Introduction
Cancer remains one of the most common causes of
death worldwide, and breast cancer is the most commonly diagnosed
cancer type, and a leading cause of cancer-related deaths in women,
particularly in western countries (1,2).
Despite numerous inspiring advances in the past years, the
underlying mechanisms of breast cancer initiation and progression
remain elusive. Moreover, once cancer cells form secondary tumors
in other organs, the disease essentially enters an incurable phase.
Metastatic tumors are currently treated as a systemic disease with
conventional therapeutics, such as chemotherapy and/or radiation.
Although these strategies ameliorate metastatic progression and
relevant implications, it is impossible to stop or reverse the
progression of metastatic disease. To this end, tremendous efforts
have been invested in search of therapeutics for breast cancer
metastasis, and some progress has been achieved to date. For
example, zoledronic acid (Zometa®) has been promisingly
used as an adjuvant therapy to treat breast cancer with metastasis,
in combination with chemotherapy, and it has significantly improved
patient prognosis (3,4). However, no metastasis-specific
therapeutics exist. Therefore, further research on breast cancer
metastasis should be conducted.
To study the metastatic process requires detailed
insights into the cellular and molecular traits for both primary
and metastatic tumors, as their differences determine the
fundamental metastatic propensity. However, it is difficult to
perform such studies with human primary breast tumors and
corresponding metastatic tumors concomitantly, since these paired
samples are mostly unavailable for laboratory investigation. Thus,
it is desirable to establish mouse models of breast cancer
metastasis. To date, several breast cancer cell lines have been
established with enhanced inclination to metastasize to distant
organs. For example, a derivative subline (termed 4175) of human
MDA-MB-231 cells was established with strong inclination for lung
metastasis in BALB/c nude mice (5,6). It
should be noted that research results from mice may not be able to
reflect corresponding changes in humans, and the mechanisms
responsible for the metastases of human breast cancer cells to
mouse organs should be distinct from those to human organs. Thus,
it would be more informative for mechanistic studies to use the
same species of breast cancer cells and recipients. To this end,
mouse breast cancer cells would be the perfect fit for this
purpose. By contrast, there are a limited number of mouse breast
cancer cell lines available for experimental studies. Among these,
4T1 cells have been widely used for the study of breast cancer
development and progression, and these cells pose a propensity to
metastasize to distant organs (7,8). When
inoculated orthotopically in mammary fat pads (MFPs), 4T1 cells
spontaneously metastasize to several organs including the lung,
liver, brain and bone (7,9,10).
However, it takes a long time (e.g. 5–6 weeks) for the parental 4T1
cells to form metastatic tumors in distant organs in mice, when
growth regression of primary tumors has occurred (3–4 weeks) with
extensive necrosis and infiltration of leukocytes (8). Moreover, the occurrence of metastases
is limited in mice after introduction of parental 4T1 cells at MFPs
(7,11). Therefore, sibling cell lines of 4T1
cells with enhanced metastatic properties are desirable for the
study of the molecular basis of tumor metastasis. In the present
study, we aimed to isolate and establish sublines of 4T1 cells that
exhibit enhanced propensity to metastasize. We purified metastatic
tumor cells from lungs, and eventually established sublines with
enhanced capability to metastasize to the lungs.
Materials and methods
Cell culture
Mouse breast cancer 4T1 cells were provided as a
gift from the Caliper Life Sciences, Inc. The genetically
engineered cells are stably tagged with the luciferase gene. Cells
were cultured in RPMI-1640 (Gibco) media, supplemented with 10%
fetal bovine serum (FBS) and 100 U/ml penicillin-streptomycin (both
from HyClone) at 37°C under 5% CO2.
Mouse experiments
All mouse experiments were approved by the Animal
Ethics Committee of the Research Center for Eco-Environmental
Sciences, Chinese Academy of Sciences. Mice with BALB/c genetic
background were purchased from the Vital River Laboratories
(Beijing, China). Mice were housed under specific pathogen-free
(SPF) environment, and surgeries were performed under sterile
conditions. For tumor cell inoculation, we followed a previously
described method (12,13). Briefly, 2.0×103 cells
were implanted into the fourth mammary fat pads (MFPs) of the mice.
