Introduction and structure of GPC-3
The glypicans, a family of proteins classified as
HSPGs, have been shown to interact with a number of growth factors
and modulate growth factor activity (1) and are linked to the extracellular side
of cell membrane by a glycosylphosphatidylinositol (GPI) anchor
(2). Members of this large family
of transmembrane proteins have been identified in both mammals and
drosophila: 6 glypicans (GPC-1 through GPC-6) in mammals, and two
others in the fly (2). Glypicans
have a core protein size of ~60–70 kDa and express an N-terminal
secretory signal peptide along with a hydrophobic domain that is
used for the addition of the GPI anchor at the C-terminus. Another
characteristic that is shared by all glypicans is the location of
the insertion sites for the heparan sulfate chains (HSC), which
seems to be restricted to the last 50 amino acids in the
C-terminus, placing the chains close to the cell membrane.
Additionally, the position of 14 cysteine residues is conserved,
further strengthening the structural relationship of the proteins
within the family (2).
Much of the literature on GPC-3 stems from its
proposed role in vivo as well as in vitro in cancer
models. Multiple studies have shown a correlation between GPC-3 and
hepatocellular carcinoma (HCC) due to the nature of the protein
being expressed during normal cellular growth and movement in
hepatocytes (1,3–6). In
patients who received surgical treatment for HCC, a decrease in
GPC-3 protein was observed (3). In
addition, in cells derived from hepatocellular carcinoma GPC-3 has
been observed to be secreted into culture media (7). GPC-3 does not seems to be expressed by
other liver cell types (hepatic stellate cells, kupffer cells, bile
duct cells, endothelial cells and fibroblasts) but it appears to be
expressed exclusively by hepatocytes. It is well known only that
its expression from hepatocytes influences the activity and
function of non-parenchymal liver cells.
GPC-3 was initially hypothesized to interact with
insulin-like growth factor in selected cell types, though this
notion has been challenged as this may not be the case in all cell
types (8). In a study done on the
loss-of-function mutation of GPC-3 in embryonic development,
researchers observed an overgrowth of organs in fetal and postnatal
development, known as Simpson-Golabi-Behmel overgrowth syndrome
(9). This evidence cast GPC-3 into
a strict role as a tumor suppressor. In another study looking at
the metastatic adenocarcinoma mammary LM3 cell line, GPC-3 was
shown to inhibit the canonical Wnt signals involved in cell
proliferation and survival. In the same study, GPC-3 was also
observed to play a role in activating the non-canonical Wnt
pathway, which directs cell morphology and migration (8). Furthermore, GPC-3 was also found to
act as a potential tumor suppressor in lung tissue (10). This has re-opened the case of GPC-3
as a strict suppressor of cell growth and introduced the
possibility of its role in stimulating growth and even
tumorigenesis in a tissue-dependent manner in some cell
lineages.
This being said, certain key elements have been
observed which give reason for upregulation of GPC-3 in certain
cell types and not others. GPC-3 is able to bind Wnt and Hedgehog
(Hh) signaling proteins and was also shown to have the ability to
bind basic growth factors such as fibroblast growth factor 2 (FGF2)
through its heparan-sulfate glycan chains (11). Additionally, expression of sulfatase
2 (SULF2), another tumor marker expressed in hepatocytes, was found
to be correlated with increased expression of GPC-3. When GPC-3 was
decreased, FGF2 binding was decreased in SULF2 expressing
hepatocellular carcinoma cells. SULF2 expression in resected HCC
tissue was also shown to have a worse prognosis and a higher rate
of recurrence after surgery (12).
Herein we synthesize the literature on GPC-3,
including its structure, expression in normal tissues, as well its
role as a serum marker as well as possible therapeutic target. The
review also includes current knowledge on GPC-3 expression as a
prognostic marker in HCC and its role in liver regeneration post
partial hepatectomy (PH). In addition, we discuss embryonic and
adult tissue expression of GPC-3. Lastly, we report the
interactions of GPC-3 with CD81 as CD81 has been shown to interact
with GPC-3 as a binding partner (13,14).
Expression pattern of GPC-3 in human and
mouse
Embryonic expression of GPC-3
Embryonic expression of GPC-3 differs greatly from
the expression map seen in health adult tissues. In most embryonic
tissues and organs, there is positive expression of GPC-3 as
illustrated by immunohistochemistry (IHC) in humans and in
situ hybridization (ISH) in mice. Organs or tissues that
display positive expression in human embryonic tissues include:
tissues of the digestive tract, the gonads, kidney, limb buds,
liver, nerve tissue, oral cavity, pancreas, respiratory syndrome,
tongue, and vertebrae (15). In
pancreatic tissue, positive expression was seen in exocrine glands,
while endocrine glands showed negative expression. In contrast, IHC
revealed negative expression of GPC-3 in human embryonic skin
tissue (15). Additional detail
regarding location and description of GPC-3 expression in human
embryonic tissue can be seen in Table
I.
 | Table I.GPC-3 expression in human and mouse
embryonic tissue. |
Table I.
GPC-3 expression in human and mouse
embryonic tissue.
