Introduction
Renal cell carcinoma (RCC) is a common malignant
tumor in the human genitourinary system that includes numerous
different pathological subtypes (1,2). The
most common subtype is clear cell renal cell carcinoma (ccRCC).
ccRCC originates from renal tubular epithelial cells and accounts
for ~60-85% of RCCs (3,4). The prevalence of ccRCC in men is
higher than that in women, and most patients are over 60 years old.
It is often asymptomatic in the early stage or only has vague
systemic symptoms such as fever and fatigue. The typical clinical
symptoms are hematuria, pain and a palpable mass in the kidney
area. It affects either kidney at an equal rate. At present, the
treatment methods are limited, mainly radical nephrectomy, but
relapse and metastasis easily occur after surgery and the fatality
rate is high (5). The etiology of
ccRCC remains unclear and possible related factors include
genetics, smoking, obesity, hypertension and antihypertensive drug
therapy (6). Therefore, it is very
important to explore the potential biomarkers and therapeutic
targets of ccRCC.
The E2F family encodes extremely important nuclear
transcription factors involved in regulating the cell cycle
(7,8). It was first discovered by Kovesdi
et al (9) in 1986 during
studies of adenovirus. There are numerous members of the E2F
family. The ones that have been discovered include E2F1 to E2F8.
According to the protein structure and function and transcription
characteristics, they are divided into transcription promotion
factors (E2F1 to E2F3) and transcription suppressors (E2F4 to E2F8)
(10). Clinical studies have found
that E2F family proteins are closely related to the occurrence,
development, proliferation and apoptosis of gastric, lung, liver,
esophageal, prostate, bladder and ovarian cancer and other
malignant tumors (7,11). In addition, E2F family proteins
exhibit complex and diverse biological functions in different
tumors and their expression levels are not consistent in different
tumors (11). However, the
expression pattern of E2F family proteins in ccRCC and their
relationship with the prognosis remains unclear.
To the best of our knowledge, bioinformatics
analysis has yet to be applied to explore the role of E2F family in
ccRCC. In the present study, RNA sequencing (RNA-Seq) data from The
Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO) were
downloaded to explore the expression characteristics of E2F family
proteins in ccRCC and their relationship with prognosis. Finally,
the results of bioinformatics analysis we reverified with clinical
samples from surgical operations. The present study may provide a
new understanding of E2F family proteins in ccRCC and help to
interpret the mechanisms underlying their functions.
Materials and methods
Raw data
RNA-Seq and clinical data (from 530 tumor tissues
and 72 normal tissues) of ccRCC from TCGA were downloaded from
https://portal.gdc.cancer.gov/. The
RNA-Seq data were reported as fragments per kilobase million. The
datasets GSE36895 (12)
(containing 72 tumor tissues and 72 normal tissues) and GSE53757
(13) (containing 29 tumor tissues
and 23 normal tissues) were downloaded from GEO (https://www.ncbi.nlm.nih.gov/geo).
Reverse transcription-quantitative
(RT-q) PCR
A total of 10 groups of tumor tissues and
paired-normal tissues were obtained from 10 patients who underwent
radical resection at The Affiliated Suqian First People's Hospital
of Nanjing Medical University between January 2020 and August 2021.
The patients were diagnosed as ccRCC by imaging and pathological
examination, and did not receive chemotherapy or radiotherapy
before operation. All experimental procedures were approved
(approval no. 2018-SL-0026) by the Ethics Committee of The
Affiliated Suqian First People's Hospital of Nanjing Medical
University (Suqian, China). Written informed consent was provided
by all patients prior to the study. All patients (age range, 55–68
years; median age, 62 years; seven men and three women) were
diagnosed with ccRCC by laboratory examination and imaging
examination. The clinical information of the patients is provided
in Table SI. The mRNA expression
of the E2Fs was examined using RT-qPCR. Total RNA was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) from the tissues. Total RNA was converted into
cDNA, and quantitated using a Fastking One Step Reverse
Transcription and Fluorescence Quantitative Kit (Tiangen Biotech
Co., Ltd.) as a fluorophore according to the manufacturer's
protocol. The primers were designed by online tool ‘primerBank’
(https://pga.mgh.harvard.edu/primerbank). β-actin was
used as the internal reference gene. The primer sequences of E2Fs
are presented in Table SII.