Thereafter, the mice were monitored for bioluminescent imaging of
luciferase activity every week using the Spectrum Imaging System
and Living Image software (Caliper Life Sciences, Inc.). Prior to
imaging, the mice were intraperitoneally administered 200 μl
luciferin (at 15 mg/ml; PerkinElmer). Afterwards, the mice were
imaged after anesthetization with isoflurane inhalation.
Immediately after the mice were sacrificed, various organs were
imaged.
Western blot analysis
Cells were collected and lysed in RIPA lysis buffer
containing protease inhibitor cocktail (Roche). The concentration
of total proteins was determined using the Lowry method. Equal
amounts of protein lysates for each sample were subjected to
SDS-PAGE. Thereafter, western blot analyses were performed
following a standard protocol, as previously described (12,13).
Antibodies used here were against GAPDH (1:1,000; Santa Cruz),
anti-Snail1 (1:500) and anti-occludin (1:500) (both from Bioss,
Inc., China). GAPDH was used as a loading control for
normalization. Chemiluminescent detection was performed by the
ChemiDoc™ XRS+ System with Image Lab™ Software (Bio-Rad, Inc.). The
intensity of the protein bands in the blots was determined with
ImageJ software (NIH, Bethesda, MD, USA).
Cloning of tumor cells from primary
tumors and metastatic tumors in the lungs
Firstly, primary tumors were gently removed and
maintained in phosphate-buffered saline (PBS). Thereafter,
metastatic tumors were carefully dissected from the surrounding
connective tissues in the lungs. We prepared a single-cell
suspension of the tumor cells following a standard protocol, as
previously described (14). Tumor
cells were then cultured as their parental 4T1 cells.
Genomic DNA extraction and PCR
analysis
To characterize the identity of the 4T1 sublines,
genomic DNA was extracted from the sublines on the basis of the
manufacturer’s protocol (Qiagen). Then, as previously described
(12), PCR analysis was carried out
under the conditions as follows; 94°C for 5 min; followed by 94°C
for 30 sec, 55°C for 35 sec and 72°C for 35 sec for 40 cycles;
finally, an extension at 72°C for 5 min. Thereafter, the PCR
products were subjected to electrophoresis on a 1.5% agarose gel,
followed by staining with ethidium bromide. Primers for PCR
reactions were: puromycin F, 5′-ATGACCGAGTACAAGCCCAC-3′ and R,
5′-GAGGCCTTCCATCTGTTGCT-3′; luciferase F,
5′-CGAGGCTACAAACGCTCTCA-3′ and R, 5′-CGAAGATGTTGGGGTGTTGC-3′.
Cell imaging for luciferase activity
Cells were collected and re-suspended in pre-warmed
PBS. An equal number of cells in 100 μl were re-seeded into
96-well plates, followed by the addition of 100 μl of
luciferin (300 μg/ml). The plates were incubated at 37°C for
2 min before imaging with the Spectrum Imaging System.
Histological analysis
Hematoxylin and eosin (H&E) staining was carried
out following the standard protocol. Briefly, lung specimens were
first fixed in 4% PBS-buffered formaldehyde, and then embedded in
paraffin. Sections were finally stained with H&E solutions.
Invasion and migration assays
The capability of tumor cell invasion and migration
was determined with the transwell method, as previously described
(13,15). Briefly, 100 μl of diluted
matrigel (at 200 μg/ml) was loaded onto the upper chambers
of transwells, followed by incubation at 37°C for 3 h. Afterwards,
5.0×104 cells were seeded onto the upper chambers in 100
μl of serum-free medium. Thereafter, 600 μl of medium
with 10% FBS was added into the bottom wells. After the cells were
cultured for 5 h, the filters were removed from the upper chambers,
and then fixed with 4% formaldehyde. The filters were stained with
5 μg/ml of 4,6-diamidino-2-phenylindole (DAPI) solution.
Cell nuclei were visualized and counted under a fluorescence
microscope.
Data analysis
Experimental data were analyzed with the SPSS
Statistics 17.0 software package. Independent t-test was used to
determine the difference between two groups. Data are shown as mean
± standard error. Statistical significance was determined at
P<0.05.
