Introduction
Aloe-emodin is a natural active compound present in
the leaves of Aloe vera (1).
Some studies have found that aloe-emodin has numerous biological
properties including antiviral, antimicrobial and hepatoprotective
activities (2). Aloe-emodin has
been reported to exhibit anticancer activity on neuroectodermal
tumors, lung squamous cell carcinoma and hepatoma cells (3–5).
Aloe-emodin has also been shown to inhibit S-phase progression in
both a transformed glia and a human glioma cell line, sensitize
HeLa cells to As2O3 via the generation of
reactive oxygen species, and affect the anticancer activity of
cisplatin by blocking the activation of extracellular
signal-regulated kinase (6–8). However, the effect of aloe-emodin on
human colon cancer cells has yet to be investigated.
Apoptosis is an actively regulated process of cell
death since its intrinsic pathway involves mitochondria (9). Mitochondrial outer membrane
permeabilization in response to cell death triggers (e.g., DNA
damage) is an important early step which is regulated by Bcl-2 and
controls the release of proteins, such as cytochrome c, from
the mitochondria to the cytoplasm where they initiate apoptosis,
ultimately leading to cell death (10). Apoptosis-inducing factor, another
mitochondrial protein that is released into the cytosol and
nucleus, induces chromatin condensation and DNA fragmentation
(11). Members of the caspase
superfamily are produced and activated, a step that hastens the
cell death process involving the caspase-dependent apoptotic
pathway (10).
Casein kinase II is a conserved and ubiquitous
protein serine/threonine kinase engaged in various functions
including normal and abnormal cell proliferation (12). Casein kinase II is localized in both
the cytoplasm and nuclear compartment of healthy cells, but is
predominantly present in the nuclear compartment of cancer cells
and has been found to be deregulated in all carcinomas studied thus
far (13). Evidence links casein
kinase II to apoptosis (14).
Casein kinase II plays a role in the modulation of caspase
susceptibility with Bid, a pro-apoptotic member of the Bcl-2
family. It was shown that casein kinase II phosphorylates Bid close
to the caspase-8 cleavage site, thereby preventing cells from
undergoing apoptosis (15). Thus,
casein kinase II is an important target for the treatment of
cancer, as the disruption of casein kinase II activity prevents the
phosphorylation of Bid, thereby allowing the cleavage of Bid by
caspases, followed by apoptosis (13,16).
Notably, an extensive down-regulation of casein kinase II by
antisense casein kinase II in prostate cancer xenografts was found
to eventually lead to a complete disappearance of the tumor
(13).
To evaluate the anticancer effect of aloe-emodin, we
employed two distinct human colon carcinoma cell lines, DLD-1 and
WiDr. The observation of DNA fragmentation and nuclear morphology,
and the examination of the release of apoptosis-inducing factor and
cytochrome c showed the apoptotic activity of aloe-emodin.
The role of casein kinase II in aloe-emodin-induced apoptosis was
also investigated. This study reports for the first time that the
natural compound aloe-emodin induces apoptosis in human colon
carcinoma cells.
Materials and methods
Aloe-emodin
Aloe-emodin
[1,8-dihydroxy-3-(hydroxymethyl)-anthraquinone; CAS registry no.
481-72-1, EU no. 2075717, purity ≥95%] was purchased from
Sigma-Aldrich Co. (St. Louis, MO, USA). It was dissolved in
dimethylsulfoxide to a concentration of 18.5 mM and stored at −20°C
until use.
Cell culture and treatments
Human colon carcinoma cell lines, DLD-1 and WiDr,
were obtained from the Food Industry Research and Development
Institute (Hsinchu, Taiwan). Cells were cultured in modified
Eagle’s medium (MEM) (Sigma-Aldrich Co.), supplemented with 10%
heat-inactivated fetal bovine serum (Moregate BioTech, Bulimba QLD,
Australia), 1% MEM non-essential amino acid, 100 U/ml penicillin G,
100 μg/ml streptomycin sulfate and 250 ng/ml amphotericin B (all
from Sigma-Aldrich Co.). The two cell lines were grown at 37°C in a
humidified atmosphere containing 5% CO2. Prior to
treatment, the cells were grown to 80–90% confluency and starved by
incubation in basal medium (MEM + 1% MEM non-essential amino acid)
for 24 h. Various concentrations of aloe-emodin (0–0.37 mM in basal
medium) and durations (0, 2, 3, 4, 6, 12, 24 and 48 h) were
applied.
Cell viability assay
Cell viability was assessed using the XTT [sodium
3′-[1-(phenylamino-carbonyl)-3,4-tetrazolium]-bis(4-methoxy-6-nitro)benzene-sulfonic
acid hydrate] assay kit (Sigma-Aldrich Co.), according to the
manufacturer’s instructions. The assay was conducted three times,
independently.
Lactate dehydrogenase activity assay
At the end of the treatment, the culture medium was
centrifuged at 250 × g for 10 min, and the supernatant was saved
for the lactate dehydrogenase activity assay. The lactate
dehydrogenase released from the lysed cells was detected using the
CytoTox 96 Non-Radioactive Cytotoxicity assay (Promega, Madison,
WI, USA), according to the manufacturer’s instructions. The assay
was conducted three times, independently.
DNA fragmentation assay
Treated cells were centrifuged and lysed in lysis
buffer [10 mM Tris-HCl (pH 8.0), 100 mM NaCl, 1% SDS, 1 mM EDTA and
2 mg/ml proteinase K] for 1 h at 65°C. Following two successive
extractions with phenol/chloroform, the DNA samples were
precipitated in ethanol. After washing with 70% ethanol, the DNA
samples were resuspended in TE buffer and subjected to 2% agarose
gel electrophoresis.