| Organ | Embryonic human
staining (IHC) (15) | Embryonic mouse
tissue (ISH) (9) | Additional
description/location of embryonic expression of GPC-3 (15) | SGBS abnormalities
(15,16,18) |
|---|
| Digestive
tract | Positive
expression | Positive
expression | Human: Smooth
muscle and gastric parietal cells; Mice: Epithelium only at 8.5
days, submucosal layer | Diastasis recti,
omphalocele and hernias |
| Gonads | Positive in male
embryos | Positive
expression | Human: Leydig
cells, seminiferous epithelium, urethra, deferent duct | Cryptorchidism,
reduced penile length, risk for testicular gonadoblastoma |
| Kidney | Positive
expression | Positive
expression | Human: Cortex
convoluted tubules and Bowman's capsule; Mice: Mesenchymal | Renal dysplasia,
nephromegaly, risk for Wilms tumors |
| Limb buds | Positive
expression | Positive
expression | Human: Mesenchymal
in P1; Mice: Mesenchymal from 9.5 to 11.5 days post-coitum. After,
only cartilage precursor of limb bones | Polydactyly,
syndactyly |
| Liver | Positive
expression | Positive
expression | – | Hepatomegaly |
| Nervous system | Positive
expression | Negative
expression | Human: Only spinal
cord motoneurons and dorsal root ganglia neurons | Hypotony,
developmental delay, CNS malformations, and high risk for
neuroblastoma and medulloblastoma |
| Oral cavity | Positive
expression | Positive
expression | Human: Palate;
Mice: Epithelium of oral cavity | Cleft palate |
| Pancreas | Positive/Negative
expression | Not reported | Human: Positive
exocrine in P1 and P2, negative in endocrine | Hyperplastic islets
of Langerhans, hypoglycemia |
| Respiratory
system | Positive
expression | Positive
expression | Human: Only
mesenchymal cells of the lungs in P1 and P2; Mice: Cartilage of
trachea, larynx and major bronchi, mesenchymal of lungs | Pneumonia |
| Skin | Negative
expression | Positive
expression | Mouse: Dermis and
hair follicles | None reported |
| Tongue | Positive
expression | Positive
expression | Human: Striated
muscle, epithelium, and connective tissue of tongue; Mouse: Only
connective tissue | Macroglossia and
midline groove |
| Vertebrae | Positive
expression | Positive
expression | Human: Mesenchymal
between caudal vertebrae; Mice: Mesenchymal cells, cartilage and
intervertebral discs | Vertebral
abnormalities |
Organs or tissues that displayed positive GPC-3
expression in mouse embryonic tissues include: digestive tract,
gonads, kidney, limb buds, liver, oral cavity, respiratory system,
skin, tongue, and vertebrae (9,15). In
contrast, ISH revealed negative expression of GPC-3 in mouse nerve
tissue. Pancreas expression in mice was not reported in this study.
While GPC-3 expression across mouse and human embryonic tissue
appears consistent in most organs, there are some tissues that
differ. While nerve tissue shows positive expression in humans, in
mice it is not expressed. On the other hand, human tissue is
negative for expression of GPC-3 in skin, but this same tissue is
positive for the protein in mice (9). Additional detail regarding location
and description of GPC-3 expression in mice embryonic tissue can be
seen in Table I.
Studies have also shown decreased GPC-3 expression
in the SGB overgrowth syndrome, causing a spectrum of congenital
defects such as macrosomia, congenital heart disease, conduction
defects, supernumerary nipples, diastasis recti/umbilical hernia,
diaphragmatic hernia, renal dysplasia/nephromegaly,
cryptorchidism/hypospadias, and hand anomalies (brachydactyly,
cutaneous syndactyly, polydactyly) (16). This change in expression and
resulting defects are due to a mutation that causes sequence
variants as well as multiexon/whole gene deletion in 37–70% of male
cases (16,17). Table
I gives a summary of the SGB syndrome abnormalities found in
various organs and tissues as a result of the GPC-3 loss of
function mutation (15,16,18).
In these cases, GPC-3 acts as a regulator for normal embryonic
development and is only found to cause problems in embryonic tissue
when it is mutated (Table I).
Expression of GPC-3 in normal adult
tissue
Table II summarizes
GPC-3 expression in adult tissues normal versus tumor, while
Table I summarizes expression of
GPC-3 in embryonic tissue in human and mice. As is shown in
Table II, the current literature
has some contradictions which must be further evaluated to better
understand the expression of GPC-3 in tissue types. The tissues
represented as having two or more sources contradicting one another
in normal tissue (expressed/not expressed) are: gastric glands,
kidney tubules, testicular germ cells, breast tissue, gall bladder,
and ovary (19–27). In addition, there are also sources
reporting no expression, trace amounts, or scarce expression in
normal tissue. These tissues are: liver, brain, colon, esophagus,
fat tissue, heart, lung, lymph, mouth and associated glandular
tissues, pancreas, prostate, skeletal and smooth muscle, small
intestine, thyroid and parathyroid, thymus, bladder, and uterus
(2,19,21,22,28–33).
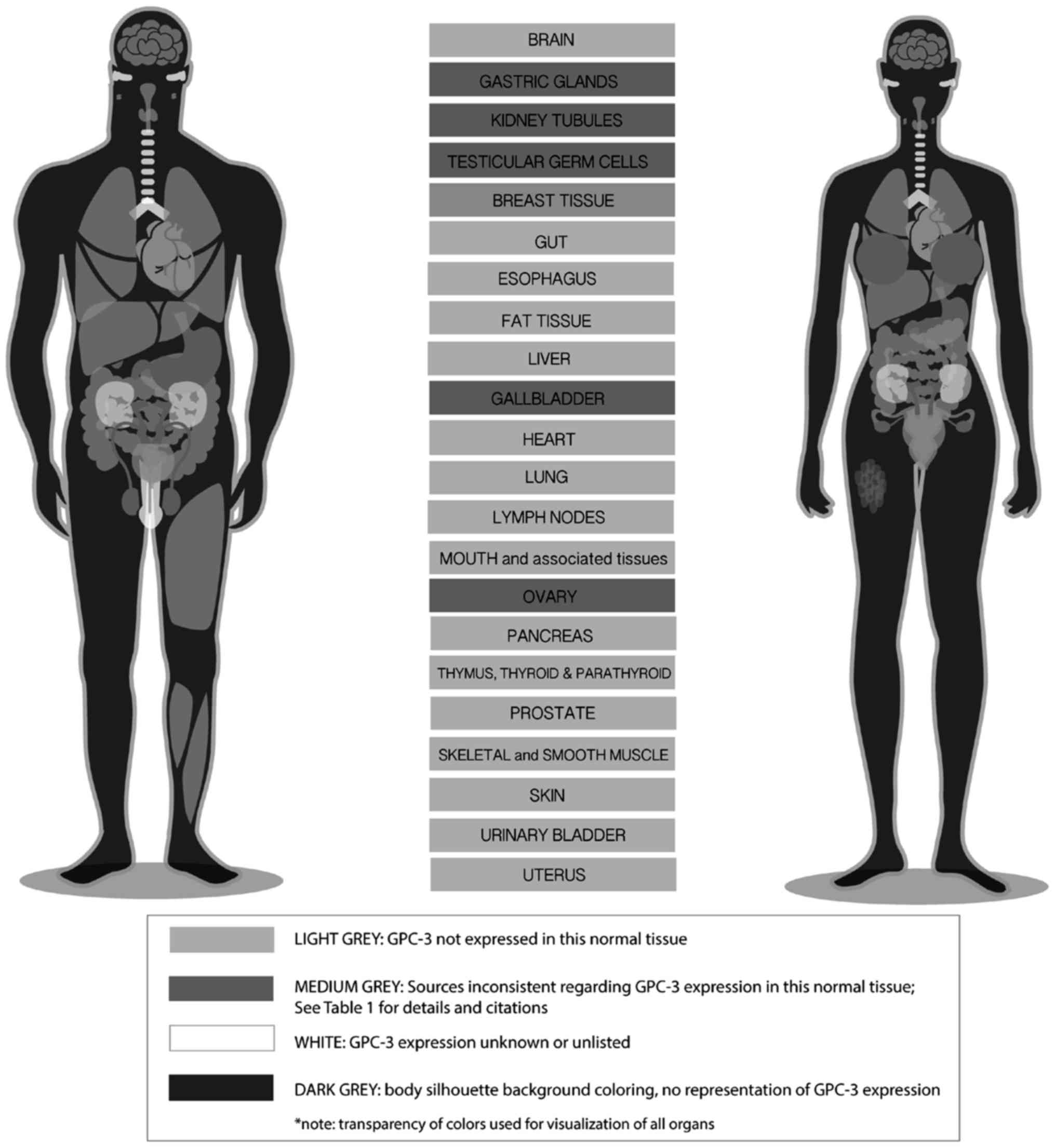
In Fig. 1, expression of GPC-3 in
normal tissue was constructed based on Table II. Different grey shades were used
to differentiate GPC-3 expression in various tissues.