Expression levels of mRNAs relative to β-actin were determined
using the 2-ΔΔCq method (14).
Definitions of clinical survival and
recurrence types
Raw counts of RNA-sequencing data (level 3) of ccRCC
from TCGA were downloaded from https://portal.gdc.cancer.gov/. A total of 3 types of
clinical survival and recurrence outcomes were selected in the
present study: overall survival (OS), disease-specific survival
(DSS), progression-free survival (PFS) and disease-free survival
(DFS) (15). Survival analysis
with the log-rank test was used to compare the survival difference
between the normal and tumor groups. The hazard ratio (HR) was used
to indicate the risk difference between the two groups.
Mutation and methylation analysis
The data were downloaded from TCGA (https://portal.gdc.cancer.gov/) and cBioPortal
(https://www.cbioportal.org/datasets),
copy number and mutation analysis were performed using cBioPortal
(https://www.cbioportal.org/) (15). MeV software (http://projects/mev-tm4) was used to create heatmaps.
Pearson's correlation analysis and mapping were performed using R
4.1.1 (https://www.r-project.org/). P<0.05
indicates a significant correlation.
Enrichment analysis
Gene enrichment analysis was performed to determine
the correlation between E2F expression and the cell cycle. The data
were downloaded from TCGA. Gene set enrichment analysis (https://www.gsea-msigdb.org/gsea) was used to
perform gene enrichment analysis in three datasets, REACTOME,
(Kyoto Encyclopedia of Genes and genomes (KEGG) and Gene Ontology
(GO). A normalized enrichment score (NES)>0 means E2F expression
is positively correlated with the cell cycle, and a false discovery
rate (FDR) <0.05 was considered to indicate a statistically
significant difference.
Statistical analysis
The Mann-Whitney U test was performed using GraphPad
Prism 9 software (GraphPad Software, Inc.) and R 4.1.1 was used to
determine E2F expression between normal tissues and tumor tissues.
Kaplan-Meier survival curves and log-rank tests were used to
evaluate the effect of E2F expression on survival. The
Kruskal-Wallis test was used to detect the differences between
groups in normal tissue + stage/grade and stage/grade. P<0.05
was considered to indicate a statistically significant
difference.
Results
E2Fs expression in patients with
ccRCC
RNA-Seq data from 530 tumors and 72 normal tissue
samples from the TCGA dataset were analyzed (Fig. 1A). In data of TCGA, compared with
normal tissues, E2F1, E2F2, E2F3, E2F4, E2F6, E2F7 and E2F8 were
overexpressed in cancer tissues, and E2F5 was expressed at low
levels in cancer tissues (Fig.
1B). In dataset GSE36985, the expression levels of E2F1, E2F2,
E2F6, E2F7 and E2F8 in tumor tissues were higher than those in
normal tissues, and the expression of E2F5 was lower than that in
normal tissues. There was no statistically significant difference
between the two groups of E2F3 and E2F4 (Fig. 1C). In dataset GSE53757, the
expression of E2F2, E2F6, E2F7 and E2F8 in tumor tissues was higher
than that in normal tissues, and the expression of E2F4 and E2F5
was lower than that in normal tissues. There was no statistically
significant difference between the two groups of E2F1 and E2F3
(Fig. 1D). In addition, the
RT-qPCR verification results of clinical specimens collected from
ccRCC surgery revealed that compared with normal tissues, E2F5
expression in cancer tissues was low, and the E2F1, E2F, E2F4,
E2F6, E2F7 and E2F8 were overexpressed (Fig. 1E).