Hoechst 33258 staining
Hoechst 33258 staining was performed as described in
a previous study (17). Hoechst
33258-positive nuclei were visualized and photographed using an
Olympus fluorescence microscope (Olympus, Tokyo, Japan).
Isolation of mitochondria and extraction
of mitochondrial proteins
Mitochondria were isolated using a cell
mitochondrial isolation kit (Sigma-Aldrich Co.), according to the
manufacturer’s instructions with minor modifications. Briefly,
treated cells were harvested, resuspended in lysis buffer
containing a protease inhibitor cocktail (Sigma-Aldrich Co.) and
incubated on ice for 1 min. Extraction buffer A (1X) containing a
protease inhibitor cocktail was added, and the solution was
centrifuged at 600 × g for 10 min at 4°C. The supernatant
(containing mitochondria) was carefully transferred to a fresh tube
and centrifuged at 11,000 × g for 10 min at 4°C. Then, the
cytosolic fraction was carefully transferred to a new tube.
Mitochondrial proteins were extracted by suspending the pellet in
CelLytic M Cell Lysis Reagent (Sigma-Aldrich Co.) with a protease
inhibitor cocktail. The samples were stored at −80°C until use. The
protein content of the cytosolic and mitochondrial fractions was
determined using a BCA protein assay kit (Pierce Biotechnology,
Inc., Rockford, IL, USA) with bovine serum albumin as a
standard.
Total protein preparation
Total proteins were extracted with an M-PER
mammalian protein extraction reagent (Pierce Biotechnology, Inc.),
according to the manufacturer’s instructions. The samples were
stored at −80°C until use. The protein concentration was measured
as described above.
Immunoblotting
Denatured protein samples were subjected to 15%
SDS-PAGE. Proteins were transferred to nitrocellulose membranes.
Blocked blots were incubated for 1 h at room temperature with the
primary antibodies monoclonal anti-apoptosis-inducing factor (E-1),
monoclonal anti-cytochrome c (A-8) (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) and polyclonal
anti-phospho Bid-Ser61 (Bethyl Laboratories, Inc., Montgomery, TX,
USA). Monoclonal anti-β-actin (mAbcam 8226) (Abcam Ltd., Cambridge,
UK) was used as a control for equal protein loading. Blots were
further incubated with secondary antibodies conjugated with
alkaline phosphatase for 1 h at room temperature and then incubated
with Lumi-Phos WB Chemiluminescent substrate (Pierce Biotechnology,
Inc.) and exposed to a Fuji medical X-ray film (Fuji Photo Film
Co., Tokyo, Japan). Image processing was performed using the
software Fuji Image Gauge. All experiments were conducted three
times, independently.
Caspase-3 activity assay
The caspase-3 activity was determined using a
caspase-3 assay kit (Sigma-Aldrich Co.). The caspase-3 inhibitor
N-acetyl-Asp-Glu-Val-Asp-CHO (Ac-DEVD-CHO; Biomol Research
Laboratories, Plymouth, PA, USA) was co-treated with aloe-emodin at
a final concentration of 200 μM to inhibit caspase-3 activity in a
parallel experiment (18). The
assay was conducted three times, independently.
Casein kinase II activity assay
Casein kinase II activity was measured using a
casein kinase assay kit (Sigma-Aldrich Co.) combined with the
PKLight HTS protein kinase assay kit (Cambrex Bio Science Rockland,
Inc., Rockland, MA, USA). Briefly, after aloe-emodin treatment,
casein kinase II substrate α-casein and a reaction buffer
containing ATP (both from the casein kinase assay kit) were added
to the cell lysates and incubated at 37°C for 15 min. The remaining
ATP was then consumed by luciferase, and light was generated from
ATP and luciferin (luciferase and luciferin were from the PKLight
HTS protein kinase assay kit). Bioluminescence was measured using a
luminometer (Berthold Detection Systems, Oak Ridge, TN, USA). The
PKLight HTS protein kinase assay kit measures the consumption of
ATP and is based on the bioluminescent measurement of the remaining
ATP present in the samples after kinase activity. The
bioluminescent signal is inversely proportional to the activity of
the kinase, that is, a higher value is indicative of an increased
amount of remaining ATP and a lower casein kinase II activity in
the cells. The assay was conducted three times, independently.
Statistical analysis
The unpaired Student’s t-test was used to identify
means that were significantly different from each other
(P<0.05).