Results
Purification of metastatic cells from
metastatic tumors in the lungs
In an effort to obtain 4T1 cells with enhanced
capability to specifically metastasize to the lungs, we endeavored
to isolate metastatic tumors using a mouse model of breast cancer
metastasis, as previously described by our research group (13). As shown in Fig. 1, a scheme was depicted for the
purification and screening of metastatic cells from the metastatic
tumors in the lungs in the present study. Six weeks post 4T1 cell
inoculation in the bilateral MFPs, primary tumors reached ~1 cm in
diameter. When the mice were sacrificed, the primary tumors were
dissociated and saved in PBS for later preparation of a single-cell
suspension. Meanwhile, the lungs with marked tumors were also
dissected, and then the tumors were carefully removed from the
surrounding connective tissues. The generation of a single-cell
suspension from the primary and metastatic tumors was performed
with a standard protocol, as previously described (14). Cells were seeded in 96-well plates,
and wells with a single cell/well were selected for subsequent
subcloning and screening.
Subcloning of tumor cells with the
capability of survival for an in vitro culture
The regimen for cell subcloning was similar to a
previously described method (16).
After colony formation for ~2 weeks, the 96-well plates were
examined for luciferase activity using the Spectrum Imaging System.
As shown in Fig. 2A, the
representative images showed a few clones with bright
bioluminescence from the primary tumors and metastatic tumors.
Afterwards, the identity of these sublines of 4T1 cells was
assessed through PCR using two markers, namely puromycin and
luciferase. Since these two markers were located in the original
plasmid that was stably transfected into the 4T1 cells, they were
detected only in the 4T1-derived cells. As shown in Fig. 2B, these cells were positive for
these two markers, suggesting that they were sublines derived from
the 4T1 cells. These sublines were accordingly named as 4T1-P1,
4T1-P2, 4T1-P3, 4T1-P4 and 4T1-P5, representing sublines from the
primary tumors, and 4T1-M1, 4T1-M2, 4T1-M3, 4T1-M4 and 4T1-M5,
denoting sublines from the metastatic tumors.
Characterization of the sublines of the
4T1 cells
Next, we characterized the metastatic propensity of
these subclones. The migration/invasion capabilities were assessed
with the transwell method, as previously described (12,13).
As reflected by the transwell results, the number of transmigrated
cells was substantially increased for sublines from the metastatic
tumors, compared to sublines from the primary tumors (Fig. 3A). Fig.
3A shows the representative images of the transmigrated cells
with DAPI staining for the 4T1-P1, in comparison to the 4T1-M1
cells. The quantitative data indicated nearly a 9-fold difference
between these two groups (Fig. 3B,
P<0.05). These data demonstrated that the in vitro
mobility and invasion capabilities were significantly enhanced in
sublines from the metastatic tumors, compared to those from the
primary tumors.
Elevated propensity of metastatic
sublines of 4T1 cells to the lungs
To validate the in vitro results, we
thereafter selected two representative sublines, i.e. 4T1-M1 and
4T1-P1, for the in vivo study. We chose these two sublines
since both displayed the strongest phenotypes in their own groups.
In other words, 4T1-M1 exhibited the greatest capability to migrate
and invade, in contrast, 4T1-P1 showed the weakest capability of
migration and invasion through the transwell assay. We
orthotopically inoculated 4T1-M1, 4T1-P1 and parental 4T1 cells
into MFPs of the mice. During the 38-day experiment, the mice from
all 3 groups developed primary tumors with 100% tumor occurrence.
Two mice in the 4T1-M1 group died at the late stage of the
experiment (Fig. 4A). Therefore,
the survival rate was 80%, in contrast to a 100% survival rate in
the other two groups (Fig. 4A),
suggesting reduced prognosis for mice upon introduction of the
4T1-M1 cells. The final tumor weight was increased in the mice
inoculated with the 4T1-M1 cells, compared to the other two groups
(Fig. 4B), despite of no
statistical significance. Furthermore, we examined the occurrence
of metastatic tumors in the lungs through bioluminescent imaging
and histological examination. As shown in Fig. 5A, all mice harbored metastatic
tumors in the lungs in the group of 4T1-M1, namely 100% lung
metastasis for 4T1-M1 cells. In contrast, only 10% of the mice
developed lung metastasis upon inoculation of the 4T1-P1 cells.