| Table II.GPC-3 expression in adult human
tissue types. |
Table II.
GPC-3 expression in adult human
tissue types.
| Tissue | Normal tissue | Cancer tissue | References |
|---|
| Liver | Not expressed
(28–30) | Expressed (1,3–6) | 1,3–6,28,29,30 |
| Gastric glands | Expressed, small
subset of gastric tissues normally express GPC-3 (19); Not expressed (20) | Expressed in
gastric carcinoma (34) | 19,20,34 |
| Kidney tubules | Expressed (19); Not expressed (20); Low expression (21,22) | Reduced expression
in renal cell carcinoma (41 mentions renal cell carcinomas exposed
to GPC-3 caused reduced proliferation); Infrequent expression in
clear cell carcinoma, oncocytoma, and papillary carcinoma of the
kidney (19) | 19–22,41 |
| Testicular germ
cells | Expressed (19); Not expressed (23) | Expressed (23) | 19,23 |
| Anus | Unknown | Expressed (33); Infrequent expression in SCC
(19) | 19,33 |
| Breast tissue | Not expressed
(19); Expressed (24); Expressed (25) | Reduced expression
(24) Silenced (25); Infrequent expression in invasive
ductal and tubular carcinoma (19) | 19,24,25 |
| Brain | Not expressed
(19) | Expressed in some
non-CNS tumors and restricted expression in atypical teratoid
rhabdoid tumors and in some craniopharyngiomas (36) | 19,36 |
| Colon | Not expressed
(2,19); Trace amounts expressed (21,22) | Expressed (1,37);
Infrequent expression in adenocarcinoma and adenoma with high-grade
dysplasia (19) |
1,2,19,21,22,37 |
| Esophagus | Not expressed
(31,19) | Expressed (31,33);
Infrequent expression in squamous cell carcinoma (19) | 19,31,33 |
| Fat tissue | Not expressed
(19) | Unknown | 19 |
| Gallbladder | Not expressed
(19); Expressed (26) | Reduced expression
in gallbladder cancer (26);
Infrequent expression in adenocarcinoma (19) | 19,26 |
| Heart | Not expressed
(19); Low expression (21,22) | Unknown | 19,21,22 |
| Lung | Not expressed
(19); Low expression (21,22) | Silenced in
mesothelioma and lung adenocarcinoma (10,11,38);
Expressed in lung squamous cell carcinoma (39); Expressed (33); Infrequent expression in
adenocarcinoma and large cell carcinoma (19) |
10,11,19,21,22,33,38,39 |
| Lymph nodes | Not expressed
(19) | Unknown | 19 |
| Mouth (mucosa) | Not expressed
(19) | Expressed on some
tongue, base/tonsil, ventral tongue/floor of mouth (33); Infrequent expression in squamous
cell carcinoma (19) | 19,33 |
| Ovary | Not expressed
(19); Low expression (21,22);
Expressed (27) | Silenced in
epithelial ovarian cancer (10,11,25,38);
Infrequent expression in Brenner tumor, endometrioid carcinoma,
mucinous carcinoma, serous carcinoma, and yolk sac tumor (19); No expression (27) |
10,11,19,21,22,25,27,38 |
| Pancreas | Not expressed
(19); Trace amounts expressed
(21,22) | Expressed (31); No expression in pancreatic
adenocarcinoma (40) | 19,21,22,31,40 |
| Parathyroid | Not expressed
(19) | Unknown | 19 |
| Parotid gland | Not expressed
(19) | Unknown | 19 |
| Penis | Unknown | No expression in
squamous cell carcinoma (33);
Infrequent expression in SCC (19) | 19,33 |
| Prostate | Not expressed
(19) in adenocarcinoma (19); No expression (42) | Infrequent
expression | 19,42 |
| Skeletal
muscle | Not expressed
(19); Trace amounts expressed
(21,22) | Infrequent
expression in rhabdomyosarcoma (19) | 19,21,22 |
| Skin | Not expressed
(19) | No expression in
squamous cell carcinoma (33);
Expression in melanoma (28) | 19,33 |
| Small intestine
(mucosa) | Not expressed
(19); Trace amounts expressed
(21,22) | Infrequent
expression in adenocarcinoma and gastrointestinal stromal tumor
(19) | 19,21,22 |
| Smooth muscle | Not expressed
(19) | Infrequent
expression in leiomyosarcoma (19) | 19 |
| Submandibular
gland | Not expressed
(19) | Infrequent
expression in acinar cell carcinoma, adenoid cystic carcinoma, and
small cell carcinoma (19) | 19 |
| Thymus | Not expressed
(19) | Unknown | 19 |
| Thyroid | Not expressed
(19); Scarcely expressed (32) | Infrequent
expression in papillary carcinoma (19); Expressed in early papillary
carcinoma (32) | 19,32 |
| Urinary
bladder | Not expressed
(19,33) | Expressed (33); Expressed in urothelial carcinoma,
mostly in high grade tumors (35);
Infrequent expression in noninvasive transitional cell carcinoma,
small cell carcinoma, and squamous cell carcinoma (19) | 19,33,35 |
| Uterus (cervix,
endometrium, myometrium) | Not expressed
(19) | Expressed (33); Infrequent expression in cervix as
SCC and endometrium as endometrioid and serous carcinoma (19) | 19,33 |
| Vagina | Unknown | Expressed (33) | 33 |
| Vulva | Unknown | No expression in
squamous cell carcinoma (33);
Expression in some tumors analyzed in SCC (19) | 19,33 |
Expression of GPC-3 in cancer
tissue
GPC-3 expression in various types of cancer is not
fully understood, though the variance may be due to the specific
type of cancer being studied. Table
II lists the reported expression of GPC-3 in cancer in various
tissue types. Expression of GPC-3 was found in the following
cancers: liver, gastric carcinoma, melanoma, high-grade urothelial
carcinoma, testicular, and some uterine and vaginal cancers
(1,3–6,23,28,33–35).
