for acute lung injury (16). In addition, G12 protein stimulation
can promote CC invasion via RhoA/ROCK/JNK activation (17). Inhibition of triple functional
domain protein expression markedly decreases the migratory and
invasive abilities of CC cells and the expression levels of the
RhoA/ROCK signal transduction factors [RhoA, ROCK and
phosphorylated (p)-LIM kinase] that in turn contribute to cell
migration and invasion (18). It
has been demonstrated that human squamous CC tissues overexpress
RhoA and ROCK-I/II, whereas RhoA promotes CC cell progression and
tumor growth (19). RhoA/ROCK is
therefore considered as an important pathway associated with
EphA2-related CC development.
The present study aimed to explore the role of
ALW-II-41-27 in human CC cells and to determine its underlying
mechanism. The findings from this study may provide further insight
into the mechanism of CC progression and offer a promising
therapeutic target for the treatment of this disease
Materials and methods
Cell culture and treatment
The human CC HeLa, CaSki and C33A cell lines, and
the non-cancerous ectocervical epithelial Ect1/E6E7 cell line, were
purchased from The Cell Bank of the Type Culture Collection of The
Chinese Academy of Sciences. These cell lines were maintained in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin/streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) and placed at 37°C in a humidified incubator
containing 5% CO2.
The EphA2 inhibitor ALW-II-41-27 (MedChemExpress)
was dissolved in sterile DMSO (10 mmol/l stock solution) and stored
in aliquots at −20°C. CaSki and HeLa cells were treated with 200,
600 or 1,000 nM ALW-II-41-27 for 24, 48 or 72 h (20). The activation of the RhoA/ROCK
pathway was achieved by pretreatment of these two cells with the
RhoA/ROCK signaling agonist phorbol 12-myristate 13-acetate (PMA;
100 ng/ml; Sigma-Aldrich; Merck KGaA) for 30 min. The expression
levels of EphA2, RhoA and ROCK were attenuated following treatment
of these two cells with the RhoA inhibitor C3 transferase (5 µg/ml;
Cytoskeleton Inc.) or ROCK blocker hydroxyfasudil (10 µM; Tocris
Bioscience) for 2 h. These compounds were selected for cell
pretreatment based on a previous study (21).
Reverse transcription-quantitative
(RT-q)PCR
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used for total RNA extraction from all
types of cells according to standard procedures. The detection of
EphA2 was performed via RT-qPCR using a high-capacity cDNA Reverse
Transcription kit (Qiagen AB) and SYBR Premix Ex Taq (Qiagen AB).
The conditions for cDNA synthesis were as follows: 42°C for 30 min
and 85°C for 5 sec. The thermocycling conditions were as follows:
95°C for 3 min; 39 cycles of 95°C for 5 sec, 56°C for 10 sec, 72°C
for 25 sec; 65°C for 5 sec; 95°C for 50 sec. The relative
expression levels were normalized to endogenous control GAPDH and
were expressed as 2−ΔΔCq (22). The sequences of the primers were as
follows: EphA2 forward, 5′-CTGGTCTGCAAGGTGTCTGA-3′ and reverse,
5′-TTGGACAACTCCCAGTAGGG-3′; and GADPH forward,
5-GATATTGTTGCCATCAATGAC-3 and reverse 5-TTGATTTTGGAGGGATCTCG-3.
Western blotting
All types of cells were separately lysed using RIPA
lysis buffer (Sigma-Aldrich; Merck KGaA) and protein concentration
was determined using the BCA method (Beyotime Institute of
Biotechnology). Proteins (20 µg/lane) were separated by 10%
SDS-PAGE and transferred onto PVDF membranes (MilliporeSigma).
Membranes were blocked in 5% milk for 2 h at room temperature. The
membranes were incubated at 4°C overnight with primary antibodies
against EphA2 (cat. no. ab185156; 1:1,000; Abcam), GTP-RhoA (cat.
no. 211164; 1:500; Abcam), ROCK1 (cat. no. 07-1458; 1:500;
MilliporeSigma), total-RhoA (cat. no. ABS620; 1:400;
MilliporeSigma) and GAPDH (cat. no. ab181602; 1:10,000; Abcam).
Next, membranes were incubated with secondary anti-rabbit (cat. no.
ab6721; 1:2,000; Abcam) or anti-mouse (cat. no. ab6728; 1:2,000;
Abcam) antibodies at room temperature for 40 min. Enhanced
chemiluminescence reagent (Pierce; Thermo Fisher Scientific, Inc.)
was used to detect the signal on the membrane. The data were
analyzed via densitometry using Image Lab™ version 3.0 software
(Bio-Rad Laboratories, Inc.) and normalized to expression of the
internal control (GAPDH).
MTT assay
CaSki and HeLa cells were harvested, seeded in
96-well plates (1×103 cells/ml per well) and grown until
80% confluence. MTT solution (5 mg/ml; Sigma-Aldrich; Merck KGaA)
was subsequently added to the cells (15 µl/well). Following 4 h of
incubation of the cells with MTT solution at 37°C, DMSO was added
(150 µl/well) to dissolve the formazan crystals. A microplate
reader (Omega Bio-Tek, Inc.) was used for spectrophotometry-based
measurements of the optical density at 490 nm.