Meanwhile, the inoculation of parental 4T1 cells caused 60% of lung
metastasis, higher than that of parental cells, but lower than that
of 4T1-M1 cells. Representative bioluminescent images for lung
metastasis in the mice are shown in Fig. 5B, and representative histological
images of metastatic tumors in lung sections are presented in
Fig. 5C. These results clearly
demonstrated that 4T1-M1 cells gained significantly enhanced
propensity to metastasize to mouse lungs, and 4T1-P1 cells
reversely lost the capability to metastasize. The parental 4T1
cells presumably stayed in between with both properties of primary
and metastatic tumors.
Enhanced epithelial-mesenchymal
transition (EMT) for metastatic sublines
To identify the molecular mechanism underlying the
elevated propensity of the metastatic sublines, we investigated the
possible induction of EMT in these cells. EMT defines the
conversion of epithelial cells into migratory and invasive
mesenchymal cells. EMT is an inherent natural process during
development for the generation of tissues and organs; however, it
is an unintentional process in cancer progression, associated with
enhanced capabilities in resistance to senescence upon chemotherapy
and immunotherapy, stem cell properties, invasion and metastasis
(17,18). Snail1, the founding member of the
Snail superfamily of zinc-finger transcription factors, plays a
central role in the regulation of EMT (19). Snail1’s EMT program is driven by a
complex network involving several targets that are associated with
reduction in cell adhesion, increase in migratory capacity,
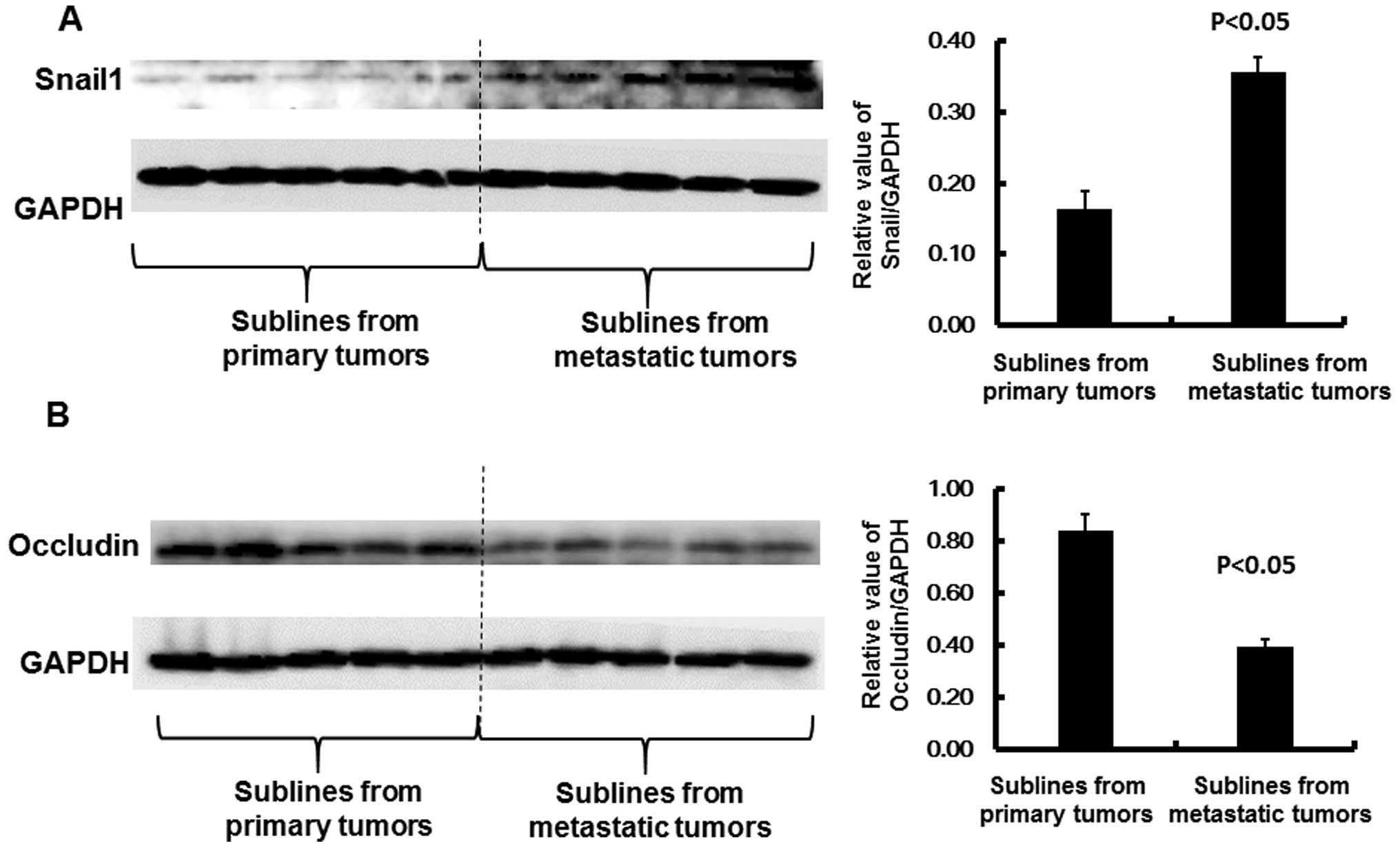
drug/immune resistance and gain of stem cell properties (20). As shown in Fig. 6A, a significant induction in Snail1
was observed in the 4T1-M cells, compared to the 4T1-P cells, as
shown by western blot analysis (P<0.05). This observation
suggested that metastatic propensity was greatly enhanced in the
4T1-M cells, relative to the 4T1-P cells. In support of this
observation, one of the Snail1 targets, occludin was accordingly
suppressed in the 4T1-M cells, compared to that in the 4T1-P cells
(Fig. 6B, P<0.05). Consistent
with our finding, a previous study revealed that the repression of