GPC-3 expression was also reported in some non-CNS tumors of the
brain (36). In various other
tissue types such as colon, lung, anal, esophageal, mouth mucosa,
pancreas, skin, thyroid, bladder, uterus, and vulvar tissue GPC-3
expression varied based on the type, and origin, and grade of
cancer [i.e. squamous cell carcinoma (SCC) versus mesothelioma of
the lung] (1,10,11,19,28,31–33,37–40).
In some cancerous tissues, reduced or even silenced
expression of GPC-3 was found; namely renal cell carcinomas, as
well as cancer of the breast, ovaries, penis, prostate, and
gallbladder (10,11,24–26,33,38,41,42).
Similarly, infrequent expression of GPC-3 was reported in some
cancers of the kidney, invasive ductal and tubular carcinomas of
the breast, some brain tumors, adenocarcinomas of the gall bladder,
prostate, and small intestine, squamous cell carcinoma of the
penis, rhabdosarcomas and leimyosarcomas of the skeletal and smooth
muscle, adeno- and small cell carcinomas of the submandibular
glands, among other cancerous tissues depending upon grade and
location within the tissue (19,33,36).
For more detail regarding GPC-3 expression in these tissues, refer
to Table II.
In human breast cancer, GPC-3 has been shown to be
silenced due to hypermethylation of its promoter region. The data
also suggest that GPC-3 can act as a negative regulator of breast
cancer growth as well a downregulator in mesotheliomas and ovarian
cancer (25). Xq26 is a region
frequently deleted in advanced ovarian cancers and is where the
GPC-3 gene is located.
In ovarian cancer, GPC-3 expression was lost in cell
lines analyzed which was also due to hypermethylation of the
promoter region. Expression of GPC-3 was restored with the
treatment of 5-aza-2′-deoxycytidinem, a demethylating agent.
Ectopic GPC-3 expression inhibited the growth of ovarian cancer
cell lines and results show that GPC-3 is frequently inactivated in
a subset of ovarian cancers which may suggest that it functions as
a tumor suppressor in the ovary (27).
In 15% or more of studied cases, GPC-3 was expressed
in tumor types of hepatocellular carcinoma, squamous cell carcinoma
of the lung, liposarcoma, testicular nonseminomatous germ cell
tumor, cervical intraepithelial neoplasia (grade 3), malignant
melanoma, adenoma of the adrenal gland, schwannoma, malignant
fibrous histiocytoma, adenocarcinoma of the stomach (intestinal
subtype), chromophobe renal cell carcinoma, invasive lobular
carcinoma of the breast, medullary carcinoma of the breast,
squamous cell carcinoma of the larynx, small cell carcinoma of the
lung, invasive transitional cell carcinoma of the urinary bladder,
mucinous carcinoma of the breast, and squamous cell carcinoma of
the cervix (19).
GPC-3 as a serum marker for early detection
of HCC
GPC-3 may be a potential serum marker in diseases
such as HCC in which expression of GPC-3 is markedly increased
(20,21). Capurro et al examined GPC-3
protein expression and serum levels using immunohistochemistry and
ELISA in HCC patients, as well as serum levels in healthy donors
and patients with hepatitis and liver cirrhosis. Results revealed
increased GPC-3 expression levels in patients with HCC, but not in
healthy hepatocytes. GPC-3 serum levels were significantly elevated
in HCC patients, but undetectable in the serum of healthy and
hepatitis-infected patients (22).
Discussing where this protein is cleaved, the
cleaved portion of the NH(2)-terminal was found to be between
Arg(358) and Ser(359) of GPC-3. In addition to this, it was
observed that soluble GPC-3 can be specifically detected in the
sera of patients with HCC using N-mAbs (23). Unfortunately, the cleavage events
and other post-translational modifications are understudied. It is
necessary to investigate in terms of function and cellular
localization the cleavage events on GPC-3. Focusing on serum marker
it is necessary to understand if the protein is cleaved and which
portion is secreted improving the knowledge of secretory pathways
involved in its sorting in and out of the cells, with major
attention on exosomal cargos.
GPC-3 was also identified as a novel diagnostic
marker for human melanoma it its early stages. In patients with
melanoma, GPC was detected. However, the protein was not found in
the sera of healthy patients, or those with benign skin lesions
(large congenital melanocytic nevus). It was also detected in the
serum of patients with stage 0, in situ melanoma (28).
In another study carried out to observe biomarkers
for HCC, preoperative serum GPC-3-N (sGPC-3N) levels were measured
alongside with serum AFP in patients with HCC. It was observed that
high serum AFP corresponded with a high sGPC-3N level. The study
states that sGPC-3N may serve as an independent prognostic
biomarker in HCC patients (24).