Results
Aloe-emodin induced cell death in a dose-
and time-dependent manner
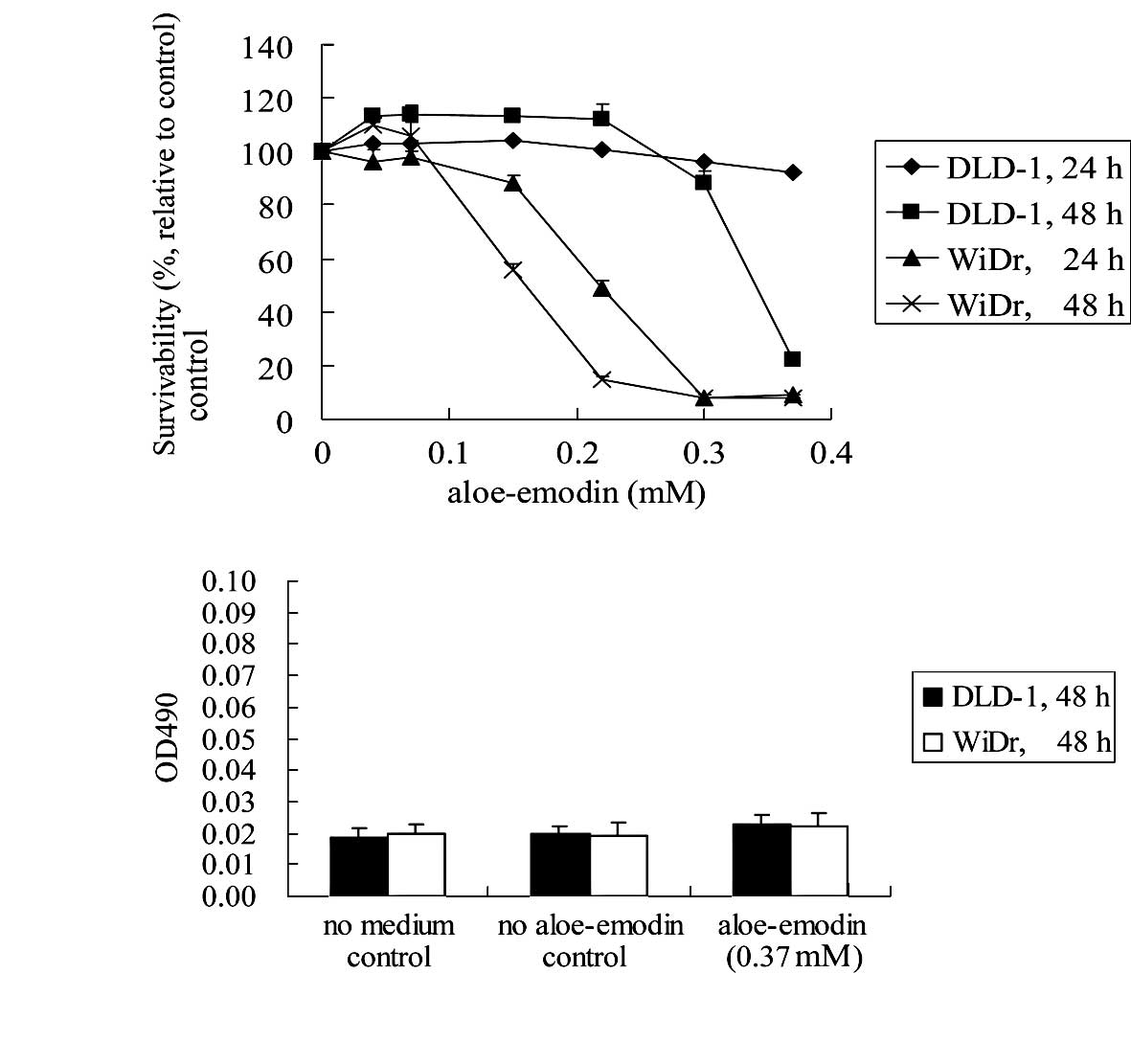
The effect of aloe-emodin on the viability of DLD-1
and WiDr colon carcinoma cells was examined using the XTT viability
assay. Exposure of DLD-1 cells to various concentrations of
aloe-emodin (0.04, 0.07, 0.15, 0.22, 0.30 and 0.37 mM) for 24 or 48
h resulted in a dose- and time-dependent decrease in cell viability
relative to control cell cultures (Fig.
1A). The IC50 value for 48 h of exposure to
aloe-emodin was 0.30–0.37 mM. A similar effect of aloe-emodin was
observed after 24 or 48 h of incubation using the WiDr cell line
(Fig. 1A). In the latter case,
however, the IC50 value for a 48-h exposure to
aloe-emodin was 0.15–0.22 mM, suggesting that the WiDr cells were
more sensitive to aloe-emodin than the DLD-1 cells.
Aloe-emodin-induced cell death was significant at a concentration
of 0.37 mM. Thus, this concentration was applied for the rest of
the experiments.
Aloe-emodin did not induce necrosis in
DLD-1 and WiDr cells
Treated cells were evaluated for the presence of
necrotic cell death by measuring the lactate dehydrogenase release
into the medium. Exposure of DLD-1 and WiDr cells to 0.37 mM
aloe-emodin for 48 h did not result in the release of lactate
dehydrogenase (Fig. 1B).
