Introduction
Epithelial ovarian cancer (EOC) accounts for 90% of
all ovarian cancer cases, and is one of the three most common
gynecological malignancies (1). EOC
has a poor prognosis and is the leading cause of death from
gynecologic cancers. Although advanced surgical techniques and
newly emerging targeted therapies have improved patient outcomes,
the 5-year survival rate for EOC patients is still low, at
approximately 30–40% (2,3). In addition, the precise molecular
mechanisms underlying the biological behavior of EOC remain
unclear. Several hypotheses have been proposed to explain the
etiology of EOC, such as the incessant ovulation hypothesis
(4), the gonadotropin theory
(5) and the sex-steroid hormone
hypothesis (6). Increasing basic
and epidemiological evidence has shown that the enriched hormonal
environment in the ovary can influence the development of EOC
(7).
Gonadotropins, including luteinizing hormone (LH)
and follicle stimulating hormone (FSH), are conducive to the growth
of ovarian cancer based on the ‘gonadotropin theory’ (5) and are regulated by
gonadotropin-releasing hormone (GnRH). GnRH agonists initially
stimulate the release of gonadotropins, while sustained
administration inhibits the release by desensitizing or
downregulating the expression of the GnRH receptor (GnRHR).
Clinically, GnRH agonists have been widely used in the treatment of
hormone-dependent tumors, such as breast (8) and prostate cancer (9). GnRH and GnRH agonists are involved in
the regulation of proliferation, apoptosis and other biological
behaviors of ovarian cancer cells (10,11).
Previous studies, including ours, have revealed that GnRH agonists
inhibit the growth and proliferation of ovarian cancer through
GnRHR (10,12–14),
indicating that GnRH agonists and the GnRHR may be used as
antitumor strategies in the future (15).
However, the effect of GnRH agonists on the
apoptosis of EOC cells and their molecular mechanisms remain
largely unidentified. In this study, we aimed to investigate the
biological function of goserelin and the underlying mechanisms
involving goserelin and EOC apoptosis using in vitro and
in vivo experiments. The present study will facilitate a
better understanding of the mechanisms underlying the effect of
GnRH agonists on EOC. Meanwhile, key factors in the regulatory
pathways may provide new therapeutic targets for the treatment of
EOC.
Materials and methods
Cell lines and reagents
The human EOC cell lines (SKOV3, SKOV3-ip and A2780)
were kindly provided by the University of Texas MD Anderson Cancer
Center (Houston, TX, USA) and were authenticated by Short Tandem
Repeat (STR) profiling. All cells were cultured in RPMI-1640 medium
with 10% fetal bovine serum (FBS) (both from Gibco, Grand Island,
NY, USA) and 100 U/ml penicillin, 100 mg/ml streptomycin at 5%
CO2 in a 37°C humidified atmosphere. The GnRH agonist
goserelin acetate was purchased from Sigma-Aldrich (St. Louis, MO,
USA) and dissolved in phosphate-buffered solution (PBS, in
vitro) or normal saline (NS, in vivo).
Goserelin stimulation
Monoplast suspensions of SKOV3, SKOV3-ip or A2780
cells were seeded into 6-well plates at a density of
2×105 cells/ml and a total volume of 2 ml. Twenty-four
hours later, EOC cells were starved in FBS-free medium for 18 h.
Then, the cells were stimulated with different concentrations
(10−9, 10−8, 10−7,
10−6, 10−5, 10−4 or
10−3 mol/l) of goserelin (for the flow cytometry) or
10−4 mol/l goserelin (for apoptosis assay, human
apoptosis gene PCR array, qRT-PCR and the relative protein assay)
for different time courses. PBS was used as a control. The
transfected cells were also starved for 18 h and then stimulated
with 10−4 mol/l goserelin for another 48 h. The cells
were harvested and used for flow cytometry; meanwhile the protein
or mRNA were extracted for western blot or qRT-PCR analyses.
Apoptosis assay
Apoptotic cells were identified by Annexin V-PE
Apoptosis Detection kit (BD Biosciences, Franklin Lakes, NJ, USA)
or the Hoechst staining kit (Beyotime Institute of Biotechnology,
Shanghai, China) according to the manufacturers instructions. EOC
cells were starved in FBS-free medium for 18 h before treatment
with goserelin or PBS. After incubating for the indicated periods
of time, the cells were harvested and washed twice with cold PBS
and resuspended in binding buffer at a concentration of
1×106 cell/ml. PE Annexin V (5 µl) and 7-AAD (5 µl) were
added and then incubation was carried out for 15 min in the dark. A
flow cytometer and FlowJo software (Flowjo LLC, Ashland, OR, USA)
were used to analyze the staining data. Meanwhile, following the
corresponding treatment, cells incubated in 6-well plates were
stained with Hoechst staining at room temperature in the dark for 5
min and subsequently observed under confocal microscopy (Leica
Microsystems, Wetzlar, Germany) with an excitation wavelength of
350 nm and an emission wavelength of 460 nm.
