Introduction
Breast cancer is the most frequent spontaneous
malignancy diagnosed in women in the western world, and is a
classical model of hormone-dependent malignancy. It may have both
its genesis and cell growth influenced by hormonal factors since
approximately one third of breast cancers is responsive to
endocrine therapies (1–5). Environmental chemicals may also be
involved in the etiology of breast cancers (6,7). Many
human tumors have been causally attributed to exposure to
environmental carcinogens, pollutants, pesticides, drugs,
ultraviolet light, radiation and tobacco (6). One of them is malathion [0, 0-dimethyl
S-(1, 2-dicarbethoxy-ethyl)-phosphorodithioate], an
organophosphorous pesticide extensively used to control a wide
range of sucking and chewing pests of field crops. According to
authors (8) exposure of the skin to
this pesticide resulted in systemic absorption. Malathion is
present in lotions and shampoos marketed for the treatment of head
lice and mites in humans. Malathion is acetylcholinesterase (AchE)
inhibitor and responsible for the hydrolysis of body choline
esters, including acetylcholine (Ach) at cholinergic synapses
(9,10). The inhibition of these enzymes
increases the availability of Ach, which in turn can stimulate
cholinergic receptors producing both nicotinic and muscarinic
effects in the organism such as muscle contractions and secretions
in many glands (9). Such effects
can be counteracted with atropine, a parasympatholytic alkaloid
used as an antidote to AchE inhibitors (9,10).
Estrogens have generally been considered beneficial,
based on a variety of hormonal effects and the potential
carcinogenic activity of estrogen-containing medications in humans
has not been recognized for many years. However, in the past 15–20
years, epidemiological studies have pointed to an increased breast
cancer risk associated with prolonged exposure to female hormones,
since early onset of menarche, late menopause, hormone replacement
therapy are associated with greater cancer incidence (9,11–16).
Estrogen administration, a risk factor for humans increases with
continuous doses of estrogen and with the length of treatment
(17). Indeed, slightly elevated
levels of circulating estrogens are also a risk factor for breast
cancer (18,19). This role of endogenous estrogen in
human breast carcinogenesis has been supported by high serum or
urine estrogen levels in breast cancer patients (20,21).
Furthermore, cohort studies have confirmed the strong relationship
between endogenous estrogen levels and breast cancer risk (19,22–25).
Estrogens are associated with carcinogenic events in
both humans and animals (26–31)
and the exact effect of estrogens in breast cancer remains unclear
at this time. Estrogens at physiologic and pharmacological levels
have been implicated causally in a variety of experimental cancers
as shown by the several investigators (15,32).
Since that time, many reports of tumor induction by estrogens have
been published, and many rodent models have been introduced
(33). The evidence for the
carcinogenic activity of estrogens in animals has been deemed
sufficient by the IARC to consider this hormone a carcinogen
(28,29,34).
This conclusion is based on numerous tests of estrogens
administered to rodents orally or subcutaneously. Such mice and
rats increased the incidence of mammary tumors (35–43).
In hamsters, a high incidence of malignant kidney tumors occurred
in intact and castrated males (26,27,44–47)
and in ovariectomized females, but not in intact females (26).
Among the major metabolites of the estrogens are the
catechol estrogens (CE), 2- and 4-hydroxylated estrogens (30). In general, the major metabolites are
the 2-CE, whereas the 4-CE is the minor one. The pathway to give
4-CE is the one leading to the endogenous carcinogenic catechol
estrogen-3, 4-quinones, 4-hydroxyestrone (4-OHE1) and
4-hydroxyestradiol (4-OHE2) are formed from estrone (E1) and
estradiol (E2), which are biochemically interconvertible by the
enzyme 17β-estradiol dehydrogenase. This inactivating pathway is in
competition with the activation of CE to semiquinones and quinones;
the oxidations are catalyzed by peroxidases and cytochrome P450
(CYP). CYP is an important component of the phase I
drug-metabolizing system (30).
Investigations on DNA damage by steroidal estrogens
via catechol estrogen metabolites have received special attention
with the reports that consider the carcinogenic activity of
4-hydroxyestradiol (4-OHE2), comparable to that of estrogen in the
hamster kidney tumor model (30).
The 2-hydroxylation (2-OHE2) of steroidal estrogens is the major
metabolic oxidation of estrogenic hormones in most mammalian
species. Such oxidation is catalyzed in human or hamster liver by
cytochrome P450 3A4 enzymes. Cytochrome P450 1A1 enzymes are the
predominant estrogen 2-hydroxylases in extra hepatic tissues
(30,31). In humans, the predominant conversion
of 17β-estradiol to 4-OHE2 has been detected in benign and
malignant mammary tumors and in normal mammary tissue (30,33).
