Introduction
Mucins are complex cell surface and secreted
glycoproteins that provide protection and lubrication to the
epithelial surface of mucosal tissues (1). The advantage of their expression in
cancer cells is likely linked to their functions in healthy tissue,
promoting epithelial resistance and resilience to toxic challenges
at mucosal surfaces. Carcinoma cells derived from epithelia
overexpress mucins to exploit their role in promoting
proliferation, survival, migration and invasion of cancer cells.
Mucins have thus been identified as markers of poor prognosis
(2) and attractive therapeutic
targets in numerous cancers (3).
The MUC13, cell surface mucin, is overexpressed in
gastric (4), colorectal (CRC)
(5–7), pancreatic (8,9), renal
(10) and ovarian (11) cancers. Usually this protein is
expressed on apical borders of epithelial cells of the intestine,
with increased cytoplasmic expression observed in response to
infection (12) and inflammation
(13). MUC13 has a relatively short
151-amino acid extracellular domain compared with other cell
surface mucins with three epidermal growth factor (EGF)-like
domains, one sea urchin sperm protein enterokinase arginine domain
within an extracellular component, followed by a short 23-amino
acid transmembrane domain and a 69-amino acid cytoplasmic domain
that includes eight serine and two tyrosine residues for potential
phosphorylation. In addition, a protein kinase C consensus
phosphorylation motif could play a role in cell signaling pathways
and regulate proliferation and cell survival (7,13,14).
The elevated cytoplasmic MUC13 expression was
reported in high-grade and metastatic CRC (6). In addition, MUC13 promotes activation
of nuclear factor κB (NFκB), thus is anti-apoptotic (15), and pro-inflammatory (16). Gupta et al (6,7)
suggested that MUC13 overexpression may influence colon
cancer tumorigenesis and metastasis via multiple oncogenic
proteins. It is considered that MUC13 may promote the survival of
CRC cells under DNA damage via numerous mechanisms. Thus, it
represents a potentially important new target for individualized
therapy in advanced patients with CRC. CRC is a multifactorial
disease resulting from individual genetic susceptibility (17,18),
environmental factors (19),
lifestyle, and inflammation (20).
However, the molecular mechanisms responsible for the
overexpression of MUC13 in CRC are poorly understood. It was
hypothesized that one possibility is dysregulation by microRNAs
(miRs) that regulate MUC13 mRNA. In our previous study on
polymorphism in miR binding sites within mucin genes, a significant
association was observed between homozygous variant genotype and
decreased CRC risk for rs1532602 in MUC13 (21). The genetic variations in the 3′
untranslated regions (UTRs) of target genes may affect miRNA
binding, ultimately imposing additional variability into the
differential mRNA and protein expression levels. Aberrant miRNA
expression and/or function are frequently observed in CRC. The
present study thus provided important insights into the role of
miR-4647 which is predicted to bind to polymorphic seed sequence
site in MUC13, SNP spot rs1532602 (Fig. 1). The MUC13-miR-4647 axis was
analyzed and pointed to their differential expression implication
in survival of patients with CRC. CRC is a worldwide health burden,
with nearly 1.2 million new cases expected each year globally
(22). The five-year survival rate
of patients with CRC in stages CRC I and II reaches 90%, while in
advanced stages decreases to 14% (23). For newly diagnosed CRC patients,
clarifying potential risk factors for prognosis and prediction of
therapy response is paramount as it could have important clinical
implications (24).
The present results rendered MUC13-miR-4647
axis as a potential therapeutic target. At present, there is a
great need to identify new diagnostic and prognostic markers, as
well as to develop novel therapeutic strategies for the treatment
of advanced CRC.
Materials and methods
Clinical samples
In total, 187 patients with sporadic CRC were
recruited and underwent surgical resection between 2011 and 2015 at
the General University Hospital (Prague, Czech Republic) and the
Teaching Hospital and Medical School of Charles University (Pilsen,
Czech Republic). All participants signed a written consent to
participate in the study and approved using their biological
samples for genetic analyses according to the Helsinki declaration.
Ethics approval (approval nos. G 09-04-09 and G 14-08-67) was
granted by the committees of the aforementioned hospitals. Written
informed consent was provided by all patients. Study subjects
provided information on their lifestyle habits, body mass index,
diabetes, and family/personal history of cancer using a structured
questionnaire to determine demographic characteristics and
potential risk factors for CRC. The clinical characteristics of
patients are presented in Table
I.
 | Table I.Clinical characteristics of the
patients. |
Table I.
Clinical characteristics of the
patients.
| Clinical
characteristics | Total number
(n=113) |
|---|
| Age, years
(range) | 65±10 (37–86) |
| Sex |
|
|
Male | 69 |
|
Female | 44 |
| SNP
stratificationa |
|
|
G:G | 30 |
|
A:G | 54 |
|
A:A | 17 |
| CRC
stratification |
|
|
Colon | 44 |
|
Rectum | 69 |
| Ta |
|
| 1 | 6 |
| 2 | 19 |
| 3 | 76 |
| 4 | 10 |
| Na |
|
| 0 | 51 |
|
>1 | 50 |
| Ma |
|
| 0 | 87 |
|
>1 | 21 |
| Gradea |
|
| 1 | 19 |
| 2 | 77 |
| 3 | 8 |
| Local
recidivea |
|
| No | 79 |
|
Yes | 33 |
| Neoadjuvant
therapya |
|
| No | 94 |
|
Yes | 16 |
| Adjuvant
therapya |
|
| No | 65 |
|
Yes | 42 |
| Living
statusa |
|
|
Dead | 30 |
|
Alive | 82 |
Tumor tissue and adjacent non-malignant mucosa
tissue (5–10 cm distant from the tumor) were resected from each
patient and deep frozen immediately after removal. The clinical
stage at diagnosis was classified by the tumor-node-metastasis
(TNM) system according to UICC (Union for International Cancer
Control) (detail characteristic in Table I).
All CRC cases were monitored until March 31st, 2021.
Overall survival (OS) was defined as the time interval from
diagnosis to death of any cause or the date of the last follow-up
used for censoring.
Tumor and adjacent mucosal tissues were homogenized
by MagNA Lyser (Roche Diagnostics). Genomic DNA, total RNA, and
small RNA were isolated from tumor tissues and adjacent
non-malignant tissue with miRVana isolation kit protocol according
to the manufacturer's instructions (Thermo Fisher Scientific, Inc.)
without small RNA enrichment. The concentration at 260 nm and
purity of RNA using the 260/280 nm ratio were determined
spectrophotometrically by measuring its optical density using
Nanodrop (Thermo Fisher Scientific, Inc.). The integrity of total
RNA was measured by Agilent 2100 Bioanalyzer with Agilent RNA 6000
Nano Kit (Agilent Technologies, Inc.).
Genotyping of human samples
Genetic polymorphism in the MUC13 gene,
rs1532602, was analyzed with TaqMan allelic discrimination assay
(Thermo Fisher Scientific, Inc.; Assay-on-demand, SNP genotyping
products: C__11906718_1_). The TaqMan genotyping reaction was
amplified on a 7500 Real-Time PCR system (Thermo Fisher Scientific,
Inc.) as follows: 95°C for 10 min, 92°C for 15 sec, and 60°C for 1
min for 40 cycles.