E2F family expression and pathological
status of patients with ccRCC
The relationship of E2F family expression with stage
and grade was analyzed, respectively in TCGA data. The expression
of E2Fs is significantly different in the staging and grading of
patients with ccRCC. The higher the pathological stage was, the
higher the expression of E2F1, E2F2, E2F3, E2F4, E2F6, E2F7 and
E2F8 and the lower the expression of E2F5 (Fig. 2A). The results of pathological
grading were similar to the results of staging (Fig. 2B).
Prognostic value of E2Fs in patients
with ccRCC
To evaluate the clinical significance of the
expression of E2Fs in the survival of patients with ccRCC, E2F
expression was assessed in 530 ccRCC clinical samples (≤4,000 days
of follow-up) from TCGA for OS, DSS and PFS. Low expression of
E2F1, E2F2, E2F3 E2F4, E2F5, E2F7 and E2F8 were significantly
associated with a longer OS time (HR=1.989, P<0.0001; HR=2.111,
P<0.0001; HR=1.779, P=0.000; HR=1.941, P<0.0001; HR=2.194,
P<0.0001; HR=2.656, P<0.0001; HR=1.616, P=0.001), a longer
DSS time (HR=3.042, P<0.0001; HR=3.121, P<0.0001; HR=2.058,
P=0.005; HR=2.433, P<0.0001; HR=2.355, P<0.0001; HR=3.746,
P<0.0001; HR=2.537, P<0.0001) and a longer PFS time
(HR=2.030, P<0.0001; HR=1.976, P<0.0001; HR=1.406, P=0.041;
HR=1.621, P=0.004; HR=1.477, P=0.033; HR=2.579, P<0.0001;
HR=1.908, P<0.0001) in patients with ccRCC, respectively
(Fig. 3). However, low expression
of E2F6 was not associated with OS (HR=1.319; P=0.068), DSS
(HR=1.314; P=0.156) or PFS (HR=0.717; P=0.077) in patients with