Colony formation assay
CaSki and HeLa cells (7×102 cells/well)
were seeded in a 6-well plate and cultured for 14 days to allow the
formation of cell colonies. Following 14 days, the colonies were
fixed with 10% formaldehyde for 30 min at room temperature and
stained with 0.5% crystal violet for 5 min at room temperature. The
images of the colonies (>50 cells) were obtained using a camera
(Olympus Corporation) and counted manually.
Transwell assay
Transwell upper chambers (pore size, 8.0 µm; BD
Biosciences) were precoated with Matrigel. A total of 100 µl CaSki
and HeLa cell suspension (1×105 cells) in serum-free
medium was separately added into the upper chamber. The lower
chamber was filled with RPMI-1640 medium supplemented with 10% FBS.
Following 20 h of incubation, non-invasive cells in the upper
chamber were removed and cells that had invaded the lower chamber
were stained using Giemsa solution at room temperature for 15 min.
The number of invading cells was estimated using a light microscope
(magnification, ×100) and quantified using ImageJ version 1.8.0
software (National Institutes of Health). The relative invasion
rate was calculated as follows: Number of invasive cells/number of
inoculated cells ×100%. All procedures were repeated three
times.
Wound healing assay
HeLa and CaSki cells (1×106 cells/well)
were separately seeded in 6-well plates (Corning, Inc.) and
cultured until they reached 95% confluence. The cell monolayer was
slowly scratched with a sterile 200-µl tip. Cellular debris was
washed off with fresh medium and the remaining cells were cultured
in serum-free RPMI-1640 medium accordingly. The images of the wound
healing process were obtained at 0 and 24 h using a light
microscope (magnification, ×100). The area of wound was quantified
using ImageJ version 1.8.0 software (National Institutes of
Health). All procedures were repeated three times.
Bioinformatics analysis
The Gene Expression Profiling Interactive Analysis
(GEPIA; http://gepia.cancer-pku.cn/) database
is a website that provides the RNA sequencing expression data of
9,736 tumors and 8,587 normal samples from The Cancer Genome Atlas
(TCGA) and the Genotype-Tissue Expression (GTEx) projects (23). This database was used to determine
the expression difference of EphA2 between patients with cervical
squamous cell carcinoma and endocervical adenocarcinoma (CESC) and
healthy controls. A total of 306 tumor samples and 13 normal
samples were obtained from GEPIA, with |Log2FC|>1 and
P<0.01 as the cutoff; where FC is fold-change. The UALCAN
(http://ualcan.path.uab.edu/) database is
a portal for tumor subgroup gene expression and survival analyses,
which provides data from TCGA, Clinical Proteomic Tumor Analysis
Consortium (CPTAC) and The Children Brain Tumor Tissue
Consortium(CBTTC) (24). This
database was also used to determine the expression difference of
EphA2 in patients with cervical squamous cell carcinoma and healthy
controls. A total of 305 tumor samples and 3 normal samples were
obtained from UALCAN. The analysis results are presented in the
form of box plots.