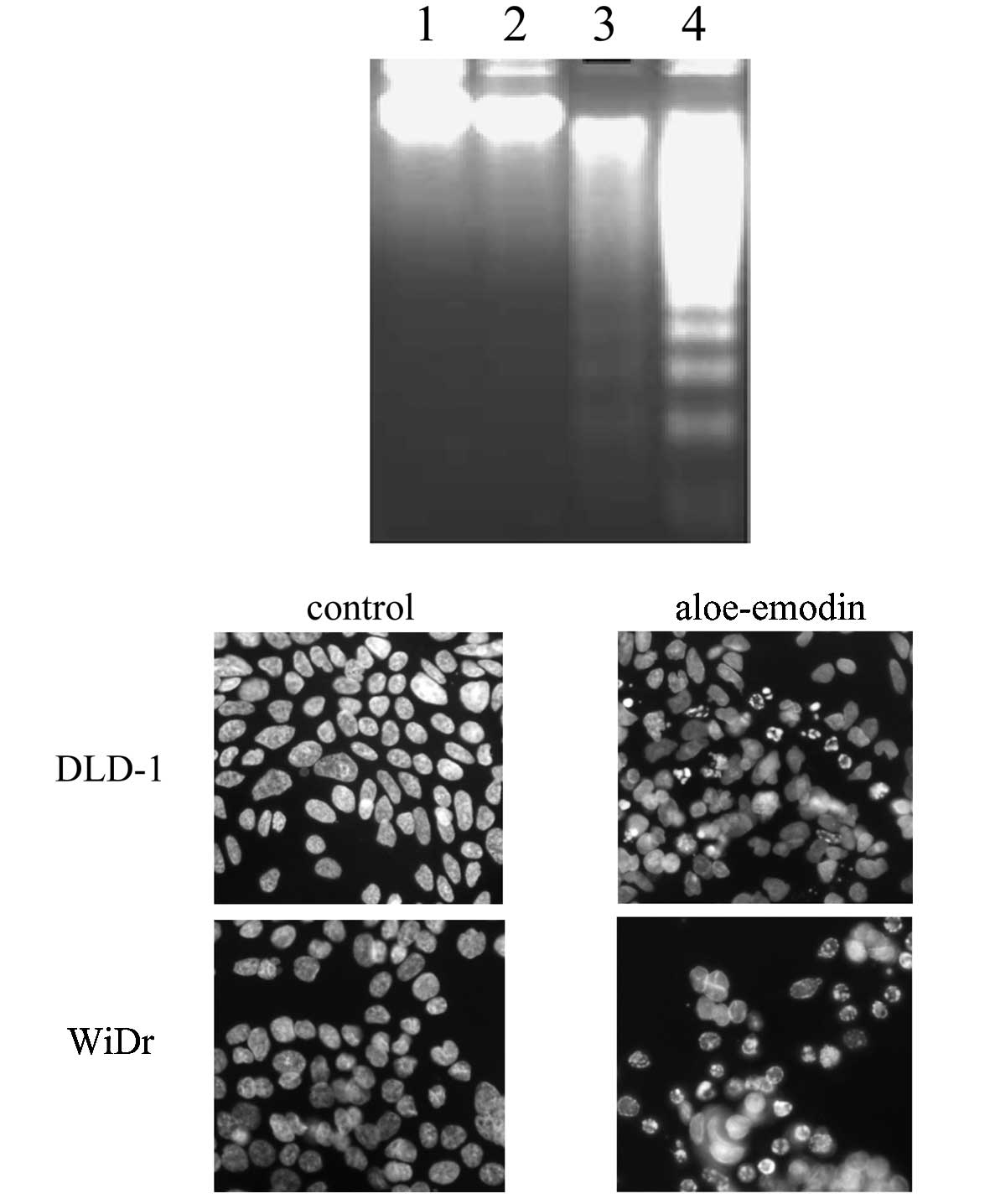
Aloe-emodin induced apoptosis in DLD-1
and WiDr cells
To investigate whether the aloe-emodin-mediated cell
death in DLD-1 and WiDr cells was due to an apoptotic mechanism,
DNA fragmentation and nuclear morphological changes that occurred
during aloe-emodin treatment were observed. Treatment of DLD-1 and
WiDr cells with 0.37 mM aloe-emodin for 24 h resulted in DNA
fragmentation (Fig. 2A). Treatment
of DLD-1 and WiDr cells with 0.37 mM aloe-emodin for 24 h also
resulted in changes in the nuclear morphology as evidenced by the
Hoechst 33258 staining (Fig. 2B), a
dye often used to label DNA in living cells and to observe
morphological and nuclear changes (17,19,20).
More fragmented nuclei were observed upon aloe-emodin treatment,
both in the DLD-1 and WiDr cells. These observations indicated that
aloe-emodin-induced cell death in DLD-1 and WiDr cells involved a
typical apoptotic pathway.
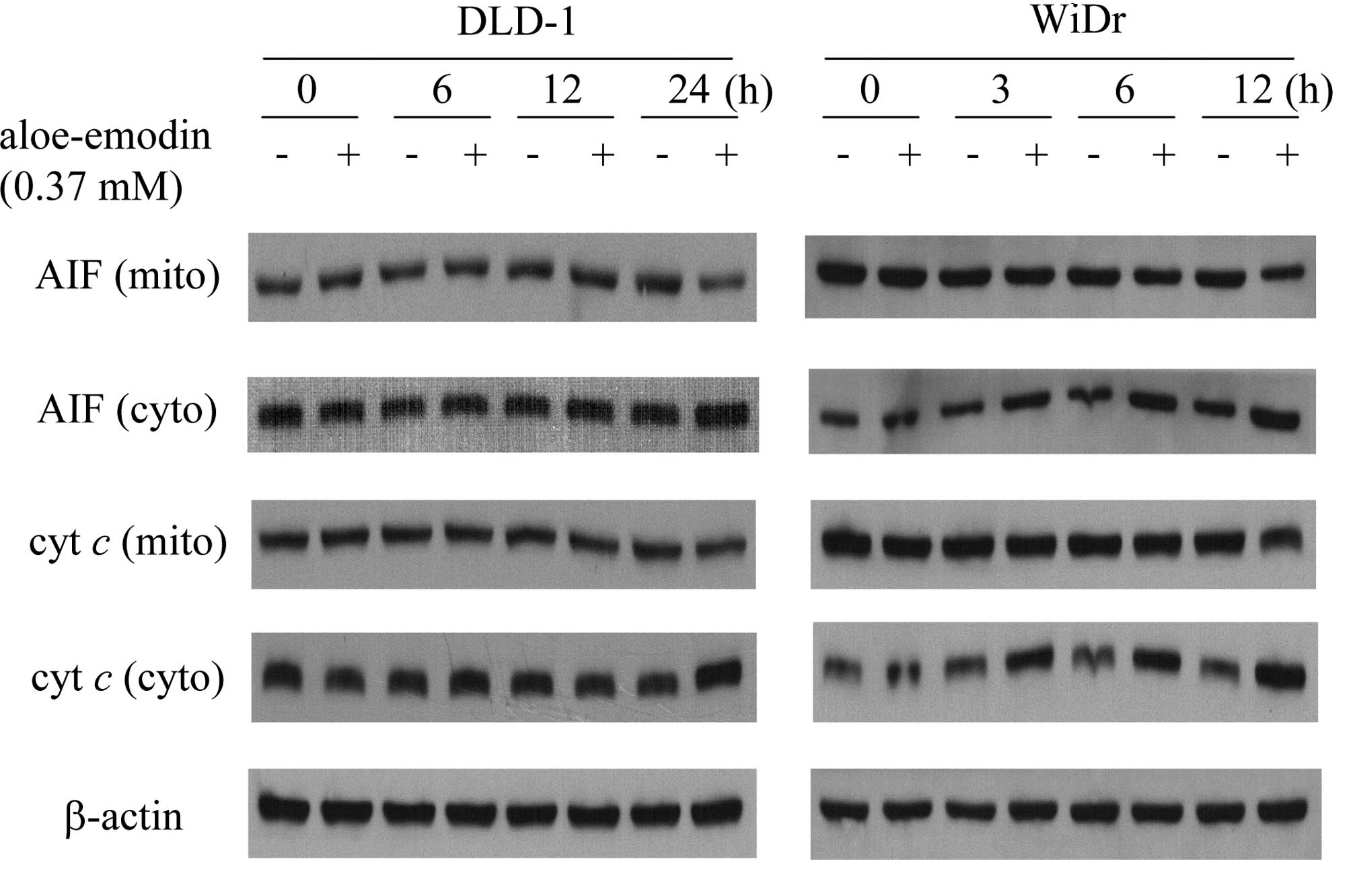
Aloe-emodin induced the release of
apoptosis-inducing factor and cytochrome c
The release of apoptosis-inducing factor and