The genotype screening was performed simultaneously
for tumor and adjacent non-malignant tissues. The genotype
correlation between the duplicate samples was >99%. The genotype
call rate ranged between 97.0 and 99.5%.
Reverse transcription-quantitative
(RT-q)PCR of the MUC13 gene
In the present study, the expression levels of
MUC13 and miR-4647 were analyzed. For MUC13 gene
detection (ID Hs00217230_m1, Table
SI), cDNA was synthesized from 80 ng of total RNA in 10 µl RT
reaction using TATAA GrandScript cDNA SuperMix kit (TATAA Biocenter
AB), according to the manufacturer's protocol in C1000 PCR cycler
(Bio-Rad Laboratories, Inc.). Before qPCR, cDNA was diluted 1:1
with nuclease-free water. A total of 10 µl of qPCR reaction
contained 2 µl of diluted cDNA, 5 µl of Taqman universal mastermix
with no UNG (Thermo Fisher Scientific, Inc.), 2.5 µl nuclease-free
water and 0.5 µl of probe assay purchased from Thermo Fisher
Scientific, Inc. The thermocycling conditions were as follows: 95°C
10 min, and 45 cycles of 15 sec at 95°C, 1 min at 60°C in CFX384
(Bio-Rad Laboratories, Inc.). The expression of MUC13 was
normalized to the ACTB gene (Hs99999903_m1). ACTB was
used as reference genes selected from TaqMan Endogenous Control
assays (ACTB, GAPDH, and 18S, Thermo Fisher Scientific,
Inc.) by Normfinder (GenEx Enterprise). All data were analyzed as a
fold change of protein expression and by the 2−ΔΔCq
method (25).
Immunohistochemistry (IHC) of MUC13
gene
Fixed specimens from the intestine wall (received
from patients with CRC) were dehydrated and embedded in paraffin
using routine procedures. Tissue blocks were cut into 5-µm
sections, mounted on Super Frost slides coated with
(3-aminopropyl)-triethoxysilane (Sigma Aldrich; Merck KGaA),
deparaffinized using xylene, rehydrated using a descending ethanol
series, and processed as follows: one section per block was stained
with haematoxylin and eosin- and one section was stained with
Verhoeff's haematoxylin and green trichrome to differentiate tissue
components. Slides with a significant proportion of cancer tissue
were selected for IHC staining. IHC was performed manually using
the primary antibody MUCIN 13 (D-5) (1:100; cat. no. sc-373857;
Santa Cruz Biotechnology, Inc.). Recommended negative and positive
controls were employed.
Histological quantification was performed using
stereological methods and Ellipse software (ViDiTo) as previously
described (26,27). A sample of the human intestinal
wall-ileum and colon-without pathological changes was used as a
positive control (Figs. S1 and
S2). The control colon sample was
used from a patient enrolled in the present study. The result of
IHC detection was in consistency with the manufacturer's
datasheets. The negative control was a control sample of the human
intestinal wall (colon; i.e. a sample containing the proven
glycoprotein mucin 13) but the reaction with the primary antibody
was left out during the IHC procedure. The preparation was negative
(mucin 13 was inconclusive) and therefore a non-specific reaction
of the detection system was ruled out.
Quantitative estimates were performed using
stereological methods and the Ellipse software (ViDiTo) as
established in the previous study on the abdominal aortic aneurysm
(28). The method is based on
counting intersections of detected structures with stereological
grids randomly superposed on the micrographs and has been
previously described (26). This
point-counting method was used for estimating the area fraction of
MUC13 within the intestine wall AA.
The microscope objective and magnification used for
the quantitative assessment of MUC13 was at the lowest setting that
permitted an exact and unambiguous identification of the counting
events with respect to the IHC detection method. The number of
counting events per sample is provided, and the resulting data are
presented as arithmetic means calculated from stochastic methods.
Quantification of MUC13 in the specimens was based on a total of
568 micrographs.
miRNA selection
miR-4647 is predicted to bind to MUC13 only
in the presence of homozygous GG genotype of rs1532602 in
MUC13 according to the freely available software:
MicroSNiper http://epicenter.ie-freiburg.mpg.de/services/microsniper/).
According to mirDIP (https://ophid.utoronto.ca/mirDIP/), Targetscan
(https://www.targetscan.org/vert_80/),
and miRWalk (http://mirwalk.umm.uni-heidelberg.de/) miR-4647 binds
to MUC13 with medium integrated score (mirDIP=0.18,
Targetscan=−0.06, miRWalk=0.85).
RT of miR-4647
For miRNA detection (miR-4647 assay ID 461900_mat),
total RNA was reversely transcribed into cDNA using a pool of
gene-specific primers designed for miRNA analysis using
TaqMan® MiR Reverse Transcription kit (Thermo Fisher
Scientific, Inc.). A pool of 5X Taqman miR primers, each diluted at
1:100, contained all target primers plus spike control miR-39 and
potential reference genes: 18S and small RNA assays: RNU6B,
RNU44, and RNU48. RNU48 (ID 001006) was used as
reference genes selected by Normfinder (GenEx Enterprise). A total
of 10 µl RT reaction contained: 8 ng/µl of RNA, 4 µl of primer
pool, 2 µl RTr enzyme, 0.2 µl of 100 mM dNTPs, 1 µl of 10X RT
buffer, 0.13 µl of RNase inhibitor and 0.67 µl of RNAse-free water.
The thermocycling conditions were as follows: 16°C for 30 min, 42°C
for 30 min and 85°C for 5 min in C1000 (Bio-Rad Laboratories,
Inc.).
cDNA preamplification of miR-4647
All samples were pre-amplified prior to use in the
high-throughput quantitative real-time PCR instrument Biomark
(Fluidigm Corporation). A preamplification pool was prepared from
20X TaqMan MiR Assays (Thermo Fisher Scientific, Inc.), each assay
diluted at 1:100. A total of 10 µl of preamplification reaction
contained: 2 µl of cDNA (not diluted), 1.5 µl of preamp pool, 5 µl
of iQ Supermix (Bio-Rad Laboratories, Inc.) and 1.5 µl RNAse-free
water. Preamplification was performed at C1000 (Bio-Rad
Laboratories, Inc.) as follows: 95°C for 3 min, 18 cycles of 95°C
for 15 sec and 59°C for 4 min. After preamplification, each
reaction was diluted at 1:20.
High-throughput qPCR of miR-4647
MicR qPCR was performed using a high-throughput
platform BioMark™ HD System (Fluidigm Corporation) and
48.48 GE Dynamic Arrays or 96.96 GE Dynamic Arrays. A total of 5 µl
of sample premix contained: 1 µl of the sample (1:20 diluted preamp
cDNA), 2.5 µl of Taqman Universal Mastermix without UNG (Thermo
Fisher Scientific, Inc.), 0.25 µl of 20X GE sample loading reagent
(Fluidigm Corporation) and 1.25 µl of water. A total of 5 µl of
assay pre-mix contained 2.5 µl of 20X Taqman miR assays and 2.5 µl
of 2X assay loading reagent (Fluidigm Corporation). The
thermocycling conditions were as follows: 95°C for 10 min, 40
cycles of 95°C for 15 sec and 60°C for 1 min. All data were
analyzed by the 2−ΔΔCq method (25).