The human estrogen-4-hydroxylase activity has been identified as
cytochrome P450 1B1, a novel extrahepatic isozyme detected
specifically in mammary tissue, ovary, adrenal gland, uterus and
several other tissues.
Materials and methods
Experimental designs
Thirty-nine-day-old virgin female Sprague-Dawley
rats were obtained from the Catholic University of Chile (Santiago,
Chile) and housed and bred in a barrier animal facility operated in
accordance with the standards outlined in Guide for the Care and
Use of Laboratory Animals (48).
All animals were allowed continuous access to a standard laboratory
chow diet (Champion, Santiago, Chile). Experimental design 1: i)
control group received saline solution, 250 μg/100 g body weight
(bw). Treated animals were injected subcutaneously (s.c.) for 5
days, twice a day with: ii) malathion (FyfanonTM,
Cheminova, Denmark) that received 22 mg/100 g bw, iii)
17β-estradiol (Sigma-Aldrich Chemical Co., Milwaukee), 30 μg/100 g
bw and iv) combination of both (M+E). The animals were sacrificed
in three periods after 30, 124, 240 and 400 days post-treatment.
Experimental design 2: i) control group received saline solution
(s.c.), 250 μg/100 g bw. Treated animals were injected
subcutaneously for 5 days, twice a day with: ii) malathion (M)
(Fyfanon, Cheminova) that received 22 mg/100 g bw, iii) atropine
(Sigma), 250 μg/100 g bw, and iv) combination of malathion and
atropine (M+A); at the dosages previously indicated. The
LD50 values of the substances for malathion was 1,000
mg/kg. However, the dose used in these experiments was 1/6th of the
LD50 for malathion, which allowed a 100% survival of
animals after a 5-day treatment. Animals were housed three per cage
and palpated weekly to detect formation of tumors. The animals were
sacrificed after 10, 20, 30, 60, 90, 124 and 240 days following a
5-day treatment. Tissues were excised and morphologically analyzed.
Animals to be sacrificed were anesthetized by intraperitoneal
injections of sodium pentobarbital (8 mg/100 g bw) and opened by a
midline incision from the pubis to the sub-maxillary area to remove
the tissues. Mammary glands and palpable tumors were fixed in 10%
neutral buffered, embedded in paraffin, then serially sectioned at
a thickness of 5 μm and stained with hematoxylin-eosin. Throughout
histological studies the severity of mammary gland damage was
evaluated by a scoring system which gave quantitative measurements.
Histomorphometry was performed using a binocular microscope, in
which a 1-mm2 grid was installed in one of the oculars.
Ten fields were studied with lens of 10x in an optical microscope
(Olympus CX31). Five slides counted per animal to determine the
number of ducts in stage of proliferation and lobules with hyaline
casts in the mammary gland from control and treated animals. The
damage was analyzed in five sectors per animal and the totality of
fields was analyzed for morphological abnormalities. Zones without
lobules filled with hyaline casts were graded with score = 0. The
damage in fifty lobules was scored according to scale from 10 to 30
points. Structures were graded as 10 when morphology and normal
structure were present and there was lack of hyaline casts in
lobules. Structures were graded as 20 when minor damage lobular
structures were present and there were small hyaline casts inside.
Structures were graded as 30 when and lobular structure had dilated
tubules and there were hyaline casts in larger sizes and lobular
structure had serious injuries and presence of large hyaline casts.
Fifty ducts were classified according to scale from 10 to 30
points. The damage was analyzed in five sectors per animal and the
totality of fields was analyzed for morphological abnormalities.
Ducts without proliferation, i.e. with two to three layers were
graded as 0; ducts with >4 layers and disorganized nuclei were
graded from 10 to 30, as layers increased in size. Comparison
between treated groups and controls were made by Anova and Dunnet’s
test, with P<0.05 and the test of hypothesis for difference of
proportions (P<0.05).
Western blot analysis
Cells were treated with 1 ml lysis buffer (pH 7.2)
(50 mM Tris base; 1 mM EDTA; 100 mM NaCl; 1 mM PMSF; 1 mM
orthovanadate; 0.1% Triton X-100) and centrifuged (10000 rpm × 10
min). The supernatant with cellular proteins was dissolved in
SDS-PAGE sample solution (60 mM Tris, pH 6.5; 10% (w/v) glycerol;
5% (w/v) β-mercaptoethanol; 20% (w/v) SDS; 0.025% (w/v) bromophenol
blue) and denatured by boiling (5 min), and vortex mixing (2×30
seg). The total amount of protein was 20 μg in each lane with
standard protein markers (Bio-Rad Laboratories, Hercules, CA).