ccRCC (Fig. 3). However, it was
found that the expression of E2F1-E2F8 was not significantly
associated with DFS. These results suggested that the mRNA
expression levels of E2F1, E2F2, E2F3 E2F4, E2F5, E2F7 and E2F8 may
be useful for the prediction of survival of patients with
ccRCC.
Mechanism of E2Fs imbalance in
patients with ccRCC
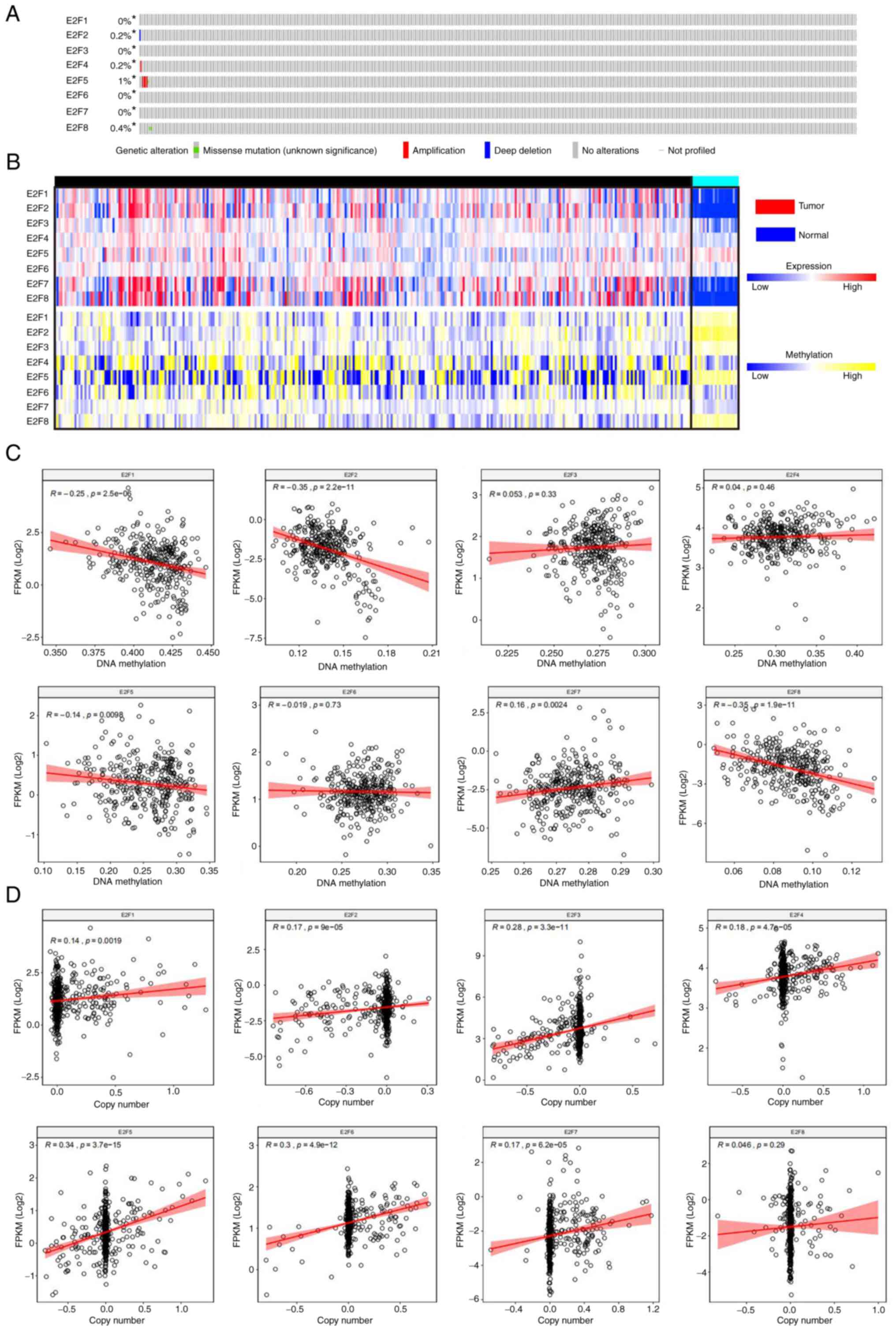
The mutation rates of E2F1-E2F8 were 0, 0.2, 0, 0.2,
1, 0, 0 and 0.4%, respectively (Fig.
4A). The expression of E2F1, E2F2, E2F5 and E2F8 was
significantly negatively correlated with DNA methylation (r=−0.25,
P<0.0001; r=−0.35, P<0.0001; r=−0.14, P<0.0001; r=−0.35,
P<0.0001, respectively), and the expression of E2F7 was
significantly positively correlated with DNA methylation (r=−0.16,
P=0.0024). The expression of E2F3, E2F4 and E2F6 was not
significantly correlated with DNA methylation (r=0.053, P=0.033;
r=0.04, P=0.46; r=−0.019, P=0.73) (Fig. 4B and C). The expression of
E2F1-E2F7 was significantly positively correlated with copy number
(r=0.14, P=0.002; r=0.17, P<0.0001; r=0.28, P<0.0001; r=0.18,
P<0.0001; r=0.34, P<0.0001; r=0.30, P<0.0001; r=0.17,
P<0.0001, respectively), and the expression of E2F8 was not
significantly correlated with copy number (r=0.046, P=0.29)
(Fig. 4B and D). The results
revealed that the mutation rate of E2Fs in patients with ccRCC is
low. E2F1, E2F2 and E2F8 expression may be increased due to
hypomethylation in tumors, and E2F4, E2F6 and E2F7 expression may
be increased due to an increased copy number in tumors.
