Introduction
Gastric cancer (GC) remains the third leading cause
of cancer-related death in both genders worldwide and causes over
723,000 deaths worldwide per year (1,2). Even
with the improvement in diagnostic techniques and advancement in
therapeutic modalities, the 5-year survival rates among patients
are still disappointing, and one of the predominant causes is
metastasis (3–5). Peritoneal metastasis, as a common mode
of metastasis for advanced GC and a common feature of the natural
history of GC, is the most frequent cause of death in patients with
advanced GC (6). It is necessary to
investigate the molecular mechanisms of GC metastasis and to
develop new strategies to prevent GC peritoneal metastasis.
Phosphatase of regenerating liver-3 (PRL-3), a
member of the PRL family of protein tyrosine phosphatases, plays an
important role in GC metastasis (7). Researchers have demonstrated that
PRL-3 induces cancer-associated phenotypes including proliferation,
migration, invasion, tumorigenesis and metastasis (8–11),
which also arise in GC (12). There
are also various experiments showing that the catalytic activity of
PRL-3 is directly involved in tumor invasion, metastasis and tumor
angiogenesis (10,13). In our previous study, we found that
in primary GC with peritoneal metastasis, PRL-3 expression was
significantly higher than that in the corresponding primary GC and
PRL-3 expression was negatively correlated with prognosis in GC
patients (14). In addition,
various studies have also indicated that upregulation of PRL-3
expression correlates with GC invasion and metastasis (15–17).
Furthermore, in our previous study, we also found that miR-495 and
miR-551a both act as tumor-suppressors by targeting the PRL-3
oncogene and inhibiting GC cell migration and invasion (18). However, the downstream mechanism by
which the signaling pathway facilitates PRL-3-promoted motility,
invasion and metastasis of GC cells remains unknown, and warrants
further research.
Phosphatase and tensin homologue (PTEN; deleted on
chromosome 10) is a tumor-suppressor gene featuring
dual-specificity phosphatase activities (19,20).
It plays crucial roles in maintaining normal cell activities and
functions (19,20). PTEN is a lipid phosphatase that
catalyzes the conversion of
phosphatidylinositol-3,4,5,-trisphosphate (PIP3) to
phosphatidylinositol-4,5-bisphosphate (PIP2) (21), which can induce apoptosis and
suppress proliferation. Loss of PTEN function leads to excessive
PIP3 accumulation at the plasma membrane and subsequent
derepression of the phosphatidylinositol-3-kinase/protein kinase B
(PI3K)/Akt pathway, which in turn stimulates cell growth,
proliferation, survival and other cellular processes (22,23).
The PI3K/Akt signaling pathway is an important
driver of cell proliferation and survival (24). PI3K signaling has been demonstrated
to be involved in GC progression, including cell growth,
metabolism, survival, metastasis and resistance to chemotherapy
(25–28). PRL-3 most likely lies upstream of
PI3K, as treatment with the PI3K inhibitor LY294002 abrogated
PRL-3-mediated AKT activation (29,30).
Additionally, PRL-3 overexpression downregulated the expression of
PTEN in DLD-1 colorectal carcinoma (29) and melanoma cells (31). Since PTEN is the most important
negative regulator of the PI3K/Akt pathway, PTEN downregulation may
be an important means of PRL-3-driven PI3K/Akt signaling, although
PI3K activation by other mechanisms may exist.
Before cancer cells metastasize to distant organs,
they must disseminate from the site of the primary tumor (32). Extracellular matrix degradation is
an essential step in tumor invasion and metastasis. Matrix
metalloproteinases (MMPs), including MMP-2 and MMP-9, are capable
of cleaving cell-surface proteins as well as degrading components
of the extracellular matrix, allowing migratory cells to invade
neighboring tissues and break through the basement membrane
(33). In a previous study, PRL-3
was found to promote the motility, invasion and metastasis of LoVo
colon cancer cells through PRL-3-integrin β1-ERK1/2 and -MMP2
signaling (34). PRL-3 also
enhanced MMP-2 secretion and cellular invasiveness via activation
of mTOR in the human colon cancer HCT116 cell line (35).
In consideration of the crucial role of PRL-3 in GC
metastasis and the emerging character of PTEN in GC, we speculated
that PRL-3 regulates the expression or post-translational
modification of PTEN and then activates the PI3K/Akt signaling
pathway, upregulating MMP-2/MMP-9 expression, further to promote GC
peritoneal metastasis by its phosphatase enzyme activity. In the
present study, we described that PRL-3 was overexpressed in GC with
peritoneal metastasis and in GC lines with high enzyme activity,
while PTEN was weakly expressed in GC with peritoneal metastasis
and in GC cell lines. The present study also showed that only PRL-3
had the enzyme activity to downregulate the expression of PTEN and
increased the ratio of p-PTEN/PTEN. We also found that PRL-3
activated the PI3K/Akt signaling pathway and further upregulated
the expression of MMP-2 and MMP-9 to promote GC peritoneal
metastasis.
Materials and methods
Tissue samples
All human gastric tissue samples including 21
specimens of normal gastric mucosa, 49 specimens of GC without
peritoneal metastasis and 23 specimens of GC with peritoneal
metastasis were obtained from the Department of General Surgery of
The First Affiliated Hospital of Nanchang University (Nanchang,
China). All specimens were immediately frozen in liquid nitrogen
and were stored at −80°C until further use. Informed consent was
obtained from all patients, and the present study was approved by
the institutional review committee. None of the patients received
radiotherapy or chemotherapy before the operation. Histological
diagnosis was confirmed for each specimen. All of the samples were
obtained with informed consent for molecular analysis and were
approved by the Ethics Committee of the The First Affiliated
Hospital of Nanchang University.
Cell culture
The human gastric mucosal cell line GES-1 was
preserved in our central laboratory. Human gastric adenocarcinoma
cancer cell lines SGC7901, MKN28, MGC803, AGS and MKN45 were
obtained from the Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China). Cells were cultured at 37°C/5%
CO2 in Dulbecco's modified Eagle's medium (DMEM;
Biological Industries, Israel), supplemented with 10% fetal bovine
serum (FBS) (MRC, China), 100 U/ml penicillin and 100 µg/ml
streptomycin.
Construction of a wild-type plasmid
EGFP-PRL-3 and a mutant-type plasmid EGFP-PRL-3 (D72A/C104S) and
transfection into SGC7901 cells
We constructed a wild-type plasmid EGFP-PRL-3 with
the enzyme activity of PRL-3 and a mutant-type plasmid EGFP-PRL-3
(D72A/C104S) containing an inactivating mutation of the essential
catalytic cytokine residue to serine at position 104 in the
phosphate active site according to related studies (13,36,38).
All of the vectors were designed and synthesized by GenePharma
Corporation (Shanghai, China). After being confirmed by DNA
sequencing, the EGFP-PRL-3 and the EGFP-PRL-3 (D72A/C104S) vectors
were transfected into SGC7901 cells using Lipofectamine 2000
reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. After screening, the
SGC7901 cancer cells stably expressed PRL-3; the wild-type
EGFP-PRL-3 with the enzyme activity was the wild-type group and the
mutant-type EGFP-PRL-3 (D72A/C104S) was the mutant-type without
enzyme activity. The enzyme activity of PRL-3 was detected by a
phosphate activity detection kit (Beyotime, Shanghai,
China/Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's instructions and confirmed that the two groups met
the requirements (39–42).
Construction of the EGFP-PRL-3 and
PRL-3-RNAi plasmids and construction of stable cell lines
To construct the EGFP-PRL-3 plasmid, human PRL-3 was
amplified by RT-PCR using the primers:
5′-TACCGGACTCAGATCTCGAGCGCCACCATGGCTCGGATGAACCGC-3′ and
5′-GATCCCGGGCCCGCGGTACCGTCATAACGCAGCACCGGGTCT-3′ and was cloned
into the GV230 vector. To acquire the generation of PRL-3 knockdown
in the SGC7901 cells, we constructed three candidate human PRL-3
siRNA target sequences and a blank plasmid control vector. All of
the vectors were designed and synthesized by GeneChem Corporation
(Shanghai, China). The PRL-3 shRNA target sequences were as
follows: 5′-GCTCACCTACCTGGAGAAA-3′ [PRL-3-RNAi (7330)],
5′-ACAGAGGCTGCGGTTCAAA-3′ [PRL-3-RNAi (7331)],
5′-CCTGTTCTCGGCACCTTAA-3′ [PRL-3-RNAi (7332)],
5′-GGATCTCGTTCTCCTCATT-3′ [PRL-3-RNAi (7333)]. All the vectors were
transfected into SGC7901 cells using Lipofectamine 2000 reagent
according to the manufacturer's protocol. We selected PRL-3-RNAi
(7332) with the highest efficacy as the suitable vector after
evaluating the interference efficiency by western blotting. After
screening, we constructed the stable expression cell lines. The
SGC7901/EGFP-PRL-3 cell line was the PRL-3 high expression group
and the SGC7901/PRL-3-RNAi cell line was the PRL-3 low expression
group.