Western blotting
Cells were harvested in RIPA lysis buffer containing
PMSF and phosphatase inhibitor (Sangon Biotech Co., Ltd., Shanghai,
China) and the protein concentration was quantified using a BCA
assay kit (Beyotime Institute of Biotechnology). Equal amounts of
protein (40 µg/lane) were separated on SDS-PAGE gels and
transferred to PVDF membranes. The membranes were blocked with 5%
non-fat milk for 1 h and incubated with primary antibodies at 4°C
overnight. After incubating with an HRP-conjugated secondary
antibody, the bands were visualized using an ECL detection system
(Thermo Fisher Scientific, Waltham, MA, USA). Antibodies including
rabbit anti-cleaved-caspase-3 monoclonal antibody (cat. no. 9664,
1:1,000), rabbit anti-p-AKT (Ser473) monoclonal antibody (cat. no.
4060, 1:1,000), rabbit anti-AKT monoclonal antibody (cat. no. 4691,
1:1,000), rabbit anti-FOXO1 monoclonal antibody (cat. no. 2880,
1:1,000), rabbit anti-BIK polyclonal antibody (cat. no. 4592,
1:1,000) and rabbit anti-TNFRSF9 polyclonal antibody (cat. no.
62634, 1:1,000) were purchased from Cell Signaling Technology
(Danvers, MA, USA). Rabbit anti-GnRHR polyclonal antibody (cat. no.
ab183079, 1:1,000), rabbit anti-cleaved-PARP monoclonal antibody
(cat. no. ab32064, 1:1,000) and rabbit anti-CIDEA polyclonal
antibody (cat. no. ab151577, 1:1,000) were purchased from Abcam
(Cambridge, UK).
siRNA and cell transfection
siRNAs were designed and purchased from Shanghai
GenePharma Co., Ltd., Technology (Shanghai, China) and are listed
in Table I. FOXO1-siRNA, AKT-siRNA
and scrambled siRNA were transfected into cells using Lipofectamine
2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturers
protocol. Briefly, 2×105 SKOV3-ip cells/well were seeded
into 6-well plates for 24 h when the cells had reached 40–50%
confluence. Before transfection, the RPMI-1640 medium without FBS
and antibiotics were replaced. A total of siRNA (75 pmol) and 5 µl
of Lipofectamine 2000 were diluted in 125 µl Opti-MEM and incubated
for 5 min at room temperature, respectively. The diluted siRNA and
diluted Lipofectamine 2000 were mixed gently and incubated for 20
min. A total of 250 µl mixed-combination was added to each well in
a final volume of 2 ml/well.
 | Table I.siRNAs used in the study. |
Table I.
siRNAs used in the study.
| Name | Sequence |
|---|
| FOXO1-siRNA-1 |
5-CCCUCGAACUAGCUAAAUTT-3 |
|
|
5-CCCUCGAACUAGCUAAAUTT-3 |
| FOXO1-siRNA-2 |
5-CCCAGUCUGUCUGAGAUAATT-3 |
|
|
5-UUAUCUCAGACAGACUGGGTT-3 |
| AKT-siRNA-1: |
5-AGGAAGUCAUCGUGGCCAATT-3 |
|
|
5-UUGGCCACGAUGACUUCCUTT-3 |
| AKT-siRNA-2 |
5-GCACUUUCGGCAAGGUGAUTT-3 |
|
|
5-AUCACCUUGCCGAAAGUGCTT-3 |
Human apoptosis gene PCR array and
qRT-PCR
SKOV3-ip cells were starved in FBS-free medium for
18 h before treatment with goserelin or PBS. After 48 h of
incubation, total RNA was extracted using an RNeasy®
Mini kit (Qiagen, Hilden, Germany) and then converted into cDNA
using an RT2 First Strand kit (Qiagen) following the
manufacturers instructions. Apoptosis-associated gene expression
profiles were evaluated using RT2 Profiler™ PCR Arrays
(Qiagen), which assess 84 genes involved in different apoptosis
pathways. In addition, qRT-PCR was performed to confirm the
differential expression of apoptosis-related genes using the ABI
7000 Real-Time PCR system (Applied Biosystems, Foster City, CA,
USA) with an RT-PCR kit (Takara Bio, Shiga, Japan) according to the
manufacturers protocol. The primers (Sangon Biotech) used in
qRT-PCR are listed in Table II.
The PCR conditions included an initial denaturation step at 95°C
for 10 min and 45 cycles of 95°C for 5 sec and 60°C for 30 sec. All
the experiments were performed in triplicate, and all the samples
were normalized to β-actin expression. The expression fold changes
were calculated using the 2−ΔΔCt method.
| Table II.The primers used in the study. |
Table II.