cytochrome c in DLD-1 and WiDr cells upon treatment with
aloe-emodin was characterized because these processes are typically
related to apoptosis (14). An
immunoblot analysis of the mitochondrial and cytosolic fractions
derived from aloe-emodin-treated DLD-1 and WiDr cells showed a
significant decrease in the amount of apoptosis-inducing factor and
cytochrome c in mitochondrial fractions. An increase in
apoptosis-inducing factor and cytochrome c in the cytosolic
fraction for the indicated time intervals was also noted (Fig. 3). Following treatment of the DLD-1
cells, the release of apoptosis-inducing factor and cytochrome
c commenced at 24 h, while that of WiDr cells started after
3 h of treatment. The results were consistent with the cell
viability data and confirm that WiDr cells are more sensitive to
aloe-emodin than DLD-1 cells.
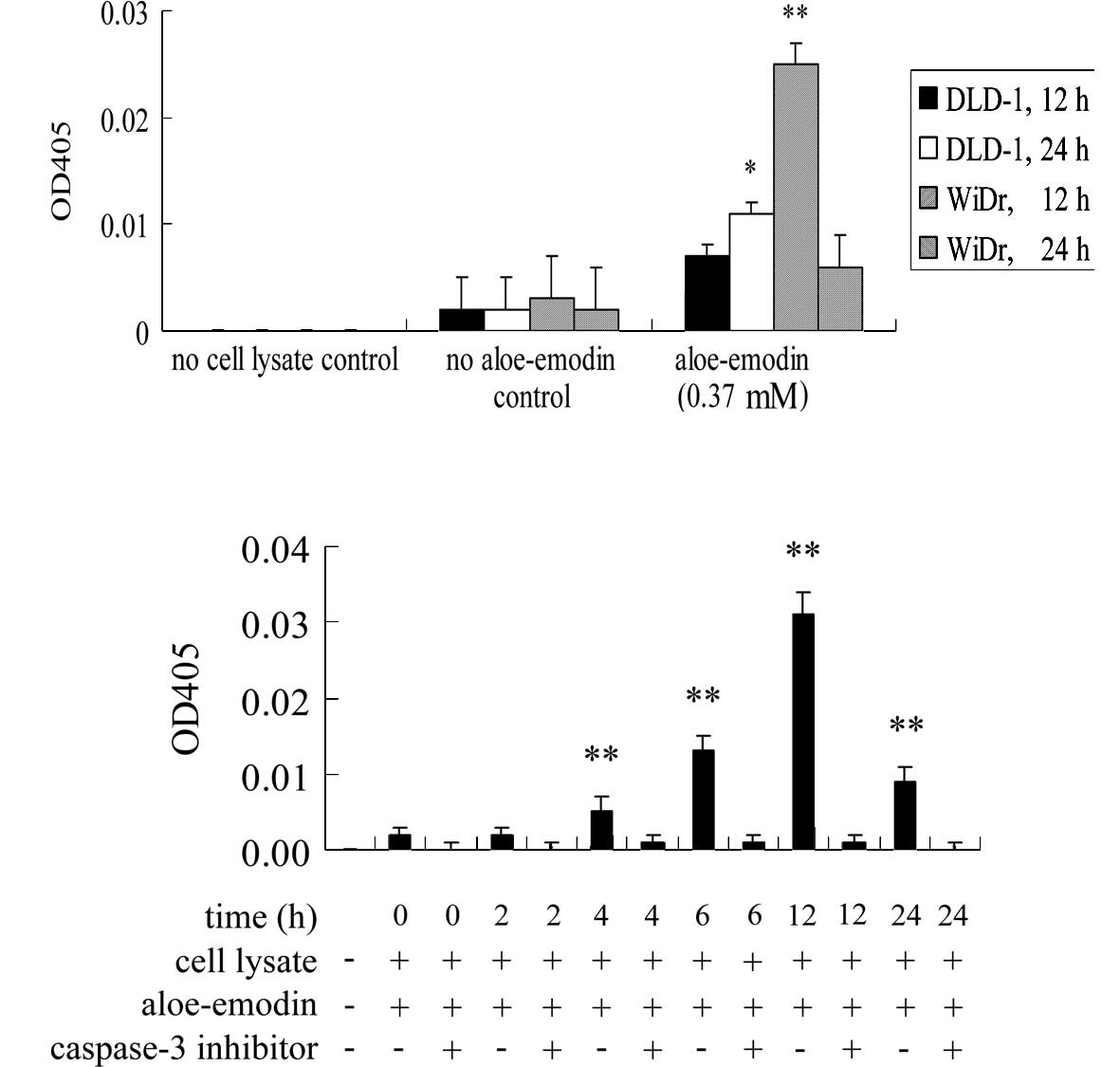
Effect of aloe-emodin on the activation
of caspase-3
The involvement of caspase-3 in aloe-emodin-induced
apoptosis in DLD-1 and WiDr cells was evaluated. The activity of
caspase-3 was significantly induced 12 h after aloe-emodin
treatment in WiDr cells (Fig. 4A).