E2Fs expression with cell cycle
The results demonstrated that the expression of
E2F1, E2F3, E2F6, E2F7 and E2F8 in REACTOME was significantly
positively correlated with the cell cycle (NES=2.22, FDR=0.004;
NES=1.81, FDR=0.01; NES=1.99, FDR<0.001; NES=2.24, FDR=0.001;
NES=2.06, FDR=0.008) (Fig. 5). The
expression of E2F1, E2F2, E2F3, E2F6, E2F7 and E2F8 in KEGG was
significantly positively correlated with the cell cycle (NES=2.08,
FDR=0.006; NES=1.73, FDR=0.04; NES=1.96, FDR=0.01; NES=2.09,
FDR=0.00; NES=2.18, FDR<0.0001; NES=2.10, FDR=0.009) (Fig. 5). The expression of E2F1, E2F2,
E2F3, E2F4, E2F6, E2F7 and E2F8 in GO was significantly positively
correlated with the cell cycle (NES=1.91, FDR=0.01; NES=1.68,
FDR=0.04; NES=1.85, FDR=0.01; NES=1.77, FDR=0.02; NES=2.18,
FDR=0.00; NES=2.16, FDR=0.001; NES=1.85, FDR=0.01) (Fig. 5).
Discussion
Studies have shown that targeted therapy can
significantly improve the survival rate of patients with metastatic
ccRCC (16). However, ccRCC
currently has fewer therapeutic targets in clinical practice, and
it is necessary to identify and explore more therapeutic targets to
provide a reference for clinical treatment.
E2F is a group of genes encoding transcription
factors in higher eukaryotes. According to their different
functions, they can be divided into transcription activators and
transcription suppressors. To date, 8 E2F family protein members
have been identified in mammals, namely, E2F1, E2F2, E2F3, E2F4,
E2F5, E2F6, E2F7 and E2F8 (7,11,17).
Among them, E2F1 is the only transcription factor among the eight
protein family members that has the dual functions of mediating
apoptosis and regulating cell proliferation. E2F1 and E2F3a are
transcriptional activators in the E2F family of proteins that bind
to target genes and participate in the regulation of the cell
cycle. E2F4 can participate in the regulation of normal cell
proliferation, differentiation, apoptosis and other physiological
processes. E2F3 to E2F5 can act as transcription inhibitors after
binding to pocket proteins related to retinoblastoma proteins
(11). E2F6 inhibits DNA
damage-induced apoptosis (8). E2F7
and E2F8 are new members of the E2F family of proteins discovered
in recent years. They are atypical family factors and can work in
concert with hypoxia-inducible factors to jointly regulate the
transcription of vascular endothelial growth factor. E2Fs have been
reported to have roles in a variety of cancer types, such as breast
(18), liver (19) and gastric cancer (20), since they can regulate numerous
cellular functions related to cell cycle progression. Although
certain E2F family members have been confirmed to play promising
roles in ccRCC, the distinct roles of E2Fs in the development,
progression and metastasis of ccRCC remain to be elucidated. In the
present study, the expression, mutation and prognostic values of
different E2Fs in patients with ccRCC were analyzed.
E2F1 is the most frequently studied gene in the E2F
family. E2F1 is an important transcription-promoting factor located
on human chromosome 20q11, ~11 kb in size, mainly comprised of six
introns and seven exons, and it can encode proteins with a size of
more than 400 amino acids (10,21).
E2F1 has a very obvious tissue specificity. It can form a
heterodimer with retinoblastoma and bind to the corresponding DNA
sequence, thereby enhancing or inhibiting the activity of E2F1. The
transcription factor E2F1 can regulate biological processes such as
the cell cycle, cell proliferation, cell apoptosis and cell
differentiation (10). In the
present study, E2F1 was highly expressed in cancer tissues and its
high expression was closely related to a worse tumor grade and
staging and a poor prognosis. In ccRCC, the expression of E2F1 is
also significantly positively correlated with the cell cycle.