Construction of the PTEN-knockdown
plasmid and transfection into the PRL-3 low expression cell
line
For the RNA interference of human PTEN, three
specific small interfering RNAs were used. The shRNA target
sequences were: 5′-GCGCTATGTGTATTATTAT-3′ [PTEN-RNAi (17260)],
5′-ACAGCTAGAACTTATCAAA-3′ [PTEN-RNAi (17261)],
5′-AGAACTTATCAAACCCTTT-3′ [PTEN-RNAi (17262)],
5′-TGCAGATAATGACAAGGAA-3′ [PTEN-RNAi (17263)], respectively. All of
the vectors were designed and synthesized by GeneChem. All of the
vectors were transfected into SGC7901 cells using Lipofectamine
2000 reagent. After screening, we constructed the stable expression
cell lines and selected the high interfering efficiency PTEN-RNAi
(17261) group.
Quantitative real-time RT-PCR
analyses
The detailed steps of RNA extraction and qRT-PCR
were described in our previous study (18). TRIzol reagent was purchased from
Invitrogen. Applied Biosystems 7500 Fast Real-Time PCR System and
SYBR-Green PCR Master Mix were purchased from Applied Biosystems
(Foster City, CA, USA), Taq PCR Master Mix was purchased
from TransGen (Beijing, China). 7900HT Fast Real-Time PCR System
was obtained from Applied Biosystems.
Western blot analysis
We prepared samples for western blotting as
described in our previous study (43). Total protein extraction and BCA
protein assay kits were both purchased from KeyGen (Nanjing,
China). Polyvinylidene fluoride (PVDF) membranes were purchased
from Millipore (Bedford, MA, USA). PRL-3 antibody [anti-PTP4A3
(ab50276)] was purchased from Abcam (Cambridge, UK) which
recognizes human PRL-3 and does not cross-react with human PRL-1
and PRL-2. PRL-3 antibody (ab50276) was also used in previous
studies (44–47). The following antibodies of PTEN,
Akt, MMP-2, MMP-9, p-Akt and p-PTEN were also purchased from Abcam.
The antibody for β-actin was purchased from Proteintech Group, Inc.
(Wuhan, China). ECL western blotting analysis system was obtained
from TransGen Biotech (Beijing, China).
Cell migration and invasion assays
Cell migration and invasion assays were performed as
previously described (18).
Transwell cell culture chambers (8.0-µm pore polycarbonate
membranes) were purchased from Becton-Dickinson (San Diego, CA,
USA). The fluorescence microscope was purchased from Olympus
(Center Valley, PA, USA) and was used to capture the images.
Statistical analysis
Each experiment was repeated at least three times.
All of the data are presented as the mean ± SD. Statistical
analysis was performed using SPSS 19.0 computer software (SPSS,
Inc., Chicago, IL, USA) and software program GraphPad Prism.
Differences among variables were assessed by χ2
analysis, Student's t-test or ANOVA. In all cases, a value of
P<0.05 was accepted as significant.
Results
Overexpression of PRL-3 is inversely
proportional to the low expression of PTEN in GC tissues
To investigate the relationship between PRL-3 and
PTEN expression in GC tissues, we analyzed the expression of PRL-3
and PTEN in 21 specimens of normal gastric mucosa, 49 specimens of
GC without peritoneal metastasis, and 23 specimens of GC with
peritoneal metastasis by qRT-PCR and western blot analysis. PRL-3
was overexpressed in the GC samples with peritoneal metastasis when
compared with the level in normal gastric mucosa both at the mRNA
(data not shown) and the protein level (Fig. 1A and B). This indicates that high
expression of PRL-3 is closely related to the tumor stage of GC.
However, there was no significant statistical difference in the
mRNA level of PTEN among the three types of gastric tissues,
implying that the downregulation of PTEN expression was a
post-transcriptional event. We also observed that weak to strong
expression of PTEN protein was noted in GC with peritoneal
metastasis, GC without peritoneal metastasis, and normal gastric
