Introduction
The search for compounds that prevent cancer has
intensified with mounting evidence that many types of cancer are
caused or triggered by factors related to lifestyle and
environment, particularly agents in foodstuffs. There is a growing
body of evidence from in vitro and in vivo
experiments that N. sativa seeds (black cumin seeds) or the
extracted volatile oils, exert anti-carcinogenic effects (1,2). N.
sativa is a common herbal plant that has been used since
ancient times by the Egyptians and Greeks for various medicinal
purposes. It has many common applications by people in the
countries surrounding the Mediterranean Sea, the Arab world,
Indians and Persians as an antimicrobial, antihelmintic, diuretic,
diaphonetic and antiasthmatic medicine. N. sativa is also
used by the Europeans, Americans and Japanese as a food additive
for cheese and bakery products (3).
A large number of in vitro and in vivo
studies have shown significant modulatory effects of N.
sativa extracts with different pathological, toxicological and
cytotoxic protocols. Previously, volatile oils from N.
sativa seeds were found to possess antitumor effects in
vitro against Ehrlich ascites carcinoma, Dalton's lymphoma
ascites and sarcoma (S-180) cell lines (4). Recently, it was postulated that
extracted proteins from N. sativa exhibit modulatory effects
on the immune system (5) with
anti-protein-oxidation activity. This effects may be rheologically
useful, particularly in pathological conditions related to free
radical stress (6). A previous
study showed that extra-virgin crude volatile oils extracted from
Egyptian N. sativa exert significant inhibitory effects on
rat colon carcinogenesis when administered in the post-initiation
phase along with a marked anti-proliferative effect on the colonic
epithelium when administered at different stages of colon
carcinogenesis (with pre-carcinogen, simultaneous or
post-carcinogen administration protocols) (7).
In the present study, a rat medium-term multi-organ
carcinogenesis bioassay (8) was
used to detect the anti-carcinogenic potential of N. sativa.
This assay was based on the two-stage concept of carcinogenesis
using five different genotoxic carcinogens applied for initiation:
diethylnitrosamine (DEN), a well-known liver-specific carcinogen
(9);
N-methyl-N-nitrosourea (MNU), which has a multi-site
carcinogenic potential particularly targeting the gastrointestinal
lumen and lung (10);
2,2′-dihydroxy-di-N-propylnitrosamine (DHPN), mainly causing
lung tumors (11);
1,2-dimethylhydrazine (DMH), a genotoxic colon cancer initiator
(12); and
N-butyl-N-(4-hydroxybutyl)nitrosamine (BBN), which is
known to initiate urinary bladder carcinogenesis (13). The rat multi-organ carcinogenesis
bioassay is widely used for the detection of carcinogenesis and the
evaluation of cancer chemopreventive agents (14,15).
This approach is beneficial for assessing the modifying effects of
chemicals on wide-range multi-organ carcinogenesis using a single
experiment within a relatively short experimental period (16).
In the last few years, numerous research results
have shown that N. sativa is able to inhibit different types
of tumors in experimental animals when administered as ground seeds
(17), as a crude extract (volatile
oil) (1) or as a single ingredient,
thymoquinone, which is the main active constituent of volatile oils
of N. sativa seeds (18,19).
However, there is currently only limited information on the actual
mechanism of action of N. sativa or its active forms.
Previous studies showed that the more significnt components of the
Egyptian N. sativa are 21% protein, 35.5% fat, 6.6% fibre,
5.5% moisture and 3.7% ash, while the remaining constituents
include total carbohydrates, minerals, vitamins and polyphenols
(20). The active ingredients are
mainly thymoquinone (primarily in essential oil), dithymoquinone
(DTQ) (nigellone), thymol (THY) and thymohydroquinone (THQ). Other
components include α-hederin (a triterpene saponin),
monosaccharides (glucose, rhamnose, xylose and arabinose) and
unsaturated fatty acids (linoleic acid and oleic acids) (21,22).
The biological activities, particularly the antioxidant or
antitumor activities, of thymoquinone and nigellamines identified
from the Egyptian N. sativa (23) or alkaloids (24) have yet to be adequately
determined.
Research studies have shown that the mechanism of
action of N. sativa may correlate with antioxidant
properties of the active ingredients found in its volatile oil such
as thymoquinone, or due to its immunosuppressive and cytotoxic
effects (25). Extracts of N.
sativa are known to exert insulin-sensitizing actions and
lipid-modulating properties (26).
These modulating effects extend to cholesterol, triglyceride and
prostaglandin levels in rats (27).
Furthermore, the same active constituents of N. sativa are
known to affect tumor insulin growth factors through the
suppression of kinase signaling pathways (28). In the present study,
bromodeoxyuridine (BrdU) labeling indices (LIs) were detected
immunohistochemically to assess the population of cells actively
synthesizing DNA (29). BrdU, a
thymidine analog which replaces (3H) thymidine (30), is incorporated into the newly
synthesized DNA strands of actively proliferating cells following
partial denaturation of double-stranded DNA. Measurement of
(3H) thymidine incorporation as cells enter the S phase
of the cell cycle is the traditional method for the detection of
cell proliferation, particularly in tumors (31). Evaluation of cell cycle progression
and cellular proliferation is essential for investigations in many
scientific fields, including cancer chemoprevention studies.
Therefore, to further elucidate the effects of crude
volatile oils extracted from N. sativa, the present
experiment was designed as a multi-organ carcinogenesis bioassay in
Wistar rats. BrdU LIs as well as plasma levels of insulin growth
factor (IGF-I), triglycerides (TG), prostaglandin E2 (PGE2) and
cholesterol were evaluated.
Materials and methods
Animals
A total of 76 male Wistar rats were obtained at 5
weeks of age from the Helwan Breeding Facility of Helwan
University, Cairo, Egypt. The rats were divided into 5 groups
according to their body weight (b.w.) to minimize inter-group
standard errors (20 rats each for groups 1–3 and 8 each for groups
4 and 5). The animals were housed 4 or 5 per plastic cage with wood
chips for bedding and maintained at a temperature of 22±2°C, with
45±4% relative humidity with a 12-h light/12-h dark cycle. The
experiment commenced after acclimatization for one week to the
animal house conditions. Diet and water were available ad
libitum, and animal growth and food consumption were measured
weekly during the experiment to assess toxicity. The Institutional
Animal Care Facility of the Zoology Department, Faculty of Science,
Tanta University, Egypt approved the experimental design.
Chemicals
DEN, DMH (purity 99%), DHPN and MNU were obtained
from Sigma-Aldrich (St. Louis, MO, USA), while BBN was obtained
from Tokyo Kasei Kogyo Co., Ltd. (Tokyo, Japan). Crude extra virgin
oil extracted from fresh N. sativa seeds was purchased from
Kahira Pharm. and Chem. Co., Cairo, Egypt. The oil was stored and
protected from light exposure in dark glass bottles at room
temperature between 15–20°C. Calculated doses of 1000 and 4000 ppm
of the oil were then mixed fresh (v/v) with basal-powdered diet and
administered ad libitum to the rats. The contents presented
in the basal diet were: crude protein, not less than ~18%; crude
lipid, not less than ~2.6%; crude fiber, not more than ~11.43%;
with an ingestion energy of not more than 284.83 kcal/100 g. The
doses of N. sativa were equivalent, respectively, to 50 and
200 mg/kg b.w. of the oil calculated according to data for food
consumption in our previous experiment (7); the doses were not associated with any
signs of toxicity.
Mid-term multi-organ carcinogenesis
bioassay
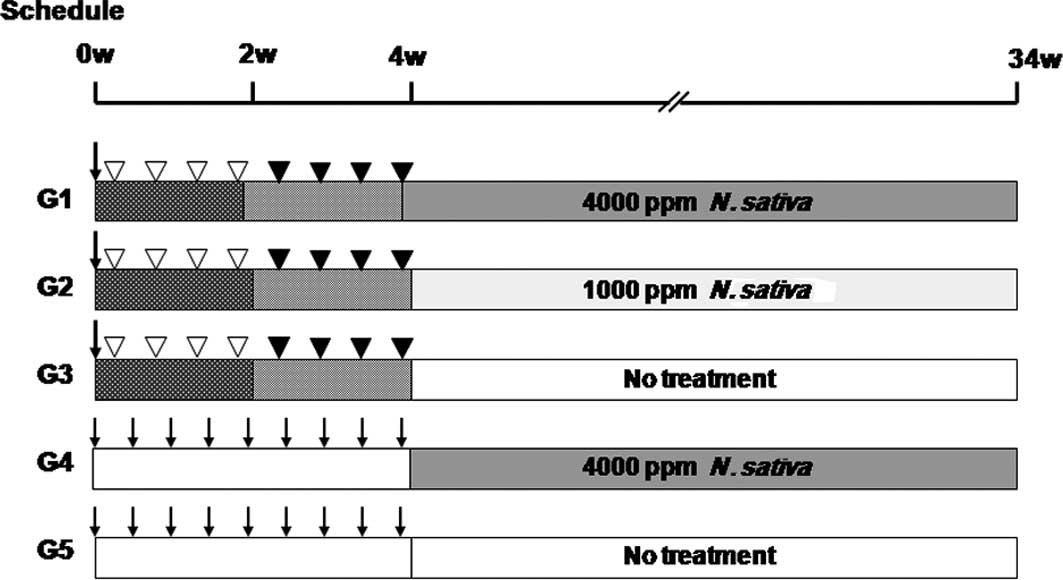
The experimental protocol is shown in Fig. 1. Rats in groups 1–3 were treated
sequentially with DEN [100 mg/kg b.w., intraperitoneally (i.p.),
single dose] at commencement; MNU, (20 mg/kg b.w., i.p.) on days 2,
5, 8 and 11; and DMH, [40 mg/kg b.w., subcutaneously (s.c.)] on
days 14, 17, 20 and 23. Animals were simultaneously given BBN,
(0.05% in the drinking water) during weeks 1 and 2 and DHPN (0.1%
in the drinking water) during weeks 3 and 4. Following this
combination treatment with DEN, MNU, BBN, DMH and DHPN (DMBDD),
animals in groups 1 and 2 were fed a powdered basal diet containing
1000 or 4000 ppm of N. sativa until the end of the
experiment, while animals in group 3 received a basal diet and
served as a carcinogen-treated control for groups 1 and 2. Animals
in groups 4 and 5 did not receive any carcinogen administration.