occludin is due to the presence of Snail1 (21). Snail1 substantially inhibits the
promoter activity of occludin by binding to the E-box located in
its promoter region. Thus, Snail1 serves as a transcriptional
repressor of occludin (22). The
reduction in occludin results in the loss of cell-to-cell adhesion
that is necessary for the EMT process (22). Therefore, these results demonstrated
that enhanced EMT could be a primary mechanism underlying the
metastatic propensity of the 4T1-M cells.
Discussion
To date, although the mechanisms underlying breast
cancer metastasis remain elusive, metastasis-specific treatments
would surely yield more effective therapeutic outcomes for breast
cancer patients with metastatic tumors. Thus, the study of
metastasis is critical. A pronounced impediment to this research is
the lack of appropriate cell models. On the one hand, human breast
cancer cells reveal poor metastasis and unstable metastatic
features in immunocompromised mice. The current xenograft models
therefore show limited ability to reflect the actual metastatic
process (23). On the other hand,
the interspecies use of human cells in mice may not be able to
faithfully represent the complex biological process, particularly
at the molecular level (24,25).
In contrast, murine breast cancer cells exhibit similar metastatic
properties in mice to those as reported in breast cancer patients
(26). In terms of mouse models,
the 4T1 mouse model of breast cancer development and metastasis has
been repeatedly used in a wide array of studies for more than 20
years. This cell line was originally established as a spontaneous
breast cancer model in mice with the BALB/c genetic background
(11). When implanted in syngeneic
mice, the parental 4T1 cell line displays limited and infrequent
metastasis to local lymph nodes and distant organs (7,11). To
address this issue, in the present study, we established cell
sublines of 4T1 cells with significantly metastatic propensity and
specific tropisms to mouse lungs. To the best of our knowledge,
this is a unique orthotopic model of mouse breast cancer metastasis
to mouse lungs. These sublines would be beneficial to recognize the
molecular events (such as gene expression signatures) responsible
for mechanisms that drive the progression of metastasis.
