Introduction
Liver cancer is the fifth most common cancer and the
seventh most frequently diagnosed cancer worldwide (1). Among primary liver cancers occurring
worldwide, hepatocellular carcinoma (HCC) represents the major
histological type, which accounts for 70–85% of cases (2). Half of the liver cancer cases and
deaths were estimated to occur in China for both men and women
(1). The high HCC rate in China and
in developing countries largely reflects the elevated prevalence of
chronic hepatitis B virus (HBV) infection, with over 8% of the
populations in these regions chronically infected with the virus
(3). Hepatitis C virus (HCV)
infection accounts for 33% of the total liver cancer cases in
developing countries, and for 20% in developed countries.
Interaction of aflatoxin B (AFB) exposure and chronic HBV infection
is known to increase the risk of liver cancer (4). Alcohol-associated cirrhosis and
possibly non-alcoholic fatty liver disease are believed to be
related to obesity and are the leading causes of liver cancer
(5). However, current
chemotherapeutic strategies have serious limitations and yield
undesirable side effects. Moreover, no vaccine is available for
HCV. Preventive strategies against HCV and complementary medicine
are needed to reduce the number of cancer deaths.
Previous studies have reported the pharmacological
properties of Coptis chinensis Franch formulation for the
treatment of various liver disorders and its major constituent
triterpenoids (6–8). One of the major constituent
phytochemicals was demonstrated to exhibit marked anti-inflammatory
and anti-proliferative effects on cancer cells. However, the
details of its mechanisms are lacking.
In the present study, the Huh7 liver cancer cell
line and the WRL68 normal liver cell line were used to study the
antitumor activity of berberine from Coptis chinensis
Franch, which is commonly used in combination with other herbs for
the treatment of various diseases. Additionally, the mode of action
of berberine in the Huh7 cell line was investigated.
Materials and methods
Chemicals and reagents
Berberine chloride was purchased from Sigma-Aldrich
Inc. (China), and Dulbecco’s phosphate-buffered saline (DPBS),
fetal bovine serum (FBS), a penicillin/streptomycin antibiotic
solution (PS), Roswell Park Memorial Institute (RPMI)-1640 medium,
and Trypsin-EDTA (1X) were purchased from Invitrogen Life
Technologies (Carlsbad, CA, USA). Minimal essential medium (MEM)
was purchased from ATCC (Manassas, VA, USA).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and sodium bicarbonate were obtained from Sigma Chemicals (St.
Louis, MO, USA). Dimethyl sulfoxide (DMSO) was purchased from
Fisher Scientific Inc. (Rockford, IL, USA). The Millex-GP filter
(0.22-μm) was purchased from Millipore (Billerica, MA, USA). Tissue
culture flasks (75-cm2) were ordered from Corning, USA.
Ninety 6-well microplates were purchased from Iwaki. Culture dishes
(100-mm) were purchased from Greiner Bio-One (Germany).
Cell treatment
WRL68 liver cells and Huh7 liver cancer cells were
grown in RPMI-1640 with 10% FBS. Cells were suspended with fresh
complete medium (5 ml) and were centrifuged at 600 × g for 3 min.
The cell pellets were washed with 1 ml of DPBS before
centrifugation at 600 × g for 3 min. Cells were transferred to a
new 75-cm2 culture flask with 12 ml of complete medium.
The flask was stored in a 37°C incubator supplied with 5% carbon
dioxide before use.
MTT assay
Huh7 and WRL68 cells (1×106) were seeded
onto a 96-well plate overnight prior to incubation with various
concentrations of berberine for 24, 48 and 72 h, respectively. MTT
solution (20 μl) was added to each well, and the plate was
maintained at 37°C with 5% carbon dioxide for 4 h. After
incubation, the plate was dried. DMSO (200 μl) was added to
dissolve the purple formazan formed in the assay prior to
measurement of absorbance at 540 nm. The IC50 value was
determined.
Detection of apoptosis by fluorescein
isothiocyanate (FITC) Annexin V and propidium iodide (PI)
co-staining
Huh7 cells were treated according to the
manufacturer’s protocol. Cells were treated with different
concentrations of berberine for 24, 48 and 72 h, respectively.