Cell culture
Both CRC cell lines [HCT-116 (cat. no.
CCL-247™) and DLD-1 (cat. no. CCL-221™; both
from American Type Culture Collection) were initially obtained from
ECACC (Sigma-Aldrich; Merck KGaA). Cell lines were cultured at 37°C
in a humidified atmosphere of 5% CO2 in Dulbecco's
Modified Eagle's Medium (DMEM; Sigma-Aldrich; Merck KGaA) and
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 1 mM L-Glutamine and 100 U/ml
penicillin/streptomycin. Both cell lines were examined with
MycoAlert (Lonza Group Ltd.) to exclude mycoplasma
contamination.
Transient cell transfection
Cells were transfected 24 h after seeding with 10 nM
hsa-miR-4647 (cat. no. 4464066; Sigma-Aldrich; Merck KGaA) mimics,
and miRNA mimics negative control with no homology to the human
genome (cat. no. HMC0003; Sigma-Aldrich; Merck KGaA) using
Lipofectamine® RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. All the
experiments were performed in three independent replicates.
The transfection efficiency, changes in miRs levels,
and their impact on MUC13 mRNA levels were confirmed by
real-time RT-qPCR as follows: Specific Silencer® Select
small interfering (si)RNA for human MUC13 (am16708) and
Silencer® Select negative control siRNA #1 (cat. no.
4464058) were purchased from Ambion; Thermo Fisher Scientific, Inc.
and transfected with Lipofectamine® RNAiMAX Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Silencing efficiency was verified by
RT-qPCR 48 h after transfection using the TaqMan® Gene
Expression Assay (Thermo Fisher Scientific, Inc.) for MUC13
(ID Hs00217230_m1).
Isolation and RT-qPCR of RNA from cell
culture samples
Total RNA, including miRNA, was extracted from cells
using Qiagen miReasy Mini Kit (Qiagen GmbH) according to the
manufacturer's protocol. The concentration of the total RNA was
measured by Nanodrop™ 8000 Spectrophotometer (Thermo
Fisher Scientific, Inc.), and the integrity of mRNA in each sample
was measured by Agilent RNA 6000 Nano kit by Agilent Bioanalyzer
2100 (Agilent Technologies, Inc.). To measure levels of miR-4647
after transfection, RT of miRNAs was performed using TaqMan Small
RNA Assay Protocols (Thermo Fisher Scientific, Inc.). cDNA for
MUC13 gene detection after transfection was synthesized from
400 ng of total RNA in 20 µl RT reaction using High-Capacity cDNA
Reverse Transcription kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol in MJ Research PTC-200
Thermal Cycler (Bio-Rad Laboratories, Inc.). cDNA was diluted 1:1
with nuclease-free water. The reaction contained 2 µl of the
sample, 10 µl of Taqman universal mastermix with no UNG (Thermo
Fisher Scientific, Inc.), 1 µl of assay and 7 µl of RNAse free
water. The thermocycling conditions were as follows: 95°C 10 min,
and 45 cycles of 15 sec at 95°C, 1 min at 60°C in 7500 Real Time
PCR System (Thermo Fisher Scientific, Inc.). The expression of
MUC13 was normalized to the ACTB gene. Expression of
miR-4647 measured using TaqMan MiR Assays at 7500 Real Time PCR
System (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: 50°C for 2 min, 95°C for 10 min, 40
cycles of 95°C for 15 sec and 60°C for 60 sec plus melting curve
analysis. The expression of miRNAs was normalized to RNU48
as previously described (29). All
data were analyzed by the 2−ΔΔCq method (25).
Genotyping and sequencing of cell
lines
The assessment of the MUC13 genotype for
rs1532602 in both cell lines was performed similarly as
aforementioned for human samples. The genotype correlation between
the duplicate samples was 100%. The genotype call rate ranged 100%.
To determine which allele of SNP MUC13 rs1532602 is actively
transcribed, total RNA was reversibly transcribed from both cell
lines into cDNA by the aforementioned standard laboratory
procedure. PCR with primers flanking the region of interest
followed afterward. Primers 5′-CCATTGGAGGGATAGAAGCA3′ and
5′-CTTTTCCTGGTAGGGCAACA-3′ designed by online tool primer3
(http://bioinfo.ut.ee/primer3-0.4.0/)
produced 233-bp long amplicon. The final product was diluted at
1:100 in a subsequent, direct sequencing reaction that was
performed using the BigDye Terminator v3.1 Cycle Sequencing kit
(Thermo Fisher Scientific, Inc.) under standard conditions. DNA
sequencing was performed on ABI PRISM 3130 Genetic Analyzer (Thermo
Fisher Scientific, Inc.), and the results were evaluated by
Mutation Surveyor software (SoftGenetics, LLC).
In vitro assays
For colony formation assay, transfected cells were
placed on six-well plates (500 cells per well) 48 h after
transfection. After 12 days, colonies were fixed at room
temperature (RT) for 20 min with 3% formaldehyde and stained with
1% crystal violet at RT for 20 min. The number of colonies (formed
from >50 cells) was counted manually. All measurements were
repeated three times. Cell migration assays were performed 48 h
after transfection using Transwell Permeable Supports 6.5-mm Insert
(24 well plate; 8-µm pore size; Corning, Inc.). Transfected cells
(1×104) were seeded in DMEM with 0.5% FBS on the top of
the Transwell chamber. The lower chamber was filled with 20%
FBS-DMEM. Cells were cultured for 24 h. After the time point, the
migratory cells were fixed with cold 3% formaldehyde for 30 min,
washed with PBS, stained at RT for 20 min with 1% crystal violet,
and counted by light microscope in five random fields under ×200
magnification. All measurements were repeated three times.
The integrity of the plasma membrane was assessed
using the ToxiLight assay (cat. LT17-217; Lonza Group Ltd.)
according to the manufacturer's protocol. This assay measures the
release of adenylate kinase in the extracellular space, which
reflects the plasma membrane's integrity. Cancer cells were
cultured in a 96-well plate (25,000 cells per well) and exposed to
a hsa-miR-4647 (cat. no. 4464066; Sigma-Aldrich; Merck KGaA) miRNA
mimic for 24 h. After incubation at 37°C, 20 µl supernatant of each
well was transferred to a new 96-well plate. Then, 50 µl of assay
buffer was added to each well. After incubation in the dark for 5
min, the luminescence was measured using a Spectramax iD3
(Molecular Devices, LLC).