After fractionation by SDS-PAGE on gels, proteins were
electro-blotted onto nitrocellulose membrane (Amersham Biosciences,
UK) using a blotting apparatus (Bio-Rad Laboratories). SDS-PAGE
blots were blocked for 2 h in 10% defatted dry milk-TBS-0.1%
Tween-20 and then incubated for 2 h at room temperature with
corresponding primary antibodies (1:200) as CYP1A1 (sc-9828), mp53
(sc-99), c-myc (sc-42) and c-fos (sc-7202) (all from Santa Cruz
Biotechnology, Santa Cruz, CA, USA) and then followed by incubation
with secondary peroxidase-conjugated (1:5000) (all from Santa Cruz
Biotechnology) in 5% defatted dry milk-TBS-0.1% Tween-20. All steps
were performed at room temperature, and blots were rinsed between
incubation steps with TBS-0.1% Tween-20. Cell blots were probed
with mouse anti-β-actin antibody as control. Immunoreactive bands
were visualized by using the ECLTM Western Blotting
Detection Reagent detection method (Amersham, Dübendorf,
Switzerland) and exposure of the membrane to X-ray film. Protein
determination was performed using the bicinchoninic acid method
(Bio-Rad Laboratories) and bovine serum albumin as standard.
Results
Results indicated that estrogen alone increased the
average number of lobules per mm2 of rat mammary glands
in comparison to control and malathion treatment alone at 30, 124,
240 and 400 days after a 5-day treatment. Fig. 1A shows the effect of estrogen,
malathion and combination of both as average number of lobules with
secretion per mm2 of rat mammary glands at 30, 124, 240
and 400 days after a 5-day treatment. Lobular carcinomas derived
from such lobules were found at 240 days of the 5-day treatment of
estradiol. Mammary gland of estrogen-treated animal had altered
lobules full of hyaline casts. The control rats had normal duct
formation. However, the malathion treated animals showed a
significant (P<0.05) increase in the size of ducts in stage of
proliferation (dsp/mm2) of the mammary gland, as well as
in the number of the epithelial layers in comparison to controls at
10 and 20 days after a 5-day treatment, as well as 30, 60, 90, 124
and 240 days (Fig. 1B). Atropine
and combination of both diminished such an effect from 10 to 240