Following berberine treatment, cells were harvested with trypsin
and stained according to the BD Pharmingen™ FITC Annexin V staining
protocol. The stained cells were analyzed using a FACSCanto™
instrument. Data were analyzed with software FlowJo 7.6.1.
Gene expression in berberine-induced
apoptotic cells
Total RNA was extracted using the RNeasy mini kit
(Qiagen, Hilden, Germany) according to the manufacturer’s protocol.
First-strand cDNA was synthesized using 1 μg of total RNA using the
Transcriptor First Strand cDNA Synthesis kit (Roche Diagnostics).
Anchored-oligo(dT)18 primer was used in the template-master mix.
Experimental cocktail was prepared as recommended by RT2 Profiler™
polymerase chain reaction (PCR) array system pathway-focused gene
expression profiling using real-time PCR protocol.
Western blot analysis of gene expression
in berberine-treated cells
Huh7 cells (1×106) were seeded onto a
100-mm culture dish overnight prior to incubation with berberine.
Following the incubation of cells with berberine for 72 h, cell
lysates were extracted using whole cell lysis buffer. The protein
concentration was determined by the DC protein assay (Bio-Rad). The
protein was transferred onto a PVDF membrane (Pall Corp.). The
blots were treated with blocking solution (5%) for 1 h, and were
probed with a specific primary antibody with 5% non-fat milk powder
in TBST at 4°C overnight. After washing with TBST, the blots were
treated with the specific HRP-linked secondary antibody for 1 h and
were washed thrice. Proteins were detected using Rodeo™ ECL western
blotting reagents (USB, Cleveland, OH, USA) according to the
manufacturer’s protocol.
Caspase cascade in berberine-treated
cells
Cells were treated with different concentrations of
berberine for 72 h. Apo-ONE® Homogeneous Caspase-3/7
assay was performed according to the ‘Promega Technical
Bulletin-Apo-ONE® Homogeneous Caspase-3/7 assay
instructions for use of products G7790, G7791 and G7792′. The plate
was incubated and shaken at room temperature for 2 h. The
fluorescence of each well was measured (excitation wavelength, 485
nm; emission wavelength, 520 nm).
Cell cycle analysis of Huh7 cells
Cells were harvested with trypsin after berberine
treatment and washed with cold DPBS twice. Cells were collected by
centrifugation and re-suspended in cold 70% (w/v) ethanol at a
concentration of 1×106 cells/ml for 24 h before being
washed by cold DPBS. Cells were re-suspended with 400 μl of PI
solution and transferred to a 5-ml polystyrene round-bottom tube at
25°C in the dark for 15 min prior to analysis by BD FACSCanto™ flow
cytometer.
Statistical analysis
FACSDiva™ software was used to operate the BD
FACSCanto™ flow cytometer. Experimental data were captured and were
analyzed by FlowJo 7.6.1. The percentage of cell populations in the
G1, G2 and S phases of the cell cycle was recorded. The experiment
was performed three times. The ratios of cells in the G0/G1,
intra-S and G2/M phases were expressed as means ± SD.
Results
Berberine induces apoptosis of
hepatocellular cells
Effects of berberine on the viability of
hepatocellular carcinoma (HCC) cells (Fig. 1) and normal liver cells (Fig. 2) were evaluated by the MTT assay.
The IC50 for Huh7 was 10 μM while the IC50
for WRL68 cells was 100 μM berberine following a 72-h treatment.
Table I summarizes the
IC50 values of the two cell lines after berberine
treatment. Apoptosis of HCC cell lines was detected by the BD
Pharmingen™ FITC Annexin V Apoptosis Detection Kit I. The results
demonstrated that berberine induced the cell death of Huh7 cells.
The results showed that cell viability of Huh7 cells was reduced
after 24, 48 and 72 h of incubation with berberine (Fig. 3). The percentage of apoptotic cells
increased with the increase in berberine concentration. The results
showed that berberine induced the apoptosis of Huh7 cells.
 | Table IIC50 (μM) values for Huh7
and WRL68 liver cell lines after incubation with berberine for
different time periods. |
Table I
IC50 (μM) values for Huh7
and WRL68 liver cell lines after incubation with berberine for
different time periods.
| Time (h) | 24 | 48 | 72 |
|---|
| Huh7 | 150 | 100 | 10 |
| WRL68 | 1000 | 120 | 100 |
Gene expression profile in the
berberine-treated cells
Gene expression profile in the HCC cell line was
assessed by PCR array and real-time PCR. PCR array was used to
profile the gene expression in the Huh7 cells. Real-time PCR was
used for the quantitative measurement of gene expression in the
berberine-treated HCC cells.