Statistical analysis
The data are presented as the mean ± standard
deviation and were analyzed using GraphPad Prism version 8.0
(GraphPad Software, Inc.). One-way ANOVA followed by Tukey's post
hoc test was used for data comparisons between multiple groups.
P<0.05 was considered to indicate a statistically significant
difference. All experiments were performed at least three
times.
Results
EphA2 expression is increased in CC
cells
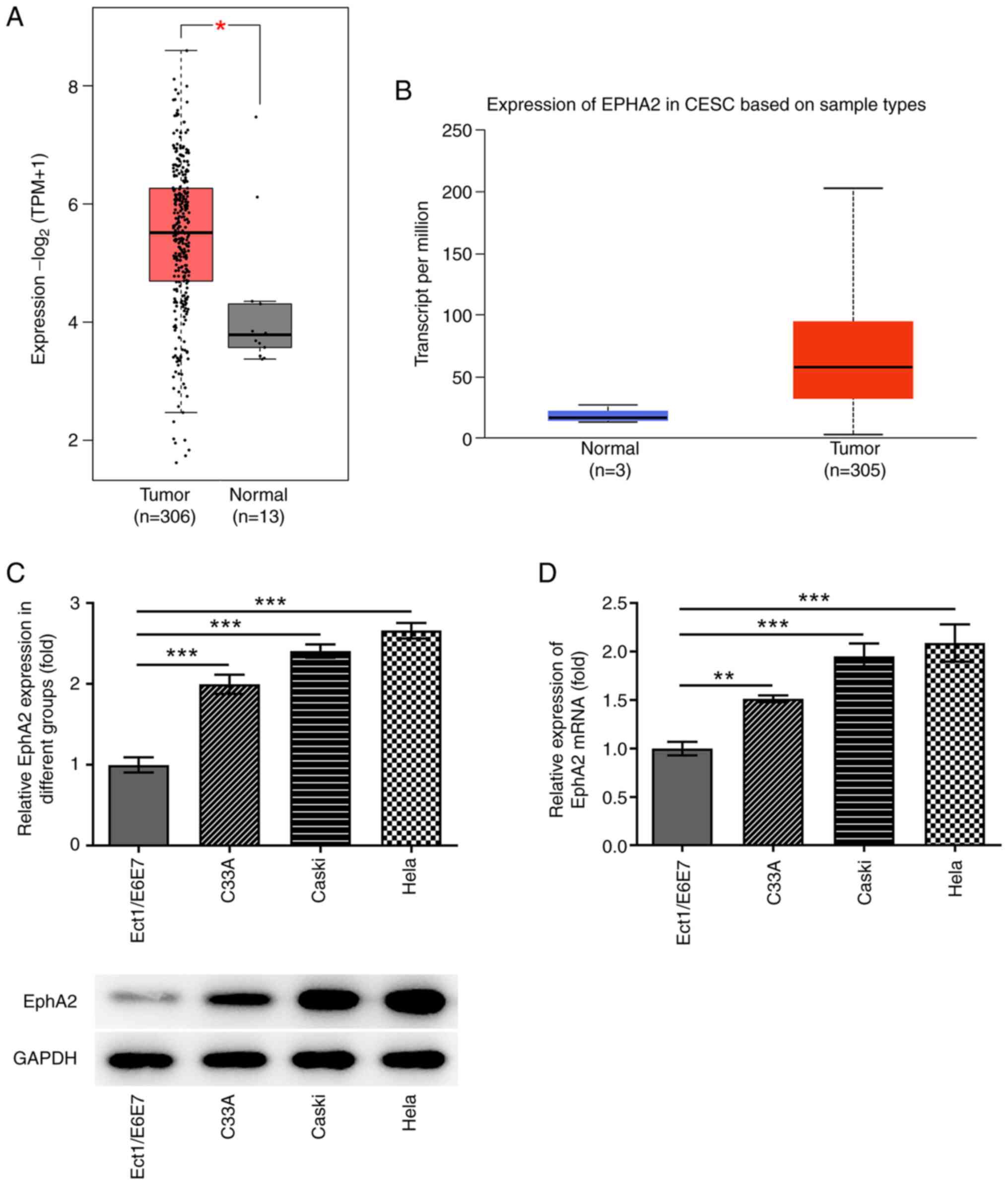
The result from GEPIA database analysis demonstrated
that the expression level of EphA2 in 306 patients with cervical
squamous cell carcinoma and endocervical adenocarcinoma was
significantly higher than that in 13 normal samples (Fig. 1A). The boxplot from the UALCAN
database of 305 patients and 3 normal samples also demonstrated the
higher expression of EphA2 in primary tumor samples compared with
that in normal samples (Fig. 1B).
Subsequently, RT-qPCR and western blotting were used to determine
EphA2 expression in HPV-positive CC cell lines (CaSki and HeLa), in
the HPV-negative CC cell line (C33A) and in immortalized human
cervical squamous cells (Ect1/E6E7). The results indicated that
EphA2 expression levels were significantly higher in C33A, CaSki
and HeLa cells than those noted in Ect1/E6E7 cells. The upward
trends in CaSki and HeLa cells were more obvious compared with that
in C33A cells (Fig. 1C and D).
Therefore, CaSki and HeLa cells were selected for the subsequent
assays.
ALW-II-41-27 inhibits the
proliferation of CC cells
To investigate the effects of ALW-II-41-27 on the
progression of CC, the proliferative capacity of CaSki and HeLa
cells was determined prior to and following treatment with
different concentrations of ALW-II-41-27. The results from an MTT
assay demonstrated that ALW-II-41-27 significantly decreased the
proliferative capacity of CaSki and HeLa cells, especially when
cells were treated for 48 and 72 h with 1,000 nM ALW-II-41-27
(Fig. 2A and B). In addition, the
number of colonies of both CaSki and HeLa cells was significantly
decreased following treatment with different concentrations of
ALW-II-41-27 (Fig. 2C and D). In
particular, the concentration of 1,000 nM exerted the most
prominent inhibitory effect on cell colony formation (Fig. 2C and D). These results demonstrated
that ALW-II-41-27 could inhibit CC cell proliferation.
ALW-II-41-27 inhibits the migration
and invasion of CC cells
CC cell migration and invasion were detected prior
to and following treatment with different concentrations of
ALW-II-41-27. The results from the wound-healing assay indicated
that ALW-II-41-27 significantly inhibited CaSki and HeLa cell
migration (Fig. 3A and C).
Furthermore, ALW-II-41-27 significantly inhibited CaSki and HeLa
cell invasive activity (Fig. 3B and
D). These findings also indicated a concentration-dependent
inhibition of CC cell progression by ALW-II-41-27, with 1,000 nM
ALW-II-41-27 exerting the most significant inhibitory effects.
These findings indicated the inhibitory effect of ALW-II-41-27 on
CC cell migration and invasion.