mucosa by western blotting (Fig. 1C and
D). The p-PTEN expression level was also lower in normal
gastric mucosa than that noted in GC with peritoneal metastasis.
Moreover, we conducted further analysis of the data by western blot
analysis. It revealed that the p-PTEN and PTEN ratio was higher in
GC cells when compared with the ratio in the normal gastric mucosal
cells (Fig. 1E). Furthermore, we
used an enzyme activity detection kit to detect the phosphatase
activity of PRL-3. From these data, the enzyme activity of PRL-3
was more active in GC with peritoneal metastasis than that noted in
the normal gastric mucosa (Fig.
1F). All of the data indicated that low expression of PTEN may
be inversely correlated to both the high enzyme activity and the
high expression of PRL-3. The high enzyme activity of PRL-3 may
increase the phosphorylation level of PTEN.
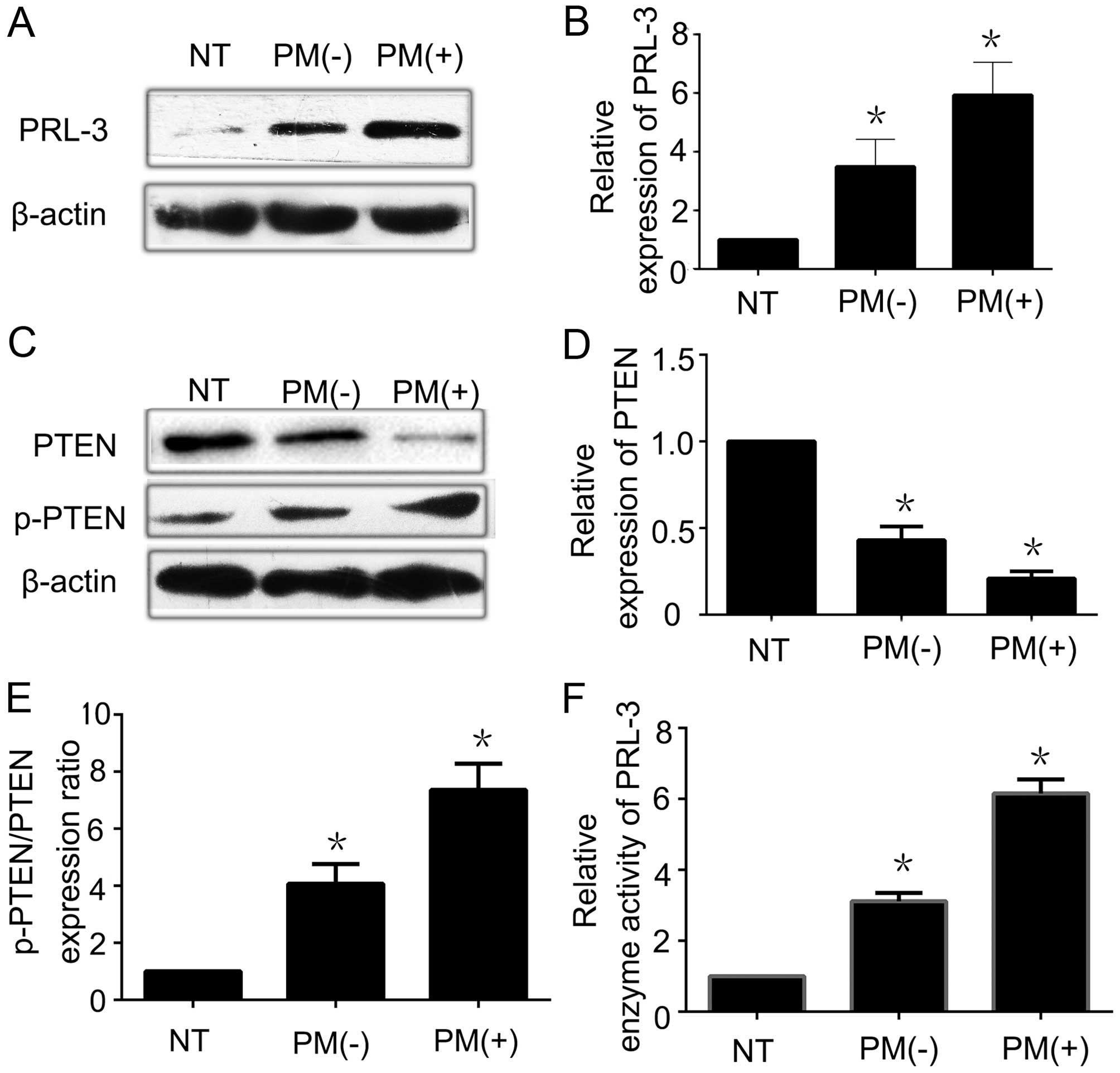 | Figure 1Overexpression of PRL-3 is inversely
proportional to the low expression of PTEN in gastric cancer
tissues. (A) PRL-3 was overexpressed in GC with peritoneal
metastasis by western blotting. (B) Relative expression of PRL-3
protein in 21 specimens of normal gastric mucosa, 49 specimens of
GC without peritoneal metastasis, and 23 specimens of GC with
peritoneal metastasis was analyzed by western blot analysis. (C)
PTEN was weakly expressed in GC with peritoneal metastasis when
compared with that in the normal gastric mucosal tissues. p-PTEN
expression was weakly expressed in the normal gastric mucosal
tissues by western blotting. (D) Relative expression of PTEN
protein in 21 specimens of normal gastric mucosa, 49 specimens of
GC without peritoneal metastasis, and 23 specimens of GC with
peritoneal metastasis was analyzed by western blot analysis. (E)
Relative p-PTEN/PTEN protein expression ratio in 21 specimens of
normal gastric mucosa, 49 specimens of GC without peritoneal
metastasis, and 23 specimens of GC with peritoneal metastasis was
analyzed by western blot analysis. (F) Relative enzyme activity of
PRL-3 in 21 specimens of normal gastric mucosa, 49 specimens of GC
without peritoneal metastasis, and 23 specimens of GC with
peritoneal metastasis was analyzed by enzyme activity detection
kit. Each bar shows the mean ± standard deviation from three
independent experiments; *P<0.05 vs. the normal
gastric mucosa tissues. NT, normal gastric mucosa tissues; PM(−),
GC without peritoneal metastasis; PM(+), GC with peritoneal
metastasis. |
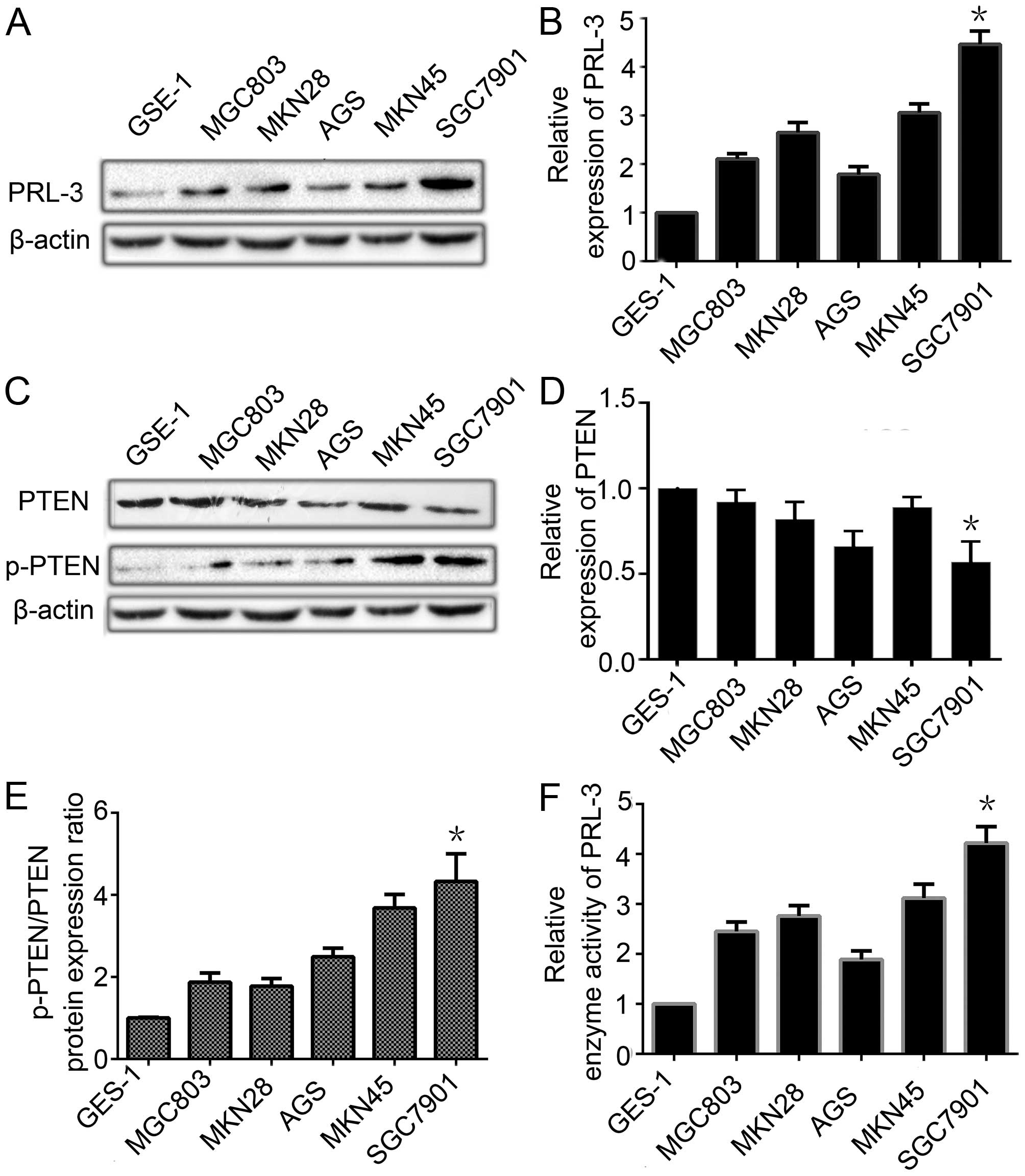
PRL-3 is overexpressed is contrast to the
low expression of PTEN in GC cell lines
To explore the relationship between the expression
of PTEN and PRL-3, we analyzed these levels in one normal human
gastric mucosal cell line GES-1 and five GC cell lines of different
metastatic potential (MGC803, MKN28, AGS, MKN45 and SGC7901) by
qRT-PCR and western blotting. The expression level of PRL-3 in the
five GC cell lines was much higher than that in the GES-1 cell line
(Fig. 2A and B). The PTEN
expression was lower in the five GC cell lines than that in normal
gastric mucosal cell line at the mRNA and protein level (Fig. 2C and D). The p-PTEN/PTEN ratio was
higher in GC cells than that in the normal gastric mucosal cell
line (Fig. 2E). We also detected
the enzyme activity level of PRL-3 by an enzyme activity detection
kit. PRL-3 phosphatase activity was higher in the GC cell lines
(Fig. 2F). The results indicated
that PRL-3, through its phosphatase activity, led to the low
expression of PTEN and phosphorylated it to facilitate the
development of peritoneal metastasis by GC. As PRL-3 decreased the
expression of PTEN and increased the ratio of p-PTEN/PTEN to a
greater extent in the SGC7901 cells, we chose the SGC7901 GC cell
line to perform the next experiment.