However, they received 0.9% saline injections instead according to
the same schedule. Animals in group 4 received N. sativa
4000 ppm as a drug treatment control and those of group 5 received
a normal basal diet as a negative control. The animals were
sacrificed under ether anesthesia at week 32 following commencement
of the experiment. A total of 5 rats from each of groups 1–3
received a single injection of BrdU (Sigma Chemicals, St. Louis,
MO, USA; 100 mg/kg b.w. i.p.) 1 h prior to sacrifice.
Biochemical analysis of blood
To assess the serum biochemical levels, blood
samples were obtained at the time of autopsy from the inferior vena
cava in tubes containing no anti-coagulant. The blood was
centrifuged at 4000 rpm for 30 min at 4°C until the serum was
obtained. Blood was collected from 10 rats of groups 1–3 and from 6
rats from each of groups 4 and 5, that had not received BrdU
injections.
Serum was collected and preserved frozen at −80°C
before measurement of different biochemical parameters. To detect
serum levels of IGF-I protein, 3-ml aliquots of the supernatant
were subjected to an immunoradiometric assay with a kit (Somatmedim
CII, Bayer) according to a previously described method (32). To determine the serum PGE2 levels, 1
ml of serum supernatant was mixed with an equal volume of 1 M
phosphate buffer (pH 6.8) containing methanol and
(3H)PGE2 (4000 d.p.m). Following mixing and
centrifugation, the samples were processed and measured for PGE2 by
radioimmunoassay (using a kit from NEN, Boston, MA, USA). Recovery
by this method was satisfactory (~80%). Total cholesterol and TG
concentrations in the serum were determined enzymatically using
commercially available reagent kits (assay kits for the TDX system;
Abbott Laboratory, Irving, TX, USA). Briefly, after the blood was
centrifuged to obtain serum, the sample was analyzed for total
cholesterol, total TG, HDL- and LDL-cholesterol levels by
precipitation techniques and modified enzymatic procedures as
previously described by Bucolo and David (33) and Allain et al (34). Total-cholesterol levels were
calculated using the Friedewald equation (35).
Tissue processing
All of the surviving animals were sacrificed under
ether anaesthesia, and all of the major organs were excised and
fixed in 10% phosphate-buffered formalin. The fixative was injected
into the head, the lumina of the alimentary canal and the urinary
bladder. The liver, kidneys, spleen and testes were excised and
quickly weighed. The organs were then wet cut into 3-mm sections,
and three liver slices (one each from the anterior, right posterior
and caudate lobes) were prepared, and fixed in the same fixative.
The alimentary tract from the esophagus to the rectum, the urinary
bladder and lungs with the trachea and thyroid gland were excised
and fixed. All of the alimentary canal areas were cut open along
the median axis from the esophagus to the anal end and were further
fixed flat between two sheets of filter paper in formalin. Other
organs, such as the pituitary glands, adrenal glands, thymus,
salivary glands and lymph nodes, were also excised, fixed in
formalin and prepared for histopathological examination. Body areas
were assessed for the presence of any treatment-related
abnormalities or lesions. Sketches were made for all of the tumors
found at any site to determine location. Tumor size (length, width
and height) was measured using calipers.
Histopathological evaluation
The liver, spleen, kidneys, lungs, testes, as well
as selected samples from all of the collected organs, lesions and
tumors were routinely embedded in paraffin, sectioned at 3-4 μm,
and prepared for histological examination following staining with
hematoxylin and eosin. The lungs were inflated with 10%
phosphate-buffered formalin. At least two sections from each lobe
(>10 sections from each rat), including any visible macroscopic
nodular lesions on the surface of the lungs, were embedded in
paraffin and routinely prepared for histopathological examination.
Microscopically, the lung epithelial lesions were classified into
three histological categories: alveolar epithelial hyperplasia,
adenoma and carcinoma. Carcinomas were classified into
adenocarcinomas and squamous cell carcinomas (SCCs).
Histopathological evaluation for the lungs and all other organs was
performed according to Boorman et al (36).
Immunohistochemical (IHC) staining for
BrdU and GST-P-positive foci
The method of Hsu et al (37) was used for the IHC staining in the
present study. For BrdU, after being deparaffinized and hydrated
through a graded ethanol series, 4 μm of the colon, lung, esophagus
and forestomach was used. Briefly, the sections were incubated with
0.3% hydrogen peroxide for 30 min to block endogenous peroxidase
activity. They were then incubated with 10% normal horse serum for
30 min at room temperature, and overnight at 4°C with the BrdU
antibody (dilution 1:500) in Tris(hydroxymethyl)
aminomethane-buffered saline (TBS). Following exposure for 30 min
at room temperature to biotinylated horse anti-mouse IgG (Vector
Laboratories, Burlingame, CA, USA), sections were incubated with
the avidin-biotin complex (ABC) at a 1:25 dilution. Washing with
TBS followed each step. To score cell proliferation, cells stained
for BrdU expression were counted in at least 2000 cells from each
organ. To generate the BrdU labeling index (LI, %), the total
number of proliferating cells was divided by the total number of
cells × 100.
Regarding IHC staining of GST-P-positive foci in the
liver, after incubation with 0.3% hydrogen peroxide, sections were
treated sequentially with 10% normal goat serum for 40 min and
incubated with anti-rabbit GST-P polyclonal antibody (Medical &
Biological Lab. Co. Ltd., Nagoya, Japan) at a dilution of 1:1000
overnight at 4°C. The sections were then treated with
biotin-labeled goat anti-rabbit IgG and the
avidin-biotin-peroxidase complex (ABC kit; Vector Laboratories).
For both BrdU- and GST-P-positive foci staining, peroxidase
activity was visualized by treatment with a 0.02% solution of
diaminobenzidine tetrahydrochloride containing 0.05% hydrogen
peroxide. The nuclei were counterstained with hematoxylin. The
numbers and areas of GST-P-positive cell foci >0.2 mm in
diameter and the total areas of the liver sections were measured
with the aid of the color video image analyzer (Qwin Image Analysis
System, Leica, Wetzlar, Germany).
Statistical analyses
The significance of differences between group mean
values for body weight, food intake, tumor size, tumor multiplicity
and immunohistochemistry data were subjected to SuperANOVA Duncan
New Multiple Range analysis (StatView v.4.5; Abacus Concepts Inc.,
Berkeley, CA, USA). Data for tumor incidence were analyzed
according to Fisher's exact probability test or the Chi-squared
test (StatView, ver. 5, SAS Institute Inc., Cary, NC, USA).
Statistical significance was estimated at P<0.05.
Results
General observations, body and organ
weights
In the DMBDD-treated groups (1–3), one
rat from group 1, two rats from group 2 and one rat from group 3
were moribund before the end point of the scheduled sacrifice.
Therefore, their data were not included in the study. The growth
rates of rats in all of the groups (Fig. 2) were constant during the experiment
with no differences among the DMBDD-administered groups 1–3 or
between groups 4 and 5. The final body weights in groups 1–3 were
less than those of groups 4 and 5, while the relative organ
weights, except for those of the testes, showed an increase in the
DMBDD-treated groups as compared to the rats in groups 4 and 5. The
mean relative right kidney weights of the rats in group 2 were
essentially similar to those of the non-carcinogen-treated rats. No
other significant inter-group variation was observed between the
comparable groups (Table I).
 | Table IData for final body (g) and relative
organ weights (%). |
Table I
Data for final body (g) and relative
organ weights (%).
| Group | Treatment | Effective no. of
rats | Final body weight
(%) | Relative liver
weight (%) | Relative kidney
weight (%) | Relative spleen
weight (g)a | Relative testis
weight (%) |
|---|
|
|---|
| R | L |
|---|
| 1 | DMBDD + 4000
ppm | 19 | 324.0±13 | 2.8 | 0.53 | 0.36 | 0.26 | 0.75 |
| 2 | DMBDD + 1000
ppm | 18 | 324.6±9 | 2.9 | 0.31b | 0.41 | 0.26 | 0.78 |
| 3 | DMBDD only | 19 | 326.9±12 | 3.0 | 0.51 | 0.42 | 0.30 | 0.78 |
| 4 | 4000 ppm only | 8 | 435.4±11 | 2.6 | 0.25 | 0.24 | 0.18 | 0.78 |
| 5 | No treatment | 8 | 429.6±11 | 2.6 | 0.23 | 0.25 | 0.19 | 0.79 |
Food consumption and N. sativa
intake
Food consumption values (Table II) in the DMBDD-initiated groups
were lower than those in the groups without initiation (4 and 5),
while water intake was similar among the groups. No inter-group
variation within the DMBDD-treated groups or between groups 4 and 5
was noted. The total (mg/experimental period) and average intake of
N. Sativa per rat (mg/rat/day) were found to be consistent
with the administration dose among the treated groups (Table II).
| Table IIAverage food consumption, water and
N. Sativa intake. |
Table II
Average food consumption, water and
N. Sativa intake.
| | | | N. sativa
intake |
|---|
| | | |
|
|---|
| Group | Treatment | Food consumption
(g/rat/day)a | Water intake
(ml/rat/day) | Total (mg) | Average
(mg/rat/day) |
|---|
| 1 | DMBDD + 4000
ppm | 12.1±5.3 | 17.3±2.3 | 10,164 | 48.4±9.2 |
| 2 | DMBDD + 1000
ppm | 12.6±6.8 | 18.2±3.3 | 2,646 | 12.6±7.5 |
| 3 | DMBDD only | 12.4±5.1 | 17.4±3.9 | 0 | 0 |
| 4 | 4000 ppm only | 14.1±6.7 | 18.9±3.4 | 11,844 | 56.4±9.5 |
| 5 | No treatment | 13.7±5.2 | 19.1±3.3 | 0 | 0 |
Histopathological examination and tumor
occurrence
Preneoplastic or neoplastic lesions were observed in
the esophagus, stomach, small intestine, colon, lungs, liver,
kidneys, urinary bladder, prostate, spleen and skin/subcutis.