ALW-II-41-27 inhibits the RhoA/ROCK
pathway in CC cells
It has been reported that overexpression of
ROCK-I/II is associated with the progression of CC (19). A previous study confirmed that RhoA
can promote the proliferation and migration of CC cells (19). These results indicate that
ALW-II-41-27 suppresses the malignant activity of CC cells. A
potential regulatory association between ALW-II-41-27 and the
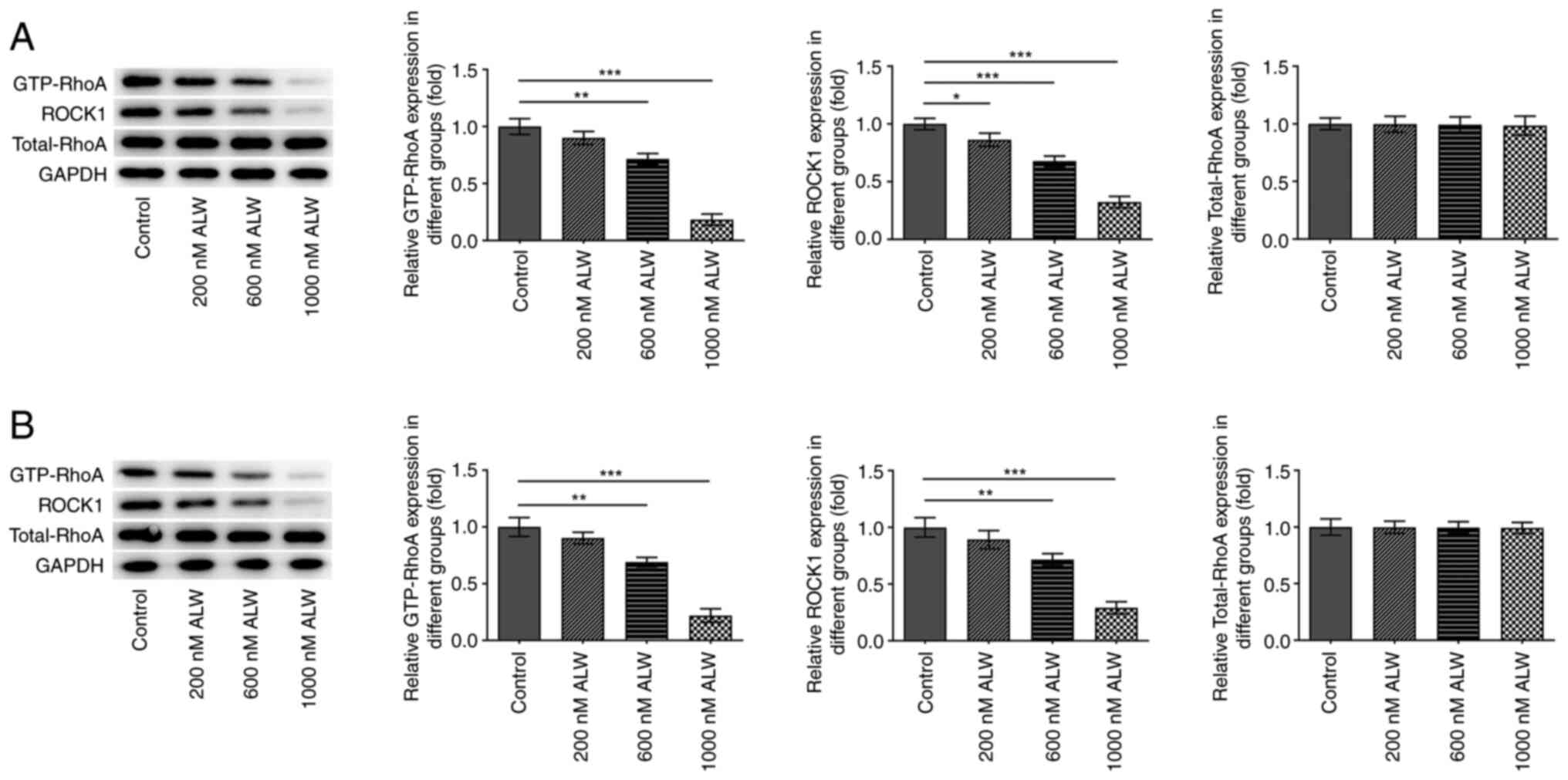
RhoA/ROCK pathway was therefore hypothesized. In the present study,
western blotting analysis was performed to determine the expression
of the proteins associated with the RhoA/ROCK pathway. The results
indicated that the protein expression of GTP-RhoA and ROCK1 was
significantly decreased by different concentrations of ALW-II-41-27
in CaSki and HeLa cells, whereas the total protein expression of
RhoA was unchanged (Fig. 4A and
B). In addition, ALW-II-41-27 at the concentration of 1,000 nM
caused the highest downregulations of GTP-RhoA and ROCK.
ALW-II-41-27 inhibits the
proliferation, invasion and migration of CC cells by blocking the
RhoA/ROCK pathway
To explore whether ALW-II-41-27 could suppress CC
cell progression via the RhoA/ROCK pathway, CaSki and HeLa cells
were co-treated with ALW-II-41-27 and PMA in the absence or
presence of C3 transferase or hydroxyfasudil. Since ALW-II-41-27
functioned in a concentration-dependent manner, the most efficient
concentration of 1,000 nM was selected for the following assays.
The results from western blotting indicated that GTP-RhoA and ROCK1
protein expression was decreased following treatment with
ALW-II-41-27 compared with that in the control group, while the
addition of PMA largely reversed this change (Fig. 5A and B). Co-treatment of the cells
with C3 transferase or hydroxyfasudil further decreased the
expression of GTP-RhoA and ROCK compared with the expression
following treatment with PMA + ALW-II-41-27, whereas that of
total-RhoA level remained the same (Fig. 5A and B).
 | Figure 5.ALW-II-41-27 inhibits the
proliferative, invasive and migratory activities of cervical cancer
cells by blocking the RhoA/ROCK pathway. Expression levels of
GTP-RhoA, ROCK1 and total-RhoA were determined by western blotting
and reverse transcription quantitative PCR analyses in (A) CaSki
and (B) HeLa cells treated with ALW-II-41-27, ALW-II-41-27 (1,000
nM) + PMA (100 ng/ml), ALW-II-41-27 + PMA + C3 transferase (5
µg/ml) or hydroxyfasudil (10 µM). The proliferation of (C) CaSki
and (D) HeLa cells was assessed using the MTT assay. The
proliferation of (E) CaSki and (F) HeLa cells was assessed using a
colony formation assay. *P<0.05, **P<0.01 and ***P<0.001.