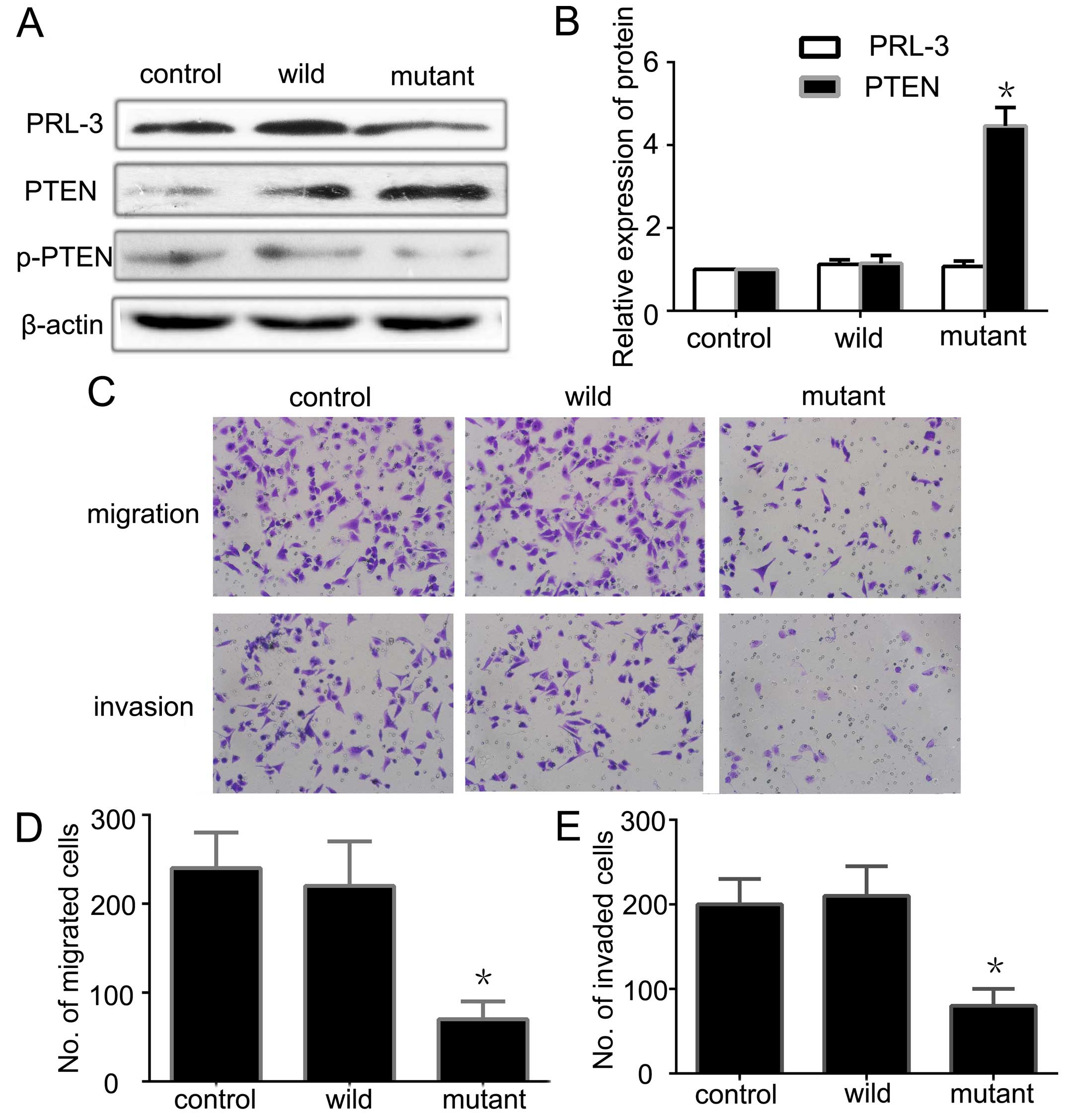
PRL-3 downregulates the expression of
PTEN through its phosphatase activity
To further investigate whether or not PRL-3 through
its enzyme activity downregulates PTEN, we constructed a wild-type
plasmid EGFP-PRL-3 and a mutant-type plasmid EGFP-PRL-3
(D72A/C104S) without the enzyme activity of PRL-3. After
transfecting the EGFP-PRL-3, EGFP-PRL-3 (D72A/C104S) and PLNCX
vector (control empty plasmid) into SGC7901 cells and screening the
stably expressed cell lines, we detected the phosphatase activity
of PRL-3 using the enzyme activity detection kit. The wild-type
with enzyme activity and the mutant-type without enzyme activity
met our requirements. Next, we detected the expression of PRL-3 and
PTEN by qRT-PCR and western blotting. There was no significant
difference between the wild-type and the mutant-type in terms of
PRL-3 expression, but both were higher than the control group
(Fig. 3A). PTEN expression was
lower in the wild-type than the level in the mutant-type, while the
ratio of p-PTEN/PTEN was higher in the wild-type (Fig. 3A and B). This shows that high
expression of PRL-3 downregulates the expression of PTEN and
increases the ratio of p-PTEN/PTEN. Further analysis indicated that
the higher enzyme activity of PRL-3 had an opposite relationship
with the expression of PTEN. PRL-3 increased the ratio of
p-PTEN/PTEN and promoted PTEN phosphorylation and inactivated it
through its phosphatase activity. We also analyzed the migration
and invasion abilities of the different groups by Transwell assay.
The mutant-type had less ability to promote GC cell migration and
invasion than the wild-type (Fig.
3C–E). All of the observations suggest that PRL-3 may
downregulate PTEN and induce PTEN inactivity through its enzyme
activity to promote GC cell metastasis.
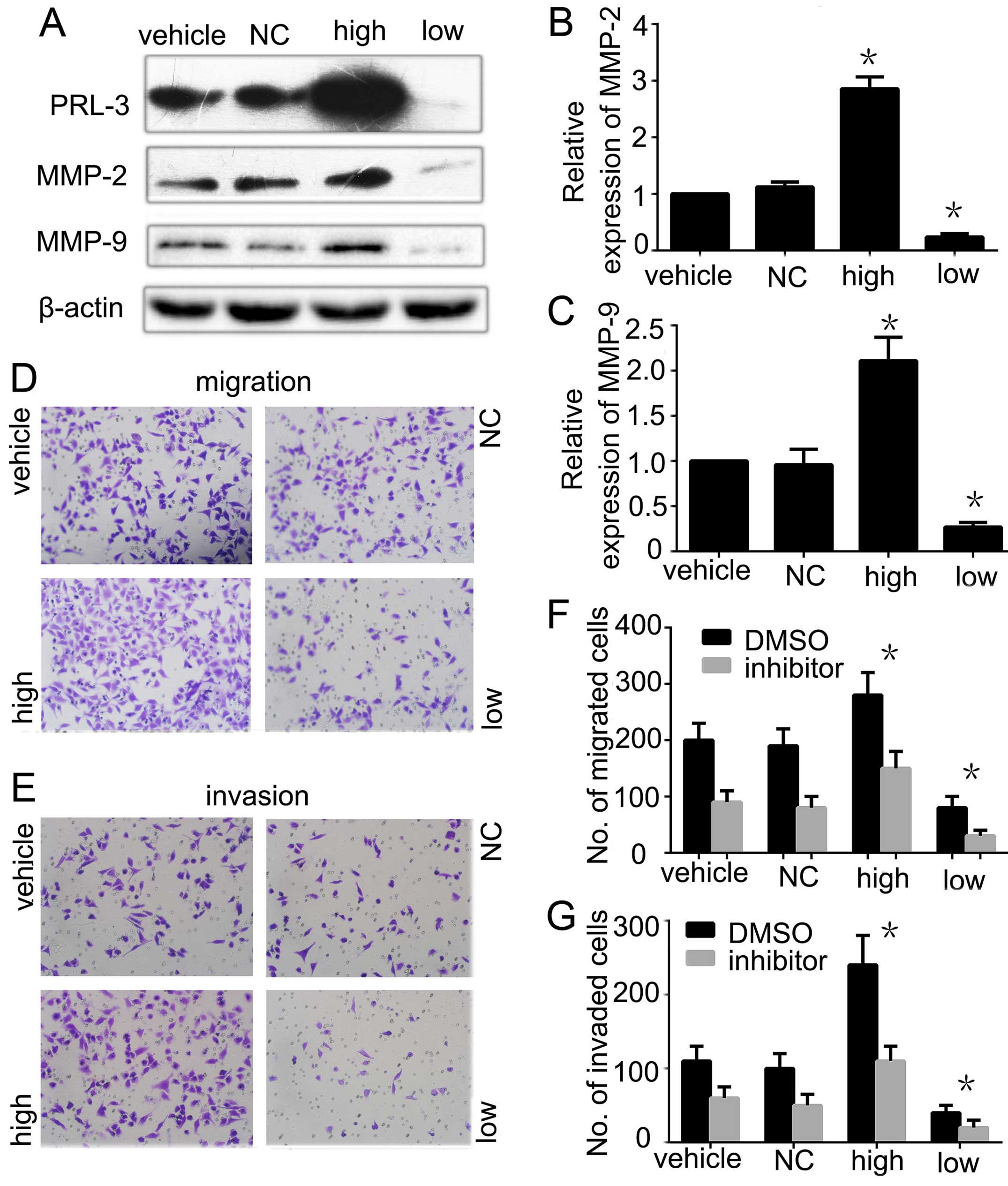
Matrix metallopeptidases (MMP-2 and
MMP-9) are involved in the invasion and migration of GC cells
regulated by PRL-3
To dissect the mechanism of MMPs in PRL-3-promoted
GC cell migration and invasion, we further analyzed the expression
of MMP-2 and MMP-9 in PRL-3 high expression and PRL-3 low
expression groups by western blot analysis. The phosphatase
activity of PRL-3 was lower in the PRL-3 low expression group than
the SGC7901 cell group and PRL-3 high expression group as detected
by the enzyme activity detection kit. The expression levels of
MMP-2 and MMP-9 were consistently higher in the high group than
that in the SGC7901 cells. When we silenced PRL-3 in the low
expression group, the expression levels of MMP-2 and MMP-9 were
reduced as expected (Fig. 4A–C). We
also evaluated the invasion and metastasis in the different groups.