days.
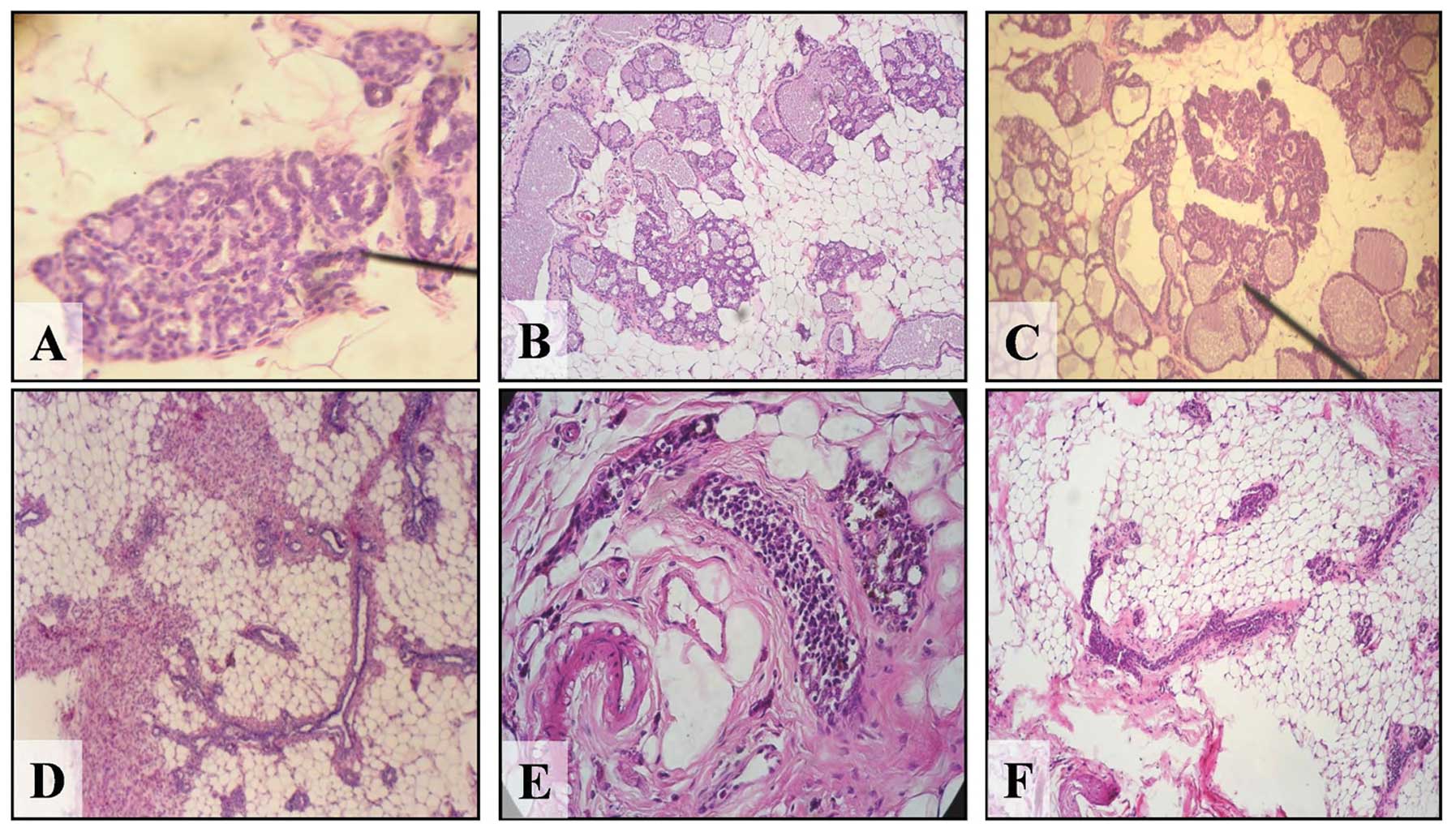
Representative images of cross section of mammary
gland of control and treated animals can be seen in Fig. 2. A normal lobule is shown in
Fig. 2A. Fig. 2B and C represent lobules filled with
hyaline casts or secretion of an estrogen-treated rat. Lobules
increased in size with the time and mammary gland tumors were
induced by the effect of estrogen alone at 240 days after a 5-day
treatment with well characterized lobule formation. Fig. 2D and E correspond to representative
images of control and malathion-treated rat, respectively. Fig. 2F shows the atrophic ducts induced by
atropine.
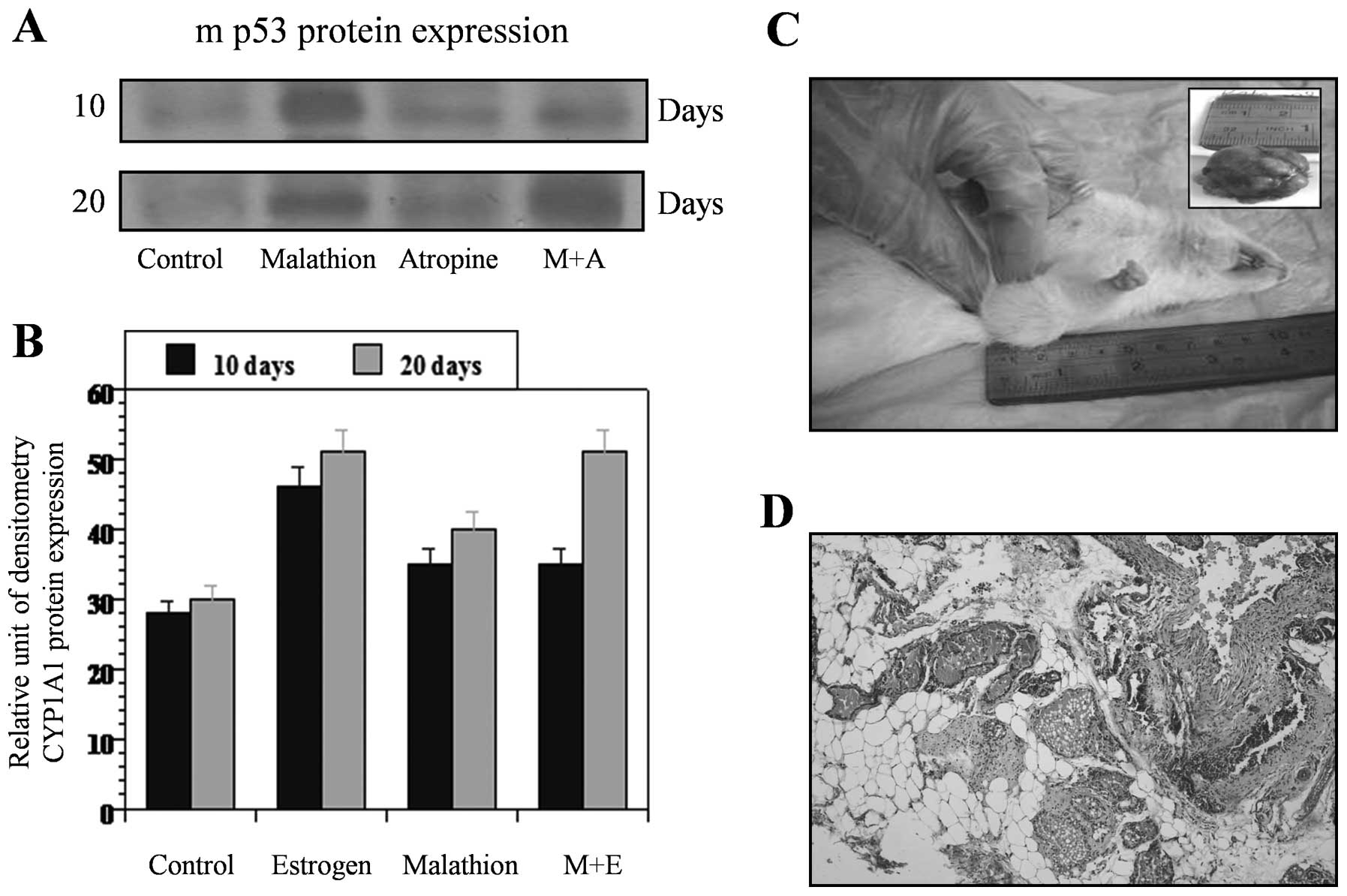
Fig. 3 shows the
effect of estrogen, malathion and combination of both on western
blot analysis of CYP 1A1 (Fig. 3A)
and mp53 (Fig. 3C) protein
expression after 124 and 240 days, respectively. Fig. 3B and D correspond to the relative
unit of densitometry of such an expression. On the other hand,
estrogen and malathion alone and in combination increased c-myc
(Fig. 4A) and c-fos (Fig. 4C) protein expression after 124 and
240 days in comparison to control as seen by western blot analysis
and its quantification (Fig. 4B and
4D). Such expressions were also higher in estrogen-treated
animals when compared to controls after 124 and 240 days.
The increase of proliferative ducts induced by the
effect of malathion after 10 and 20 days, was coincident with the
increase in mutant p53 protein expression by western blot analysis
as quantified by the relative unit of densitometry (Fig. 5A). Atropine and combination of both
diminished such effect after 20 days of a 5-day treatment. Control
animals did not develop any kind of tumors. Combination of
malathion and estrogen induced greater cellular changes in the rat
mammary glands than estrogen or malathion alone (Fig. 5A). Increased amount of proliferative
ducts and secretory lobules were induced by these two substances
showing rat mammary gland tumor formation at 240 days after a 5-day
treatment. Mammary gland tumor formed by the effect of these
substances was characterized by the presence of ducts in stage of
proliferation and secretory lobules (Fig. 5C and D). Analysis of heart,