Significant inter-group differences in the incidences and
multiplicities of lesions were found mainly in the colon, lungs,
esophagus and forestomach. No tumors were found in any rat from the
control groups 4 and 5. In the colon, tumors were mainly adenomas
and adenocarcinomas, with a few signet ring cell carcinomas,
mucinous adenocarcinomas and fibrous histiosarcomas. N.
sativa significantly decreased the incidences and
multiplicities of the colon tumors (Table III). The effect was similar for
both treatment doses. Colon tumor sizes were significantly
decreased by the N. sativa treatment in a dose-dependent
manner (Table IV).
| Table IIIIncidence (%) and multiplicity of the
colon tumors. |
Table III
Incidence (%) and multiplicity of the
colon tumors.
| DMBDD + 4000 ppm
(G1: n=19) | DMBDD + 1000 ppm
(G2: n=18) | DMBDD only (G3:
n=19) |
|---|
| Colon tumors | Incidencea |
Multiplicityb | Incidencea |
Multiplicityb | Incidencea |
Multiplicityb |
|---|
| Adenoma | 1 (5.3)c | 0.05±0.01c | 1 (5.6)c | 0.06±0.01c | 8 (42.1) | 0.40±1.68 |
| Adenocarcinoma | 5 (26.3) | 0.32±0.31c | 4 (22.2)c | 0.28±0.40c | 12 (63.2) | 0.40±1.90 |
| Signet-ring cell
carcinoma | 0 | 0 | 0 | 0 | 1 (5.3) | 0.04±0.01 |
| Mucinous
adenocarcinoma | 0 | 0 | 1 (5.6) | 0.06±0.01 | 1 (5.3) | 0.05±0.01 |
| Fibrous
histiosarcoma | 1 (5.3) | 0.05±0.01 | 0 | 0 | 0 | 0 |
| All tumors | 7 (36.8)c | 0.47±1.2c | 7 (38.8)c | 0.44±0.60c | 15 (79.0) | 1.85±0.1.8 |
| Table IVAverage colon tumor sizes
(mm3) and BrdU labeling indices. |
Table IV
Average colon tumor sizes
(mm3) and BrdU labeling indices.
| | | BrdU labeling
indices (%) |
|---|
| | |
|
|---|
| Group | Treatment | Tumor sizea | Mucosa | Adenoma | Carcinoma |
|---|
| 1 | DMBDD + 4000
ppm | 12.1±11.85b | 8.4b | 9.2b | 9.4b |
| 2 | DMBDD + 1000
ppm | 24.5±18.24 | 8.1b | 9.4b | 10.1b |
| 3 | DMBDD only | 45.6±47.40 | 12.2 | 14.8 | 14.9 |
In the lungs, epithelial lesions were mainly
alveolar epithelial hyperplasias, alveolar cell adenomas, SCCs and
adenocarcinomas. Table V shows data
for incidences and multiplicities of the lesions. In groups 1 and 2
treated with N. sativa after DMBDD, the incidences and
multiplicities of the lung tumors were generally less than those
found in the DMBDD only-treated group 3. Significant values were
obtained in the multiplicities of adenomas and adenocarcinomas in
the 1000-ppm N. sativa-treated group and in the incidences
and multiplicities of the 4000-ppm-treated group. When the
incidences and multiplicities of the tumors (adenoma + carcinomas)
were collectively calculated together, a significant decrease was
evident for the two N. sativa treatment doses as compared to
the corresponding data obtained from group 3. The multiplicities of
lung cancer (data for the two cancer types together; SCCs +
adenocarcinomas) were significantly decreased by the two treatment
doses of N. sativa while the incidences also decreased,
although this did not reach statistical significance (Table V).
| Table VIncidence (%) and multiplicity of the
lung neoplasias. |
Table V
Incidence (%) and multiplicity of the
lung neoplasias.
| DMBDD + 4000 ppm
(Group 1: n=19) | DMBDD + 1000 ppm
(Group 2: n=18) | DMBDD only (Group
3: n=19) |
|---|
| Lung lesions | Incidencea |
Multiplicityb | Incidencea |
Multiplicityb | Incidencea |
Multiplicityb |
|---|
| Adenoma (alveolar
cell type) | 8 (42.1) | 0.61±0.92d | 6 (33.3) | 0.44±1.54d | 12 (63.2) | 1.10±2.90 |
| Adenocarcinoma | 1 (5.3)d | 0.05±0.01d | 2 (11.1) | 0.11±0.24d | 5 (26.3) | 0.40±2.00 |
| Squamous cell
carcinoma | 1 (5.3) | 0.11±0.20 | 0 | 0 | 1 (5.3) | 0.05±0.01 |
| Adenoma +
carcinoma | 8 (42.1)d | 0.47±1.20d | 8 (44.4)d | 0.56±0.60d | 16 (84.2) | 1.45±0.18 |
| Cancer
incidencec | 2 (10.5) | 0.16±0.28d | 2 (11.1) | 0.17±0.24d | 5 (26.3) | 0.45±2.20 |
In the esophagus, the neoplastic lesions found
[papillary or nodular (P/N) hyperplasias, squamous cell papillomas
and SCCs] had lower incidences and decreased numbers in the N.
sativa-treated groups 1 and 2 as compared to the data obtained
for group 3. The incidences and multiplicities of the sum of the
lesions calculated together were significantly decreased by the two
N. sativa treatment doses. Additionally, the multiplicities
of the papillomas and SCCs of the 4000 ppm-treated group were also
significantly inhibited (Table
VI).
| Table VIIncidence (%) and multiplicity of
neoplastic lesions in the esophagus and forestomach. |
Table VI
Incidence (%) and multiplicity of
neoplastic lesions in the esophagus and forestomach.
| DMBDD + 4000 ppm
(Group 1: n=19) | DMBDD + 1000 ppm
(Group 2: n=18) | DMBDD only (Group
3: n=19) |
|---|
| Incidencea |
Multiplicityb | Incidencea |
Multiplicityb | Incidencea |
Multiplicityb |
|---|
| Esophagus |
| P/N
hyperplasia | 5 (26.3) | 0.37±0.6 | 5 (27.8) | 0.28±0.50 | 9 (47.4) | 0.45±0.5 |
| Papilloma | 5 (26.3) | 0.26±0.5c | 4 (22.2) | 0.28±0.58 | 9 (47.4) | 0.60±0.8 |
| Squamous cell
carcinoma | 3 (15.8) | 0.16±0.4c | 4 (22.2) | 0.22±0.40 | 7 (36.8) | 0.40±0.6 |
| All lesions | 7 (36.8)c | 0.42±0.9c | 8 (44.4)c | 0.51±1.1c | 16 (84.2) | 1.00±0.9 |
| Forestomach |
| P/N
hyperplasia | 2 (10.5)c | 0.16±0.4c | 8 (44.4) | 0.28±0.5 | 8 (42.1) | 0.45±0.5 |
| Papilloma | 2 (10.5)c | 0.32±0.1c | 5 (27.8) | 0.28±0.6c | 9 (47.4) | 0.60±0.8 |
| Squamous cell
carcinoma | 2 (10.5)c | 0.26±0.5 | 4 (22.2) | 0.22±0.4 | 8 (42.1) | 0.45±0.6 |
| All lesions | 4 (21.1)c | 0.50±0.7c | 9 (50.0) | 0.20±0.7 | 15 (79.0) | 1.10±0.6 |
In the forestomach, a dose-dependent inhibition of
tumor incidences and multiplicities was observed. The majority of
the lesions were significantly inhibited by the 4000 ppm dose,
while for the 1000 ppm-treated dose, significance was only noted
for the multiplicity of papillomas (Table VI). Data for the incidences and
multiplicities of the tumors found in the other organs of groups 1,
2 and 3 are shown in Table VII.