Invasion and metastasis were significantly inhibited in the PRL-3
low expression group with low expression of MMP-2 and MMP-9
compared with the control group (Fig.
4D and E). To further confirm that PRL-3 promotes GC metastasis
by upregulating the expression of MMP-2 and MMP-9, we added 20
µM MMP-2/MMP-9 inhibitor I into the different groups before
performing the migration and invasion assays. The data showed that
MMP-2/MMP-9 inhibitor I inhibited the PRL-3-promoted invasion and
metastasis abilities of the SGC7901 cells (Fig. 4F and G). In conclusion, all the
results indicated that PRL-3 promoted GC cell migration and
invasion by upregulating the expression of MMP-2 and MMP-9.
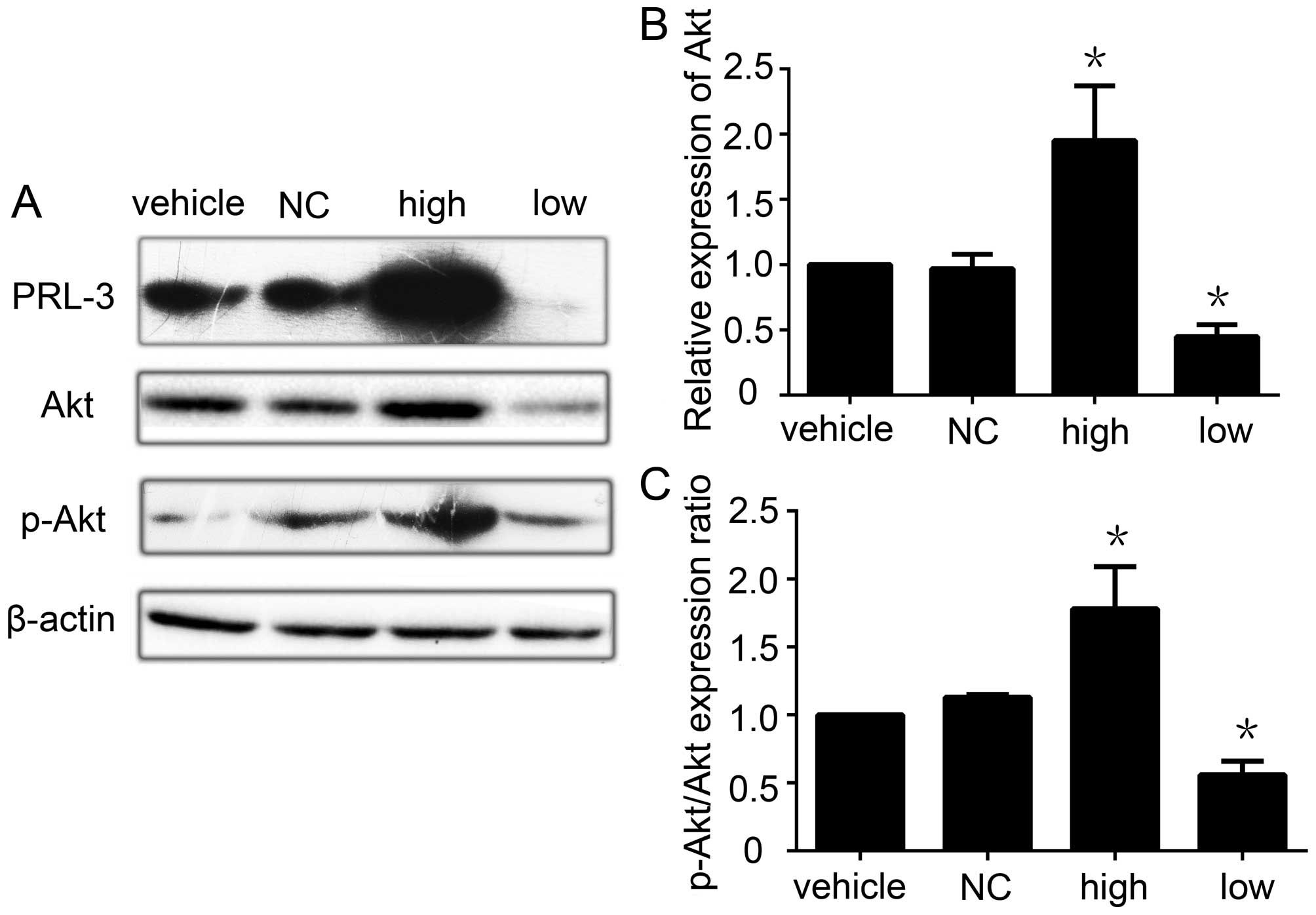
The Akt pathway is involved in the
upregulation of MMP-2 and MMP-9 and the invasion and migration of
GC cells mediated by PRL-3
The PI3K/Akt pathway plays a critical role in the
control of tumor cell growth, survival, progression, apoptosis,
invasion and metastasis, and AKT is the central mediator of the
PI3K/Akt pathway (48). Since
researchers have reported that PRL-3 activates the PI3K/Akt pathway
to promote cancer cell invasion and metastasis (30), we next detected the effect of PRL-3
expression on AKT activation in GC. PRL-3 increased the expression
of Akt in the PRL-3 high expression group. When we knocked down
PRL-3 by RNAi, the expression of Akt was also decreased (Fig. 5A and B). We also found that the
p-Akt/Akt ratio was higher in the PRL-3 high expression group than
that in the control group. Following knockdown of PRL-3 in the
SGC7901 GC cell line, the ratio of p-Akt/Akt was also decreased
(Fig. 5C). When we added the PI3K
inhibitor LY294002 into the PRL-3 high expression group, the
effects of PRL-3 on Akt, MMP-2, MMP-9 was blocked, with concomitant
inhibition of invasion and metastasis (data not shown). All of the
data show that PRL-3 upregulates the PI3K/Akt signaling pathway to
promote GC metastasis.
Ablation of PTEN expression restores the
migration/invasion phenotype of PRL-3-silenced cancer cells
To further confirmed that the reduced migration and
invasion were caused by the downregulation of PTEN by PRL-3, we
next tested whether knockdown of PTEN in PRL-3-silenced cancer
cells (the PRL-3 low expression group) reactivated the
PI3K/Akt/MMP-2/MMP-9 signaling pathway and restored their
migration/invasion phenotype. As expected, silencing of PTEN with
RNAi in the PRL-3 low expression group restored the expression of
PTEN and the Akt expression (Fig. 6A
and B). Meanwhile, the ratio of p-Akt/Akt and the expression of
MMP-2 and MMP-9 were also restored (Fig. 6A and B). Furthermore, the cell
migration and invasion ability was also partly restored in the
PRL-3 low expression group when we transfected the PTEN-RNAi vector
into the cells (Fig. 6C–F). These
results suggested that PRL-3 decreases the expression of PTEN to
promote GC migration and invasion.