RhoA, Ras homolog family member A; ROCK1, Rho-associated protein
kinase; PMA, phorbol 12-myristate 13-acetate; ALW,
ALW-II-41-27. |
The results from the MTT assay revealed that
ALW-II-41-27 significantly decreased CC cell proliferation compared
with the control and that PMA hindered its effect. Co-treatment
with PMA and C3 transferase or hydroxyfasudil significantly
decreased ALW-II-41-27 + PMA-promoted cell proliferation at 72 h.
CaSki and HeLa cells presented similar trends (Fig. 5C and D). The results of the colony
formation assay indicated that co-treatment with PMA and C3
transferase or hydroxyfasudil inhibited cell colony formation,
which was consistent with the results of the MTT assay (Fig. 5E and F).
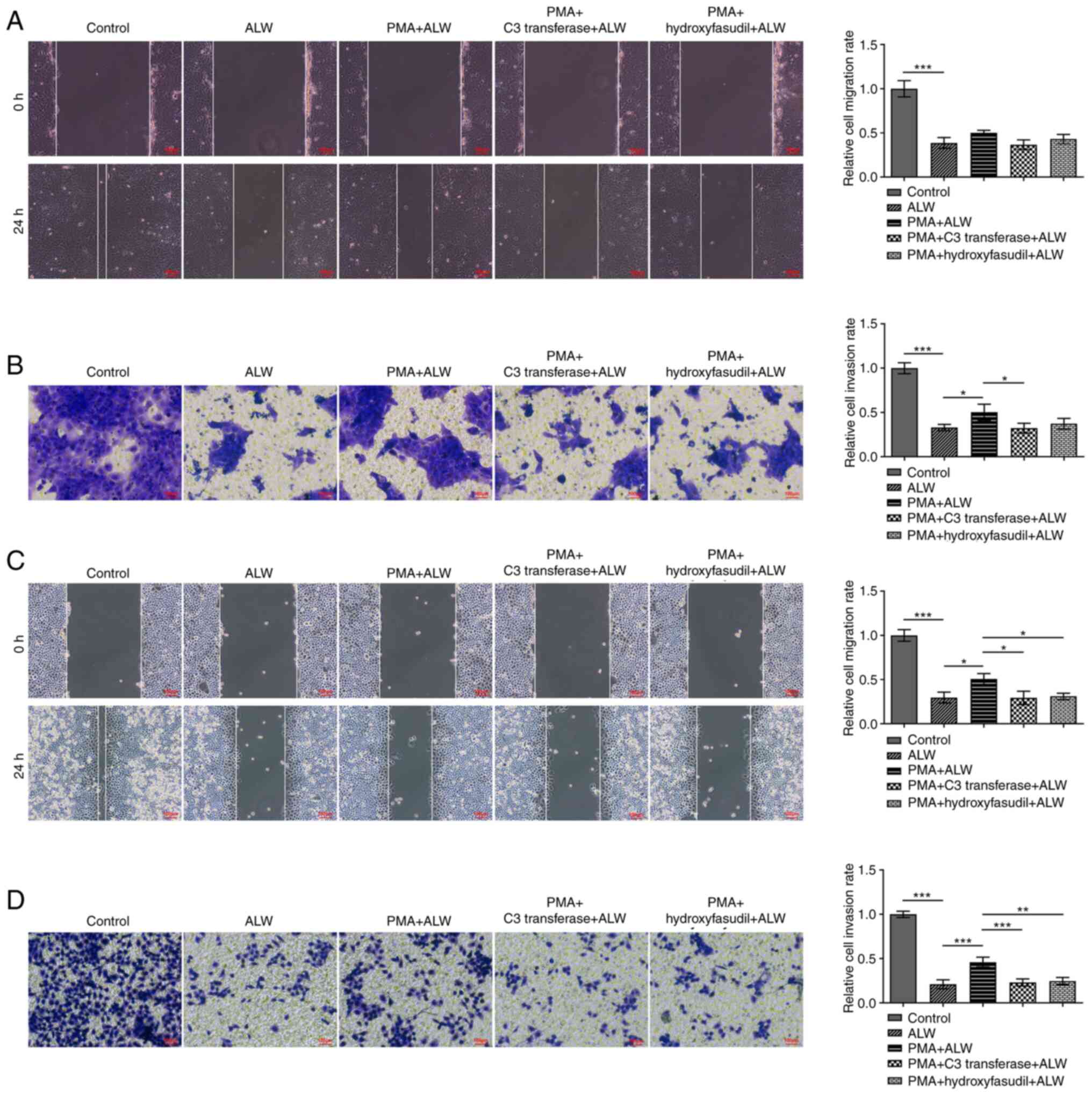
The results from the wound healing and Transwell
assays revealed that ALW-II-41-27-treated CaSki and HeLa cells
exhibited lower migratory and invasive activities compared with the
control, while activities were partially restored in the PMA and
ALW-II-41-27 co-treatment groups. The migratory activities of
ALW-II-41-27-treated CC cells detected by wound healing assays were
decreased following co-treatment with PMA and C3 transferase or
hydroxyfasudil (Fig. 6A and C).