intestinal tract and ovaries did not show any tumors. However, lung
and kidney were affected as was previously reported (data not
shown).
Discussion
The high level of cell proliferation and
differentiation that occurs during mammary gland development makes
this organ an attractive experimental animal model for examining
its susceptibility to different carcinogenic actions. Cell
proliferation in the mammary gland is not a random event, but is
intimately related to both topography of the mammary parenchyma and
specific stages of the gland development that are modulated by age,
hormonal variations and parity history. The mammary gland is a
complex organ that undergoes continuous changes under the influence
of body growth as well as cyclic hormonal stimulation from birth to
senescence. It is composed of a single primary or main lactiferous
duct that branches into three to five secondary ducts at birth and
during the first week of postnatal life. The ducts are narrow and
straight and end in small club-shaped terminals, called terminal
end buds. Different compartments, such as ducts, ductules, and
intralobular terminal ducts, have been observed in rats. The
intralobular terminal duct is equivalent to the terminal ductal
lobular unit in the human breast, considered the site of origin of
human breast carcinomas (49,50).
Mammary gland tumors formed in the presence of
either estrogen or malathion are similar to lobular and ductal
carcinomas, respectively described in breast biopsies according to
World Health Organization. Our results showed transformation in the
epithelium of ducts while they were in the process of development.
These results indicated that the proliferative changes observed in
such structures may have induced the formation of mammary ductal
carcinomas.
In contrast to potent carcinogens, which induced
mammary carcinomas in 100% of intact females Sprague-Dawley rats by
chemical carcinogens, such as dimethylbenz[α]anthracene
(DMBA), organophosphorous pesticides seem to have a slow and less
infiltrating and potent effect. Tumor incidence of 94–100% was
obtained when DMBA was administered to rats 30–55 days of age.