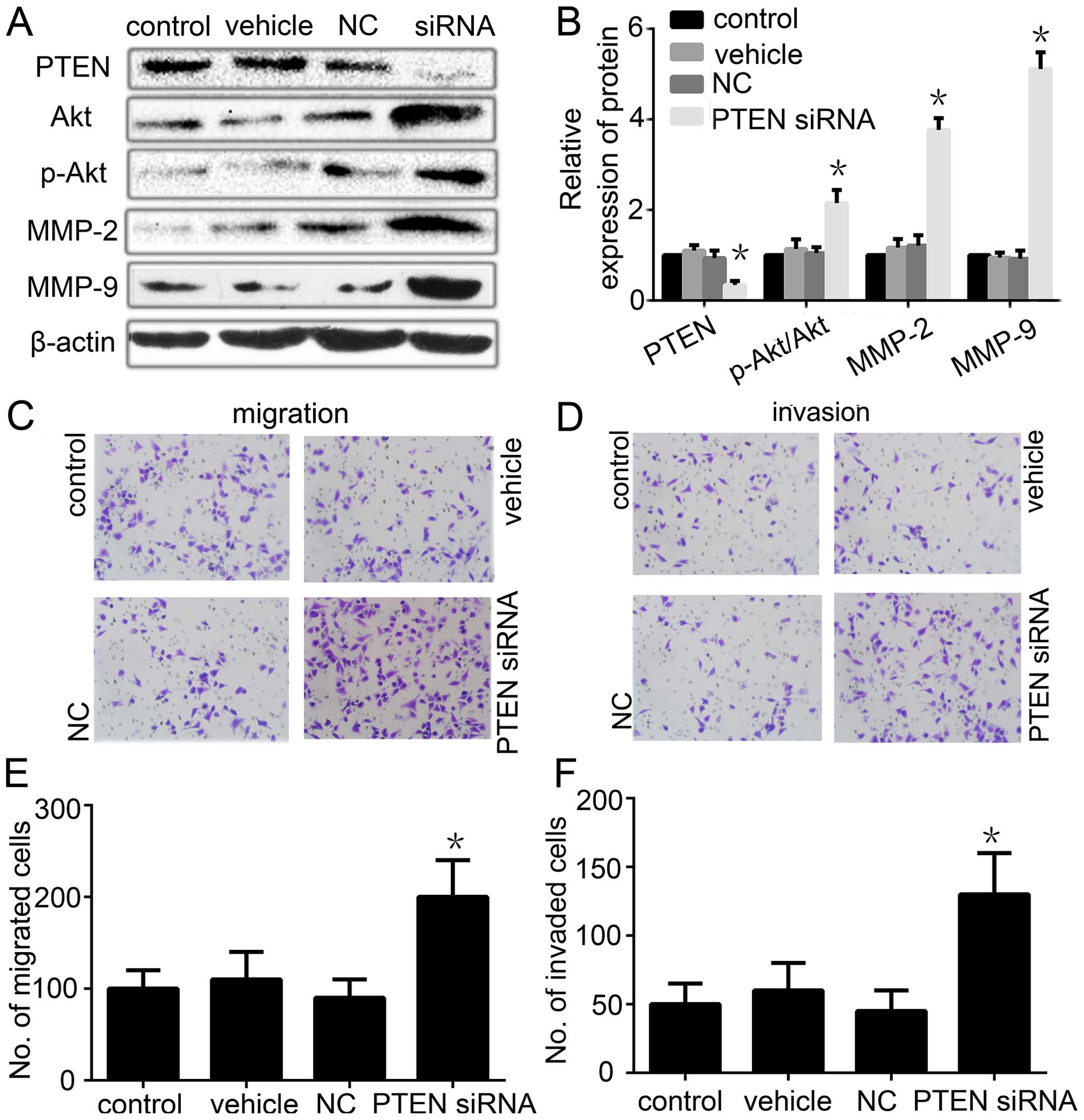 | Figure 6Ablation of PTEN expression restores
the migration/invasion phenotype in PRL-3-silenced cancer cells.
(A) The expression of PTEN, Akt, p-Akt, MMP-2 and MMP-9 was
analyzed by western blotting after PTEN RNAi was transfected into
the PRL-3 stable low expression cells. (B) Relative expression of
PTEN, Akt, p-Akt, MMP-2 and MMP-9 in A was determined as a ratio to
β-actin. (C) Cell migration and (D) invasion were respectively
analyzed. Original magnification, ×200. (E) Migration and (F)
invasion cell numbers are indicated in the bar graph. Each value is
the number of stained cells. Control, PRL-3 low expression SGC7901
cells; vehicle, PRL-3 low expression SGC7901 cells (only used
Lipofectamine 2000 reagent); NC, PRL-3 low expression SGC7901 cells
transfected with empty control plasmid; siRNA, PRL-3 low expression
SGC7901 cells transfected with PTEN siRNA. Control and vehicle
groups were used as a control. The experiments were repeated at
least three times independently. Each bar shows the mean ± standard
deviation from three independent experiments;
*P<0.05. |
Discussion
Phosphatase of regenerating liver-3 (PRL-3) is a
metastasis-promoting phosphatase, which has been reported to
promote the metastasis of a variety of cells, including gastric
cancer (GC) SGC7901 (14,49), colon cancer LoVo (50), salivary adenoid cystic carcinoma
SACC-83 (8) and lung cancer cells
(42). In our previous study, we
found that PRL-3 was highly expressed in GC (14), and miR-495 and miR-551a both act as
tumor-suppressors by targeting the PRL-3 oncogene and inhibiting GC
cell migration and invasion (18).
In the present study, we also found that PRL-3 was overexpressed in
GC tissues with peritoneal metastasis when compared with that in
normal gastric tissues. High expression of PRL-3 was also found in
GC cells that had a high degree of metastatic potential, as
compared with corresponding normal cells. The results were in
accordance with previous research studies (16,51,52).
PTEN is a tumor-suppressor gene that occupies a key
position in the regulation of cell growth, proliferation,
apoptosis, mobility and signal transduction (53), while its expression and activity can
be regulated at almost all levels: transcriptional, translational
and post-translational (54). The
function of PTEN is commonly lost in a large proportion of human
cancers through somatic mutations, gene silencing or epigenetic
mechanisms (55). In a previous
study, researchers found that PRL-3 downregulated PTEN in the DLD-1
colorectal cell line (29). From
our data, the PTEN mRNA expression was not statistically different
between the GC tissues or the normal gastric mucosal tissues.
However, the PTEN protein expression was lower in GC with
peritoneal metastasis than that noted in normal gastric mucosal
tissues. The present study also found that there was a higher level
of PTEN phosphorylation in GC tissues than that noted in the normal
gastric tissues. It showed that the downregulation of PTEN
expression was a post-transcriptional regulation event. We also
found the same phenomenon in the GC cell lines and the gastric
mucosa cell line GES-1. The enzyme activity of PRL-3 was higher in
GC with peritoneal metastasis, as well as in the GC cell lines,
than that in the normal gastric mucosa and the gastric mucosal cell
line GES-1. All of the data indicate that PRL-3 may downregulate
the expression of PTEN and induce PTEN phosphorylation through its
enzyme activity.
To further explore the relationship between the
enzyme activity of PRL-3 and downregulation of PTEN expression and
high level phosphorylation of PTEN, we constructed a wild-type
plasmid EGFP-PRL-3 and a mutant-type plasmid EGFP-PRL-3
(D72A/C104S) without enzyme activity and transfected the plasmids
into the SGC7901 cells. PRL-3 with high enzyme activity
significantly downregulated PTEN expression and induced PTEN
phosphorylation compared to the no enzyme activity cells. All of
the data indicated that PRL-3 downregulated the expression of PTEN
and induced PTEN phosphorylation to inactivite it through its
phosphatase activities. Meanwhile, silencing of PTEN with RNAi in
PRL-3-silenced cells restored the cell migration and invasion
ability, which further demonstrated that PRL-3 through its
phosphatase activity downregulated PTEN, which led to GC cell
migration and invasion. However, the mechanism behind the PRL-3
downregulation of PTEN remains unknown. As a phosphatase of PRL-3,
it is almost not possible to induce PTEN phosphorylation. We
speculate that, on one hand, PRL-3 may decrease the expression of
PTEN. On the other hand, PRL-3 may through its enzyme activity
dephosphorylate a certain kinase and improve its enzyme activity,
and then the kinase phosphorylates PTEN and makes it inactive. All
of the speculations need further validation.
Metastasis is a key step in cancer progression that
indicates a more advanced stage and a poorer prognosis. Tumor cells
escape from the primary site and migrate into the circulatory
system which are important steps in cancer metastasis (56). Extracellular matrix degradation
plays an important role in tumor invasion and metastasis, which is
mainly mediated by MMP-2 and MMP-9 (57). In our previous study, we found that
overexpression of PRL-3 promoted GC invasion and metastasis
(14). Thus, it was necessary to
investigate whether the involvement of PRL-3 in invasion and
migration is also correlated with MMP-2 and MMP-9 in GC. The data
indicated that PRL-3 increased the expression of MMP-2/MMP-9 in the
PRL-3 high expression group, while silencing of PRL-3 with RNAi
resulted in reduction of MMP-2/MMP-9 expression. Moreover,
MMP-2/MMP-9 inhibitor I obstructed the migration and invasion
abilities in the PRL-3 high expression group. These data suggest
that PRL-3 may upregulate MMP-2 and MMP-9 expression leading to GC
metastasis. Of course, it may be more appropriate to detect the
expression of MMP-2/MMP-9 in secreted media (35), and not only in cellular lysates.
This is a limitation of the present study.