E2F2 and E2F3 are involved in regulation of the cell
cycle and are highly expressed in breast cancer tissues (18). At present, little is known about
the expression and role of E2F2 in ccRCC. In the present study, it
was revealed that the expression of E2F2 in ccRCC tissues is higher
than that in normal tissues, and this expression is significantly
related to the tumor stage and grade of patients with ccRCC. In
addition, in all patients with ccRCC, high expression of E2F2 was
significantly associated with a poor OS, DSS and PFS, which
appeared to be consistent with the role of E2F2 as an oncogene.
Notably, high expression of E2F3 in cancer tissues was observed in
TCGA, but this phenomenon was not identified in the two GEO
datasets or the clinical sample validation.
The present study showed that E2F4 may have
carcinogenic effects and the high expression of E2F4 is related to
poor prognostic factors of ccRCC, such as high TNM stage and high
grade. Furthermore, it has been reported that the expression of
E2F4 in prostate (22) and breast
cancer (18) is higher than that
in normal tissues. Although E2F4 is traditionally classified as a
cell cycle inhibitor, its pro-proliferation and anti-apoptotic
activities have been confirmed in various human cell lines.
Notably, gene enrichment analysis in the present study revealed
that the positive correlation between E2F4 and the cell cycle was
not significant, and the possible relationship of E2F4 with the
early stages of carcinogenesis needs to be clarified.
E2F5 has different expression levels in different
types of tumors. Studies have reported that E2F5 is overexpressed
in glioblastoma (23) and prostate
cancer (24). However, E2F5 was
downregulated in MCF7 human breast cancer cells, significantly
impairing cell proliferation, migration and invasion in
vitro and increasing cell cycle arrest in the G0/G1 phase
(25). In the present study, it
was proved that the expression of E2F5 in ccRCC tissues is lower
than that in normal tissues. According to previous studies, the
expression of E2F6 in breast cancer tissues is lower than that in
normal tissues, but this expression was not correlated with tumor
stage in patients with breast cancer (18,26).
In the present study, it was identified that the expression of E2F6
in ccRCC tissues was higher than that in normal tissues, and the
expression level was significantly positively correlated with the
copy number but not significantly correlated with the prognosis of
patients with ccRCC. Studies have found that E2F7 and E2F8 are
unique inhibitory genes that have a vital inhibitory effect on cell
proliferation (27–29). However, the present study
demonstrated that E2F7 and E2F8 are highly expressed in ccRCC
tissues, suggesting that they may play different biological roles
in different cell types.
The change of DNA methylation status is an important
factor in tumorigenesis. This change includes the decrease of the
overall methylation level of the genome and the abnormal increase
of the local methylation level of CpG island, resulting in the
instability of the genome and the low expression of tumor
suppressor genes (30). It was
revealed that E2F1, E2F2 and E2F8 expression may be increased due
to hypomethylation in tumors, and E2F4, E2F6 and E2F7 expression
may be increased due to an increased copy number in tumors. In
addition, the present study showed that the expression of E2F
family in ccRCC was significantly positively correlated with cell
cycle.
In conclusion, except for the E2F3 and E2F5, all E2F
family members are highly expressed in ccRCC tissues, and their
expression is closely related to the pathological status and
survival prognosis of patients with ccRCC, but different members of
the family have differences in different tissue samples. The
present findings suggested that E2F family genes may be potential
targets for molecular diagnosis and targeted therapy of ccRCC,
transcriptional E2F1-5, 7, and 8 were potential prognostic markers
for the improvement of ccRCC survival and prognostic accuracy.
Targeted therapy against single or combined E2F family proteins may
improve the therapeutic effect and patient outcomes. It is expected
that the findings of the present study will contribute to available
knowledge, improve treatment designs, and enhance the accuracy of
prognosis for patients with ccRCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Medical Research Project
of Jiangsu Provincial Health Commission (grant no. Z2020074), the
Six Talent Peaks Project in Jiangsu (grant no. WSN-342), the
Jiangsu Provincial Medical Youth Talent (grant no. QNRC2016480),
Suqian Guiding Science and Technology Project (grant no. Z2018174)
and the Science and Technology Project of Suqian (grant no.