Serum α-fetoprotein is the common diagnostic marker
to detect HCC. Several studies showed that the combination of
α-fetoprotein and GPC-3 increase significantly the sensitivity of
diagnostic value compared to their sensitivity analyzed
independently (43). Several
markers are under investigation to ameliorate the sensitivity and
specificity of HCC detection such as: des-γ carboxyprothrombin,
lens culinaris agglutinin-reactive α-fetoprotein, Golgi protein 73
(43,44). However, GPC-3 seems to be more
specific due to its peculiar expression in hepatocytes as showed
before, and this represents the main advantage for its use in HCC
detection even if its mechanism of transport in the serum is still
unknown. For its peculiar localization and post-translational
modifications GPC-3 could represent an ideal target for antibody
therapeutic approach with the possibility to bind its membrane form
thus inhibiting its activity. Moreover, a chemical approach could
obtain the cleavage events that determine GPC-3 functional
modulation and cellular localization. Consequent to its expression
in pre-neoplastic hepatocytes, this offers the unique advantage of
allowing an early and specific therapeutic approach.
GPC-3 in resection, regeneration and liver
diseases
Few studies have demonstrated the expression of
GPC-3 pre- and post-hepatic resection. Due to hepatic progenitor
cells expressing cytokeratin 19 (CK19) and GPC-3 at varying
phenotypes, cell samples were analyzed from patients who had a
liver resection and were statistically compared with each phenotype
of expression in Table III. It
was found that CK19+/GPC-3+ HCC was the most
aggressive subtype, followed by the
CK19−/GPC-3+ HCC and finally with the
CK19−/GPC-3− HCC subtype being the least
aggressive from the subtypes observed (45). Therefore, a poorer prognosis may be
associated with patients undergoing a hepatectomy who have the
expression of both CK19 and GPC-3.
| Table III.Phenotype of expression (25). |
Table III.
Phenotype of expression (25).
| Order of
severity | Cell marker
phenotype CK19/GPC-3 HCC | Cell marker
description |
|---|
| 1 | + + | HCC subtype
transformed from hepatic progenitor cells (HPC) |
| 2 | − + | Immature
hepatocyte |
| 3 | − - | Terminal
differentiated hepatocyte |
Liu et al, on the other hand, described
transgenic GPC-3 mice, which were under the control of the albumin
promoter gene, overexpressed GPC-3 and actually had a suppression
of hepatocyte proliferation and liver regeneration following
partial hepatectomy (30,46). These mice developed normally and the
authors claim that GPC-3 may play a negative regulatory role in
hepatocyte proliferation, showing a contrasting role of GPC-3 in
the hepatocytes. Further investigation using models post-PH is
needed to uncover various important aspects of GPC-3 expression
such as the density and location of GPC-3 expression (i.e. the cut
margin or other location). This research may be instrumental in
further elucidation of the role and mechanisms of GPC-3 in liver
growth and regeneration post-resection and even injury
(mechanically, inflammatory, or otherwise). However, limited
studies have been conducted on the regeneration of the liver.
At this time there is no evidence on a possible
function and presence of GPC-3 in other liver diseases such as
fibrosis, fatty liver disease or liver cirrhosis. The peculiarity
of its expression in neoplastic hepatocytes makes GPC-3 not only a
unique HCC marker but also a possible therapeutic target. However,
studies in cirrhotic livers without HCC are necessary to rule out
the role of GPC-3 or the presence in these circumstances.
GPC-3 as a therapeutic target for HCC
Therapeutic targets are especially important in the
treatment of various types of cancer. GC33, which is a humanized
monoclonal antibody, was shown to bind human GPC-3. GC33 has been
observed to contain antitumor properties and targets GPC-3
specifically. It was noted that GC33 was well tolerated in advanced
HCC and provided some benefit in the clinical treatment (47). In addition, human (MDX-1414 and HN3)
and humanized mouse (GC33 and YP7) antibodies that also target
GPC-3 are under different stages of clinical development and could
also aid in the treatment of HCC (21). miR-219-5p is a microRNA (miRNA)
which could exert tumor suppression on hepatic cells expressing
GPC-3. miR-219-5p reduced both the mRNA and protein levels of GPC-3
and exerted tumor-suppressive effects in HCC (48). Finally, nanoparticles are also under
development to target and bind GPC-3 in cells that could provide
useful for further imaging and targeting of GPC-3 (49).
Research has also been conducted in regards to
peptide sequences which bind GPC-3. Feng et al described a
12-mer peptide sequence (DHLASLWWGTEL) which was able to act as a
probe and bind GPC-3 for HCC detection (21). The peptide allows visualization of
the specific sequence via near-infrared fluorescence (50). Another therapeutic target on GPC-3
is mir717, a miRNA, which is located on intron 3 of GPC-3 on the X
chromosome. It has been shown to play a regulatory role in renal
osmoregulation. mir717 might be connected in some way with obesity
regulation, one of the risk factors of HCC (51,52).
It has been proposed that GPC-3 acts as a negative
regulator in the Hedgehog signaling pathway during development and
as a result, a non-functional GPC-3 protein could be the cause of
overgrowth or a GPC3 modified by post-translational modifications.
GPC-3 has also been associated with the binding of Wnt to its
signaling receptor, Frizzled (53–56).
Both the Hh and canonical/non-canonical pathways of Wnt could be
potential therapeutic targets as GPC-3 may play a role in
proliferation and/or growth suppression, depending on the type of
tissue and the stage of development (8,57). In
addition, the peptide sequences
Arg-Leu-Asn-Val-Gly-Gly-Thr-Tyr-Phe-Leu-Thr-Thr-Arg-Gln and
Tyr-Phe-Leu,Thr-Thr-Arg-Gln showed selective binding of GPC-3
(58) which could provide useful
for a target that is needing to bind onto GPC-3 for regulation.