Western blot analysis
Proteins (20 µg) were loaded and separated in 12%
SDS-PAGE gels at 15 mA for 60 min, and the separated proteins were
then transferred to 0.45-µm Amersham Protran Nitrocellulose
Blotting Membrane (Cytiva) in methanol transfer buffer, using Mini
Trans-Blot Cell (Bio-Rad Laboratories, Inc.). The membranes were
blocked with 5% bovine serum albumin in Tris-buffered saline
containing Tween 20 (TBST; 20 mM Tris-HCl at pH 7.4, 0.15 M NaCl
and 0.1% Tween 20) for 1 h and incubated with anti-MUC13 (1:100;
cat. no. ab235450; Abcam) and anti-GAPDH (1:500; cat. no. ab8245;
Abcam) at 4°C overnight, followed by incubation for 1 h at RT with
goat anti-rabbit secondary antibody (Abcam) conjugated with
horseradish peroxidase. The membranes were developed with
SupersignalWest Pico Chemiluminescent Substrate (Pierce; Thermo
Fisher Scientific, Inc.) and visualized by Azure c600 (Azure
Biosystems, Inc.).
Statistical analysis
Statistical analysis for expression levels was
conducted by SPSS Statistics 20 (IBM Corp.). All qPCR reactions
were run in triplicates. The expression level of each sample was
normalized using pre-selected reference genes and then averaged.
The expression of miR-4647 in patients with CRC was normalized with
the expression of RNU48. The expression of MUC13 was
normalized to the ACTB gene. Expression levels of all
studied genes did not follow a normal distribution in the study
population, as analyzed by the Kolmogorov-Smirnov test. Data were
logarithmically transformed and the paired non-parametric Wilcoxon
and non-parametric Man-Whitney tests were used for statistical
analyses to compare medians. The relationships between the examined
variables and survival were investigated using the Spearman's
correlation, expressed by Spearman's rho (ρ). The survival analysis
was performed using the log-rank test and Kaplan-Meier plots
approach. For in vitro tests, the Man-Whitney test was used
for comparing unpaired data, and the ANOVA test was used for
comparing multiple groups with the Tukey's post hoc test. All
statistical tests were conducted at a 95% confidence level and
P<0.05 was considered to indicate a statistically significant
difference.
Results
Human-based research
Genes differentially expressed in CRC
tissues
Expression levels of MUC13 and miR-4647 were
successfully analyzed. Overall, MUC13 and miR-4647 were
differentially expressed in the present study group: Significantly
higher expression levels for MUC13 were observed in tumor
tissues when compared with adjacent mucosal tissue while for
miR-4647 an opposite trend was observed: Lower expression levels in
tumor tissues (MUC13: 0.32-fold change-32% increase; P=0.02,
Fig. 2A, miR-4647:0.17-fold
change-17% decrease, P=0.05, respectively, Fig. 2B). A negative correlation between
MUC13 and miR-4647 expression levels was observed in tumor
tissue as well as in adjacent non-malignant mucosa (Spearman
correlation r=−0.21, P=0.02 and r=−0.25, P=0.006, respectively;
Fig. 3A and B). Significantly
higher miR-4647 expression levels were observed in women compared
with men (0.20-fold change-20% increase; P=0.009). Patients not
receiving neoadjuvant therapy evinced significantly lower
expression levels of miR-4647 when compared with those receiving
neoadjuvant treatment (0.53-fold change-53% decrease; P=0.05,
respectively). No other associations with clinicopathological data
were observed. All data are summarized in Table II and Fig. S3A-F.
| Table II.Fold change difference in expression
of log2 scale of analyzed genes by non-parametric
Wilcoxon test and Mann-Whitney test. |
Table II.
Fold change difference in expression
of log2 scale of analyzed genes by non-parametric
Wilcoxon test and Mann-Whitney test.
|
| MUC13 | microRNA-4647 |
|---|
|
|
|
|
|---|
| Covariate | Fold change | P-value | Fold change | P-value |
|---|
| Relative expression
(tumor vs. normal mucosa) | 0.32 | 0.02 | −0.17 | 0.05 |
| Sex (female vs.
male) | 0.85 | 0.23 | 0.20 | 0.009 |
| Stratification
(colon vs. rectum) | 0.58 | 0.13 | −0.29 | 0.81 |
| Neoadjuvant therapy
(no vs. yes) | −0.04 | 0.64 | −0.53 | 0.05 |
miR-4647 is differentially expressed
in CRC tissues
To further support the potential importance of
analyzing miRNAs and MUC13 in CRC, their relation to OS was
analyzed. It was observed that higher expression levels of
MUC13 (Fig. 4A, log-rank
P=0.05) and miR-4647 (Fig. 4B,
log-rank P=0.03) were associated with worse survival of
patients.
In silico, miR-4647 is predicted to bind to
the homozygous GG genotype for rs1532602 in the MUC13 gene.
However, in the presence of the variant homozygous AA genotype,
miR-4647 does not bind to the MUC13 gene (Fig. 1). Previously, it has been observed
that the homozygous variant AA genotype of rs1532602 in the miRNA
binding site of MUC13 was associated with a decreased risk
of CRC (21). Notably, patients
carrying the homozygous GG genotype for rs1532602 in MUC13,
together with higher levels of miR-4647, displayed worse OS
(log-rank P=0.04, Fig. 5A). This
could explain the controversial observation of higher miR-4647
expression levels and worse survival of patients. No other
significant association with patient's survival was observed for
heterozygous or variant genotypes (Fig.
5B and C). Additionally, survival analyses were also performed
dividing patients according to the miRSNP rs1532602. No significant
association with prognosis of patients was observed (data not
shown).
IHC
In total, samples from 44 patients included in the
study were analyzed for MUC13 positivity in tumor and adjacent
non-malignant tissue. However, this procedure was not commercially
available for miR-4647.
Control specimens
The control specimens of non-malignant human ileum,
colon ascending and rectum were analyzed. In all specimens, IHC
revealed the MUC13 positivity in columnar epithelial cells
(enterocytes) and goblet cells (base of cells). MUC13 was localized
in the apex of enterocytes and on their surface. In the
non-malignant tunica mucosa of ileum, MUC13 was detected at
Lieberküns' crypts base predominantly, on the top of villi of ileum
in the cytoplasm of enterocytes and also in the base of goblet
cells with intense MUC 13 positivity (Fig. 6A). In the non-malignant colon
tissue, a change in positivity for MUC13 was observed. The
expression of MUC13 was more evenly distributed in the crypts and,
on the contrary, was more pronounced in enterocytes close to
intestinal lumen (Fig. 6B). In the
rectum, more pronounced positivity was observed in the crypts and
the enterocytes (Fig. 6C).
CRC tissue and its stroma
Higher MUC13 expression intensity was observed in
cancer tissue compared with its adjacent non-malignant mucosa
(32.57 vs. 5.04%). In the majority, MUC13 positivity was detected
in the cytoplasm of the cancer cells and in some cells in the
stroma as well (Fig. 6D). In
certain cancer tissue samples, positive vessels for MUC13 were also
detected. Endothelial cells, layer on top of the luminal part of
endothelium and leiomyocytes in larger vessel wall were positive
for MUC13, too (Fig. 6E). Mucinous
tumors expressed MUC13, but at a lower level, based on the lower
staining intensity than adenocarcinomas without mucinous components
(25 vs. 34%).
To investigate whether there is a differential
distribution in MUC13 expression in tumor tissue and non-malignant
mucosa, the population was stratified for the rs1532602 genotype of
the MUC13 gene. No significant distribution of MUC13
expression was observed. However, the patients with homozygous AA
variant genotype evinced lower MUC13 expression levels both in
tumor tissue and non-malignant mucosa when compared with homozygous
GG genotype (Fig. S4A and B).