However, the highest number of tumors per animal was observed when
the carcinogen was given to animals when they were 40–46 days of
age, a period when terminal end buds were most actively
differentiating into alveolar buds (49). The administration of DMBA to virgin
rats of different ages induced tumors with an incidence that was
directly proportional to the density of highly proliferative
structures.
Our previous study (51) examined whether pesticides were able
to induce malignant transformation of the rat mammary gland and
whether alterations induced by these substances increased the
cholinergic activation influencing such transformation. We have
demonstrated that parathion and malathion, formed 14.3 and 24.3% of
rat mammary tumors, respectively, that correlated with the greater
density of ducts in the mammary gland present in the 44 day-old
treated animals. We postulated that it is possibly through
acetylcholinesterase inhibition since atropine decreased such
effects, as corroborated here. These results showed that malathion
increased cell proliferation of terminal end buds of the 44-day-old
mammary gland of rats, followed by formation of 24.3% of mammary
carcinomas, respectively, after ~28 months. At the same time,
acetylcholinesterase activity decreased in the serum of these
animals from 9.78±0.78 U/ml in the control animals to 3.88±0.44
U/ml in malathion-treated groups. However, atropine alone induced a
significant (P<0.05) decrease in the acetylcholinesterase
activity from the control value of 9.78±0.78 to 4.38±0.10 for
atropine alone, to 2.39±0.29 for atropine with malathion, and there
was no mammary tumor formation. These results indicated that
organophosphorous pesticides induce changes in the epithelium of
mammary gland influencing the process of carcinogenesis, and such
alterations occur at the level of the nervous system by increasing
the cholinergic stimulation.
Estrogen in combination with an organophosphorous
pesticide such as malathion can produce several injuries also in
kidney tissue, e.g. increase in glomerular hypertrophy, damage in
the convoluted tubules, and malignant proliferation in cortical and
hilium areas. It is possible that the oxidative stress caused by
malathion exposure enhances the deleterious effects of
17β-estradiol (52). The level of
injuries found in kidney tissue suggested an increased oxidative
stress caused by treatment of malathion in combination with
estrogen. The grade of damage can be serious and can develop tumors
in urothelium of the hilium. Previous studies also showed that
17β-estradiol in combination with pesticides as parathion and
malathion induced malignant transformation of lung in rats
(53).
In conclusion, the combination of the environmental
substance malathion, extensively used in Latin American and in many
other countries, and an endogenous substance such as estrogen can
induce deleterious effects in the humans by inducing breast cancer.
Thus, these studies are relevant to understanding the possible
effects of these agents that through their control can contribute
to a decrease in this disease.
Acknowledgements
The technical assistance of Guiliana Rojas Ordoñez
and Georgina Vargas Marchant are greatly appreciated. The authors
are sincerely thankful by support provided by FONDECYT grant no.
1080482 (GMC) and Convenio de Desempeño Universidad de
Tarapacá-Mineduc, Chile.
References
|
1
|
Dickson RB, Gelmann EP, Knabbe C, et al:
Mechanisms of estrogenic and antiestrogenic regulation of growth of
human breast carcinoma. Hormonal Manipulation of Cancer: Peptides,
Growth Factors, and New (Anti) Steroidal Agents. Klijn JGM: Raven
Press; New York, NY: pp. 381–403. 1987
|
|
2
|
Clark GM and McGuire WL: Steroid receptors
and other prognostic factors in primary breast cancer. Semin Oncol.
18:20–25. 1989.
|
|
3
|
Clarke RB, Howell A, Potten CS, et al:
Dissociation between steroid receptor expression and cell
proliferation in the human breast. Cancer Res. 57:4987–4991.
1997.PubMed/NCBI
|
|
4
|
Henderson BE, Pike MC and Ross RK:
Epidemiology and risk factors. Breast Cancer: Diagnosis and
Management. Bonadonna G: John Wiley and Sons Ltd; New York, NY: pp.
1–17. 1984
|
|
5
|
Henderson BE, Ross RK and Pike MC:
Hormonal chemoprevention of cancer in women. Science. 259:633–638.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perera FP: Environment and cancer: who are
susceptible? Science. 278:1068–1073. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McLachlan JA: Functional toxicology: a new
approach to detect biologically active xenobiotics. Environ Health
Perspect. 101:386–387. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Taylor P: Anticholinesterase agents. The
Pharmacological Basis of Therapeutics. Goodman Gilman A, Rall TW,
Nies AS and Taylor P: Pergamon Press Inc; New York, NY: pp.
131–147. 1990
|
|
9
|
Klaasen C: Nonmetallic environmental
toxicants: air pollutants, solvents and vapors, and pesticides. The
Pharmacological Basis of Therapeutics. Goodman Gilman A, Rall TW,
Nies AS and Taylor P: Pergamon Press Inc; New York, NY: pp.