None of the data showed any significant inter-group
differences.
| Table VIITumor incidences in other organs. |
Table VII
Tumor incidences in other organs.
| DMBDD + 4000 ppm
(G1: n=19) | DMBDD + 1000 ppm
(G2: n=18) | DMBDD only (G3:
n=19) |
|---|
| Spleen |
| Malig.
lymphoma/leukemia | 1 (5.3) | 1 (5.6) | 3 (15.8) |
|
Hemangiosarcoma | 0 | 0 | 1 (5.3) |
| Metastatic
fibrosarcoma | 0 | 1 (5.6) | 4 (21.0) |
| All tumor
incidences | 1 (5.3) | 2 (11.1) | 4 (21.0) |
| Kidney(s) |
|
Nephroblastoma | 13 (68.4) | 9 (50.0) | 14 (73.7) |
| Renal cystic
adenoma | 1 (5.3) | 0 | 0 |
| Renal cell
adenoma | 0 | 0 | 5 (26.3) |
| Transitional cell
adenoma | 1 (5.3) | 0 | 0 |
| Renal cell
carcinoma | 2 (10.5) | 3 (16.7) | 6 (31.6) |
| Malignant
fibrosarcoma | 1 (5.3) | 0 | 1 (5.3) |
| All tumor
incidences | 17 (89.5) | 12 (66.6) | 17 (89.5) |
| Urinary
bladder |
| P/N
hyperplasia | 8 (42.1) | 11 (61.1) | 9 (47.4) |
| Paoilloma | 0 | 4 (22.2) | 3 (15.8) |
| TCC | 1 (5.3) | 0 | 2 (10.5) |
| Squamous cell
carcinoma | 1 (5.3) | 1 (5.6) | 0 |
| All tumor
incidences | 2 (10.5) | 5 (27.8) | 5 (26.3) |
| Prostate |
| Adenoma | 0 | 1 (5.6) | 1 (5.3) |
| Skin/subcutis |
| Squamous cell
carcinoma | 0 | 1 (5.6) | 0 |
| Small
intestine |
| Adenoma | 2 (10.5) | 1 (5.6) | 1 (5.3) |
|
Adenocarcinoma | 5 (26.3) | 3 (16.7) | 7 (36.8) |
BrdU LIs
The IHC examination of the lesions in the colons,
lungs, esophagus and forestomach showed lower BrdU LIs in the N.
sativa-treated groups when compared to the DMBDD only-treated
group 3. In the colon, (Table IV)
administration of N. sativa (groups 1 and 2) significantly
reduced the BrdU LIs in the mucosal crypts, adenomas and
adenocarcinomas as compared with the group 3 values (P<0.01).
The inhibition was similar for the two doses of N. sativa.
In the lungs, esophagus and forestomach, the values for the BrdU
LIs were significantly reduced in lesions of the rats treated with
4000 ppm when compared to the values of the DMBDD only-treated
group 3 (Table VIII). However,
for the 1000 ppm-treated groups, a significant decrease in the BrdU
LIs was more evident in the forestomach as compared to the other
two organs (Table VIII).
| Table VIIIMean BrdU labeling indices (%) in the
lung, esophagus and forestomach neoplasia. |
Table VIII
Mean BrdU labeling indices (%) in the
lung, esophagus and forestomach neoplasia.
| Treatment
group | DMBDD + 4000
ppma | DMBDD + 1000
ppm | DMBDD only |
|---|
| Lung |
| Adenomab | 4.00±3.0d | 5.70±3.7 | 8.90±6.5 |
|
Adenocarcinoma | 6.20±5.1d | 8.10±6.6 | 10.3±7.0 |
| Esophagus |
| Mucosac | 7.30±3.0d | 7.10±4.3d | 10.5±6.0 |
| Papilloma | 7.10±5.1d | 9.00±4.3 | 11.7±7.1 |
| Squamous cell
carcinoma | 9.80±5.1 | 11.9±4.3 | 14.2±7.1 |
| Forestomach |
| Mucosa | 6.40±4.1d | 9.82±3.5 | 11.7±8.9 |
| Papilloma | 10.6±7.2d | 9.80±6.2d | 16.0±6.5 |
| Squamous cell
carcinoma | 12.2±6.8d | 11.9±9.0d | 18.7±7.0 |
GST-P-positive foci in the liver
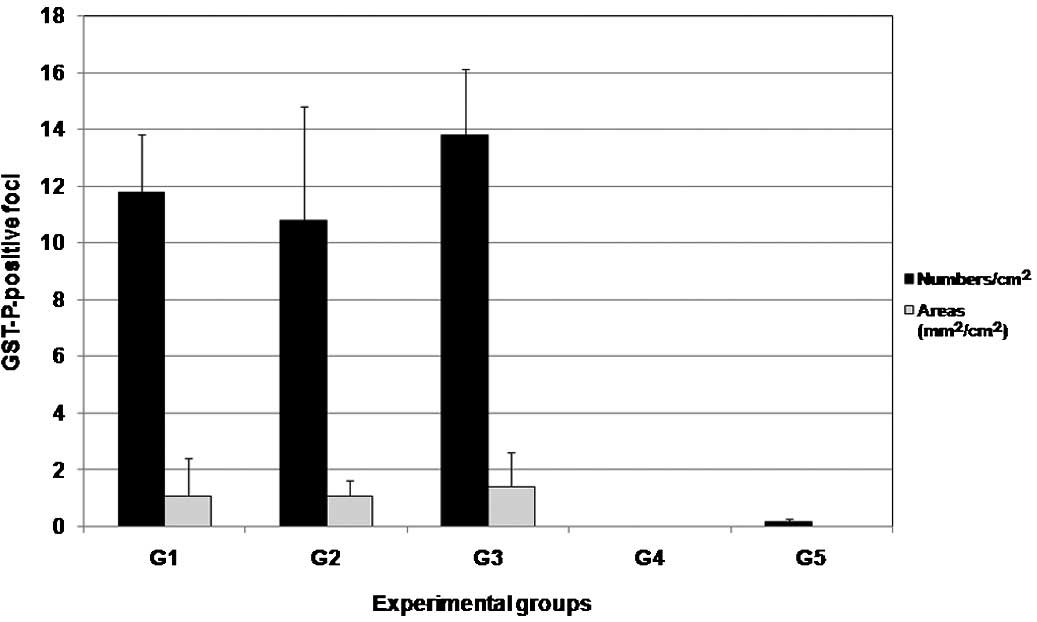
Results of the quantitative evaluation of
GST-P-positive foci in the livers of the DMBDD-initiated groups are
shown in Fig. 3. In the N.
sativa-treated groups (1 and 2), the numbers and areas of
GST-P-positive foci were fewer than those found in the DMBDD
only-treated group 3, albeit without statistical significance. In
group 5, IHC staining revealed a positive GST-P focus in one
rat.
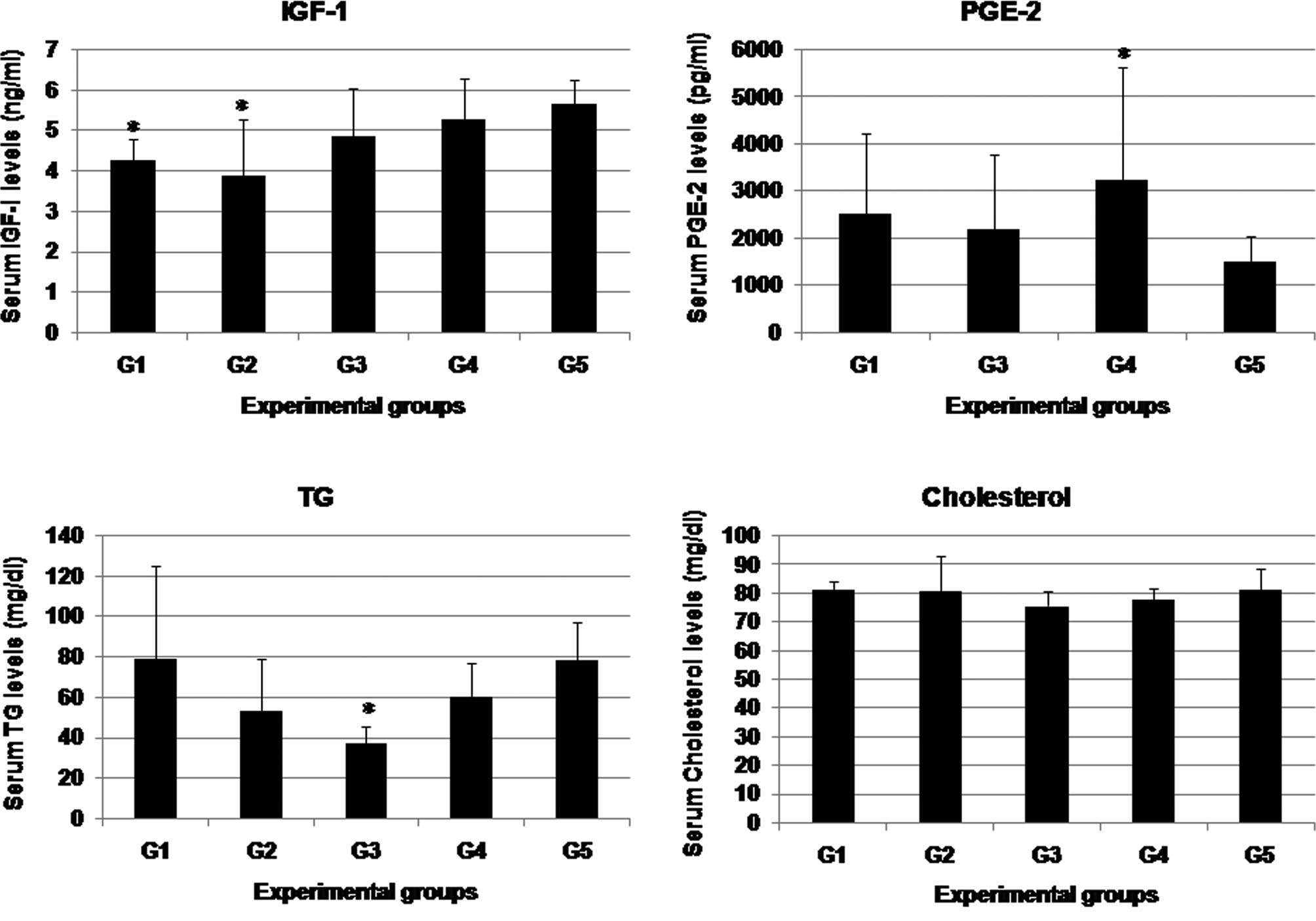
Serum biochemistry
Fig. 4 shows the
serum biochemical levels of IGF-I, PGE2, TG and cholesterol in the
studied groups. IGF-I levels of groups 1–3 were generally lower
than those of control groups 4 and 5. N. sativa treatment in
groups 1 and 2 significantly decreased IGF-I levels to below the
control levels. No differences in IGF-I levels were found between
animals treated with N. sativa in group 4 and those of group
5. PGE2 levels were evaluated in groups 1, 3, 4 and 5. The N.
sativa administration significantly lowered the PGE2 levels
close to the normal control levels. Additionally, serum TG levels
were significantly restored close to the normal control levels,
particularly in the rats of group 1 treated with 4000 ppm N.
sativa following carcinogen administration. Serum cholesterol
levels did not vary significantly among the studied groups.