MUC13 relation to OS was analyzed to support further
the potential importance of MUC13 expression levels in CRC.
Although it was observed that higher expression levels of
MUC13 (Fig. S4C, log-rank
P=0.5) were associated with worse survival of patients, this
association was not significant.
Characteristics of histological sections and
microscopic image fields representing each colorectal tumor sample
used for estimating the parameters are presented in Tables SII and SIII, and Fig.
6F.
Cell culture-based research
Determination of MUC13 genotype
status
To assess the MUC13 genotype for rs1532602 in
both cell lines, genotype screening by TaqMan allelic
discrimination assay was performed. Both cell lines, HCT-116 and
DLD-1, revealed heterozygous GA genotype for MUC13
rs1532602. To ascertain whether cell lines transcribe both alleles,
the corresponding MUC13 cDNA was sequenced. It was confirmed
that both cell lines expressed both alleles (G and A allele) of
MUC13 rs1532602 in the same concentration (Fig. 7).
Determination of transfection
efficiency
To analyze the effects of miR4647 on cell migration
and colony forming, the transfection process of corresponding miRs
precursors was first optimized. HCT-116 and DLD-1 cells were
transfected using Lipofectamine RNAiMAX and efficiency was
subsequently determined by RT-qPCR at 24, 48, 72 and 96 h after
transfection (Fig. 8A and B).
Significantly increased levels of tumor suppressive miR-4647 were
recorded (more than 1,000-fold increase was achieved in both cell
lines, n=3). In the majority, the most eminent effect was observed
24 h after transfection. The silenced MUC13 protein
expression by miR-4647 was confirmed by western blot analysis
(Fig. 8C).
To further examine the MUC13-miR4647 axis,
CRC cell lines were transfected with miR-4647 to detect the
MUC13 mRNA and protein expression levels. The data revealed
that overexpression of miR-4647 inhibited MUC13 mRNA
expression compared with the negative control in the DLD-1
(Fig. 9A, P=0.05) and HCT-116 cells
(Fig. 9B, P=0.003).
To determine whether analyzed miR-4647 affects cell
homeostasis, the HCT-116 and DLD-1 cells were transfected with
miR-4647 precursor and adenylate kinase enzyme in cell medium was
measured 48 h after transfection. It was observed that the
upregulated expression of miR-4647 did not affect the cellular
homeostatic network (Fig. 9C and
D).
miR-4647 inhibits colony formation and
migration of CRC cells
To determine the effect of miRs on cell survival,
the anchorage dependence of proliferation using colony formation
assay was assessed. After 12 days, HCT-116 and DLD-1 cells
transfected with tested miR formed significantly fewer colonies
than cells transfected with control oligonucleotide (miR-4647:
HCT-116 with 20% decrease, P=0.03; DLD-1 with 57% decrease, P=0.05,
respectively; Figs. 10A and B and
S5A).
Migration assay was performed to investigate the
effect of miR on the invasive behavior of cancer cells. The
migratory abilities of CRC cells after transfection with miR-4647
were reduced in both cell lines (miR-4647: HCT-116 with 37%
decrease, P=0.002; DLD-1 with 46% decrease, P=0.001, respectively;
Figs. 10C and D, and S5B).
Silencing of MUC13 influences colony
formation and migration of CRC cells
To evaluate whether the downregulation of
MUC13 is responsible for the observed effects in CRC cells,
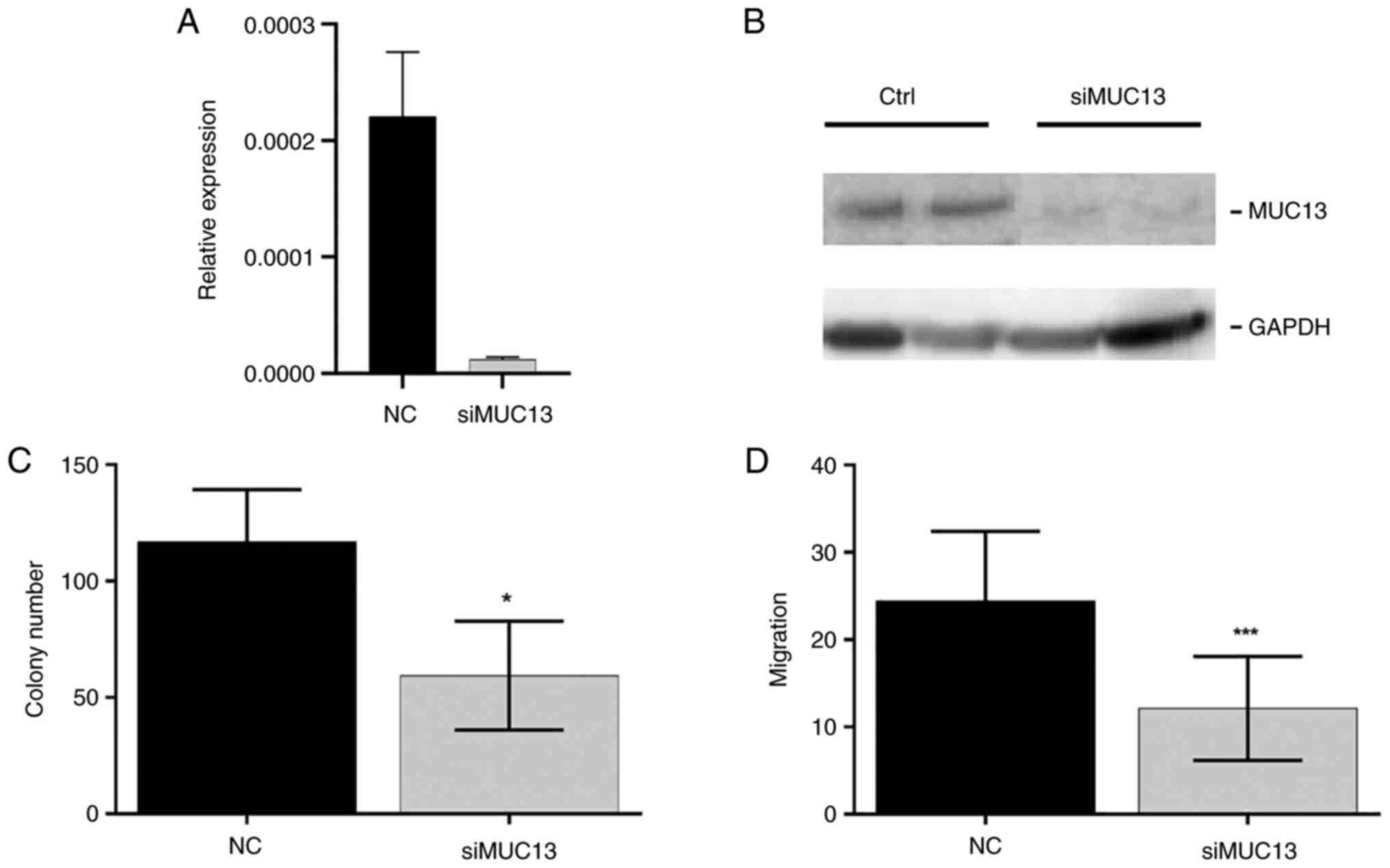
MUC13 was silenced by siRNA in HCT-116 cells. The silenced
MUC13 mRNA levels and protein expression in HTC-116 cells
were confirmed by RT-qPCR and western blot analysis (Figs. 11A and B, and S5C). Lower expression levels of
MUC13 confirmed the suppression of colony formation (33%
decrease, P=0.02, Fig. 11C) and
cell migration (64% decrease, P=0.0003, Fig. 11D). This effect was also observed
after artificial overexpression of miR-4647, thus these data may
indicate the role of MUC13 in CRC cells.