This effect was also observed in DLD-1 cells albeit only
significantly after 24 h of aloe-emodin treatment (Fig. 4A). To illustrate the sequence of
caspase-3 activation and the release of apoptosis-inducing factor
and cytochrome c, a time course of caspase-3 activation
after aloe-emodin treatment was performed in WiDr cells. As shown
in Fig. 4B, caspase-3 activity was
significantly induced after 4 h of aloe-emodin treatment. The
results showed that the release of apoptosis-inducing factor and
cytochrome c started before caspase-3 activation. To
demonstrate whether the activation of caspase-3 is essential for
aloe-emodin-mediated apoptosis, the influence of the peptidyl
inhibitor of caspase-3, Ac-DEVD-CHO, a generally used caspase-3
inhibitor, was analyzed. Aloe-emodin-induced caspase-3 activation
in WiDr cells was blocked when co-treated with Ac-DEVD-CHO
(Fig. 4B). These data showed that
the activation of caspase-3 was an essential step in
aloe-emodin-induced apoptosis in the two cell lines. However, the
DLD-1 cells appeared to be less sensitive than the WiDr cells (data
not shown).
Effect of aloe-emodin on casein kinase II
activity and Ser61 phosphorylation of Bid
To investigate the role of casein kinase II in
aloe-emodin-induced apoptosis, the activity of casein kinase II was
examined, as well as the Ser61 phosphorylation of Bid, a downstream
substrate of casein kinase II (Ser61 is one of the phosphorylation
sites of Bid). The activity of casein kinase II started to decrease
at 3 h of exposure to aloe-emodin and continued to decrease in a
time-dependent manner in WiDr cells (Fig. 5A). A similar effect was observed in
the DLD-1 cells (Fig. 5A); however,
the activity of casein kinase II started to decrease after 12 h of
exposure to aloe-emodin. Using anti-phospho Bid-Ser61 polyclonal
antibodies, immunoblotting was used to confirm the kinase activity.
The aloe-emodin-mediated inhibition of casein kinase II activity
caused an inhibition of the Ser61 phosphorylation of Bid (Fig. 5B). These results suggest that
aloe-emodin induces apoptosis in DLD-1 and WiDr cells through
inhibition of the phosphorylation of Bid.
Discussion
The present study demonstrated that aloe-emodin
exhibits anticancer activity in DLD-1 and WiDr colon carcinoma
cells and that the cytotoxic mechanism involves the induction of
apoptosis. The latter was characterized by the release of
apoptosis-inducing factor and cytochrome c from the
mitochondria to the cytosol, activation of caspase 3 and inhibition
of the activity of casein kinase II, with the subsequent inhibition
of the downstream phosphorylation of Bid. It was reported that,
using the human lung squamous carcinoma cell line CH27, apoptosis
induced by aloe-emodin was associated with changes in the
expression of members of the Bcl-2 family apoptosis regulators
(i.e., increase in the Bax level and decrease in Bcl-XL).
Aloe-emodin was found to cause cytochrome c release from
mitochondria (21). In addition, it
was observed that aloe-emodin-induced apoptosis in human
hepatocellular carcinoma cell lines HepG2 and Hep3B was accompanied
by the induction of p53 and p21 expression (5). Another report suggests that the
decrease in the expression of protein kinase Cδ and ɛ isoforms
plays a critical role in aloe-emodin-induced apoptosis (21). A similar role of protein kinase C
was observed in U-937 glioma cells (6). With the exception of protein kinase
Cα, most of the protein kinase C isozymes (protein kinase Cα, βI,
γ, δ, ɛ, θ, ξ and μ) were found to decrease upon aloe-emodin
treatment. Notably, two hydrophobic residues unique to casein
kinase II, Val66 and Ile174, were found to be essential for the
interaction with aloe-emodin; a Val66Ala casein kinase II variant
exhibited a substantially affected interaction with aloe-emodin
(22,23).
In the present study, aloe-emodin-induced apoptosis
in colon carcinoma cells involved inhibition of casein kinase II
activity and blockage of the phosphorylation of Bid. We also
demonstrated that aloe-emodin resulted in the release of
apoptosis-inducing factor and cytochrome c from
mitochondria, followed by the activation of caspase-3. It is known
that phosphorylated Bid is cleaved by caspase-8, and truncated Bid
binds to mitochondria to induce the release of apoptosis-inducing
factor and cytochrome c from mitochondria (10,24,25).
It is also known that, in the cytosol, cytochrome c mediates
the allosteric activation of Apf-1, which is required for the
proteolytic maturation of caspase-9 and -3 (10,26).