Discussion
The present investigation found that two doses of
N. sativa volatile oil, 1000 ppm and 4000 ppm in the diet,
significantly inhibited tumor development in multiple sites of the
rat body. To the best of our knowledge, this is the first study to
simultaneously show the multi-potent effects of the N.
sativa crude extract in various organs. Benign and malignant
colon tumor incidences, multiplicities and sizes, and tumor
incidences and multiplicities in the lungs as well as in different
parts of the alimentary canal, particularly the esophagus and
forestomach, were all significantly reduced by the two doses of
N. sativa.
Previous and recent in vitro and in
vivo studies suggest that constituents of N. sativa
seeds have clear anticancer properties (4,19).
N. sativa volatile oil was found to suppress ferric
nitrilotriacetate-induced two-stage renal carcinogenesis in rats
(1) and reduce tumors and markers
of apoptosis and oxidative stress in
7,12-di-methylbenz(a)anthracene (DMBA)-induced rat mammary
carcinomas (38). N. sativa
volatile oil constituents such as thymoquinone inhibited
20-methylcholanthrene-induced fibrosarcoma tumorigenesis in male
Swiss albino mice, most likely through antioxidant activity and
interference with DNA synthesis, coupled with the enhancement of
detoxification processes (18).
Previously, the post-initiation application of N. sativa oil
was shown to exert potent chemopreventive potential on DMBA-induced
skin carcinogenesis (39) and on
methylcholanthrene-induced soft tissue sarcoma development in
female BALB/c mice (40). More
recently, pre-exposure to the N. sativa extract and
thymoquinone, before gemcitabine or oxaliplatin in vitro,
resulted in a 60–80% growth inhibition of pancreatic cancer cells
compared with 15–25% when gemcitabine or oxaliplatin were used
alone (19).
The results of the present study confirmed previous
results on the chemopreventive potential of N. sativa
volatile oil on DMH-induced colonic aberrant crypt foci (7) and on its modulatory effects on
AOM-induced colon carcinogenesis in a short-term period in rats
(41). Notably, tumor sizes were
decreased dose-dependently by the two doses of N. sativa.
Previously, a combination of bee honey and N. sativa ground
seeds was found to inhibit MNU-induced colon carcinogenesis and
suppress oxidative stress markers such as nitric oxide and
malondialdehyde in vivo (17). The majority of available data on the
chemopreventive effects of N. sativa, and/or its bioactive
extracts, were reported for colon cancer cells in vitro. N.
sativa extracts had significant inhibitory effects on human
colon cancer cells such as HT-29 (42) and HCT-116 (43). When its anticancer effect was
compared with that of 5-fluorouracil (5-FU), the chemotherapeutic
gold-standard for the treatment of colon cancer, the N.
sativa extract had the same effects on colon cancer SW-626
cells as 5-FU but with significantly fewer side effects (44). Gali-Muhtasib et al (43) showed that extracts from N.
sativa affected human colorectal cancer cells by triggering
apoptotic cell death via a p53-dependent mechanism.
In the present study, marked inhibitory effects on
the lung tumors were noted. To the best of our knowledge, this is
the first report on the effect of the chemopreventive
potential of N. sativa on lung tumors in vivo. Even
when subgroup data did not reach significant statistical values
(perhaps due to the relatively small number of tumors studied in
each case), the number of tumors decreased when compared to the
control group 3. Previously, Kumara and Huat (45) showed the antitumor activity of a
column fraction 5 (CC-5) of an ethanolic extract of the N.
sativa seeds against s.c.-implanted LL/2 Lewis lung carcinoma
cells and i.p.-implanted murine P388 leukemia in BDF1 mice at doses
of 200 and 400 mg/kg b.w. The extract yielded significant tumor
inhibition rate values of 70% (P<0.001) and 60% (P<0.001),
respectively. In another study (46), α-hederin and thymoquinone, the two
principal bioactive constituents of N. sativa volatile oil,
separately demonstrated dose- and time-dependent cytotoxic and
apoptotic/necrotic effects on implanted tumors from four human
cancer cell lines [A549 (lung carcinoma), HEp-2 (larynx epidermoid
carcinoma), HT-29 (colon adenocarcinoma) and MIA PaCa-2m
(pancreatic carcinoma)].
Numerous studies have demonstrated various
modulatory effects of N. sativa volatile oils on respiratory
diseases in experimental animals (47–53).
N. sativa reduced the peripheral blood eosinophil count,
IgG1 and IgG2a levels, cytokine profiles and inflammatory cells in
lung tissues in a mouse model of allergic airway inflammation
(47). Thymoquinone affected the
cyclooxygenase expression and PGE2 production of airway
inflammation in mice (48), and
protected against lung injury from exposure to sulfur mustard in
Guinea pigs (49). Furthermore, a
preventive effect of thymoquinone was detected on tracheal
responsiveness and inflammatory cells in the lung lavage of
sensitized Guinea pigs (50). The
seed extract ameliorated lung tissue damage in rats after
experimental pulmonary aspiration (51), and attenuated airway inflammation in
a mouse model of allergic asthma by downregulating leukotriene
biosynthesis (52). In addition,
the N. sativa seed aqueous extract showed prophylactic
effects on respiratory symptoms and pulmonary function tests in
chemical war victims (53).
Therefore, the effects of N. sativa crude oil and its
bioactive constituents on lung cancer warrant further study.
N. sativa was found to exert inhibitory
effects on the tumors that appeared in the epithelia of the
esophagus and forestomach. After a search of the available data in
PubMed, the present study is the first to show a chemopreventive
potential of N. sativa volatile oil on esophageal tumors
in vivo. Notably, in the forestomach, the incidences and
multiplicities of P/N hyperplasias, papillomas, SCCs separately,
and the sum of the lesions were significantly inhibited by N.
sativa treatment. The results are in accordance with those of
Badary et al (54) who found
that thymoquinone inhibited mice forestomach tumor development,
most likely through its antioxidant and anti-inflammatory
activities, coupled with the enhancement of detoxification
processes. The modulatory effects of N. sativa extracts on
gastropathies and ulcers were noted. An aqueous suspension of black
seed significantly prevented gastric ulcer formation induced by
necrotizing agents in pylorus-ligated Shay rats (55). This effect of N. sativa was
shown to be prostaglandin-mediated and/or through its antioxidant
activities. In other studies (56,57),
N. sativa volatile oil and thymoquinone, particularly the
oil, protected gastric mucosa from acute alcohol-induced mucosal
injury through antiperoxidative, antioxidant and antihistaminic
effects. Another study (58) showed
that treatment with N. sativa oil and thymoquinone
normalized the level of gastric lesions induced in male Wistar rats
by ischaemia/reperfusion, which is known to be linked to free
radical formation. On the other hand, N. sativa oil alone
tended to normalize the levels of lactate dehydrogenase, reduced
glutathione and superoxide dismutase in the stomach mucosa which
may be related to the conservation of the gastric mucosal redox
state.
Rat GST-P, which is related to human GST-π in
enzymatic and immunological properties, is used by many
investigators as a reliable marker for preneoplastic lesions, since
it is strongly and specifically expressed in the very early phase
of chemically induced hepatocarcinogenesis, but not in normal
hepatocytes (59). The degree of
induction of GST-P-positive foci and nodules in the present
bioassay system for hepatocarcinogenesis has been proven to
correspond with the incidence of hepatocellular carcinomas observed
in long-term in vivo assays (60,61).
In the present study, the mean number of GST-P-positive foci per
cm2 and the average areas (mm2) per
cm2 were decreased by the two N. sativa-treated
doses, although without statistical significance. Iddamaldeniya
et al (62) previously
demonstrated that the combined effect of N. sativa seeds,
Hemidesmus indicus root bark and Smilax glabra
rhizome resulted in complete inhibition of overt liver tumors or
histopathological changes leading to tumor development such as
GST-P-positive foci in Wistar rats treated with DEN after long-term
(16 months) post-carcinogen treatment. In addition, N.
sativa and thymoquinone were found to protect against
hepatocellular toxicity in vitro through induction of
detoxifying agents including quinone reductase and glutathione
transferase (63). N. sativa
protected against liver toxicity induced by various agents, such as
carbon tetrachloride, by downregulating hepatic cytochrome P450
isozyme mRNA and protein expression, partly due to the
downregulation of NO production and the upregulation of
anti-inflammatory IL-10 in rats (64). The difference between the present
data on the liver and that of Iddamaldeniya et al (62) is due to the difference in the
combination of the chemopreventive agents and the different
protocols used for the two studies. The data of this study showed
no clear inhibitory effects on liver carcinogenesis by N.
sativa that may be explained, in part, by the interference of
different active metabolites and the challenge of different
biochemical functions in the livers of rats in this experiment.