Discussion
Recently, it has been observed that the homozygous
variant AA genotype of rs1532602 in the miR binding site of
MUC13 was associated with a decreased risk of CRC (21). Mucinous CRCs have been found to have
worse survival and a higher TNM stage at diagnosis. These results
inspired the authors to investigate the role of miRNAs regulation
of MUC13. The in silico predicted
MUC13-miR-4647 axis was analyzed and the association of
their differential expression in survival of patients was pointed
out.
Firstly, it was confirmed that MUC13
expression levels decreased after ectopic overexpression of
miR-4647 by RT-qPCR. Further, the decreased miR-4647 expression was
observed in human CRC tissues compared with matching adjacent
non-malignant tissue, while MUC13 was found to be
overexpressed in CRC tumors.
The main result of the present study is the
observation of higher expression levels of MUC13 in
association with poor survival of patients with CRC. In
vitro functional tests confirmed this finding where CRC cells
with attenuated MUC13 expression levels evinced decreased
ability to form colonies. Besides, CRC cells with overexpressed
miR-4647 formed significantly fewer colonies and had a reduced
ability to migrate. MiR-4647 is predicted to bind to the G allele
in polymorphic seed sequence site in MUC13, SNP spot
rs1532602. This observation pointed out that the poorer survival of
CRC cells may be linked to the MUC13 gene and not to
miR-4647 overexpression. Higher expression levels of miR-4647 in
CRC tumors also increased the risk of death in patients with the
homozygous GG genotype of rs1532602 in MUC13. To the best of
our knowledge, the effect of miR-4647 expression levels in patients
with CRC as well as on patient's survival has not been analyzed in
available literature yet. However, this miRNA was recently
presented at ESMO Annals of Oncology and identified as a biomarker
for the selection of patients with chronic myeloid leukemia that
can qualify for achieving deep molecular response and can be
considered for imatinib discontinuation trial (30). Similarly, the analysis of MUC13 in
relation to association with relevant miRNA is, to the best of our
knowledge, performed in the present study for the first time.
The presence of SNPs could also contribute to a
different combination of miRNAs interacting in the region. This
fact may additionally affect the modulation of post-transcriptional
regulation mediated by a single SNP. Therefore, at present, it
cannot be excluded that the observed clinical phenotypes may be the
result of different combinations of miRNAs binding to one of such
predicted SNP. Moreover, a recent study (31) suggested that variations in gene
regions other than 3′UTRs may also affect the binding of
miRNAs.
Notably, higher expression levels of miR-4647 were
observed in association with poor survival of patients with CRC.
This controversial association with the present data should be
considered with caution. One of the hypotheses could be the fact
that according to Targetscan, the other target for miR-4647 is the
tumor necrosis factor receptor superfamily, member 13C
(TNFRSF13C), which plays an important role in B cell
homeostasis, immune system processes, adaptive immune response and
the tumor necrosis factor-mediated signaling pathway. MiRNAs work
to fine-tuning translation through specific mRNA binding. Negative
regulation of TNFRSF13C gene expression by miR-4647 inhibits
its translation and causes degradation of target mRNA. The
association of poorer survival with higher levels of miR-4647 could
be, thus, an explanation of the observed effect. Notably, Sheng
et al observed that MUC13 promoted TNF-induced NF-κB
activation by interacting with TNFR1 and the E3 ligase, cIAP1,
which increased the ubiquitination of RIPK1 (15). However, patients carrying the
homozygous GG genotype for rs1532602 in MUC13 together with
higher levels of miR-4647, displayed worse OS. This could explain
the observation of higher miR-4647 expression levels and worse
patients' survival.
In a healthy colon, MUC13 was detected as a thin
layer on the apical surface of glands. In cancer tissue, higher
MUC13 expression intensity was observed compared with their
adjacent non-malignant mucosa. MUC13 positivity was detected in the
cytoplasm of the cancer cells. On the other hand, in our study,
mucinous tumors expressed MUC13, but at a lower level, based on the
lower staining intensity than adenocarcinomas (25 vs. 34%). In the
present study, we have observed the overexpression of MUC13 in
tumors compared with an adjacent colon. Further studies are
required to clarify the relationships between MUC13 expression and
colon cancer stages and prognosis.
Dysregulated expression of MUC13 has been
shown in ovarian, pancreatic, gastric and colorectal cancers
(4–6,9,11). We
have confirmed that CRC patients with higher expression levels of
MUC13 in tumor tissue displayed worse survival than those
with lower expression levels. These observations agree with the
studies of Gupta et al (6,7). On
the contrary, Packer et al reported that the mRNA level of
MUC13 was decreased in colon cancer; however, this was a
small study with only 23 samples of colon cancer and 6
non-malignant colon tissue samples (32). MUC13 mRNA was also detected
in the blood of CRC patients; however, MUC13 mRNA was also
identified in the blood of healthy individuals (33). Sheng et al (13) observed that MUC13 may protect
epithelial cells in the colon from apoptosis and through targeting
MUC13 and MUC13-regulated pathways sensitize cancer cells to death
and therefore may present an attractive target for cancer treatment
(15).
Chauhan et al (9) showed that MUC13 expression increased
the expression of HER2 in multiple cell types and MUC13 knockdown
resulted in the downregulation of HER2 expression. HER2 belongs to
the human EGF receptor (EGFR/ErbB) family of receptor tyrosine
kinases that have been widely implicated in human cancers (34) and cancer pathogenesis (35). Although it is understood that
receptor activation requires interactions between their specific
ligands, so far, no soluble ligand has been identified for HER2.
However, previous studies suggested that activation of ErbB
receptors can be potentiated by proteins such as mucins, like MUC1
and MUC4 (36). Expression of MUC13
on the surface of CRC cells may also influence the growth
characteristics via interactions with c-erbB growth factor
receptors, modulate adhesion and interfere with immune recognition.
Duan et al (37) suggested
that MUC13 expressed on platelets in rats is involved in the
interaction of platelets with endothelial cells, and, possibly by
the same mechanism, MUC13 on cancer cells interacts with
endothelial ligands during metastasis (14). MUC13 released from the surface of
cancer cells may be a helpful serum diagnostic target in patients
with gastrointestinal cancers.