GPC-3 and co-receptors
GPC-3 has been observed to interact with a number of
co-receptors that further modulate cellular expression. Liu et
al showed that GPC-3 and CD81 levels were significantly
upregulated in general in the transgenic mice following partial
hepatectomy (30,46). They showed that the negative
regulatory role was somehow associated with GPC-3 and CD81 and that
there is enhanced association between GPC-3 and CD81. GPC-3 and
CD81 have both been associated as binding partners in which
expression could influence the Hh (13). Co-localization of GPC-3 and CD81 was
also shown to occur 2–6 days after hepatectomy (30). In contrast, another study observed
that GPC-3 binds to members of the Hh pathway and prevents their
interactions with the patched-1 receptor. There was decreased
binding of GPC-3 with Hh and CD81 following PH and GPC-3/CD81 may
play a role in the termination of liver growth following liver
regeneration (14,46). Dysregulation of the association
between GPC-3/CD81, which may occur during hepatitis C infection
can result in dysregulated signaling and proliferation in infected
hepatocytes (59). The structure
and character of GPC-3 as a transmembrane protein, in conjunction
with the current literature, lends itself to the strong possibility
of many associations or interactions with other co-receptors and
proteins, not only in hepatocytes, but possible in other normal or
cancerous tissue types. It is important to emphasize again the
paucity in the current literature in regard to this, and many other
aspects of GPC-3.
Conclusion
GPC-3 is a molecule that is still not fully
understood in its role in the proliferation and suppression of cell
growth in normal and abnormal or cancerous tissue and also in
structural and post-translational modifications. The question still
remains as to why it is normally expressed in some tissues, while
remaining silenced in others. Future research that looks into the
upstream and downstream cell signaling pathways and how GPC-3 may
be involved could provide further answers. In addition, studies
investigating a more complete picture and analysis of GPC-3
structure are warranted, as glycosylation, sulfonation, or other
structural components of the protein may be important in
understanding its regulatory function within differing tissue
types. In this same vein, continued insight into the cleavage of
GPC-3 and the function of the cleaved versus non-cleaved form of
the molecule could lead to a more complete comprehension of
function. Other research paths may include developing the theory of
GPC-3 as a dual tumor suppressor and oncogene, as dependent upon
its structure. With exosome research brightening the horizons, more
attention and detail must be paid to uncover the importance of
exosomal GPC-3 as a serum marker or as possible therapeutic target
in HCC. GPC-3 is a molecule that could further connect the missing
links in liver cancer research and lead to an abundance of new
intercellular relationships to reveal important aspects of the
biology of this disease.
Glossary
Abbreviations
Abbreviations:
|
CK19
|
cytokeratin 19
|
|
FGF2
|
fibroblast growth factor 2
|
|
GPC-3
|
glypican-3
|
|
GPI
|
glycosylphosphatidylinositol
|
|
HCC
|
hepatocellular carcinoma
|
|
HPC
|
hepatic progenitor cell
|
|
HSC
|
heparin sulfate chain
|
|
HSPG
|
heparan sulfate proteoglycan
|
|
Hh
|
hedgehog
|
|
IHC
|
immunohistochemistry
|
|
ISH
|
in situ hybridization
|
|
PH
|
partial hepatectomy
|
|
SGB
|
Simpson-Golabi-Behmel
|
|
SULF2
|
sulfatase 2
|
|
sGPC-3N
|
serum glypican-3 N-terminal
|
|
miRNA
|
microRNA
|
|
SCC
|
squamous cell carcinoma
|
References
|
1
|
Midorikawa Y, Ishikawa S, Iwanari H,
Imamura T, Sakamoto H, Miyazono K, Kodama T, Makuuchi M and
Aburatani H: Glypican-3, overexpressed in hepatocellular carcinoma,
modulates FGF2 and BMP-7 signaling. Int J Cancer. 103:455–465.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Filmus J and Selleck SB: Glypicans:
Proteoglycans with a surprise. J Clin Invest. 108:497–501. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakatsura T, Yoshitake Y, Senju S, Monji
M, Komori H, Motomura Y, Hosaka S, Beppu T, Ishiko T, Kamohara H,
et al: Glypican-3, overexpressed specifically in human
hepatocellular carcinoma, is a novel tumor marker. Biochem Biophys
Res Commun. 306:16–25. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang XY, Degos F, Dubois S, Tessiore S,
Allegretta M, Guttmann RD, Jothy S, Belghiti J, Bedossa P and
Paradis V: Glypican-3 expression in hepatocellular tumors:
Diagnostic value for preneoplastic lesions and hepatocellular
carcinomas. Hum Pathol. 37:1435–1441. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Montalbano M, Rastellini C, Wang X,
Corsello T, Eltorky MA, Vento R and Cicalese L: Transformation of
primary human hepatocytes in hepatocellular carcinoma. Int J Oncol.
48:1205–1217. 2016.PubMed/NCBI
|
|
6
|
Montalbano M, Curcurù G, Shirafkan A,
Vento R, Rastellini C and Cicalese L: Modeling of hepatocytes
proliferation isolated from proximal and distal zones from human
hepatocellular carcinoma lesion. PLoS One. 11:e01536132016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sung YK, Hwang SY, Park MK, Farooq M, Han
IS, Bae HI, Kim JC and Kim M: Glypican-3 is overexpressed in human
hepatocellular carcinoma. Cancer Sci. 94:259–262. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stigliano I, Puricelli L, Filmus J,
Sogayar MC, Bal de Kier Joffé E and Peters MG: Glypican-3 regulates
migration, adhesion and actin cytoskeleton organization in mammary
tumor cells through Wnt signaling modulation. Breast Cancer Res
Treat. 114:251–262. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pellegrini M, Pilia G, Pantano S, Lucchini
F, Uda M, Fumi M, Cao A, Schlessinger D and Forabosco A: Gpc3
expression correlates with the phenotype of the
Simpson-Golabi-Behmel syndrome. Dev Dyn. 213:431–439. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim H, Xu GL, Borczuk AC, Busch S, Filmus
J, Capurro M, Brody JS, Lange J, D'Armiento JM, Rothman PB, et al:
The heparan sulfate proteoglycan GPC3 is a potential lung tumor
suppressor. Am J Respir Cell Mol Biol. 29:694–701. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ho M and Kim H: Glypican-3: A new target
for cancer immunotherapy. Eur J Cancer. 47:333–338. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lai JP, Sandhu DS, Yu C, Han T, Moser CD,
Jackson KK, Guerrero RB, Aderca I, Isomoto H, Garrity-Park MM, et
al: Sulfatase 2 up-regulates glypican 3, promotes fibroblast growth
factor signaling, and decreases survival in hepatocellular
carcinoma. Hepatology. 47:1211–1222. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhave VS, Mars W, Donthamsetty S, Zhang X,
Tan L, Luo J, Bowen WC and Michalopoulos GK: Regulation of liver
growth by glypican 3, CD81, hedgehog, and Hhex. Am J Pathol.