This process thus requires confirmation.
A large number of studies have investigated the
precise mechanism of tumor inhibition by N. sativa volatile
oil. For instance, thymoquinone was found to have antioxidant
effects in animal models (54,58,65).
These antioxidant effects were also thought to protect tissues from
radiation injury (66). It has been
found that polyunsaturated fatty acids constitute 83.7% of the
total fat in black seeds (67),
with linoleic acid (cis-C18:2 ω-6) as the major component (60.8%)
and oleic acid (cis-C18:1 ω-9) constituting 21.9% of total fats. It
is known that ω-3 polyunsaturated fatty acids exhibit inhibitory
effects on various types of tumors in animal experiments, and
epidemiological studies have shown that these fatty acids may exert
a protective influence in humans (68). Tocopherols, phospholipids,
phytosterols and phenols are important natural antioxidants in
crude oils. Processing of vegetable oils may induce the formation
of antioxidants. Previously, N. sativa crude seed oil and
oil fractions showed a strong radical scavenging activity, and this
activity correlated well with the total content of polyunsaturated
fatty acids, unsaponifiables and phospholipids, as well as the
initial peroxide values of crude oil (69,70).
The presence of antioxidants such as tocopherols and polyphenols in
oil extracted from N. sativa seeds may be of significance.
The total tocopherol (α-, β- and γ-tocopherols) content in N.
sativa seeds was found to be ~340 μg/g, while total polyphenols
are present at 1744 μg/g (67).
Naturally occurring plant phenols are widely distributed and a
number of these phenols have been found to protect against chemical
carcinogenesis (71). Certain plant
phenols inhibit the activity of cytochrome P-450-dependent enzymes
that metabolize drugs and carcinogens (72). Therefore, the possibility that
polyphenols in N. sativa oil (67) are involved in the present
chemopreventive findings deserves further investigation.
N. sativa markedly reduced the BrdU LIs in
colonic, lung, esophagus and forestomach lesions. Modulation of
cell proliferation by cancer chemopreventive agents in colon
epithelium is widely considered to be a mechanism underlying
protective actions (73). Thus, the
present inhibitory effects of N. sativa oil on BrdU LIs may
be explained by the antioxidant and antiproliferative effects of
the constituents found in the volatile oil of N. sativa,
particularly polyphenols, polyunsaturated fatty acids and
thymoquinone. The antioxidant effects of N. sativa extracts
ameliorate the hyperproliferative response of cells in rat
two-stage renal carcinogenesis (1).
Eleostearic acid (α-ESA), a conjugated linolenic acid, blocked
ER-negative MDA-MB-231 (MDA-wt) and ER-positive MDA-ERα7 human
breast cancer cell proliferation and induced apoptosis through a
mechanism that may be oxidation-dependent (74). Carotenoids with antioxidant
properties such as astaxanthin decreased colon carcinogenesis,
cellular proliferation and increased levels of lipid peroxidation
(75). Thymoquinone administered to
mice reduced the incidence of stomach tumors (54) via mechanisms that included
inhibition of DNA synthesis and the promotion of apoptosis by
inhibiting cell growth in the G1 phase. It also inhibited cellular
proliferation and induced apoptosis in colon cancer cells by
triggering a p53-dependent mechanism (43). Further investigation of the
relevance of other genetic or biochemical factors, particularly
those related to induction of apoptosis, may be of great
significance.
Blood analysis data in the present study showed that
N. sativa restored IGF-I, PGE2 and TG levels in the rats of
groups 1 and 2 to almost control levels. Insulin growth factor I
(IGF-I) is mainly related to kinase signaling and tumor progression
(28). Prostaglandins are potent
proinflammatory mediators generated through arachidonic acid
metabolism by cyclooxygenase (COX)-1 and -2 in response to
different stimuli, and play important roles in modulating
inflammatory responses under a number of conditions. They are
highly correlated to the progression of different types of tumors
(76). The active constituents of
N. sativa have been found to exhibit anti-inflammatory
effects throughout the allergic response in the lung by inhibition
of PGE2 synthesis and attenuation of immune responses (48). LDL and HDL were not measured
separately in the present study but triglyceride levels were lowest
in the DMBDD-only group and N. sativa restored these levels
almost to control values. Lower LDL cholesterol levels and total TG
are known to correlate well with favorable survival of cancer
patients (77). Time-dependent
measurements of TG, LDL and HDL should be performed in a similar
multi-organ carcinogenesis protocol in the future. Malignant cells
maximize their ability to acquire and metabolize nutrients
necessary for their rapid growth and progression by fine-tuning a
double (lipogenic/lipolytic)-edged metabolic sword in a manner
conducive to proliferation as opposed to efficient ATP production
(78). Thus, the present inhibitory
effects of N. sativa on various tumors may correlate partly
or indirectly to the three studied hematological factors. Studies
on the effects of N. sativa on the genes controlling lipid
metabolism, kinase signaling, COX-1 and -2, lipid peroxidation,
glutathione metabolism, growth, cell cycle regulation and apoptosis
are warranted.
Although the exact mechanisms of the
chemopreventive effects of N. sativa oil and its
constituents should be determined more precisely, particularly at
the genetic and molecular levels, the present data indicate a
promising cancer chemopreventive potential. In particular, it
should be stressed that the doses used in the present study exerted
no unfavorable side effects.
Acknowledgements
The author wishes to express his thanks to
Professor Shoji Fukushima, Director of the Japan Bioassay Research
Center, Kanagawa, Japan, for the useful discussions throughout the
work and revision of the final manuscript, and to Professor
Mohammed H. Mona, Head of the Department of Zoology, Faculty of
Science, Tanta University, Egypt for providing the technical
facilities for this experiment.
Abbreviations:
|
ABC
|
avidin biotin complex
|
|
BrdU
|
5-bromo 2′-deoxyuridine
|
|
b.w.
|
body weight
|
|
DMBDD
|
combination of the following 5
carcinogens: diethylnitrosamine (DEN),
N-methyl-N-nitrosourea (MNU),
N-butyl-N-(4-hydroxybutyl)nitrosamine (BBN),
2,2′-dihydroxy-di-N-propylnitrosamine (DHPN) and
1,2-dimethylhydrazine dihydrochloride (DMH)
|
|
GST-P
|
glutathione s transferase-positive
foci
|
|
HDL-cholesterol
|
high density cholesterol
|
|
I.g
|
intragastroluminal gavage
|
|
IgG
|
immunoglobulin G
|
|
IGF-I
|
insulin growth factor 1
|
|
IHC
|
immunohistochemistry
|
|
i.p.
|
intraperitoneal
|
|
LDL-cholesterol
|
low-density cholesterol; N.
sativa, Nigella sativa
|
|
PGE2
|
prostaglandin E2
|
|
ppm
|
part per million
|
|
P/N
|
papillary or nodular hyperplasia
|
|
s.c.
|
subcutaneously
|
|
SCC
|
squamous cell carcinoma
|
|
TBS
|
Tris buffered solution
|
|
TG
|
triglycerides
|
|
v/v
|
volume per volume
|
References
|
1
|
Khan N and Sultana S: Inhibition of two
stage renal carcinogenesis, oxidative damage and hyperproliferative
response by Nigella sativa. Eur J Cancer Prev. 14:159–168.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Farah IO: Assessment of cellular responses
to oxidative stress using MCF-7 breast cancer cells, black seed
(N. Sativa L.) extracts and H2O2. Int
J Environ Res Public Health. 2:411–419. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanafy M and Hatem M: Studies on the
antimicrobial activity of Nigella sativa seed (black cumin).
J Ethnopharmacol. 34:275–278. 1991. View Article : Google Scholar
|
|
4
|
Salomi N, Nair S, Jayawardhanan K,
Varghese D and Panikkar K: Anti-tumor principles from Nigella
sativa seeds. Cancer Lett. 63:41–46. 1992. View Article : Google Scholar
|
|
5
|
Landa P, Marsik P, Havlik J, Kloucek P,
Vanek T and Kokoska L: Evolution of antimicrobial and
anti-inflammatory activities of seed extracts from six
Nigella species. J Med Food. 12:408–415. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suboh SM, Bilto YY and Aburjai TA:
Protective effects of selected medicinal plants against protein
degradation, lipid peroxidation and deformability loss of
oxidatively stressed human erythrocytes. Phytother Res. 18:280–284.
2004. View Article : Google Scholar
|
|
7
|
Salim EI and Fukushima S: Chemopreventive
potential of volatile oil from black cumin (Nigella sativa
L.) seeds against rat colon carcinogenesis. Nutr Cancer.
45:195–202. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ito N, Shirai S and Fukushima S:
Medium-term for carcinogens using multiorgan models. In:
Modification of Tumor Development in Rodents. Progress in
Experimental Tumor Research. Ito N and Sugano H: Karger; Basel: pp.
41–57. 1991
|
|
9
|
Ito N, Tsuda H, Tatematsu M, et al:
Enhancing effect of various hepatocarcinogens on induction of
preneoplastic glutathione S-transferase placental form positive
foci in rats – an approach for a new medium-term bioassay system.