The results of invasive activities from the Transwell assays
presented similar trends among the different groups (Fig. 6B and D). These findings indicated
that ALW-II-41-27 could suppress the malignant activity of CC cells
by blocking the RhoA/ROCK signaling pathway.
Discussion
CC ranks fourth among the most lethal gynecological
malignancies worldwide (25).
Despite the efforts made to improve the efficacy of standard
treatment for CC, including surgical excision assisted by
chemotherapy and radiotherapy (26,27),
the 5-year overall survival rate remains low in metastatic cases
(28). Therefore, in-depth
exploration and understanding of the pathogenesis and developmental
process of CC are crucial for the development of novel therapeutic
drugs and the search for novel treatment strategies.
EphA2 is expressed at high levels in several human
tumors, such as prostate, bladder, skin and lung tumors (29). Furthermore, cancer cells of the
highest tumor stage exhibit the highest expression levels of EphA2
(30). In addition, a previous
study demonstrated that EphA2 overexpression was closely associated
with a poor prognosis, a higher probability of metastasis and a
shorter overall survival time in Ewing sarcoma (31). Furthermore, EphA2 plays a novel
role in atherosclerosis, regulating both plaque inflammation and
progression to advanced atherosclerotic lesions (32). EphA2 might also serve as a novel
target for bladder cancer therapy (33). In addition, high expression of
EphA2 together with EphrinA-1 is negatively correlated with the
overall survival and prognosis of squamous cell CC (12). In the present study, EphA2
expression levels were assessed by western blotting and RT-qPCR
analyses in CC cell lines. A significant increase in EphA2
expression was demonstrated in both HPV-positive CC cell lines and
the HPV-negative CC cell line. Therefore, CaSki and HeLa cell lines
were selected for subsequent experiments in the present study. To
evaluate the function of EphA2 in the progression of CC cells, the
EphA2 inhibitor ALW-II-41-27 was used to treat the CaSki and HeLa
cell lines. The results from MTT and colony formation assays
indicated that ALW-II-41-27 decreased the proliferative activity of
the CaSki and HeLa cells in a concentration-dependent manner.
Subsequently, Transwell and wound-healing assays were performed.
The results demonstrated that ALW-II-41-27 could also inhibit the
migratory and invasive activities of CaSki and HeLa cells, and that
1,000 nM ALW-II-41-27 exhibited an optimal inhibitory effect.
Investigation of the molecular mechanism of
ALW-II-41-27 in inhibiting the growth and metastasis of CC was
focused on its potential to regulate the RhoA/ROCK pathway. A
previous study reported that the RhoA/ROCK pathway could mediate
cardiomyocyte apoptosis via induction of oxidative stress and
activation of the p38 MAPK and JNK pathways in vitro. These
experiments were performed in primary cardiomyocytes isolated from
ventricles of neonatal rats (34).
In addition, it has been demonstrated that Shroom family member 2
inhibits tumor metastasis via the RhoA/ROCK pathway in
nasopharyngeal carcinoma (35).
Targeting the RhoA/ROCK signaling pathway is also considered as an
effective method to suppress breast cancer progression and lung
cancer metastasis (36).
Furthermore, a previous study reported that vascular endothelial
growth factor-C facilitates CC metastasis via RhoA/ROCK-2-regulated
activation of the moesin protein (37). Quercetin can induce apoptosis and
loss of proliferation, migration and angiogenesis of SiHa CC cells
by downregulating the expression levels of specific
tumor-regulating genes, such as Akt1, Bcl-2-like 1, cyclin D1 gene,
cyclin-dependent kinase 4, Polo-like kinase 1 and RhoA (38). In the present study, the protein
expression of GTP-RhoA and ROCK1 was downregulated in a
concentration-dependent manner following ALW-II-41-27 treatment,
whereas the expression of total-RhoA did not change. Furthermore,
to investigate the involvement of the RhoA/ROCK pathway in the
effects of ALW-II-41-27 on CC cell progression, the RhoA/ROCK
pathway agonist PMA, the RhoA inhibitor C3 transferase and the ROCK
blocker hydroxyfasudil were used. Although PMA restored
ALW-II-41-27-mediated inhibition of CC cell proliferation,
migration and invasion, both C3 transferase and hydroxyfasudi could
reverse the changes caused by PMA treatment. These results
indicated that ALW-II-41-27 could inhibit CC cell proliferation,
migration and invasion by inhibiting the RhoA/ROCK pathway. It is
notable that the ROCK inhibitor Y-27632 has been shown to induce
upregulation of p63 expression in CC cells (39). This protein is considered to play a
fundamental role in cancer types arising from multi-layered
epithelium, for example, p63 participates in the malignant
phenotypes of esophageal squamous cell cancer cells (40,41).
In addition, a previous study demonstrated that ROCK kinase could
impede p63 nuclear transport in squamous cell carcinoma (42). The results from these two studies
are fairly different compared with those from the aforementioned
studies, as well as with the findings from the present study,
suggesting that additional investigation is required to determine
the exact underlying mechanism of ALW-II-41-27.
In summary, the present study demonstrated that
EphA2 was expressed at abnormally high levels in CC cells.