183:153–159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Michalopoulos GK: Regenerative responses
to liver injury. Eur J Med Res. 19 Suppl 1:S12014.doi:
10.1186/2047-783X-19-S1-S1. View Article : Google Scholar :
|
|
15
|
Iglesias BV, Centeno G, Pascuccelli H,
Ward F, Peters MG, Filmus J, Puricelli L and de Kier Joffé EB:
Expression pattern of glypican-3 (GPC3) during human embryonic and
fetal development. Histol Histopathol. 23:1333–1340.
2008.PubMed/NCBI
|
|
16
|
Golabi M, Leung A and Lopez C:
Simpson-Golabi-Behmel syndrome type 1.
GeneReviews®[Internet]. Pagon RA, Adam MP, Ardinger HH,
et al: University of Washington; Seattle, Seattle, WA: pp.
1993–2016. 2006 June 23–2011
|
|
17
|
Khan S, Blackburn M, Mao DL, Huber R,
Schlessinger D and Fant M: Glypican-3 (GPC3) expression in human
placenta: Localization to the differentiated syncytiotrophoblast.
Histol Histopathol. 16:71–78. 2001.PubMed/NCBI
|
|
18
|
DeBaun MR, Ess J and Saunders S: Simpson
Golabi Behmel syndrome: Progress toward understanding the molecular
basis for overgrowth, malformation, and cancer predisposition. Mol
Genet Metab. 72:279–286. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baumhoer D, Tornillo L, Stadlmann S,
Roncalli M, Diamantis EK and Terracciano LM: Glypican 3 expression
in human nonneoplastic, preneoplastic, and neoplastic tissues: A
tissue microarray analysis of 4,387 tissue samples. Am J Clin
Pathol. 129:899–906. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao H, Li K, Tu H, Pan X, Jiang H, Shi B,
Kong J, Wang H, Yang S, Gu J, et al: Development of T cells
redirected to glypican-3 for the treatment of hepatocellular
carcinoma. Clin Cancer Res. 20:6418–6428. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng M and Ho M: Glypican-3 antibodies: A
new therapeutic target for liver cancer. FEBS Lett. 588:377–382.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hsu HC, Cheng W and Lai PL: Cloning and
expression of a developmentally regulated transcript MXR7 in
hepatocellular carcinoma: Biological significance and
temporospatial distribution. Cancer Res. 57:5179–5184.
1997.PubMed/NCBI
|
|
23
|
Zynger DL, Dimov ND, Luan C, Teh BT and
Yang XJ: Glypican 3: A novel marker in testicular germ cell tumors.
Am J Surg Pathol. 30:1570–1575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Castillo L, Huvelle MAL, Fujita A, Lobba
ARM, Tascon R and Peters MG: Expression of glypican-3 (GPC3) in
malignant and non-malignant human breast tissues. Open Cancer J.
8:12–23. 2015.https://benthamopen.com/contents/pdf/TOCJ/TOCJ-8-12.pdf
View Article : Google Scholar
|
|
25
|
Xiang YY, Ladeda V and Filmus J:
Glypican-3 expression is silenced in human breast cancer. Oncogene.
20:7408–7412. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Man XB, Tang L, Zhang BH, Li SJ, Qiu XH,
Wu MC and Wang HY: Upregulation of Glypican-3 expression in
hepatocellular carcinoma but downregulation in cholangiocarcinoma
indicates its differential diagnosis value in primary liver
cancers. Liver Int. 25:962–966. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin H, Huber R, Schlessinger D and Morin
PJ: Frequent silencing of the GPC3 gene in ovarian cancer cell
lines. Cancer Res. 59:807–810. 1999.PubMed/NCBI
|
|
28
|
Nakatsura T, Kageshita T, Ito S, Wakamatsu
K, Monji M, Ikuta Y, Senju S, Ono T and Nishimura Y: Identification
of glypican-3 as a novel tumor marker for melanoma. Clin Cancer
Res. 10:6612–6621. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luo JH, Ren B, Keryanov S, Tseng GC, Rao
UN, Monga SP, Strom S, Demetris AJ, Nalesnik M, Yu YP, et al:
Transcriptomic and genomic analysis of human hepatocellular
carcinomas and hepatoblastomas. Hepatology. 44:1012–1024. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu B, Paranjpe S, Bowen WC, Bell AW, Luo
JH, Yu YP, Mars WM and Michalopoulos GK: Investigation of the role
of glypican 3 in liver regeneration and hepatocyte proliferation.
Am J Pathol. 175:717–724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ahmed SS, Ola A, Ahmed E and Mohammed A:
Seroprevalence of Glypican-3 (GPC-3) in patients with pancreatic,
gastric and esophageal cancers. NY Sci J. 4:45–50. 2011.http://www.sciencepub.net/newyork/ny0407/08_6074ny0407_45_50.pdf
|
|
32
|
Yamanaka K, Ito Y, Okuyama N, Noda K,
Matsumoto H, Yoshida H, Miyauchi A, Capurro M, Filmus J and Miyoshi
E: Immunohistochemical study of glypican 3 in thyroid cancer.
Oncology. 73:389–394. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gailey MP and Bellizzi AM:
Immunohistochemistry for the novel markers glypican 3, PAX8, and
p40 (∆Np63) in squamous cell and urothelial carcinoma. Am J Clin
Pathol. 140:872–880. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ushiku T, Uozaki H, Shinozaki A, Ota S,
Matsuzaka K, Nomura S, Kaminishi M, Aburatani H, Kodama T and
Fukayama M: Glypican 3-expressing gastric carcinoma: Distinct
subgroup unifying hepatoid, clear-cell, and
alpha-fetoprotein-producing gastric carcinomas. Cancer Sci.