PCR array was used as a preliminary screening
procedure for genes that were significantly affected by berberine
(Fig. 4). The relative gene
expression levels in the cell samples were plotted against the
control in a scatter plot using the program provided by the
manufacturer. The middle line indicates relative fold changes. The
left and right lines indicate the fold-change in gene expression
threshold, which was defined as 3-fold. Outliers of the left line
were found to be genes upregulated by at least 3-fold, which
included BCL2, BCL2L1, CASP14, CD40, CIDEA, FASLG, GADD45A, HRK,
LTA, TNFRSF10A, TNFRSF10B, CD27, TNFRSF9 and CD70. The outlier of
the right line was BCLAF1, which was downregulated by at least
3-fold. Bax, Bid, CIDEA, HRK and p21 were found to be upregulated
by berberine in a dose-dependent manner, while AKT and Bcl-2 were
found to be downregulated by berberine in a dose-dependent manner.
The gene expression of survivin decreased with the increase in
berberine concentration. The gene expression of Bcl-2, CIDEA and
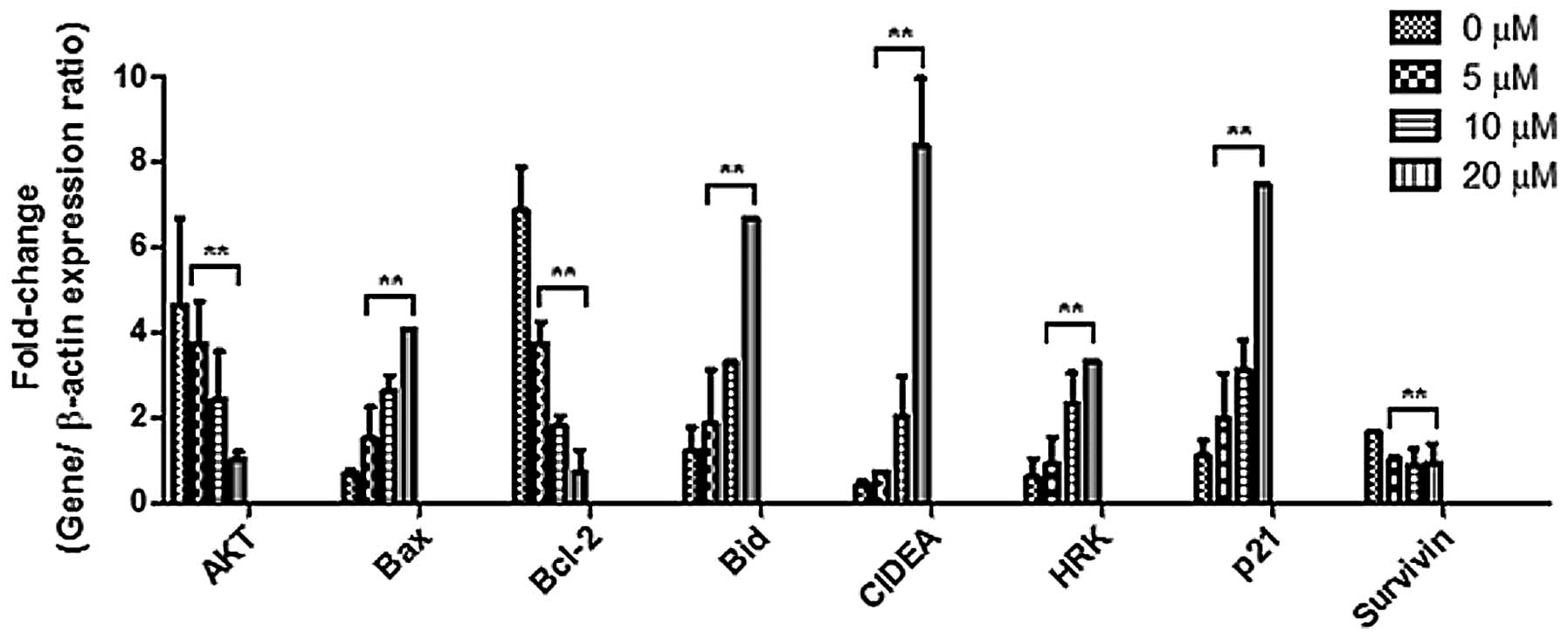
HRK was validated by real-time PCR (Fig. 5). The real-time PCR results
corroborated with the PCR array analysis in regards to the CIDEA
gene.