Downregulation of EphA2 expression levels was induced following CC
cell treatment with the inhibitor ALW-II-41-27, resulting in
decreased cell proliferation, migration and invasion. ALW-II-41-27
may therefore impact the aforementioned aspects of CC cells,
including proliferation, migration and invasion, by blocking the
RhoA/ROCK pathway. The present study involved basic in vitro
experiments. Additional in-depth mechanistic studies and in
vivo experiments are therefore required to confirm these
findings. The present study explored the association between EphA2
and CC, and identified the crucial role of EphA2 in disease
progression. Inhibition of EphA2 expression may thus be considered
as a promising therapeutic strategy for the treatment or the
control of CC. In addition, this study demonstrated that RhoA/ROCK
may be a key pathway in the regulation of EphA2 and should
therefore be examined further in the future to improve the
understanding of CC pathogenesis.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and DL contributed equally to this work and made
substantial contributions to conception and design, acquisition of
data and analysis and interpretation of data. RM made substantial
contributions to the conception and design of the study, and the
analysis and interpretation of data, and was involved in drafting
and critically revising the manuscript for important intellectual
content. XL, DL and RM confirm the authenticity of the data, and
have read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pimple SA and Mishra GA: Global strategies
for cervical cancer prevention and screening. Minerva Ginecol.
71:313–320. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bhatla N and Singhal S: Primary HPV
screening for cervical cancer. Best Pract Res Clin Obstet Gynaecol.
65:98–108. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stumbar SE, Stevens M and Feld Z: Cervical
cancer and its precursors: A preventative approach to screening,
diagnosis, and management. Prim Care. 46:117–134. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun Y, Yang X, Liu M and Tang H:
Corrigendum to ‘B4GALT3 up-regulation by miR-27a contributes to the
oncogenic activity in human cervical cancer cells.’ [Canc. Lett.
375 (2016) 284-292]. Cancer Lett. 493:16–18. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zelinski DP, Zantek ND, Stewart JC,
Irizarry AR and Kinch MS: EphA2 overexpression causes tumorigenesis
of mammary epithelial cells. Cancer Res. 61:2301–2306.
2001.PubMed/NCBI
|
|
8
|
Walker-Daniels J, Coffman K, Azimi M, Rhim
JS, Bostwick DG, Snyder P, Kerns BJ, Waters DJ and Kinch MS:
Overexpression of the EphA2 tyrosine kinase in prostate cancer.
Prostate. 41:275–280. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miyazaki T, Kato H, Fukuchi M, Nakajima M
and Kuwano H: EphA2 overexpression correlates with poor prognosis
in esophageal squamous cell carcinoma. Int J Cancer. 103:657–663.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kinch MS, Moore MB and Harpole DH Jr:
Predictive value of the EphA2 receptor tyrosine kinase in lung
cancer recurrence and survival. Clin Cancer Res. 9:613–618.
2003.PubMed/NCBI
|
|
11
|
Zeng G, Hu Z, Kinch MS, Pan CX, Flockhart
DA, Kao C, Gardner TA, Zhang S, Li L, Baldridge LA, et al:
High-level expression of EphA2 receptor tyrosine kinase in
prostatic intraepithelial neoplasia. Am J Pathol. 163:2271–2276.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu D, Suo Z, Kristensen GB, Li S, Troen G,
Holm R and Nesland JM: Prognostic value of EphA2 and EphrinA-1 in
squamous cell cervical carcinoma. Gynecol Oncol. 94:312–319. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng L, Li K, Wei H, Hu J, Jiao L, Yu S
and Xiong Y: A novel EphA2 inhibitor exerts beneficial effects in
PI–IBS in vivo and in vitro models via Nrf2 and NF-κB signaling
pathways. Front Pharmacol. 9:2722018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hong HN, Won YJ, Shim JH, Kim HJ, Han SH,
Kim BS and Kim HS: Cancer-associated fibroblasts promote gastric
tumorigenesis through EphA2 activation in a ligand-independent
manner. J Cancer Res Clin Oncol. 144:1649–1663. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ishigaki H, Minami T, Morimura O, Kitai H,
Horio D, Koda Y, Fujimoto E, Negi Y, Nakajima Y, Niki M, et al:
EphA2 inhibition suppresses proliferation of small-cell lung cancer
cells through inducing cell cycle arrest. Biochem Biophys Res
Commun. 519:846–853. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feng G, Sun B, Liu HX, Liu QH, Zhao L and
Wang TL: EphA2 antagonism alleviates LPS-induced acute lung injury
via Nrf2/HO-1, TLR4/MyD88 and RhoA/ROCK pathways. Int
Immunopharmacol. 72:176–185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan B, Cui J, Wang W and Deng K: Gα12/13
signaling promotes cervical cancer invasion through the
RhoA/ROCK-JNK signaling axis. Biochem Biophys Res Commun.
473:1240–1246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hou C, Zhuang Z, Deng X, Xu Y, Zhang P and
Zhu L: Knockdown of Trio by CRISPR/Cas9 suppresses migration and
invasion of cervical cancer cells. Oncol Rep. 39:795–801.
2018.PubMed/NCBI
|
|
19
|
Liu X, Chen D and Liu G: Overexpression of
RhoA promotes the proliferation and migration of cervical cancer
cells. Biosci Biotechnol Biochem. 78:1895–1901. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng Q, Chen L, Wu W, Wang J, Zheng X,
Chen Z, Jiang Q, Han J, Wei L, Wang L, et al: EPH receptor A2
governs a feedback loop that activates Wnt/β-catenin signaling in
gastric cancer. Cell Death Dis. 9:11462018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu XP, He HL, Hu SL, Han JB, Huang LL, Xu
JY, Xie JF, Liu AR, Yang Y and Qiu HB: Ang II-AT2R increases
mesenchymal stem cell migration by signaling through the FAK and
RhoA/Cdc42 pathways in vitro. Stem Cell Res Ther. 8:1642017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res.