There is a significant clinical need for biomarkers
to provide an early indication of CRC. Such markers could act as an
adjunct in CRC detection and allow a suitable use of chemotherapy
and monitoring response to treatment (38). Investigations to identify biomarkers
can also identify novel targets for therapy. MiRNAs not only
contribute to diverse biological processes but are also implicated
in the progression and metastasis of human cancers (39). Falzone et al (40) identified that miR-183-5p, miR-21-5p,
miR-195-5p and miR-497-5p are directly related to CRC through the
interaction with the mismatch repair pathway. In another recent
study, clusters miR-17/92a-1, miR-106a/363, miR-106b/93/25 and
miR-183/96/182 showed the strongest association with metastasis
occurrence and poor patient survival (41). Similarly, Wang et al
(42) observed that the
miR-99a-HS3ST2-miR-100 axis, a novel miRNA-mRNA network, was
associated with the presence of lymph node metastasis in CRC.
Insights into the roles of miRNAs in cancer have established them
as targets for novel therapeutic approaches. Previously, several
miRNA-targeted therapeutics have reached clinical development
(43). Consistent with the
aforementioned findings, in the present study, miR-4647 suppressed
proliferation in vitro.
Significantly higher miR-4647 expression levels
were observed in women in comparison with men. Male and female
patients have different endocrine backgrounds. It can be
hypothesized that the changes observed in the expression levels of
miR-4647 may be thus related to sex of patients instead of MUC13 or
CRC.
There are certain limitations to the present study,
such as the absence of non-cancerous cell lines as a control.
However, as the aim of the study was to identify prognostic markers
as well as to disclose novel therapeutic strategies for the
treatment of advanced CRC, the exclusion of non-cancerous cell
lines was not of utmost importance.
There is proof-of-principle evidence that miRNA
SNPs can play a critical role in predicting cancer risk, treatment
response and outcome. Understanding the factors contributing to
cancer risk can represent a powerful future tool for clinicians and
genetic counselors, as well as in advancing our understanding of
cancer biology. If one risk allele or a signature of alleles were
identified, clinicians could advise a specific group of patients to
begin earlier, more frequent, and intensive screening, or even
stronger preventative measures, in hopes of preventing disease or
diagnosing it at an earlier and more treatable stage. More notably,
as miRNAs are stimulated by external stimuli, it is also possible
to manage patients with such SNPs by modifying lifestyle factors to
maintain homeostasis of their inherited differences. This path of
active research may prove the most promising. While assessing an
individual's risk can be a valuable tool for diagnosing cancer at
an earlier stage, the question of the best treatment for individual
patients remains open.
In summary, the present study investigated a
MUC13-miR-4647axis in CRC. The present data revealed the essential
role of MUC13 in survival of patients and the
MUC13-miR-4647axis pathway may provide novel therapeutic
approaches. It is expected that MUC13 may hold significant
potential in cancer screening, diagnosis and treatment.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
The authors would like to thank Dr Ondrej Daum
(Department of Pathology, Faculty Hospital and Faculty of Medicine
in Pilsen, Charles University in Prague, Czech Republic) for the
consultation and help, and Dr Marketa Slajerova and Mrs Jaroslava
Berankova (both from the Department of Histology and Embryology,
Faculty of Medicine in Pilsen, Charles University in Prague, Czech
Republic) for their technical help.
Funding
The present study was supported by the National Science
Foundation (grant no. 22-05942S), the Czech Health Research council
of the Ministry of Health of the Czech Republic (grant no.
NV19-09-00237), the Cooperation Program, research area ‘Oncology
and Haematology’ (grant no. LX22NPO5102), the National institute
for cancer research and the Cooperation Program, research area SURG
and research area MED/DIAG.
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
VV and PV conceptualized the present study. LS, AO,
LB, JH, MU, VKr, JB and OK developed methodology. VKo, VN, JS, LV
and VV conducted formal analysis. LS, JB, VL, MS and JS provided
resources. LS and VV prepared the original draft. LS, AO, VKo, VN,
VKr, JB, VL, MS, LV, MU, JS, PV and VV wrote, reviewed and edited
the manuscript. PV, VL and JS supervised the study. VV, VL, LV, JS,
MS and PV conceived the study. VV, JB, VL, LV, PV and VKr acquired
funding. All authors read and approved the final version of the
manuscript. AO and VKo confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Ethics approval (approval nos. G 09-04-09 and G
14-08-67) was granted by the committees of the General University
Hospital (Prague; Czech Republic) and the Teaching Hospital and
Medical School of Charles University (Pilsen; Czech Republic).
Written informed consent was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
McGuckin MA, Linden SK, Sutton P and
Florin TH: Mucin dynamics and enteric pathogens. Nat Rev Microbiol.
9:265–278. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jonckheere N, Skrypek N and Van Seuningen
I: Mucins and tumor resistance to chemotherapeutic drugs. Biochim
Biophys Acta. 1846:142–151. 2014.PubMed/NCBI
|
|
3
|
Kufe DW: Mucins in cancer: Function,
prognosis and therapy. Nat Rev Cancer. 9:874–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shimamura T, Ito H, Shibahara J, Watanabe
A, Hippo Y, Taniguchi H, Chen Y, Kashima T, Ohtomo T, Tanioka F, et
al: Overexpression of MUC13 is associated with intestinal-type
gastric cancer. Cancer Sci. 96:265–273. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Walsh MD, Young JP, Leggett BA, Williams
SH, Jass JR and McGuckin MA: The MUC13 cell surface mucin is highly
expressed by human colorectal carcinomas. Hum Pathol. 38:883–892.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gupta BK, Maher DM, Ebeling MC, Sundram V,
Koch MD, Lynch DW, Bohlmeyer T, Watanabe A, Aburatani H, Puumala
SE, et al: Increased expression and aberrant localization of mucin
13 in metastatic colon cancer. J Histochem Cytochem. 60:822–831.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta BK, Maher DM, Ebeling MC, Stephenson
PD, Puumala SE, Koch MR, Aburatani H, Jaggi M and Chauhan SC:
Functions and regulation of MUC13 mucin in colon cancer cells. J
Gastroenterol. 49:1378–1391. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khan S, Ebeling MC, Zaman MS, Sikander M,
Yallapu MM, Chauhan N, Yacoubian AM, Behrman SW, Zafar N, Kumar D,
et al: MicroRNA-145 targets MUC13 and suppresses growth and
invasion of pancreatic cancer. Oncotarget. 5:7599–7609. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chauhan SC, Ebeling MC, Maher DM, Koch MD,
Watanabe A, Aburatani H, Lio Y and Jaggi M: MUC13 mucin augments
pancreatic tumorigenesis. Mol Cancer Ther. 11:24–33. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sheng Y, Ng CP, Lourie R, Shah ET, He Y,
Wong KY, Seim I, Oancea I, Morais C, Jeffery PL, et al: MUC13
overexpression in renal cell carcinoma plays a central role in
tumor progression and drug resistance. Int J Cancer. 140:2351–2363.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chauhan SC, Vannatta K, Ebeling MC,
Vinayek N, Watanabe A, Pandey KK, Bell MC, Koch MD, Aburatani H,
Lio Y and Jaggi M: Expression and functions of transmembrane mucin
MUC13 in ovarian cancer. Cancer Res. 69:765–774. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Linden SK, Sutton P, Karlsson NG, Korolik
V and McGuckin MA: Mucins in the mucosal barrier to infection.