S201720).
Availability of data and materials
The datasets generated and analyzed during the
current study are available in TCGA (portal.gdc.cancer.gov) and GEO
(ncbi.nlm.nih.gov/geo/).
Authors' contributions
ZL, YS and HG conceptualized and designed the
research and performed bioinformatics analysis. YW and XYZ
performed the experiments. JS, XY, XCZ, HL and XJY analyzed and
interpreted the data. ZL and JS drafted and edited the manuscript.
XY and XCZ supervised the project. ZL, YS and HG confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved (approval no.
2018-SL-0026) by the Ethics Committee of The Affiliated Suqian
First People's Hospital of Nanjing Medical University (Suqian,
China). Written informed consent was provided by all patients prior
to the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gray RE and Harris GT: Renal cell
carcinoma: Diagnosis and management. Am Fam Physician. 99:179–184.
2019.PubMed/NCBI
|
|
2
|
Linehan WM and Ricketts CJ: The cancer
genome atlas of renal cell carcinoma: Findings and clinical
implications. Nat Rev Urol. 16:539–552. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Makhov P, Joshi S, Ghatalia P, Kutikov A,
Uzzo RG and Kolenko VM: Resistance to systemic therapies in clear
cell renal cell carcinoma: Mechanisms and management strategies.
Mol Cancer Ther. 17:1355–1364. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lucarelli G, Loizzo D, Franzin R,
Battaglia S, Ferro M, Cantiello F, Castellano G, Bettocchi C,
Ditonno P and Battaglia M: Metabolomic insights into
pathophysiological mechanisms and biomarker discovery in clear cell
renal cell carcinoma. Expert Rev Mol Diagn. 19:397–407. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tegos T, Tegos K, Dimitriadou A and
Dimitriadis G: Current and emerging first-line systemic therapies
in metastatic clear-cell renal cell carcinoma. J BUON.
24:1340–1353. 2019.PubMed/NCBI
|
|
6
|
Schodel J, Grampp S, Maher ER, Moch H,
Ratcliffe PJ, Russo P and Mole DR: Hypoxia, hypoxia-inducible
transcription factors, and renal cancer. Eur Urol. 69:646–657.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang H, Wang X, Xu L, Zhang J and Cao H:
Integrated analysis of the E2F transcription factors across cancer
types. Oncol Rep. 43:1133–1146. 2020.PubMed/NCBI
|
|
8
|
Pennycook BR, Vesela E, Peripolli S, Singh
T, Barr AR, Bertoli C and de Bruin RAM: E2F-dependent transcription
determines replication capacity and S phase length. Nat Commun.
11:35032020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kovesdi I, Reichel R and Nevins JR: Role
of an adenovirus E2 promoter binding factor in E1A-mediated
coordinate gene control. Proc Natl Acad Sci USA. 84:2180–2184.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ertosun MG, Hapil FZ and Nidai OO: E2F1
transcription factor and its impact on growth factor and cytokine
signaling. Cytokine Growth Factor Rev. 31:17–25. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kent LN and Leone G: The broken cycle: E2F
dysfunction in cancer. Nat Rev Cancer. 19:326–338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng L, Dou X, Song H, Gao R and Tang X:
TRPV1 acts as a tumor suppressor and is associated with immune cell
infiltration in clear cell renal cell carcinoma: Evidence from
integrated analysis. J Cancer. 11:5678–5688. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang W, Zhang M, Wang Q, Gu D, Huang Z,
Wang H, Xiang Y, Xia Q, Cui Z and Jin X: The SLC family are
candidate diagnostic and prognostic biomarkers in clear cell renal
cell carcinoma. Biomed Res Int. 2020:19329482020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng Y, Fang YC and Li J: PD-L1
expression levels on tumor cells affect their immunosuppressive
activity. Oncol Lett. 18:5399–5407. 2019.PubMed/NCBI
|
|
15
|
Wang H, Yan C and Ye H: Overexpression of
MUC16 predicts favourable prognosis in MUC16-mutant cervical cancer
related to immune response. Exp Ther Med. 20:1725–1733. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Martinez-Saez O, Borau PG, Alonso-Gordoa
T, Molina-Cerrillo J and Grande E: Targeting HIF-2 alpha in clear
cell renal cell carcinoma: A promising therapeutic strategy. Crit
Rev Oncol Hematol. 111:117–123. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Attwooll C, Denchi EL and Helin K: The E2F
family: Specific functions and overlapping interests. EMBO J.