PI3K signaling is a crucial regulator of many
essential cellular processes, including cell growth, metabolism,
survival, metastasis and resistance to chemotherapy (58). PI3K activity is regulated by the
lipid phosphatase and tensin homolog (PTEN), a tumor-suppressor
gene that encodes a lipid phosphatase that downregulates the PI3K
signal by converting PIP3 back to PIP2 (59). Loss of PTEN function, as well as
PI3K activation, results in accumulation of PI3P triggering the
activation of its downstream effection (60). In the present study, we found that
PRL-3 downregulated PTEN, phosphorylated PTEN and inactivated it.
However, whether the PI3K/Akt signaling pathway is also involved in
the promotion of GC metastasis by PRL-3, also needs to be
investigated. In our research, PRL-3 overexpression in the PRL-3
high expression group increased the expression of Akt and increased
the ratio of p-Akt/Akt, accompanied by upregulation of MMP-2/MMP-9
expression and promoted the GC cell migration and invasion. The
expression levels of Akt, MMP-2 and MMP-9 were blocked by the PI3K
inhibitor LY294002. These data suggest that PRL-3 may signal
through the PI3K/Akt pathway to upregulate MMP-2/MMP-9 leading to
migration and invasion of GC.
In summary, we identified that PRL-3 downregulates
PTEN and induces PTEN phosphorylation then activates, at least
partly, the PI3K/Akt signaling pathway, as well as upregulation of
MMP-2 and MMP-9, to promote the peritoneal metastasis of GC.
However, whether or not PRL-3 directly downregulates PTEN protein
remains unknown, and warrants further investigation. In conclusion,
the present study revealed the potential mechanism of PRL-3-mediate
GC invasion and metastasis, which will provide new strategies for
the prevention and treatment of GC peritoneal metastasis.
Acknowledgments
The present study was supported by the National
Science Foundation of China (no. 81160304/81360362), the Education
Department of Jiangxi Province Science and Technology Research
Projects (no. GJJ13126), the Training Program for Young Scientists
of Jiangxi Province (no. 20133BCB23028), and the Graduate Student
Innovation Special Fund Project of Nanchang University (no.
cx2015189). We thank the Department of Gastrointestinal Surgery of
The First Affiliated Hospital of Nanchang University for providing
human gastric tissue samples. We also thank The Key Laboratory of
Basic Pharmacology of Nanchang University and the Department of
Histology and Embryology of Medical College of Nanchang University
for technical assistance in the experiments.
References
|
1
|
Globocan: 2012, Estimated Cancer
Incidence, Mortality and Prevalence Worldwide in 2012. http://globocan.iarc.fr/Pages/fact_sheets_cancer.aspx.
Accessed March 31, 2014.
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Vita F, Vecchione L, Galizia G, Di
Martino N, Fabozzi T, Catalano G, Ciardiello F and Orditura M:
Perspectives in adjuvant therapy of gastric cancer. Oncology.
77(Suppl 1): S38–S42. 2009. View Article : Google Scholar
|
|
4
|
Kagawa S, Shigeyasu K, Ishida M, Watanabe
M, Tazawa H, Nagasaka T, Shirakawa Y and Fujiwara T: Molecular
diagnosis and therapy for occult peritoneal metastasis in gastric
cancer patients. World J Gastroenterol. 20:17796–17803.
2014.PubMed/NCBI
|
|
5
|
Bando E, Yonemura Y, Takeshita Y,
Taniguchi K, Yasui T, Yoshimitsu Y, Fushida S, Fujimura T,
Nishimura G and Miwa K: Intraoperative lavage for cytological
examination in 1,297 patients with GC. Am J Surg. 178:256–262.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Turaga KK, Gamblin TC and Pappas S:
Surgical treatment of peritoneal carcinomatosis from gastric
cancer. Int J Surg Oncol. 2012:4056522012.PubMed/NCBI
|
|
7
|
Bessette DC, Qiu D and Pallen CJ: PRL
PTPs: Mediators and markers of cancer progression. Cancer
Metastasis Rev. 27:231–252. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong Q, Ding X, Chang B, Wang H and Wang
A: PRL-3 promotes migration and invasion and is associated with
poor prognosis in salivary adenoid cystic carcinoma. J Oral Pathol
Med. 45:111–118. 2016. View Article : Google Scholar
|
|
9
|
Kato H, Semba S, Miskad UA, Seo Y, Kasuga
M and Yokozaki H: High expression of PRL-3 promotes cancer cell
motility and liver metastasis in human colorectal cancer: A
predictive molecular marker of metachronous liver and lung
metastases. Clin Cancer Res. 10:7318–7328. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo K, Li J, Tang JP, Koh V, Gan BQ and
Zeng Q: Catalytic domain of PRL-3 plays an essential role in tumor
metastasis: Formation of PRL-3 tumors inside the blood vessels.
Cancer Biol Ther. 3:945–951. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu X, Zeng H, Zhang X, Zhao Y, Sha H, Ge
X, Zhang M, Gao X and Xu Q: Phosphatase of regenerating liver-3
promotes motility and metastasis of mouse melanoma cells. Am J
Pathol. 164:2039–2054. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dai N, Lu AP, Shou CC and Li JY:
Expression of phosphatase regenerating liver 3 is an independent
prognostic indicator for gastric cancer. World J Gastroenterol.
15:1499–1505. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng Q, Dong JM, Guo K, Li J, Tan HX, Koh
V, Pallen CJ, Manser E and Hong W: PRL-3 and PRL-1 promote cell
migration, invasion, and metastasis. Cancer Res. 63:2716–2722.
2003.PubMed/NCBI
|
|
14
|
Li ZR, Wang Z, Zhu BH, He YL, Peng JS, Cai
SR, Ma JP and Zhan WH: Association of tyrosine PRL-3 phosphatase
protein expression with peritoneal metastasis of GC and prognosis.
Surg Today. 37:646–651. 2007. View Article : Google Scholar
|
|
15
|
Xing X, Lian S, Hu Y, Li Z, Zhang L, Wen
X, Du H, Jia Y, Zheng Z, Meng L, et al: Phosphatase of regenerating
liver-3 (PRL-3) is associated with metastasis and poor prognosis in
GC. J Transl Med. 11:309. 2013. View Article : Google Scholar
|
|
16
|
Miskad UA, Semba S, Kato H and Yokozaki H:
Expression of PRL-3 phosphatase in human gastric carcinomas: Close
correlation with invasion and metastasis. Pathobiology. 71:176–184.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z, Cai SR, He YL, Zhan WH, Chen CQ,
Cui J, Wu WH, Wu H, Song W, Zhang CH, et al: High expression of
PRL-3 can promote growth of gastric cancer and exhibits a poor
prognostic impact on patients. Ann Surg Oncol. 16:208–219. 2009.
View Article : Google Scholar
|
|
18
|
Li Z, Cao Y, Jie Z, Liu Y, Li Y, Li J, Zhu
G, Liu Z, Tu Y, Peng G, et al: miR-495 and miR-551a inhibit the
migration and invasion of human gastric cancer cells by directly
interacting with PRL-3. Cancer Lett. 323:41–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Waite KA and Eng C: Protean PTEN: Form and
function. Am J Hum Genet. 70:829–844. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bermúdez Brito M, Goulielmaki E and
Papakonstanti EA: Focus on PTEN regulation. Front Oncol. 5:1662015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ye B, Jiang LL, Xu HT, Zhou DW and Li ZS:
Expression of PI3K/AKT pathway in gastric cancer and its blockade
suppresses tumor growth and metastasis. Int J Immunopathol
Pharmacol. 25:627–636. 2012.PubMed/NCBI
|
|
22
|
Myers MP, Pass I, Batty IH, Van der Kaay
J, Stolarov JP, Hemmings BA, Wigler MH, Downes CP and Tonks NK: The
lipid phosphatase activity of PTEN is critical for its tumor
supressor function. Proc Natl Acad Sci USA. 95:13513–13518. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stambolic V, Suzuki A, de la Pompa JL,
Brothers GM, Mirtsos C, Sasaki T, Ruland J, Penninger JM,
Siderovski DP and Mak TW: Negative regulation of PKB/Akt-dependent
cell survival by the tumor suppressor PTEN. Cell. 95:29–39. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cully M, You H, Levine AJ and Mak TW:
Beyond PTEN mutations: The PI3K pathway as an integrator of
multiple inputs during tumorigenesis. Nat Rev Cancer. 6:184–192.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wadhwa R, Song S, Lee JS, Yao Y, Wei Q and
Ajani JA: Gastric cancer-molecular and clinical dimensions. Nat Rev
Clin Oncol. 10:643–655. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li NA, Wang W, Xu B and Gong H: miR-196b
regulates gastric cancer cell proliferation and invasion via
PI3K/AKT/mTOR signaling pathway. Oncol Lett. 11:1745–1749.