45(W1):W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cancer Genome Atlas Research Network,
Albert Einstein College of Medicine; Analytical Biological
Services; Barretos Cancer Hospital; Baylor College of Medicine;
Beckman Research Institute of City of Hope; Buck Institute for
Research on Aging; Canada's Michael Smith Genome Sciences Centre;
Harvard Medical School, . Helen F, et al: Integrated genomic and
molecular characterization of cervical cancer. Nature. 543:378–384.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yee GP, de Souza P and Khachigian LM:
Current and potential treatments for cervical cancer. Curr Cancer
Drug Targets. 13:205–220. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Olusola P, Banerjee HN, Philley JV and
Dasgupta S: Human papilloma virus-associated cervical cancer and
health disparities. Cells. 8:6222019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goodman A: HPV testing as a screen for
cervical cancer. BMJ. 350:h23722015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tandon M, Vemula SV and Mittal SK:
Emerging strategies for EphA2 receptor targeting for cancer
therapeutics. Expert Opin Ther Targets. 15:31–51. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wykosky J and Debinski W: The EphA2
receptor and ephrinA1 ligand in solid tumors: Function and
therapeutic targeting. Mol Cancer Res. 6:1795–1806. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garcia-Monclus S, Lopez-Alemany R,
Almacellas-Rabaiget O, Herrero-Martín D, Huertas-Martinez J,
Lagares-Tena L, Alba-Pavón P, Hontecillas-Prieto L, Mora J, de
Álava E, et al: EphA2 receptor is a key player in the metastatic
onset of Ewing sarcoma. Int J Cancer. 143:1188–1201. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Finney AC, Funk SD, Green JM, Yurdagul A
Jr, Rana MA, Pistorius R, Henry M, Yurochko A, Pattillo CB, Traylor
JG, et al: EphA2 expression regulates inflammation and
fibroproliferative remodeling in atherosclerosis. Circulation.
136:566–582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abraham S, Knapp DW, Cheng L, Snyder PW,
Mittal SK, Bangari DS, Kinch M, Wu L, Dhariwal J and Mohammed SI:
Expression of EphA2 and Ephrin A-1 in carcinoma of the urinary
bladder. Clin Cancer Res. 12:353–360. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou H, Sun Y, Zhang L, Kang W, Li N and
Li Y: The RhoA/ROCK pathway mediates high glucose-induced
cardiomyocyte apoptosis via oxidative stress, JNK, and p38MAPK
pathways. Diabetes Metab Res Rev. 34:e30222018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yuan J, Chen L, Xiao J, Qi XK, Zhang J, Li
X, Wang Z, Lian YF, Xiang T, Zhang Y, et al: SHROOM2 inhibits tumor
metastasis through RhoA-ROCK pathway-dependent and -independent
mechanisms in nasopharyngeal carcinoma. Cell Death Dis. 10:582019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li D, Wang H, Ding Y, Zhang Z, Zheng Z,
Dong J, Kim H, Meng X, Zhou Q, Zhou J, et al: Targeting the
NRF-2/RHOA/ROCK signaling pathway with a novel aziridonin, YD0514,
to suppress breast cancer progression and lung metastasis. Cancer
Lett. 424:97–108. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He M, Cheng Y, Li W, Liu Q, Liu J, Huang J
and Fu X: Vascular endothelial growth factor C promotes cervical
cancer metastasis via up-regulation and activation of
RhoA/ROCK-2/moesin cascade. BMC Cancer. 10:1702010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Khumkhrong P, Piboonprai K, Chaichompoo W,
Pimtong W, Khongkow M, Namdee K, Jantimaporn A, Japrung D,
Asawapirom U, Suksamrarn A and Iempridee T: Crinamine induces
apoptosis and inhibits proliferation, migration, and angiogenesis
in cervical cancer SiHa cells. Biomolecules. 9:4942019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suprynowicz FA, Upadhyay G, Krawczyk E,
Kramer SC, Hebert JD, Liu X, Yuan H, Cheluvaraju C, Clapp PW,
Boucher RC Jr, et al: Conditionally reprogrammed cells represent a
stem-like state of adult epithelial cells. Proc Natl Acad Sci USA.
109:20035–20040. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Masuda Y, Takahashi H and Hatakeyama S:
TRIM29 regulates the p63-mediated pathway in cervical cancer cells.
Biochim Biophys Acta. 1853:2296–2305. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hazawa M, Lin DC, Handral H, Xu L, Chen Y,
Jiang YY, Mayakonda A, Ding LW, Meng X, Sharma A, et al: ZNF750 is
a lineage-specific tumour suppressor in squamous cell carcinoma.
Oncogene. 36:2243–2254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hazawa M, Lin DC, Kobayashi A, Jiang YY,
Xu L, Dewi FRP, Mohamed MS, Hartono .Nakada M, Meguro-Horike M, et
al: ROCK-dependent phosphorylation of NUP62 regulates p63 nuclear
transport and squamous cell carcinoma proliferation. EMBO Rep.
19:73–88. 2018. View Article : Google Scholar : PubMed/NCBI
|