Carcinogenesis. 9:387–394. 1988.PubMed/NCBI
|
|
10
|
Kurata Y, Hagiwara A, Tamano S, Shibata M,
Tanaka H, Yamada M, Ito H and Fukushima S: Modifying effects of
beraprost sodium (TRK-100) on N-methyl-N-nitrosourea (MNU)
carcinogenesis in F344 rats. J Toxicol Sci. (Suppl 1): 1–39. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Konishi Y, Yamamoto K, Eimoto H, Tsutsumi
M, Sugimura M, Nii H and Mori Y: Carcinogenic activity of
endogenously synthesized N-nitrosobis(2-hydroxypropyl)amine in
rats. IARC Sci Publ. 105:318–321. 1991.PubMed/NCBI
|
|
12
|
Bird RP: Observation and quantification of
aberrant crypts in the murine colon treated with a colon
carcinogen: preliminary findings. Cancer Lett. 37:147–151. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salim EI, Wanibuchi H, Morimura K, Kim S,
Yano Y, Yamamoto S and Fukushima S: Inhibitory effects of
1,3-diaminopropane, an ornithine decarboxylase inhibitor, on rat
two-stage urinary bladder carcinogenesis initiated by
N-butyl-N-(4-hydroxybutyl)nitrosamine. Carcinogenesis. 21:195–203.
2000. View Article : Google Scholar
|
|
14
|
Spinardi-Barbisan AL, Kaneno R, Rodrigues
MA, Fávero Salvadori DM, Trindade Moreira EL, Barbisan LF and Viana
de Camargo JL: Lymphoproliferative response and T lymphocyte
subsets in a medium-term multiorgan bioassay of carcinogenesis in
Wistar rats. Cancer Lett. 154:121–129. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Doi K, Sakai K, Tanaka R, Toma K,
Yamaguchi T, Wei M, Fukushima S and Wanibuchi H: Chemopreventive
effects of 13alpha, 14alpha-epoxy-3beta-methoxyserratan-21beta-ol
(PJJ-34), a serratane-type triterpenoid, in a rat multi-organ
carcinogenesis bioassay. Cancer Lett. 289:161–169. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hagiwara A, Tanaka H, Imaida K, Tamano S,
Fukushima S and Ito N: Correlation between medium-term multi-organ
carcinogenesis bioassay data and long-term observation results in
rats. Jpn J Cancer Res. 84:237–245. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mabrouk GM, Moselhy SS, Zohny SF, Ali EM,
Helal TE, Amin AA and Khalifa AA: Inhibition of methylnitrosourea
(MNU) induced oxidative stress and carcinogenesis by orally
administered bee honey and Nigella grains in Sprague Dawely
rats. J Exp Clin Cancer Res. 21:341–346. 2002.PubMed/NCBI
|
|
18
|
Badary OA and Gamal El-Din AM: Inhibitory
effects of thymoquinone against 20-methylcholanthrene-induced
fibrosarcoma tumorigenesis. Cancer Detect Prev. 25:362–368.
2001.PubMed/NCBI
|
|
19
|
Banerjee S, Kaseb AO, Wang Z, Kong D,
Mohammed M, Padhye S, Sarkar FH and Mohammed RM: Antitumor activity
of gemcitabine and oxaliplatin is augmented by thymoquinone in
pancreatic cancer. Cancer Res. 69:5575–5583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Babayan VK, Koottungal D and Halaby GA:
Proximate analysis, fatty acid, and amino acid composition of
Nigella sativa L. seeds. J Food Sci. 43:1314–1319. 1978.
View Article : Google Scholar
|
|
21
|
Ali BH and Blunden G: Pharmacological and
toxicological properties of Nigella sativa. Phytother Res.
17:299–305. 2003. View Article : Google Scholar
|
|
22
|
Ghosheh OA, Houdi AA and Crooks PA: High
performance liquid chromatographic analysis of the
pharmacologically active quinines and related compounds in the oil
of the black seed (Nigella sativa L.). J Pharm Biomed Anal.
19:757–762. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morikawa T, Xu F, Ninomiya K, Matsuda H
and Yoshikawa M: Nigellamines A3, A4, A5, and C, new
dolabellane-type diterpene alkaloids, with lipid
metabolism-promoting activities from the Egyptian medicinal food
black cumin. Chem Pharm Bull. 52:494–497. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu YM, Yang JS and Liu QH: A new alkaloid
and its artificial derivative with an indazole ring from Nigella
glandulifera. Chem Pharm Bull. 52:454–455. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Islam SN, Begum P, Ahsan T, Huque S and
Ahsan M: Immunosuppressive and cytotoxic properties of Nigella
sativa. Phytother Res. 18:395–398. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Le PM, Benhaddou-Andaloussi A, Elimadi A,
Settaf A, Cherrah Y and Haddad PS: The petroleum ether extract of
Nigella sativa exerts lipid-lowering and insulin-sensitizing
actions in the rat. J Ethnopharmacol. 94:251–259. 2004.PubMed/NCBI
|
|
27
|
Kocyigit Y, Atamer Y and Uysal E: The
effect of dietary supplementation of Nigella sativa L. on
serum lipid profile in rats. Saudi Med J. 30:893–896.
2009.PubMed/NCBI
|
|
28
|
Yi T, Cho SG, Yi Z, Pang X, Rodriguez M,
Wang Y, Sethi G, Aggarwal BB and Liu M: Thymoquinone inhibits tumor
angiogenesis and tumor growth through suppressing AKT and
extracellular signal-regulated kinase signalling pathways. Mol
Cancer Ther. 7:1789–1796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chakraborty T, Chatterjee A, Rana A,
Srivastawa S, Damodaran S and Chatterjee M: Cell proliferation and
hepatocarcinogenesis in rat initiated by diethylnitrosamine and
promoted by phenobarbital: potential roles of early DNA damage and
liver metallothionein expression. Life Sci. 81:489–499. 2007.
View Article : Google Scholar
|
|
30
|
Gratzner HG: Monoclonal antibody to
5-bromo-2 and 5-iododeoxyuridine: a new reagent for detection of
DNA replication. Science. 218:474–475. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McGinley JN, Knott KK and Thompson HJ:
Effect of fixation and epitope retrieval on BrdU indices in mammary
carcinomas. J Histochem Cytochem. 48:355–362. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rosenfeld RG, Wilson DM, Lee PD and Hintz
RL: Insulin-like growth factors I and II in evaluation of growth
retardation. J Pediatr. 109:428–433. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bucolo G and David H: Quantitative
determination of serum triglycerides by the use of enzymes. Clin
Chem. 19:476–482. 1973.PubMed/NCBI
|
|
34
|
Allain CC, Poon LS, Chan CS, Richmond W
and Fu PC: Enzymatic determination of total serum cholesterol. Clin
Chem. 20:470–475. 1974.PubMed/NCBI
|
|
35
|
Wasan KM, Holtorf L, Subramanian R,
Cassidy SM, Pritchard PH, Stewart DJ, Novak E and Moghadasian MH:
Assessing plasma pharmacokinetics of cholesterol following oral
co-administration with a novel vegetable stanol mixture to fasting
rats. J Pharm Sci. 90:23–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Boorman GA, Eustis SL, Elwell MR,
Montgomry CA and MacKenzie WF: Pathology of the Fischer Rat
Reference & Atlas. Academic Press; San Diego: 1990
|
|
37
|
Hsu SM, Raine L and Fanger H: Use of
avidin-biotin-peroxidase complex (ABC) in immunoperoxidase
techniques: a comparison between ABC and unlabeled antibody (PAP)
procedures. J Histochem Cytochem. 29:577–580. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
El-Aziz MA, Hassan HA, Mohamed MH, Meki
AR, Abdel-Ghaffar SK and Hussein MR: The biochemical and
morphological alterations following administration of melatonin,
retinoic acid and Nigella sativa in mammary carcinoma: an
animal model. Int J Exp Pathol. 86:383–396. 2005.PubMed/NCBI
|
|
39
|
Salomi MJ, Nair SC and Pannikar KR:
Inhibitory effects of Nigella sativa and Saffron (Crocus
sativus) on chemical carcinogenesis in mice. Nutr Cancer.
16:67–72. 1991.
|
|
40
|
El-Mofty MM, Abdel-Galil AM, Shwaireb MH,
et al: Prevention of skin tumors induced by
7,12-dimethylbenz(a)anthracene in mice by black seed oil. Oncol
Rep. 4:139–141. 1997.PubMed/NCBI
|
|
41
|
Al-Johar D, Shinwari N, Arif J, Al-Sanea
N, Jabbar AA, El-Sayed R, Mashhour A, Billedo G, El-Doush I and
Al-Saleh I: Role of Nigella sativa and a number of its
antioxidant constituents towards azoxymethane-induced genotoxic
effects and colon cancer in rats. Phytother Res. 22:1311–1323.
2008.
|
|
42
|
Rooney S and Ryan MF: Effects of
alpha-hederin and thymoquinone, constituents of Nigella
sativa, on human cancer cell lines. Anticancer Res.
25:2199–2204. 2005.PubMed/NCBI
|
|
43
|
Gali-Muhtasib H, Diab-Assaf M, Boltze C,
Al-Hmaira J, Hartig R, Roessner A and Schneider-Stock R:
Thymoquinone extracted from black seed triggers apoptotic cell
death in human colorectal cancer cells via a p53-dependent
mechanism. Int J Oncol. 25:857–866. 2004.PubMed/NCBI
|
|
44
|
Norwood AA, Tan M, May M, Tucci M and
Benghuzzi H: Comparison of potential chemotherapeutic agents,
5-fluoruracil, green tea, and thymoquinone, on colon cancer cells.