Caspase cascade and the cell cycle
distribution in the berberine-treated Huh7 cell line
The caspase cascade was evaluated by SDS-PAGE
(Fig. 6) and western blot analysis
(Fig. 7). The expression of Bcl-2
protein family members including Bcl-2 and Bid, and PARP and PCNA
was altered following treatment with berberine (Fig. 8).
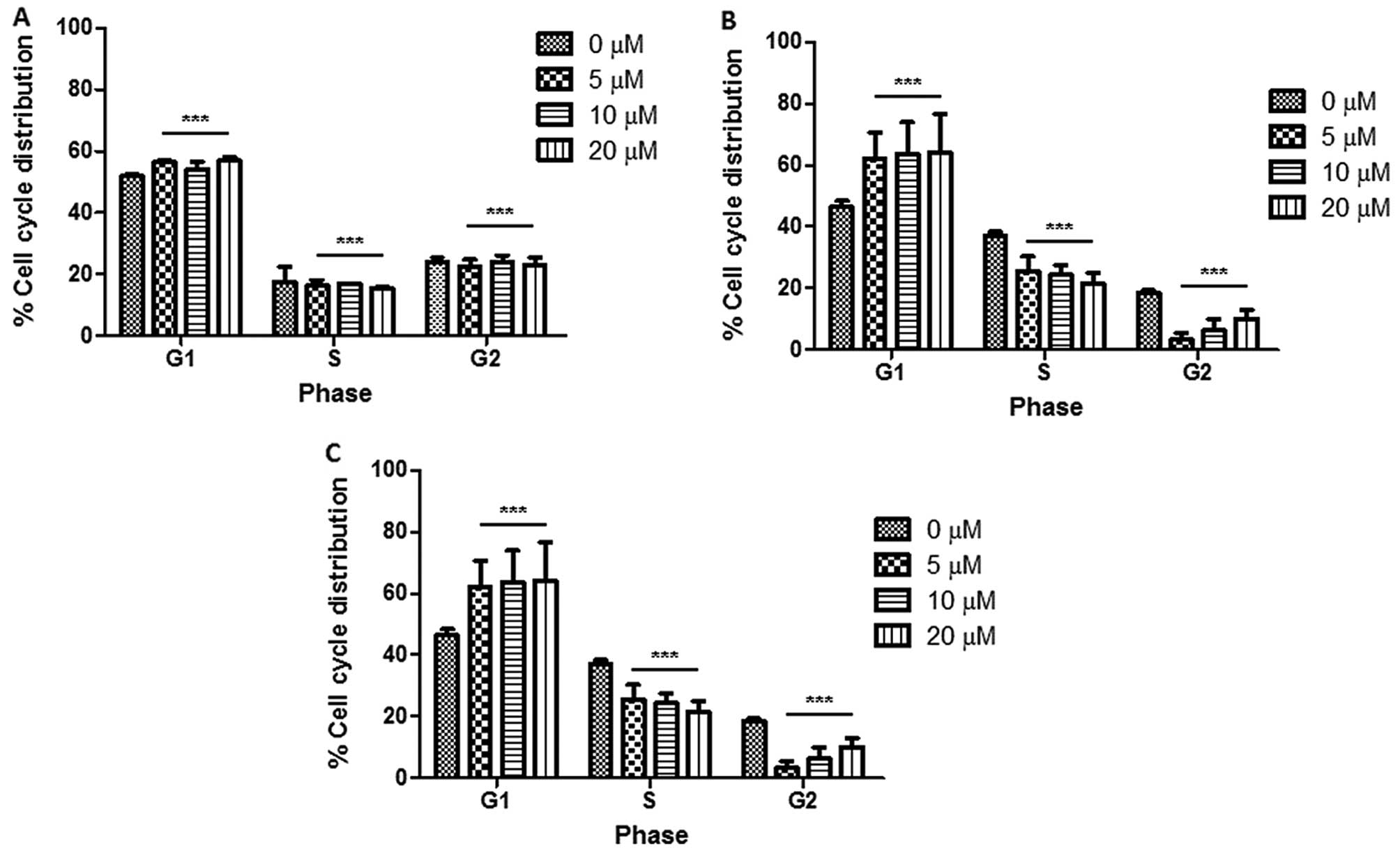
Cell cycle analysis showed that there was an
increase in the G1 cell population with an increase in the
concentration of berberine, suggesting that berberine caused G1
phase cell cycle arrest in the Huh7 cells. The cell cycle
distribution was investigated at three time points: 24, 48 and 72
h, respectively (Fig. 9). Fig. 9A and B shows that there was a slight
increase in the percentage of cells in the G1 phase with an
increase in berberine concentration. Fig. 9B–D reveals that the percentage of
cells in the G1 phase following treatment with 5, 10 and 20 μM
increased by 10% compared with the percentage of cells in the
control without berberine treatment. The distribution of the cell
cycle is shown in Fig. 10.