As a main cause of cancer-related death, metastasis
involves a serial of events: escape of cancer cells from a primary
tumor, spread via the circulation and eventually colonization at
distant sites, where some tumor cells acquire dormant or actively
dividing phenotypes (27). EMT is
considered as a driving force of breast cancer metastasis, as EMT
changes cytoskeleton remodeling from apical-basolateral polarities
indicative of epithelial cells to the generation of actin stress
fibers characteristic of mesenchymal cells (18). As a result, EMT leads to the
production of invasive and migratory tumor cells, with stem
cell-like and chemoresistant properties (18,28).
In support of a contributive role of EMT in metastasis, clinical
studies in patients with breast cancers indicate that enhanced
concentration of mesenchymal markers in tumor cells is more
predictive of poor prognosis than epithelial cytokeratin markers
alone (29–33). EMT is a complex process involving a
large number of genes, of which Snail1 plays a crucial role in this
event (20,27). The primary contribution of Snail1 in
EMT is to regulate the expression of genes necessary for EMT
including the repression of E-cadherin and occludin, leading to
compromised cell adhesion and enforced migratory capacity (20). Increased cell motility and
invasiveness in a primary tumor would certainly contribute to the
dissemination of cancer cells to circulation and distant organs
(34). E-cadherin is a
transmembrane adhesion glycoprotein, which is exclusively expressed
in epithelial cells (35). For
example, E-cadherin is highly expressed in normal mammary ductal
epithelial cells, whereas its expression is diminished in
metastatic breast cancer cells (36). E-cadherin crucially governs
calcium-dependent intercellular adhesion under normal settings
(37). However, its expression is
abnormally altered in cancer cells, and reduced expression
oftentimes represents an invasive and metastatic phenotype
(38–40). Previous studies indicate that
aberrant expression of E-cadherin is associated with more invasive
and metastatic breast cancers. Thus, reduced expression of
E-cadherin has been reported to be associated with the development
of distant metastases in patients with breast cancer, and
E-cadherin has been recognized as a tumor-suppressor (38–40).
Occludin is an integral transmembrane protein with four hydrophobic
domains, and it is crucial for the integrity of tight junctions
(41). Occludin is an important
component at tight junctions in epithelial and endothelial cells
(41), and loss of occludin causes
loose cell-to-cell adhesion that would accelerate the EMT process
during cancer progression (20). In
the present study, Snail1 was identified to be greatly induced in
the metastatic sublines, compared to cell lines from primary
tumors. As the master regulator in EMT, its downstream target
occludin was significantly repressed in metastatic sublines,
compared to the control cell lines. These changes would greatly
induce EMT and contribute to metastasis, supporting the enhanced
propensity of metastasis for the metastatic sublines.
To summarize, we established a novel orthotopic
model of breast cancer metastasis to mouse lungs. Metastatic
sublines of 4T1 cells were characterized with substantial
metastatic propensity to lungs, and these cells also displayed
enhanced EMT features. This novel model allows detailed
investigation of the molecular bases responsible for site-specific
metastasis of breast cancer to the lungs. The present study also
opens an emerging avenue for therapeutic studies in a more
clinically relevant model.
Acknowledgments
This study was supported by a grant under the
National ‘973’ Program (grant no. 2014CB932000), the Strategic
Priority Research Program of the Chinese Academy of Sciences (grant
no. XDB14000000), and grants from the National Natural Science
Foundation of China (grant nos. 21377159, 21177151 and 21321004).
We thank all the laboratory members for their great assistance with
the experiments and reagents.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis C, Siegel R, Bandi P and Jemal A:
Breast cancer statistics, 2011. CA Cancer J Clin. 61:409–418. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lluch A, Cueva J, Ruiz-Borrego M and Ponce
J: Pérez-Fidalgo JA Zoledronic acid in the treatment of metastatic
breast cancer. Anticancer Drugs. 25:1–7. 2014. View Article : Google Scholar
|
|
4
|
Hayashi K, Aono M, Shintam K and Kazuki K:
Bisphosphonate-related atypical femoral fracture with bone
metastasis of breast cancer Case report and review. Anticancer Res.