23:4709–4716. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun CC, Li SJ, Hu W, Zhang J, Zhou Q, Liu
C, Li LL, Songyang YY, Zhang F, Chen ZL, et al: Comprehensive
analysis of the expression and prognosis for E2Fs in human breast
cancer. Mol Ther. 27:1153–1165. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiao Y, Li Y, Fu Z, Hou L, Chen Q, Cai Y,
Jiang P, He M and Yang Z: OGDHL expression as a prognostic
biomarker for liver cancer patients. Dis Markers. 2019:90371312019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wei WY, Yan LH, Wang XT, Li L, Cao WL,
Zhang XS, Zhan ZX, Yu H, Xie YB and Xiao Q: E2F-1 overexpression
inhibits human gastric cancer MGC-803 cell growth in vivo. World J
Gastroenterol. 21:491–501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fang Z, Lin M, Li C, Liu H and Gong C: A
comprehensive review of the roles of E2F1 in colon cancer. Am J
Cancer Res. 10:757–768. 2020.PubMed/NCBI
|
|
22
|
Yin H, Lowery M and Glass J: In prostate
cancer C/EBPalpha promotes cell growth by the loss of interactions
with CDK2, CDK4, and E2F and by activation of AKT. Prostate.
69:1001–1016. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fang DZ, Wang YP, Liu J, Hui XB, Wang XD,
Chen X and Liu D: MicroRNA-129-3p suppresses tumor growth by
targeting E2F5 in glioblastoma. Eur Rev Med Pharmacol Sci.
22:1044–1050. 2018.PubMed/NCBI
|
|
24
|
Li SL, Sui Y, Sun J, Jiang TQ and Dong G:
Identification of tumor suppressive role of microRNA-132 and its
target gene in tumorigenesis of prostate cancer. Int J Mol Med.
41:2429–2433. 2018.PubMed/NCBI
|
|
25
|
Xu H, Fei D, Zong S and Fan Z:
MicroRNA-154 inhibits growth and invasion of breast cancer cells
through targeting E2F5. Am J Transl Res. 8:2620–2630.
2016.PubMed/NCBI
|
|
26
|
Lafta IJ: E2F6 is essential for cell
viability in breast cancer cells during replication stress. Turk J
Biol. 43:293–304. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moreno E, Toussaint MJM, van Essen SC,
Bongiovanni L, van Liere EA, Koster MH, Yuan R, van Deursen JM,
Westendorp B and de Bruin A: E2F7 is a potent inhibitor of liver
tumor growth in adult mice. Hepatology. 73:303–317. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kent LN, Rakijas JB, Pandit SK, Westendorp
B, Chen HZ, Huntington JT, Tang X, Bae S, Srivastava A, Senapati S,
et al: E2f8 mediates tumor suppression in postnatal liver
development. J Clin Invest. 126:2955–2969. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo L, Jones MC, Liu Y, Yv S, Zhu Y and
Guo Y: Cross-cultural validation of the student nurse stress index
scale: A descriptive survey targeting student nurses in China. J
Affect Disord. 251:31–38. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klutstein M, Nejman D, Greenfield R and
Cedar H: DNA methylation in cancer and aging. Cancer Res.
76:3446–3450. 2016. View Article : Google Scholar : PubMed/NCBI
|