1615–1635. 1990
|
|
10
|
Silman I and Futerman A: Modes of
attachment of acetylcholinesterase to the surface membrane. Eur J
Biochem. 170:11–20. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krieger N: Rising incidence of breast
cancer. J Natl Cancer Inst. 80:2–3. 1988. View Article : Google Scholar
|
|
12
|
Krieger N: Exposure, susceptibility, and
breast cancer risk: a hypothesis regarding exogenous carcinogens,
breast tissue development, and social gradients, including
black/white differences, in breast cancer incidence. Breast Cancer
Res Treat. 13:205–223. 1989. View Article : Google Scholar
|
|
13
|
Kelsey JL, Gammon MD and John EM:
Reproductive factors and breast cancer. Epidemiol Rev. 15:36–47.
1993.PubMed/NCBI
|
|
14
|
Kelsey JL and Bernstein L: Epidemiology
and prevention of breast cancer. Annu Rev Public Health. 17:47–67.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lippman M, Bolan G and Huff K: The effects
of estrogens and antiestrogens on hormone-responsive human breast
cancer in long-term tissue culture. Cancer Res. 36:4595–4601.
1976.PubMed/NCBI
|
|
16
|
Dickson RB and Lippman ME: Control of
human breast cancer by estrogen, growth factors, and oncogenes.
Cancer Treat Res. 40:119–165. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Key TJ and Pike MC: The dose-effect
relationship between ‘unopposed’ oestrogens and endometrial mitotic
rate: its central role in explaining and predicting endometrial
cancer risk. Br J Cancer. 57:205–212. 1988.
|
|
18
|
Feigelson HS and Henderson BE: Estrogens
and breast cancer. Carcinogenesis. 17:2279–2284. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bernstein L: The epidemiology of breast
cancer. LOWAC J. 1:7–13. 1998.
|
|
20
|
Toniolo PG, Levitz M, Zeleniuch-Jacquotte
A, et al: A prospective study of endogenous estrogens and breast
cancer in postmenopausal women. J Natl Cancer Inst. 87:190–197.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adlercreutz H, Gorbach SL, Goldin BR, et
al: Estrogen metabolism and excretion in Oriental and Caucasian
women. J Natl Cancer Inst. 86:1076–1082. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Berrino F, Muti P, Micheli A, et al: Serum
sex hormone levels after menopause and subsequent breast cancer. J
Natl Cancer Inst. 88:291–296. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shimizu H, Ross RK, Bernstein L, et al:
Serum oestrogen levels in postmenopausal women: comparison of
American whites and Japanese in Japan. Br J Cancer. 62:451–453.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bernstein L, Yuan JM, Ross RK, et al:
Serum hormone levels in pre-menopausal Chinese women in Shanghai
and white women in Los Angeles: results from two breast cancer
case-control studies. Cancer Causes Control. 1:51–58. 1990.
View Article : Google Scholar
|
|
25
|
Bernstein L, Ross RK, Pike MC, et al:
Hormone levels in older women: a study of post-menopausal breast
cancer patients and healthy population controls. Br J Cancer.
61:298–302. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kirkman H: Estrogen-induced tumors of the
kidney. III Growth characteristics in the Syrian hamster. Natl
Cancer Inst Monogr. 1:1–57. 1959.PubMed/NCBI
|
|
27
|
Li JJ, Li SA, Klicka JK, et al: Relative
carcinogenic activity of various synthetic and natural estrogens in
the Syrian hamster kidney. Cancer Res. 43:5200–5204.
1983.PubMed/NCBI
|
|
28
|
International Agency for Research on
Cancer (IARC). IARC Monographs on the Evaluation of Carcinogenic
Risks to Humans. 21. Scientific Publications, IARC; Lyon: pp.
279–362. 1979
|
|
29
|
International Agency for Research on
Cancer (IARC). IARC Monographs on the Evolution of Carcinogenic
Risks to Humans. IARC; (Suppl 7)Lyon: pp. 280–285. 1987
|
|
30
|
Cavalieri EL, Stack DE, Devanesan PD, et
al: Molecular origin of cancer: catechol estrogen-3,4-quinones as
endogenous tumor initiators. Proc Natl Acad Sci USA.
94:10937–10942. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chakravarti D, Mailander P, Cavalieri E,
et al: Estrogen-DNA damage in mouse skin H-ras gene is mutated by
error prone repair. Proc Am Assoc Cancer Res. 41:1072000.
|
|
32
|
Segaloff A and Maxfield WS: The synergism
between radiation and estrogen in the production of mammary cancer
in the rat. Cancer Res. 31:166–168. 1971.PubMed/NCBI
|
|
33
|
Calaf GM and Garrido F: Catechol estrogens
as biomarkers for mammary gland cancer. Int J Oncol. 39:177–183.