Thus, the inhibition of casein kinase II activity following
aloe-emodin treatment may lead to the activation of caspase-3
through the phosphorylation of Bid and release of
apoptosis-inducing factor and cytochrome c from
mitochondria. A similar pathway was described for Radix Paeoniae
Alba extract-induced apoptosis in HL-60 leukemic cells and for
quercitin-mediated cell apoptosis in murine melanoma B16-BL6 cells
(27,28). Ahmad et al found that the
down-regulation of casein kinase II by antisense RNA elicited
intracellular hydrogen peroxide production, which was associated
with caspase-3 activation (29).
Although not relative to aloe-emodin, this is another possible
pathway between casein kinase II and caspase-3. Thus, further
studies are required to elucidate the relationship between casein
kinase II and caspase-3 activities.
Notably, in all of the assays used in the present
study, the WiDr colon carcinoma cells were significantly more
sensitive to aloe-emodin than the DLD-1 cells. The reason for this
difference in sensitivity remains unclear. Controversial data have
been analyzed to attempt to correlate differences in response to a
given drug with the differentiation state of a given cell line. In
an in vitro system using epidermal cells, maturing basal
cells were found to undergo apoptosis upon drug treatment, whereas
stem cells appeared to be resistant (30). In addition, it has been suggested
that undifferentiated keratinocyte stem cells are protected from
spontaneous apoptosis (31).
Similarly, differentiated human neural crest-derived tumor cells
exhibit an increased resistance to potassium ionophore-mediated
apoptosis. Cells and human keratinocytes that are undifferentiated
are more sensitive to H2O2-induced cell death
than their differentiated counterparts (32,33).
However, it was shown that the TNF-related apoptosis-inducing
ligand triggers apoptosis in undifferentiated but not in
differentiated human keratinocytes (34). Microscopy analyses have demonstrated
that DLD-1 cells are less differentiated than WiDr cells (data not
shown). Thus, our results strongly suggest that the degree of
differentiation of colon carcinoma cells affects the cellular
response to aloe-emodin. In addition to the degree of
differentiation of culture cells, evidence shows that p53-mutant
cell lines are less sensitive to aloe-emodin than their p53
wild-type counterparts (35). In
the present study, the amount of mutant p53 in the DLD-1 cells was
seven times higher than that in the WiDr cells (1.96 and 0.27 ng/ml
cell lysate, respectively). Our data are consistent with the
above-mentioned study and suggest that the p53 phenotype affects
the cellular response to aloe-emodin.
In conclusion, using two human colon carcinoma cell
lines, we investigated the anticancer effect of the natural
product, aloe-emodin, isolated from Aloe vera leaves. We
demonstrated for the first time that aloe-emodin exhibits an
anticancer effect against colon carcinoma.
Acknowledgements
The authors would like to thank Dr Sheng-Hsuan Chen,
Dr Chun-Chao Chang and Dr Chia-Lang Fang for the helpful comments
and review of the manuscript.
References
|
1
|
Reynolds T: The compounds in Aloe leaf
exudates: a review. Bot J Linn Soc. 90:157–177. 1985. View Article : Google Scholar
|
|
2
|
Eshun K and He Q: Aloe vera: a valuable
ingredient for the food, pharmaceutical and cosmetic industries – a
review. Crit Rev Food Sci Nutr. 44:91–96. 2004.PubMed/NCBI
|
|
3
|
Pecere T, Gazzola MV, Mucignat C, Parolin
C, Vecchia FD, Cavaggioni A, Basso G, Diaspro A, Salvato B, Carli M
and Palu G: Aloe-emodin is a new type of anticancer agent with
selective activity against neuroectodermal tumors. Cancer Res.
60:2800–2804. 2000.PubMed/NCBI
|
|
4
|
Lee HZ, Hsu SL, Liu MC and Wu CH: Effects
and mechanisms of aloe-emodin on cell death in human lung squamous
cell carcinoma. Eur J Pharmacol. 431:287–295. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuo PL, Lin TC and Lin CC: The
antiproliferative activity of aloe-emodin is through p53-dependent
and p21-dependent apoptotic pathway in human hepatoma cell lines.
Life Sci. 71:1879–1892. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
MAcevedo-Duncan M, Russell C, Patel S and
Patel R: Aloe-emodin modulates PKC isoenzymes, inhibits
proliferation and induces apoptosis in U-373MG glioma cells. Int
Immunopharmacol. 4:1775–1784. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yi J, Yang J, He R, Gao F, Sang H, Tang X
and Ye RD: Emodin enhances arsenic trioxide-induced apoptosis via
generation of reactive oxygen species and inhibition of survival
signaling. Cancer Res. 64:108–116. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mijatovic S, Maksimovic-Ivanic D, Radovic
J, Miljkovic D, Kaludjerovic GN, Sabo TJ and Trajkovic V: Aloe
emodin decreases the ERK-dependent anticancer activity of
cisplatin. Cell Mol Life Sci. 62:1275–1282. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Penninger JM and Kroemer G: Mitochondria,
AIF and caspases – rivaling for cell death execution. Nat Cell
Biol. 5:97–99. 2003.
|
|
10
|
Kim R, Emi M and Tanabe K: Role of
mitochondria as the gardens of cell death. Cancer Chemother
Pharmacol. 57:545–553. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Susin SA, Lorenzo HK, ZamZami N, et al:
Molecular characterization of mitochondrial apoptosis-inducing
factor. Nature. 397:441–446. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kikkawa U, Mann SK, Firtel RA and Hunter
T: Molecular cloning of casein kinase II alpha subunit from
Dictyostelium discoideum and its expression in the life
cycle. Mol Cell Biol. 12:5711–5723. 1992.PubMed/NCBI
|
|
13
|
Ahmad KA, Wang G, Slaton J, Unger G and
Agmed K: Targeting CK2 for cancer therapy. Anti-Cancer Drugs.