34:1245–1249. 2014.PubMed/NCBI
|
|
5
|
Minn AJ, Gupta GP, Padua D, Bos P, Nguyen
DX, Nuyten D, Kreike B, Zhang Y, Wang Y, Ishwaran H, et al: Lung
metastasis genes couple breast tumor size and metastatic spread.
Proc Natl Acad Sei USA. 104:6740–6745. 2007. View Article : Google Scholar
|
|
6
|
Minn AJ, Gupta GP, Siegel PM, Bos PD, Shu
W, Gin DD, Viale A, Olshen AB, Gerald WL and Massagué J: Genes that
mediate breast cancer metastasis to lung. Nature. 436:518–524.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aslakson CJ and Miller FR: Selective
events in the metastatic process defined by analysis of the
sequential dissemination of subpopulations of a mouse mammary
tumor. Cancer Res. 52:1399–1405. 1992.PubMed/NCBI
|
|
8
|
Tao K, Fang M, Alroy J and Sahagian GG:
Imagable 4T1 model for the study of late stage breast cancer. BMC
Cancer. 8:2282008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pulaski BA and Ostrand-Rosenberg S:
Reduction of established spontaneous mammary carcinoma metastases
following immunotherapy with major histocompatibility complex class
II and B71 cell-based tumor vaccines. Cancer Res. 58:1486–1493.
1998.PubMed/NCBI
|
|
10
|
Eckhardt BL, Parker BS, van Laar RK,
Restall CM, Natoli AL, Tavaria MD, Stanley KL, Sloan EK, Moseley JM
and Anderson RL: Genomic analysis of a spontaneous model of breast
cancer metastasis to bone reveals a role for the extracellular
matrix. Mol Cancer Res. 3:1–13. 2005.PubMed/NCBI
|
|
11
|
Miller FR, Miller BE and Heppner GH:
Characterization of metastatic heterogeneity among subpopulations
of a single mouse mammary tumor Heterogeneity in phenotypic
stability. Invasion Metastasis. 3:22–31. 1983.
|
|
12
|
Liu S, Goldstein RH, Scepansky EM and
Rosenblatt M: Inhibition of rho-associated kinase signaling
prevents breast cancer metastasis to human bone. Cancer Res.
69:8742–8751. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu S, Li S and Du Y: Polychlorinated
biphenyls (PCBs) enhance metastatic properties of breast cancer
cells by activating Rho-associated kinase (ROCK). PLoS One.
5:ell2722010.
|
|
14
|
Brocks C, Graefe H, Frenzel H, Pries R and
Wollenberg B: Isolation of human myeloid dendritic cells from tumor
tissue and peripheral blood. In Vivo. 20:239–242. 2006.PubMed/NCBI
|
|
15
|
Padua D, Zhang XH, Wang Q, Nadal C, Gerald
WL, Gomis RR and Massagué J: TGFbeta primes breast tumors for lung
metastasis seeding through angiopoietin-like 4. Cell. 133:66–77.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lelekakis M, Moseley JM, Martin TJ, Hards
D, Williams E, Ho P, Lowen D, Javni J, Miller FR, Slavin J, et al:
A novel orthotopic model of breast cancer metastasis to bone. Clin
Exp Metastasis. 17:163–170. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nieto MA: The ins and outs of the
epithelial to mesenchymal transition in health and disease. Annu
Rev Cell Dev Biol. 27:347–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Foubert E, De Craene B and Berx G: Key
signalling nodes in mammary gland development and cancer The
Snaill-Twistl conspiracy in malignant breast cancer progression.
Breast Cancer Res. 12:2062010. View
Article : Google Scholar
|
|
20
|
Kaufhold S and Bonavida B: Central role of
Snail1 in the regulation of EMT and resistance in cancer: A target
for therapeutic intervention. J Exp Clin Cancer Res. 33:622014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kajita M, McClinic KN and Wade PA:
Aberrant expression of the transcription factors snail and slug
alters the response to genotoxic stress. Mol Cell Biol.
24:7559–7566. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ikenouchi J, Matsuda M, Furuse M and
Tsukita S: Regulation of tight junctions during the
epithelium-mesenchyme transition: Direct repression of the gene
expression of claudins/occludin by Snail. J Cell Sci.