100:626–632. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aydin O, Yildiz L, Baris S, Dundar C and
Karagoz F: Expression of Glypican 3 in low and high grade
urothelial carcinomas. Diagn Pathol. 10:342015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chan ES, Pawel BR, Corao DA, Venneti S,
Russo P, Santi M and Sullivan LM: Immunohistochemical expression of
glypican-3 in pediatric tumors: An analysis of 414 cases. Pediatr
Dev Pathol. 16:272–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gonzalez AD, Kaya M, Shi W, Song H, Testa
JR, Penn LZ and Filmus J: OCI-5/GPC3, a glypican encoded by a gene
that is mutated in the Simpson-Golabi-Behmel overgrowth syndrome,
induces apoptosis in a cell line-specific manner. J Cell Biol.
141:1407–1414. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Murthy SS, Shen T, De Rienzo A, Lee WC,
Ferriola PC, Jhanwar SC, Mossman BT, Filmus J and Testa JR:
Expression of GPC3, an X-linked recessive overgrowth gene, is
silenced in malignant mesothelioma. Oncogene. 19:410–416. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu X, Li Y, Chen SW, Shi Y and Xu F:
Differential expression of glypican-3 (GPC3) in lung squamous cell
carcinoma and lung adenocarcinoma and its clinical significance.
Genet Mol Res. 14:10185–10192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mounajjed T, Zhang L and Wu TT: Glypican-3
expression in gastrointestinal and pancreatic epithelial neoplasms.
Hum Pathol. 44:542–550. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Valsechi MC, Oliveira AB, Conceição AL,
Stuqui B, Candido NM, Provazzi PJ, de A, raújo LF, Silva WA Jr,
Calmon Mde F and Rahal P: GPC3 reduces cell proliferation in renal
carcinoma cell lines. BMC Cancer. 14:6312014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang L, Liu H, Sun L, Li N, Ding H and
Zheng J: Glypican-3 as a potential differential diagnosis marker
for hepatocellular carcinoma: A tissue microarray-based study. Acta
Histochem. 114:547–552. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang TS, Shyu YC, Turner R, Chen HY and
Chen PJ: Diagnostic performance of alpha-fetoprotein, lens
culinaris agglutinin-reactive alpha-fetoprotein, des-gamma
carboxyprothrombin, and glypican-3 for the detection of
hepatocellular carcinoma: A systematic review and meta-analysis
protocol. Syst Rev. 2:372013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang Y, Yang H, Xu H, Lu X, Sang X, Zhong
S, Huang J and Mao Y: Golgi protein 73, not Glypican-3, may be a
tumor marker complementary to α-Fetoprotein for hepatocellular
carcinoma diagnosis. J Gastroenterol Hepatol. 29:597–602. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feng J, Zhu R, Chang C, Yu L, Cao F, Zhu
G, Chen F, Xia H, Lv F, Zhang S, et al: CK19 and Glypican 3
expression profiling in the prognostic indication for patients with
HCC after surgical resection. PLoS One. 11:e01515012016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu B, Bell AW, Paranjpe S, Bowen WC,
Khillan JS, Luo JH, Mars WM and Michalopoulos GK: Suppression of
liver regeneration and hepatocyte proliferation in
hepatocyte-targeted glypican 3 transgenic mice. Hepatology.
52:1060–1067. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu AX, Gold PJ, El-Khoueiry AB, Abrams
TA, Morikawa H, Ohishi N, Ohtomo T and Philip PA: First-in-man
phase I study of GC33, a novel recombinant humanized antibody
against glypican-3, in patients with advanced hepatocellular
carcinoma. Clin Cancer Res. 19:920–928. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang N, Lin J, Ruan J, Su N, Qing R, Liu
F, He B, Lv C, Zheng D and Luo R: MiR-219-5p inhibits
hepatocellular carcinoma cell proliferation by targeting
glypican-3. FEBS Lett. 586:884–891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park JO, Stephen Z, Sun C, Veiseh O,
Kievit FM, Fang C, Leung M, Mok H and Zhang M: Glypican-3 targeting
of liver cancer cells using multifunctional nanoparticles. Mol
Imaging. 10:69–77. 2011.PubMed/NCBI
|
|
50
|
Zhu D, Qin Y, Wang J, Zhang L, Zou S, Zhu
X and Zhu L: Novel glypican-3-binding peptide for in vivo
hepatocellular carcinoma fluorescent imaging. Bioconjug Chem.
27:831–839. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kunej T, Skok DJ, Horvat S, Dovc P and
Jiang Z: The glypican 3-hosted murine mir717 gene: Sequence
conservation, seed region polymorphisms and putative targets. Int J
Biol Sci. 6:769–772. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kew MC: Obesity as a cause of
hepatocellular carcinoma. Ann Hepatol. 14:299–303. 2015.PubMed/NCBI
|
|
53
|
Filmus J and Capurro M: The role of
glypican-3 in the regulation of body size and cancer. Cell Cycle.
7:2787–2790. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Filmus J and Capurro M: Glypican-3: A
marker and a therapeutic target in hepatocellular carcinoma. FEBS
J. 280:2471–2476. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tsuda M, Kamimura K, Nakato H, Archer M,
Staatz W, Fox B, Humphrey M, Olson S, Futch T, Kaluza V, et al: The
cell-surface proteoglycan Dally regulates Wingless signalling in
Drosophila. Nature. 400:276–280. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
56
|
Song HH, Shi W, Xiang YY and Filmus J: The
loss of glypican-3 induces alterations in Wnt signaling. J Biol
Chem. 280:2116–2125. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Capurro MI, Xiang YY, Lobe C and Filmus J:
Glypican-3 promotes the growth of hepatocellular carcinoma by
stimulating canonical Wnt signaling. Cancer Res. 65:6245–6254.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lee YL, Ahn B-C, Lee Y, Lee S-W, Cho J-Y
and Lee J: Targeting of hepatocellular carcinoma with
glypican-3-targeting peptide ligand. J Pept Sci. 17:763–769. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yorita K, Takahashi N, Takai H, Kato A,
Suzuki M, Ishiguro T, Ohtomo T, Nagaike K, Kondo K, Chijiiwa K, et
al: Prognostic significance of circumferential cell surface
immunoreactivity of glypican-3 in hepatocellular carcinoma. Liver
Int. 31:120–131. 2011. View Article : Google Scholar : PubMed/NCBI
|