16:1037–1043. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ahmed K, Gerber DA and Cochet C: Joining
the cell survival squad: an emerging role for protein kinase CK2.
Trends Cell Biol. 12:226–230. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Desagher S, Osen-Sand A, Montessuit A,
Magnenat E, Vibois F, Hochmann A, Journot L, Antonsson B and
Martinou JC: Phosphorylation of bid by casein kinases I and II
regulates its cleavage by caspase 8. Mol Cell. 8:601–611. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Unger GM, Davis AT, Slaton JW and Ahmed K:
Protein kinase CK2 as regulator of cell survival: implications for
cancer therapy. Curr Cancer Drug Targets. 4:77–84. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin KY, Wang HH, Lai ST, Pan JP and Chiang
AN: β2-glycoprotein I protects J774A.1 macrophages and
human coronary artery smooth muscle cells against apoptosis. J Cell
Biochem. 94:485–496. 2005.
|
|
18
|
Chatterjee PK, Todorovic Z, Sivarajah A,
Mota-Filipe H, Brown PAJ, Stewart KN, Cuzzocrea S and Thiemermann
C: Differential effects of caspase inhibitors on the renal
dysfunction and injury caused by ischemia-reperfusion of the rat
kidney. Eur J Pharmacol. 503:173–183. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Martin RM, Leonhardt H and Cardoso MC: DNA
labeling in living cells. Cytometry Part A. 67A:45–52. 2005.
View Article : Google Scholar
|
|
20
|
Sharma S, Dube A and Bose B:
Pharmacokinetics and phototoxicity of purpurin-18 in human colon
carcinoma cells using liposomes as delivery vehicles. Cancer
Chemother Pharmacol. 57:500–506. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee HZ: Protein kinase C involvement in
aloe-emodin- and emodin-induced apoptosis in lung carcinoma cell.
Br J Pharmacol. 134:1093–1103. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sarno S, Moro S, Meggio F, Zagotto G, Ben
DD, Ghisellini P, Battistutta R, Zanotti G and Pinna LA: Toward the
rational design of protein kinase casein kinase-2 inhibitors.
Pharmacol Ther. 93:159–168. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Litchfield DW: Protein kinase CK2:
structure, regulation and role in cellular decisions of life and
death. Biochem J. 369:1–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chau DHW, Yuan J, Zhang H, Cheung P, Lim
T, Liu Z, Sall A and Yang D: Coxsackievirus B3 proteases 2A and 3C
induce apoptotic cell death through mitochondria injury and
cleavage of eIL4GI but not DAP5/p97/NAT1. Apoptosis. 12:513–524.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shulga N and Pastorino JG: Acyl coenzyme
A-binding protein augments Bid-induced mitochondrial damage and
cell death by activating μ-calpain. J Biol Chem. 281:30824–30833.
2006.PubMed/NCBI
|
|
26
|
Garrido C, Galluzzi L, Brunet M, Puig PE,
Didelot C and Kroemer G: Mechanisms of cytochrome c release from
mitochondria. Cell Death Differ. 13:1423–1433. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kwon KB, Kim EK, Han MJ, et al: Induction
of apoptosis by Radix Paeoniae Alba extract through
cytochrome c release and the activations of caspase-9 and caspase-3
in HL-60 cells. Biol Pharm Bull. 29:1082–1086. 2006.
|
|
28
|
Zhang XM, Chen J, Xia YG and Xu Q:
Apoptosis of murine melanoma B16-BL6 cells induced by quercetin
targeting mitochondria, inhibiting expression of PKC-α and
translocating PKC-δ. Cancer Chemother Pharmacol. 55:251–262.
2005.PubMed/NCBI
|
|
29
|
Ahmad KA, Wang G and Ahmed K:
Intracellular hydrogen peroxide production is an upstream event in
apoptosis induced by down-regulation of casein kinase 2 in prostate
cancer cells. Mol Cancer Res. 4:331–338. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morris RJ, Fischer SM, Klein-Szanto AJ and
Slaga TJ: Subpopulations of primary adult murine epidermal basal
cells sedimented on density gradients. Cell Tissue Kinet.
23:587–602. 1990.PubMed/NCBI
|
|
31
|
Tiberio R, Marconi A, Fila C, et al:
Keratinocytes enriched for stem cells are protected from anoikis
via an integrin signaling pathway in a Bcl-2 dependent manner. FEBS
Lett. 524:139–144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Teplova V, Jaaskelainen E,
Salkinoja-Salonen M, Saris NE, Serlachius M, Li FY and Andersson
LC: Differentiated Paju cells have increased resistance to toxic
effects of potassium ionophores. Acta Biochim Pol. 51:539–544.
2004.PubMed/NCBI
|
|
33
|
Zuliani T, Denis V, Nobless E, Schnebert
S, Andre P, Dumas M and Ratinaud MH: Hydrogen peroxide-induced cell
death in normal human keratinocytes is differentiation dependent.
Free Radic Biol Med. 38:307–316. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jansen BJ, van Ruissen F, Cerneus S, Cloin
W, Bergers M, van Erp PE and Schalkwijk J: Tumor necrosis factor
related apoptosis inducing ligand triggers apoptosis in dividing
but not in differentiating human epidermal keratinocytes. J Invest
Dermatol. 121:1433–1439. 2003. View Article : Google Scholar
|
|
35
|
Pecere T, Sarinella F, Salata C, et al:
Involvement of p53 in specific anti-neuroectodermal tumor activity
of aloe-emodin. Int J Cancer. 106:836–847. 2003. View Article : Google Scholar : PubMed/NCBI
|