116:1959–1967. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bibby MC: Orthotopic models of cancer for
preclinical drug evaluation: Advantages and disadvantages. Eur J
Cancer. 40:852–857. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Parsons JT, Zetter B and Mohla S: Shifting
paradigms in tumor metastasis: Challenges and opportunities. Cancer
Biol Ther. 1:582–585. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Goldstein RH, Weinberg RA and Rosenblatt
M: Of mice and (wo)men: Mouse models of breast cancer metastasis to
bone. J Bone Miner Res. 25:431–436. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vernon AE, Bakewell SJ and Chodosh LA:
Deciphering the molecular basis of breast cancer metastasis with
mouse models. Rev Endocr Metab Disord. 8:199–213. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ansieau S: EMT in breast cancer stem cell
generation. Cancer Lett. 338:63–68. 2013. View Article : Google Scholar
|
|
29
|
Gradilone A, Raimondi C, Nicolazzo C,
Petracca A, Gandini O, Vincenzi B, Naso G, Aglianò AM, Cortesi E
and Gazzaniga P: Circulating tumour cells lacking cytokeratin in
breast cancer: The importance of being mesenchymal. J Cell Mol Med.
15:1066–1070. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Roxanis I: Occurrence and significance of
epithelial-mesenchymal transition in breast cancer. J Clin Pathol.
66:517–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Taube JH, Herschkowitz JI, Komurov K, Zhou
AY, Gupta S, Yang J, Hartwell K, Onder TT, Gupta PB, Evans KW, et
al: Core epithelial-to-mesenchymal transition interactome
gene-expression signature is associated with claudin-low and
metaplastic breast cancer subtypes. Proc Natl Acad Sci USA.
107:15449–15454. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hennessy BT, Gonzalez-Angulo AM,
Stemke-Hale K, Gilcrease MZ, Krishnamurthy S, Lee JS, Fridlyand J,
Sahin A, Agarwal R, Joy C, et al: Characterization of a naturally
occurring breast cancer subset enriched in
epithelial-to-mesenchymal transition and stem cell characteristics.
Cancer Res. 69:4116–4124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Herschkowitz JI, Simin K, Weigman VJ,
Mikaelian I, Usary J, Hu Z, Rasmussen KE, Jones LP, Assefnia S,
Chandrasekharan S, et al: Identification of conserved gene
expression features between murine mammary carcinoma models and
human breast tumors. Genome Biol. 8:R762007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu S: The ROCK signaling and breast
cancer metastasis. Mol Biol Rep. 38:1363–1366. 2011. View Article : Google Scholar
|
|
35
|
Twiss F and de Rooij J: Cadherin
mechanotransduction in tissue remodeling. Cell Mol Life Sci.
70:4101–4116. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schneider MR and Kolligs FT: E-cadherin’s
role in development, tissue homeostasis and disease: Insights from
mouse models: Tissue-specific inactivation of the adhesion protein
E-cadherin in mice reveals its functions in health and disease.
Bioessays. 37:294–304. 2015. View Article : Google Scholar
|
|
37
|
Nelson WJ, Dickinson DJ and Weis WI: Roles
of cadherins and catenins in cell-cell adhesion and epithelial cell
polarity. Prog Mol Biol Transl Sci. 116:3–23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jeanes A, Gottardi CJ and Yap AS:
Cadherins and cancer: How does cadherin dysfunction promote tumor
progression? Oncogene. 27:6920–6929. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gheldof A and Berx G: Cadherins and
epithelial-to-mesenchymal transition. Prog Mol Biol Transl Sci.
116:317–336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Farahani E, Patra HK, Jangamreddy JR,
Rashedi I, Kawalec M, Rao Pariti RK, Batakis P and Wiechec E: Cell
adhesion molecules and their relation to (cancer) cell stemness.
Carcinogenesis. 35:747–759. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Furuse M, Hirase T, Itoh M, Nagafuchi A,
Yonemura S and Tsukita S and Tsukita S: Occludin: A novel integral
membrane protein localizing at tight junctions. J Cell Biol.
123:1777–1788. 1993. View Article : Google Scholar : PubMed/NCBI
|