Mucosal Immunol. 1:183–197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sheng YH, Lourie R, Linden SK, Jeffery PL,
Roche D, Tran TV, Png CW, Waterhouse N, Sutton P, Florin TH and
McGuckin MA: The MUC13 cell-surface mucin protects against
intestinal inflammation by inhibiting epithelial cell apoptosis.
Gut. 60:1661–1670. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Williams SJ, Wreschner DH, Tran M, Eyre
HJ, Sutherland GR and McGuckin MA: Muc13, a novel human cell
surface mucin expressed by epithelial and hemopoietic cells. J Biol
Chem. 276:18327–18336. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sheng YH, He Y, Hasnain SZ, Wang R, Tong
H, Clarke DT, Lourie R, Oancea I, Wong KY, Lumley JW, et al: MUC13
protects colorectal cancer cells from death by activating the NF-κB
pathway and is a potential therapeutic target. Oncogene.
36:700–713. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sheng YH, Triyana S, Wang R, Das I,
Gerloff K, Florin TH, Sutton P and McGuckin MA: MUC1 and MUC13
differentially regulate epithelial inflammation in response to
inflammatory and infectious stimuli. Mucosal Immunol. 6:557–568.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Naccarati A, Pardini B, Hemminki K and
Vodicka P: Sporadic colorectal cancer and individual
susceptibility: A review of the association studies investigating
the role of DNA repair genetic polymorphisms. Mutat Res.
635:118–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tomlinson IP, Dunlop M, Campbell H, Zanke
B, Gallinger S, Hudson T, Koessler T, Pharoah PD, Niittymäki I,
Tuupanen S, et al: COGENT (COlorectal cancer GENeTics): An
international consortium to study the role of polymorphic variation
on the risk of colorectal cancer. Br J Cancer. 102:447–454. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rattray NJW, Charkoftaki G, Rattray Z,
Hansen JE, Vasiliou V and Johnson CH: Environmental influences in
the etiology of colorectal cancer: The premise of metabolomics.
Curr Pharmacol Rep. 3:114–125. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao Y, Zhang W, Huo M, Wang P, Liu X,
Wang Y, Li Y, Zhou Z, Xu N and Zhu H: XBP1 regulates the protumoral
function of tumor-associated macrophages in human colorectal
cancer. Signal Transduct Target Ther. 6:3572021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vymetalkova V, Pardini B, Rosa F,
Jiraskova K, Di Gaetano C, Bendova P, Levy M, Veskrnova V, Buchler
T, Vodickova L, et al: Polymorphisms in microRNA binding sites of
mucin genes as predictors of clinical outcome in colorectal cancer
patients. Carcinogenesis. 38:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hao M, Wang K, Ding Y, Li H, Liu Y and
Ding L: Which patients are prone to suffer liver metastasis? A
review of risk factors of metachronous liver metastasis of
colorectal cancer. Eur J Med Res. 27:1302022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krizkova V, Dubova M, Susova S, Vycital O,
Bruha J, Skala M, Liska V, Daum O and Soucek P: Protein expression
of ATP-binding cassette transporters ABCC10 and ABCC11 associates
with survival of colorectal cancer patients. Cancer Chemother
Pharmacol. 78:595–603. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Witter K, Tonar Z, Matejka VM, Martinca T,
Jonák M, Rokosný S and Pirk J: Tissue reaction to three different
types of tissue glues in an experimental aorta dissection model: A
quantitative approach. Histochem Cell Biol. 133:241–259. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eberlová L, Tonar Z, Witter K, Křížková V,
Nedorost L, Korabečná M, Tolinger P, Kočová J, Boudová L, Třeška V,
et al: Asymptomatic abdominal aortic aneurysms show histological
signs of progression: A quantitative histochemical analysis.
Pathobiology. 80:11–23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cervena K, Novosadova V, Pardini B,
Naccarati A, Opattova A, Horak J, Vodenkova S, Buchler T, Skrobanek
P, Levy M, et al: Analysis of MicroRNA expression changes during
the course of therapy in rectal cancer patients. Front Oncol.
11:7022582021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guru SA, Sumi MP, Najar IA, Mir AR and
Saxena A: MO10-6 miR-4647 an early biomarker of outcome in chronic
myeloid leukaemia patients. Ann Oncol. 33 (Suppl 6):S4882022.
View Article : Google Scholar
|
|
31
|
Liu C, Rennie WA, Carmack CS, Kanoria S,
Cheng J, Lu J and Ding Y: Effects of genetic variations on
microRNA: Target interactions. Nucleic Acids Res. 42:9543–9552.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Packer LM, Williams SJ, Callaghan S,
Gotley DC and McGuckin MA: Expression of the cell surface mucin
gene family in adenocarcinomas. Int J Oncol. 25:1119–1126.
2004.PubMed/NCBI
|
|
33
|
Lauriola M, Ugolini G, Rosati G, Zanotti
S, Montroni I, Manaresi A, Zattoni D, Rivetti S, Mattei G, Coppola
D, et al: Identification by a digital gene expression displayer
(DGED) and test by RT-PCR analysis of new mRNA candidate markers
for colorectal cancer in peripheral blood. Int J Oncol. 37:519–525.
2010.PubMed/NCBI
|
|
34
|
Settleman J: Predicting response to HER2
kinase inhibition. Oncotarget. 6:588–589. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chaturvedi P, Singh AP, Chakraborty S,
Chauhan SC, Bafna S, Meza JL, Singh PK, Hollingsworth MA, Mehta PP
and Batra SK: MUC4 mucin interacts with and stabilizes the HER2
oncoprotein in human pancreatic cancer cells. Cancer Res.
68:2065–2070. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Senapati S, Das S and Batra SK:
Mucin-interacting proteins: From function to therapeutics. Trends
Biochem Sci. 35:236–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Duan Y, Naruse T, Nakamura M, Yamaguchi Y,
Kawashima T, Morikawa Y, Kitamura T and Suda T: Expression and
functional analysis of a hemopoietic progenitor antigen, NJ-1
(114/A10), in the megakaryocytic lineage. Biochem Biophys Res
Commun. 253:401–406. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu J, Xu Y, Liu S, Qiao L, Sun J and Zhao
Q: MicroRNAs associated with colon cancer: New potential prognostic
markers and targets for therapy. Front Bioeng Biotechnol.
8:1762020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Detassis S, Grasso M, Del Vescovo V and
Denti MA: microRNAs make the call in cancer personalized medicine.
Front Cell Dev Biol. 5:862017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Falzone L, Scola L, Zanghi A, Biondi A, Di
Cataldo A, Libra M and Candido S: Integrated analysis of colorectal
cancer microRNA datasets: Identification of microRNAs associated
with tumor development. Aging (Albany NY). 10:1000–1014. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pidíková P and Herichová I: miRNA clusters
with up-regulated expression in colorectal cancer. Cancers (Basel).
13:29792021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang X, Gao G, Chen Z, Chen Z, Han M, Xie
X, Jin Q, Du H, Cao Z and Zhang H: Identification of the miRNA
signature and key genes in colorectal cancer lymph node metastasis.
Cancer Cell Int. 21:3582021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|