Discussion
Berberine was found to reduce the cell viability of
Huh7 cells in a dose-dependent manner. It exhibited a relatively
reduced effect on normal liver cells. The results suggest that
berberine exhibited differential effects on normal and cancer
cells. The difference in inhibitory activities suggests that
berberine initiates a different signaling process in normal and
cancer cells.
FITC Annexin V and PI co-staining revealed that
berberine caused apoptosis in Huh7 HCC cells (Fig. 3). The Cide family is known as the
inhibitory subunits of endonucleases, cleavage of which is induced
by downstream caspases (9). The
results support the notion that berberine induced the apoptosis of
cancer cells. Fig. 5 shows that
Bax, Bid, CIDEA, HRK and p21 were upregulated, while AKT and Bcl-2
were downregulated in Huh7 cells following treatment with
berberine. The change in the gene expression profile suggests that
berberine induced the apoptosis of Huh7 cells through their
intrinsic pro-apoptotic activity which was reported to be
associated with mitochondrial dysfunction in cell death (9). The p21 protein is known to serve as a
regulator of cell cycle progression at the G1 phase (10). The increase in p21 gene expression
(Fig. 6) suggests that p21
regulated the cell cycle progression following treatment with
berberine. The AKT protein was reported to participate in the
regulation of tumor cell survival and proliferation by stabilizing
p21 protein (11). Bcl-2 gene
expression was decreased in the present study. The results suggest
that berberine inhibited the gene expression of the Bcl-2 gene in
the HCC cells. An increased gene expression of Bid implies an
increased protein expression of Bid in the cytosol. It was reported
that survivin is an anti-apoptosis gene expressed in cancer and
lymphoma (12,13). The present results indicate that
survivin negatively regulated apoptosis by inhibiting caspase
activation. The findings demonstrated that berberine may inhibit
survivin gene expression in Huh7 cells. The real-time PCR results
agreed with the PCR array analysis regarding the HRK gene, which
was reported to be inhibited by Bcl-2 and Bcl-xL (13). Previous studies have reported that
increased Bcl-2 protein expression causes resistance to
chemotherapeutic drugs and radiation therapy, while decreasing
Bcl-2 expression promotes apoptosis induced by anticancer drugs
(10). The results reflect the fact
that an overexpression of Bcl-2 may cause accumulation of cells in
the G0 phase of the cell cycle distribution, resulting in
chemoresistance (14). The present
findings provide experimental evidence that berberine modulated the
protein expression of Bcl-2 associated with caspase-3/7 activities
in HCC cells. Higher caspase-3/7 activities were observed in Huh7
cells. It has been reported that cytochrome c interacts with
Apaf-1, which activates caspase-9 (15). The effector caspases, caspase-7 and
caspase-3, are downstream targets of caspase-9. Poly(ADP-ribose)
polymerase (PARP) is a well-known downstream target of active
caspase-3 (16). PARP was reported
to produce poly(ADP-ribosyl)ation of nuclear proteins with NAD as a
substrate. PARP is inactivated by cleavage. The present findings
demonstrated that the protein expression of full-length PARP
decreased while the cleaved form increased. Since berberine
decreased the protein expression of procaspase-9 and its downstream
effector caspases, procaspase-3 and procaspase-7, it is possible
that berberine cleaves caspase-9, caspase-3 and caspase-7. Cleaved
caspases become active executioners of the intrinsic apoptotic
pathway. The protein expression of proliferating cell nuclear
antigen (PCNA) was found to be downregulated by berberine in Huh7
cells. The experimental results indicate that as the expression of
PCNA was reduced by berberine, fewer cells were able to repair
damaged DNA.