Biomed Sci Instrum. 42:350–356. 2006.PubMed/NCBI
|
|
45
|
Kumara SS and Huat BT: Extraction,
isolation and characterisation of antitumor principle,
alpha-hederin, from the seeds of Nigella sativa. Planta Med.
67:29–32. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Farah N, Benghuzzi H, Tucci M and Cason Z:
The effects of isolated antioxidants from black seed on the
cellular metabolism of A549 cells. Biomed Sci Instrum. 41:211–216.
2005.PubMed/NCBI
|
|
47
|
Abbas AT, Abdel-Aziz MM, Zalata KR and
el-D Abd Al-Galel T: Effect of dexamethasone and Nigella
sativa on peripheral blood eosinophil count, IgG1 and IgG2a,
cytokine profiles and lung inflammation in a murine model of
allergic asthma. Egyp J Immunol. 12:95–102. 2005.
|
|
48
|
El Mezayen R, El Gazzar M, Nicolls MR,
Marecki JC, Dreskin SC and Nomiyama H: Effect of thymoquinone on
cyclooxygenase expression and prostaglandin production in a mouse
model of allergic airway inflammation. Immunol Lett. 106:72–81.
2006.PubMed/NCBI
|
|
49
|
Hossein BM, Nasim V and Sediqa A: The
protective effect of Nigella sativa on lung injury of sulfur
mustard-exposed Guinea pigs. Exp Lung Res. 34:183–194. 2008.
|
|
50
|
Keyhanmanesh R, Boskabady MH, Eslamizadeh
MJ, Khamneh S and Ebrahimi MA: The effect of thymoquinone, the main
constituent of Nigella sativa, on tracheal responsiveness
and white blood cell count in lung lavage of sensitized Guinea
pigs. Planta Med. 76:2187–222. 2009.
|
|
51
|
Kanter M: Effects of Nigella sativa
seed extract on ameliorating lung tissue damage in rats after
experimental pulmonary aspirations. Acta Histochem. 111:393–403.
2009.
|
|
52
|
El Gazzar M, El Mezayen R, Nicolls MR,
Marecki JC and Dreskin SC: Downregulation of leukotriene
biosynthesis by thymoquinone attenuates airway inflammation in a
mouse model of allergic asthma. Biochim Biophys Acta.
1760:1088–1095. 2006.PubMed/NCBI
|
|
53
|
Boskabady MH and Farhadi J: The possible
prophylactic effect of Nigella sativa seed aqueous extract
on respiratory symptoms and pulmonary function tests on chemical
war victims: a randomized, double-blind, placebo-controlled trial.
J Altern Complement Med. 14:1137–1144. 2008.
|
|
54
|
Badary OA, Al-Shabanah OA, Nagi MN,
Al-Rikabi AC and Elmazar MM: Inhibition of benzo(a)pyrene-induced
forestomach carcinogenesis in mice by thymoquinone. Eur J Cancer
Prev. 8:435–440. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Al Mofleh IA, Alhaider AA, Mossa JS,
Al-Sohaibani MO, Al-Yahya MA, Rafatullah S and Shaik SA:
Gastroprotective effect of an aqueous suspension of black cumin
Nigella sativa on necrotizing agent-induced gastric injury
in experimental animals. Saudi J Gastroenterol. 14:128–134.
2008.PubMed/NCBI
|
|
56
|
Kanter M, Demir H, Karakaya C and Ozbek H:
Gastroprotective activity of Nigella sativa L oil and its
constituent, thymoquinone, against acute alcohol-induced gastric
mucosal injury in rats. World J Gastroenterol. 11:6662–6666.
2005.
|
|
57
|
Kanter M, Coskun O and Uysal H: The
antioxidative and antihistaminic effect of Nigella sativa
and its major constituent, thymoquinone, on ethanol-induced gastric
mucosal damage. Arch Toxicol. 80:217–224. 2006.PubMed/NCBI
|
|
58
|
El-Abhar HS, Abdallah DM and Saleh S:
Gastroprotective activity of Nigella sativa oil and its
constituent, thymoquinone, against gastric mucosal injury induced
by ischaemia/reperfusion in rats. J Ethnopharmacol. 84:251–258.
2003.
|
|
59
|
Morimura S, Susuki T, Hochi S, Yuki A,
Nomura K and Kitagawa T: Transactivation of GST-P gene during
hepatocarcinogenesis of rat. Proc Natl Acad Sci USA. 90:2065–2068.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ogiso T, Tatematsu M, Tamano S, Tsuda H
and Ito N: Comparison of dose dependent effects of chemical
carcinogens on induction of glutathione S-transferase placental
form positive foci in a short term assay and of hepatocellular
carcinomas in a long-term assay. Toxicol Pathol. 13:257–265. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ogiso T, Tatematsu M, Tamano S, Hasegawa R
and Ito N: Correlation between medium-term bioassay system data and
results of long-term testing of rats. Carcinogenesis. 11:561–566.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Iddamaldeniya SS, Thabrew MI,
Wickramasinghe SM, Ratnatunge N and Thammitiyagodage MG: A
long-term investigation of the anti-hepatocarcinogenic potential of
an indigenous medicine comprised of Nigella sativa,
Hemidesmus indicus and Smilax glabra. J Carcinog.
9:5–11. 2006.PubMed/NCBI
|
|
63
|
Nagi MN and Almakki HA: Thymoquinone
supplementation induces quinone reductase and glutathione
transferase in mice liver: possible role in protection against
chemical carcinogenesis and toxicity. Phytother Res. 23:1295–1298.
2009. View Article : Google Scholar
|
|
64
|
Ibrahim ZS, Ishizuka M, Soliman M, ElBohi
K, Sobhy W, Muzandu K, Elkattawy AM, Sakamoto KQ and Fujita S:
Protection by Nigella sativa against carbon
tetrachloride-induced downregulation of hepatic cytochrome P450
isozymes in rats. Jpn J Vet Res. 56:119–128. 2008.
|
|
65
|
Ait Mbarek L, Ait Mouse H, Elabbadi N,
Bensalah M, Gamouh A, Aboufatima R, Benharref A, Chait A, Kamal M,
Dalal A and Zyad A: Anti-tumor properties of black seed (Nigella
sativa L.) extracts. Braz J Med Biol Res. 40:839–847.
2007.PubMed/NCBI
|
|
66
|
Cemek M, Enginar H, Karaca T and Unak P:
In vivo radioprotective effects of Nigella sativa L oil and
reduced glutathione against irradiation-induced oxidative injury
and number of peripheral blood lymphocytes in rats. Photochem
Photobiol. 82:1691–1696. 2006.
|
|
67
|
Nergiz C and Otles S: Chemical composition
of Nigella sativa L. seeds. Food Chem. 48:259–261. 1993.
View Article : Google Scholar
|
|
68
|
Bartsch H, Nair J and Owen RW: Dietary
polyunsaturated fatty acids and cancers of the breast and
colorectum: emerging evidence for their role as risk modifiers.
Carcinogenesis. 20:2209–2218. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Badary OA, Taha RA, Gamal El-Din AM and
Abdel-Wahab MH: Thymoquinone is a potent superoxide anion
scavenger. Drug Chem Toxicol. 26:87–98. 2003. View Article : Google Scholar
|
|
70
|
Ramadan MF, Kroh LW and Mörsel JT: Radical
scavenging activity of black cumin (Nigella sativa L.),
coriander (Coriandrum sativum L.), and niger (Guizotia
abyssinica Cass.) crude seed oils and oil fractions. J Agric
Food Chem. 51:6961–6969. 2003.PubMed/NCBI
|
|
71
|
Wattenberg LW: Chemoprevention of cancer
by naturally occur-ring and synthetic compounds. Cancer
Chemoprevention. Wattenberg L, Lipkin M, Boone CW and Keloff GJ:
CRC Press; Boca Raton: pp. 19–39. 1992
|
|
72
|
Mukhtar H, Das M and Bickers DR:
Inhibition of 3-methyl-cholanthrene-induced skin tumorigenesis in
BALB/c mice by oral feeding of trace amounts of ellagic acid in
drinking water. Cancer Res. 46:2262–2265. 1986.PubMed/NCBI
|
|
73
|
Ames BN, Gold LS and Willet WC: The causes
and prevention of cancer. Proc Natl Acad Sci USA. 92:5258–5265.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Grossmann ME, Mizuno NK, Dammen ML,
Schuster T, Ray A and Cleary MP: Eleostearic acid inhibits breast
cancer proliferation by means of an oxidation-dependent mechanism.
Cancer Prev Res. 2:879–886. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Prabhu PN, Ashokkumar P and Sudhandiran G:
Antioxidative and antiproliferative effects of astaxanthin during
the initiation stages of 1,2-dimethyl hydrazine-induced
experimental colon carcinogenesis. Fundam Clin Pharmacol.
23:225–234. 2009. View Article : Google Scholar
|
|
76
|
Beales IL and Ogunwobi OO: Microsomal
prostaglandin E synthase-1 inhibition blocks proliferation and
enhances apoptosis in oesophageal adenocarcinoma cells without
affecting endothelial prostacyclin production. Int J Cancer.
126:2247–2255. 2010.
|
|
77
|
Abu-Bedair FA, El-Gamal BA, Ibrahim NA and
El-Aaser AA: Serum lipids and tissue DNA content in Egyptian female
breast cancer patients. Jpn J Clin Oncol. 33:278–282. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Menendez JA: Fine-tuning the
lipogenic/lipolytic balance to optimize the metabolic requirements
of cancer cell growth: molecular mechanisms and therapeutic
perspectives. Biochim Biophys Acta. 1801:381–391. 2010. View Article : Google Scholar
|