2016.PubMed/NCBI
|
|
27
|
Akrami H, Mahmoodi F, Havasi S and Sharifi
A: PlGF knockdown inhibited tumor survival and migration in gastric
cancer cell via PI3K/Akt and p38MAPK pathways. Cell Biochem Funct.
34:173–180. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun L, Liu L, Liu X, Wang Y, Li M, Yao L,
Yang J, Ji G, Guo C, Pan Y, et al: MGr1-Ag/37LRP induces cell
adhesion-mediated drug resistance through FAK/PI3K and MAPK pathway
in gastric cancer. Cancer Sci. 105:651–659. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang H, Quah SY, Dong JM, Manser E, Tang
JP and Zeng Q: PRL-3 down-regulates PTEN expression and signals
through PI3K to promote epithelial-mesenchymal transition. Cancer
Res. 67:2922–2926. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang H, Vardy LA, Tan CP, Loo JM, Guo K,
Li J, Lim SG, Zhou J, Chng WJ, Ng SB, et al: PCBP1 suppresses the
translation of metastasis-associated PRL-3 phosphatase. Cancer
Cell. 18:52–62. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fang XY, Song R, Chen W, Yang YY, Gu YH,
Shu YQ, Wu XD, Wu XF, Sun Y, Shen Y, et al: PRL-3 promotes the
malignant progression of melanoma via triggering dephosphorylation
and cytoplasmic localization of NHERF1. J Invest Dermatol.
135:2273–2282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: A tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Peng L, Xing X, Li W, Qu L, Meng L, Lian
S, Jiang B, Wu J and Shou C: PRL-3 promotes the motility, invasion,
and metastasis of LoVo colon cancer cells through PRL-3-integrin
beta1-ERK1/2 and-MMP2 signaling. Mol Cancer. 8:1102009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ye Z, Al-Aidaroos AQ, Park JE, Yuen HF,
Zhang SD, Gupta A, Lin Y, Shen HM and Zeng Q: PRL-3 activates
mTORC1 in cancer progression. Sci Rep. 5:170462015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li WJ, Xing XF, Qu LK, Meng L and Shou CC:
Construction and expression of PRL-3 plasmid with C104S point
mutation and CAAX deletion. Beijing Da Xue Xue Bao. 41:516–520.
2009.In Chinese. PubMed/NCBI
|
|
37
|
Pascaru M, Tanase C, Vacaru AM, Boeti P,
Neagu E, Popescu I and Szedlacsek SE: Analysis of molecular
determinants of PRL-3. J Cell Mol Med. 13:3141–3150. 2009.
View Article : Google Scholar
|
|
38
|
Kozlov G, Cheng J, Ziomek E, Banville D,
Gehring K and Ekiel I: Structural insights into molecular function
of the metastasis-associated phosphatase PRL-3. J Biol Chem.
279:11882–11889. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
McParland V, Varsano G, Li X, Thornton J,
Baby J, Aravind A, Meyer C, Pavic K, Rios P and Köhn M: The
metastasis-promoting phosphatase PRL-3 shows activity toward
phosphoinositides. Biochemistry. 50:7579–7590. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Matter WF, Estridge T, Zhang C, Belagaje
R, Stancato L, Dixon J, Johnson B, Bloem L, Pickard T, Donaghue M,
et al: Role of PRL-3, a human muscle-specific tyrosine phosphatase,
in angiotensin-II signaling. Biochem Biophys Res Commun.
283:1061–1068. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fiordalisi JJ, Keller PJ and Cox AD: PRL
tyrosine phosphatases regulate rho family GTPases to promote
invasion and motility. Cancer Res. 66:3153–3161. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jian M, Nan L, Guocheng J, Qingfu Z,
Xueshan Q and Enhua W: Downregulating PRL-3 inhibit migration and
invasion of lung cancer cell via RhoA and mDia1. Tumori.
98:370–376. 2012.PubMed/NCBI
|
|
43
|
Li Z, Zhan W, Wang Z, Zhu B, He Y, Peng J,
Cai S and Ma J: Inhibition of PRL-3 gene expression in gastric
cancer cell line SGC7901 via microRNA suppressed reduces peritoneal
metastasis. Biochem Biophys Res Commun. 348:229–237. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu Y, Zheng P, Liu Y, Ji T, Liu X, Yao S,
Cheng X, Li Y, Chen L, Xiao Z, et al: An epigenetic role for PRL-3
as a regulator of H3K9 methylation in colorectal cancer. Gut.
62:571–581. 2013. View Article : Google Scholar
|
|
45
|
Molleví DG, Aytes A, Padullés L,
Martínez-Iniesta M, Baixeras N, Salazar R, Ramos E, Figueras J,
Capella G and Villanueva A: PRL-3 is essentially overexpressed in
primary colorectal tumours and associates with tumour
aggressiveness. Br J Cancer. 99:1718–1725. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mayinuer A, Yasen M, Mogushi K, Obulhasim
G, Xieraili M, Aihara A, Tanaka S, Mizushima H, Tanaka H and Arii
S: Upregulation of protein tyrosine phosphatase type IVA member 3
(PTP4A3/PRL-3) is associated with tumor differentiation and a poor
prognosis in human hepatocellular carcinoma. Ann Surg Oncol.
20:305–317. 2013. View Article : Google Scholar :
|
|
47
|
Khapare N, Kundu ST, Sehgal L, Sawant M,
Priya R, Gosavi P, Gupta N, Alam H, Karkhanis M, Naik N, et al:
Plakophilin3 loss leads to an increase in PRL3 levels promoting K8
dephosphorylation, which is required for transformation and
metastasis. PLoS One. 7:e385612012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang S, Zhang Y, Zhuang Y, Wang J, Ye J,
Zhang S, Wu J, Yu K and Han Y: Matrine induces apoptosis in human
acute myeloid leukemia cells via the mitochondrial pathway and Akt
inactivation. PLoS One. 7:e468532012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bessette DC, Wong PC and Pallen CJ: PRL-3:
A metastasis-associated phosphatase in search of a function. Cells
Tissues Organs. 185:232–236. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xu H, Lai W, Zhang Y, Liu L, Luo X, Zeng
Y, Wu H, Lan Q and Chu Z: Tumor-associated macrophage-derived IL-6
and IL-8 enhance invasive activity of LoVo cells induced by PRL-3
in a KCNN4 channel-dependent manner. BMC Cancer. 14:3302014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Miskad UA, Semba S, Kato H, Matsukawa Y,
Kodama Y, Mizuuchi E, Maeda N, Yanagihara K and Yokozaki H: High
PRL-3 expression in human gastric cancer is a marker of metastasis
and grades of malignancies: An in situ hybridization study.
Virchows Archiv. 450:303–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Z, He YL, Cai SR, Zhan WH, Li ZR, Zhu
BH, Chen CQ, Ma JP, Chen ZX, Li W, et al: Expression and prognostic
impact of PRL-3 in lymph node metastasis of gastric cancer: Its
molecular mechanism was investigated using artificial microRNA
interference. Int J Cancer. 123:1439–1447. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xu W, Yang Z, Zhou SF and Lu N:
Posttranslational regulation of phosphatase and tensin homolog
(PTEN) and its functional impact on cancer behaviors. Drug Des
Devel Ther. 8:1745–1751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bononi A and Pinton P: Study of PTEN
subcellular localization. Methods. 77–78:92–103. 2015. View Article : Google Scholar
|
|
55
|
Milella M, Falcone I, Conciatori F, Cesta
Incani U, Del Curatolo A, Inzerilli N, Nuzzo CM, Vaccaro V, Vari S,
Cognetti F, et al: PTEN: Multiple functions in human malignant
tumors. Front Oncol. 5:242015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Su Z, Yang Z, Xu Y, Chen Y and Yu Q:
Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol
Cancer. 14:482015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Khasigov PZ, Podobed OV, Gracheva TS,
Salbiev KD, Grachev SV and Berezov TT: Role of matrix
metalloproteinases and their inhibitors in tumor invasion and
metastasis. Biochemistry. 68:711–717. 2003.PubMed/NCBI
|
|
58
|
Willems L, Tamburini J, Chapuis N, Lacombe
C, Mayeux P and Bouscary D: PI3K and mTOR signaling pathways in
cancer: New data on targeted therapies. Curr Oncol Rep. 14:129–138.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ortega-Molina A and Serrano M: PTEN in
cancer, metabolism, and aging. Trends Endocrinol Metab. 24:184–189.
2013. View Article : Google Scholar
|
|
60
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|