2011.PubMed/NCBI
|
|
34
|
International Agency for Research on
Cancer (IARC). IARC Monographs on the Evolution of Carcinogenic
Risks to Humans. 72. Hormonal Contraception and Postmenopausal
Hormone Therapy. IARC; Lyon: 1999
|
|
35
|
Huseby RA: Demonstration of a direct
carcinogenic effect of estradiol on Leydig cells of the mouse.
Cancer Res. 40:1006–1013. 1980.PubMed/NCBI
|
|
36
|
Highman B, Norwell MJ and Shellenberger
TE: Pathological changes in female C3H mice continuously fed diets
containing diethylstilbestrol or 17beta-estradiol. J Environ Pathol
Toxicol. 1:1–30. 1978.
|
|
37
|
Highman B, Greenman DL, Norvell MJ, Farmer
J and Shellenberger TE: Neoplastic and preneoplastic lesions
induced in female C3H mice by diets containing diethylstilbestrol
or 17beta-estradiol. J Environ Pathol Toxicol. 4:81–95. 1980.
|
|
38
|
Highman B, Roth SI and Greenman DL:
Osseous changes and osteosacomas in mice continuously fed diets
containing diethylstilbestrol or 17beta-estradiol. J Natl Cancer
Inst. 67:653–662. 1981.PubMed/NCBI
|
|
39
|
Nagasawa H, Mori T and Nakajima Y:
Long-term effects of progesterone or diethylstilbestrol with or
without estrogen after maturity on mammary tumorigenesis in mice.
Eur J Cancer. 16:1583–1589. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Inoh A, Kamiya K, Fujii Y and Yokoro K:
Protective effects of progesterone and tamoxifen in
estrogen-induced mammary carcinogenesis in ovariectomized W/Fu
rats. Jpn J Cancer Res. 76:699–704. 1985.PubMed/NCBI
|
|
41
|
Noble RL, Hochachka BC and King D:
Spontaneous and estrogen-produced tumors in Nb rats and their
behavior after transplantation. Cancer Res. 35:766–780.
1975.PubMed/NCBI
|
|
42
|
Shull JD, Spady TJ, Snyder MC, Johansson
SL and Pennington KL: Ovary-intact, but not ovariectomized female
ACI rats treated with 17beta-estradiol rapidly develop mammary
carcinoma. Carcinogenesis. 18:1595–1601. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Welsch CW: Host factors affecting the
growth of carcinogen-induced rat mammary carcinomas: a review and
tribute to Charles Brenton Huggins. Cancer Res. 45:3415–3443.
1985.PubMed/NCBI
|
|
44
|
Li JJ and Li SA: Estrogen-induced
tumorigenesis in hamsters: roles for hormonal and carcinogenic
activities. Arch Toxicol. 55:110–118. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li JJ and Li SA: Estrogen carcinogenesis
in Syrian hamster tissues: role of metabolism. Fed Proc.
46:1858–1863. 1987.PubMed/NCBI
|
|
46
|
Bhat HK, Calaf GM, Hei TK, et al: Critical
role of oxidative stress in estrogen-induced carcinogenesis. Proc
Natl Acad Sci USA. 100:3913–3918. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liehr JG: Genotoxic effects of estrogens.
Mutat Res. 238:269–276. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Institute of Animal Laboratory Resources.
Guide for the Care and Use of Laboratory Animals. National Academy
Press; Washington, DC: 1996
|
|
49
|
Russo J and Russo IH: Mammary
tumorigenesis. Prog Exp Tumor Res. 33:175–191. 1991.
|
|
50
|
Wellings SR, Jensen HM and Marcum RG: An
atlas of subgross pathology of the human breast with special
reference to possible precancerous lesions. J Natl Cancer Inst.
55:231–273. 1975.PubMed/NCBI
|
|
51
|
Cabello G, Valenzuela M, Vilaxa A, et al:
A rat mammary tumor model induced by the organophosphorous
pesticides parathion and malathion, possibly through
acetylcholinesterase inhibition. Environ Health Perspect.
109:471–479. 2001. View Article : Google Scholar
|
|
52
|
Alfaro-Lira S, Pizarro-Ortiz M and Calaf
GM: Malignant transformation of rat kidney induced by environmental
substances and estrogen. Int J Environ Res Public Health. (In
Press).
|
|
53
|
Echiburu-Chau C and Calaf GM: Rat lung
cancer induced by malathion and estrogen. Int J Oncol. 33:603–611.
2008.PubMed/NCBI
|