The results suggest that berberine may halt HCC
cells from dividing, thereby prohibiting deregulated cell
proliferation and demonstrate that berberine inhibits the growth of
HCC cells by inducing the intrinsic apoptotic pathway, and by
arresting the cell cycle at the G1/S phase. Berberine induces
apoptotic events in HCC cells through procaspase-9, and its
effector caspases, procaspase-3 and procaspase-7. Our findings
revealed that berberine possesses potent anticancer activity in
human hepatocellular carcinoma.
Acknowledgements
This study was supported by Tissue Mgfy Ltd. through
grant no. 6903292.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2
|
Perz JF, Armstrong GL, Farrington LA,
Hutin YJ and Bell BP: The contributions of hepatitis B virus and
hepatitis C virus infections to cirrhosis and primary liver cancer
worldwide. J Hepatol. 45:529–538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Campo RA, Rowland JH, Irwin ML, Nathan PC,
Gritz ER and Kinney AY: Cancer prevention after cancer: changing
the paradigm - a report from the American Society of Preventive
Oncology. Cancer Epidemiol Biomarkers Prev. 20:2317–2324. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Parkin DM: The global health burden of
infection-associated cancers in the year 2002. Int J Cancer.
118:3030–3044. 2006.PubMed/NCBI
|
|
5
|
El-Serag HB: Epidemiology of
hepatocellular carcinoma in USA. Hepatol Res. 37(Suppl 2): S88–S94.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mahata S, Bharti AC, Shukla S, Tyagi A,
Husain SA and Das BC: Berberine modulates AP-1 activity to suppress
HPV transcription and downstream signaling to induce growth arrest
and apoptosis in cervical cancer cells. Mol Cancer. 10:392011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo Y, Hao Y, Shi TP, Deng WW and Li N:
Berberine inhibits cyclin D1 expression via suppressed binding of
AP-1 transcription factors to CCND1 AP-1 motif. Acta Pharmacol Sin.
29:628–633. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu HL, Hsu CY, Liu WH and Yung BY:
Berberine-induced apoptosis of human leukemia HL-60 cells is
associated with down-regulation of nucleophosmin/B23 and telomerase
activity. Int J Cancer. 81:923–929. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo Y, Hurwitz J and Massagué J:
Cell-cycle inhibition by independent CDK and PCNA binding domains
in p21Cip1. Nature. 375:159–161. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y, Dowbenko D and Lasky LA: AKT/PKB
phosphorylation of p21Cip/WAF1 enhances protein
stability of p21Cip/WAF1 and promotes cell survival. J
Biol Chem. 277:11352–11361. 2002.PubMed/NCBI
|
|
12
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inohara N, Ding L, Chen S and Núñez G:
harakiri, a novel regulator of cell death, encodes a protein that
activates apoptosis and interacts selectively with
survival-promoting proteins Bcl-2 and Bcl-X(L). EMBO J.
16:1686–1694. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reed JC: Bcl-2 family proteins: regulators
of apoptosis and chemoresistance in hematologic malignancies. Semin
Hematol. 34:9–19. 1997.PubMed/NCBI
|
|
15
|
Earnshaw WC, Martins LM and Kaufmann SH:
Mammalian caspases: structure, activation, substrates, and
functions during apoptosis. Ann Rev Biochem. 68:383–424. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boulares AH, Yakovlev AG, Ivanova V,
Stoica BA, Wang G, Iyer S and Smulson M: Role of poly(ADP-ribose)
polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP
mutant increases rates of apoptosis in transfected cells. J Biol
Chem. 274:22932–22940. 1999. View Article : Google Scholar : PubMed/NCBI
|