The primers used in the study.
| Gene | Primer | Sequence (5–3) |
|---|
| FOXO1 | Forward |
CGGGTATGTAACTGAACTTG |
|
| Reverse |
GCTGTAGTTGCCTCTTTAAT |
| AKT1 | Forward |
CTTCAAGCCCCAGGTCAC |
|
| Reverse |
CGCTGTCCACACACTCCAT |
| GAPDH | Forward |
ACCACAGTCCATGCCATCAC |
|
| Reverse |
CCACCACCCTGTTGCTGTAG |
| BCL2L10 | Forward |
GGATTCCGTGCTCTCCGACA |
|
| Reverse |
TCGCCCTCCTGCTCCTTTAG |
| BIK | Forward |
GGAGGTTCTTGGCATGACTGA |
|
| Reverse |
GCTCACGTCCATCTCGTCC |
| CASP14 | Forward |
CCACGGTAGAGGGATACATCG |
|
| Reverse |
GAGGGTGCTTTGGATTTCAGG |
| FASLG | Forward |
CTTCCCTGTCCAACCTCTGTG |
|
| Reverse |
CTACCAAGGCAACCAGAACCA |
| IL10 | Forward |
AAGCTGAGAACCAAGACCCAG |
|
| Reverse |
ATCGATGACAGCGCCGTAG |
| PYCARD | Forward |
TTGGACCTCACCGACAAGC |
|
| Reverse |
ATGTCGCGCAGCACGTTA |
| TNFRSF1B | Forward |
CACCGGGAGCTCAGATTCTTC |
|
| Reverse |
CACTGTGAGCTGTGGTCAGA |
| TNFSF8 | Forward |
TGTTGGTCGTTCAGAGGACG |
|
| Reverse |
TGGAGGTAGGCCCATGACTT |
| TP73 | Forward |
GGAGGGACTTCAACGAAGGA |
|
| Reverse |
GGGTCATCCACATACTGCGA |
In vivo study
Five-week-old specific-pathogen free (SPF) female
nude mice, weighing 18–20 g, were purchased from Shanghai SLAC
Laboratory Animal Corporation. The mice were cared for in
accordance with the criteria of the Medical Laboratory Animal
Administrative Committee of Shanghai and the Guide for the Care and
Use of Laboratory Animals. All experimental procedures were
approved by the Ethics Committee of Obstetrics and Gynecology
Hospital of Fudan University.
SKOV3 (1×107/ml) cells were
heterotransplanted into 14 nude mice, 6 of which were injected with
SKOV3 cells labelled with CM-DiI (Invitrogen). Six CM-DiI-labelled
nude mice and the other 8 nude mice were divided randomly into 2
groups respectively: the Goserelin group was subcutaneously
injected daily with 100 µg of goserelin in 200 µl of NS (14,16,17);
the NS group was subcutaneously injected with 200 µl of NS per day.
Subcutaneous xenograft tumors in nude mice were monitored by an
in vivo imaging system. After 19 days of treatment, the mice
were sacrificed, and the xenograft tumors were fixed in 4%
paraformaldehyde for immunohistochemistry and TUNEL staining. The
weights of the nude mice, TUNEL staining and immunohistochemical
staining for FOXO1, AKT/p-AKT and GnRHR expression in the xenograft
tumors were compared between the two groups.
CM-DiI for long-term cellular
labeling
Stock solutions of CM-DiI were prepared in DMSO at 1
mg/ml. SKOV3 cells were harvested and washed twice with PBS and
resuspended in PBS at a concentration of 1×106 cells/ml.
An appropriate 1 mg/ml stock solution was added into the cell
suspension at a 1 µg/ml final working concentration and incubated
for 6 min at 37°C and then for an additional 15 min at 4°C.
Labelled SKOV3 cells were washed twice with PBS again and
resuspended in PBS at a concentration of 1×107 cells/ml.
A drop of labelled SKOV3 cells was placed on a slide and the
efficiency of cell labelling was determined to be 98% under a
fluorescence microscope.
Immunohistochemistry (IHC) and TUNEL
staining
The xenograft tumors were fixed in 4%
paraformaldehyde and dehydrated through a serial alcohol gradient
and then embedded in paraffin. After sectioning (5-µm thick), the
tissues were immunostained with anti-FOXO1 (1:50), anti-AKT
(1:200), anti-p-AKT (1:50) and anti-GnRHR (1:300) antibodies using
a Histostain-Plus IHC kit (NeoBioscience, Shanghai, China) and
subjected to TUNEL staining using a TUNEL Apoptosis Assay kit
(Roche Diagnostics, Indianapolis, IN, USA) following the
manufacturers protocol. The immunostaining was evaluated by
determining the immunoreactive score (IRS). The IRS was calculated
by multiplying the staining intensity (SI) by the percentage of
positive cells (PP). SI was defined as 0 (negative), 1 (weak), 2
(moderate) and 3 (strong). PP was defined as 0 (negative), 1 (≤10%
positive cells), 2 (11–50% positive cells), 3 (51–75% positive
cells) and 4 (>75% positive cells). IRS = SI × PP, and an IRS
>3 was defined as positive (18).
Statistical analysis
Stata 14.0 (StataCorp LP, College Station, TX, USA)
and GraphPad Prism 6.0 (GraphPad Software Inc., La Jolla, CA, USA)
were used for the statistical analyses. Continuous data are
expressed as the mean ± SD, and analyzed by independent t-test
between two groups or one-way ANOVA among multiple groups. The
categorical data were compared using the Chi-squared or Fishers
exact tests as appropriate. Differences were considered
statistically significant at P<0.05.
Results
Goserelin promotes the apoptosis of
EOC cells in vitro
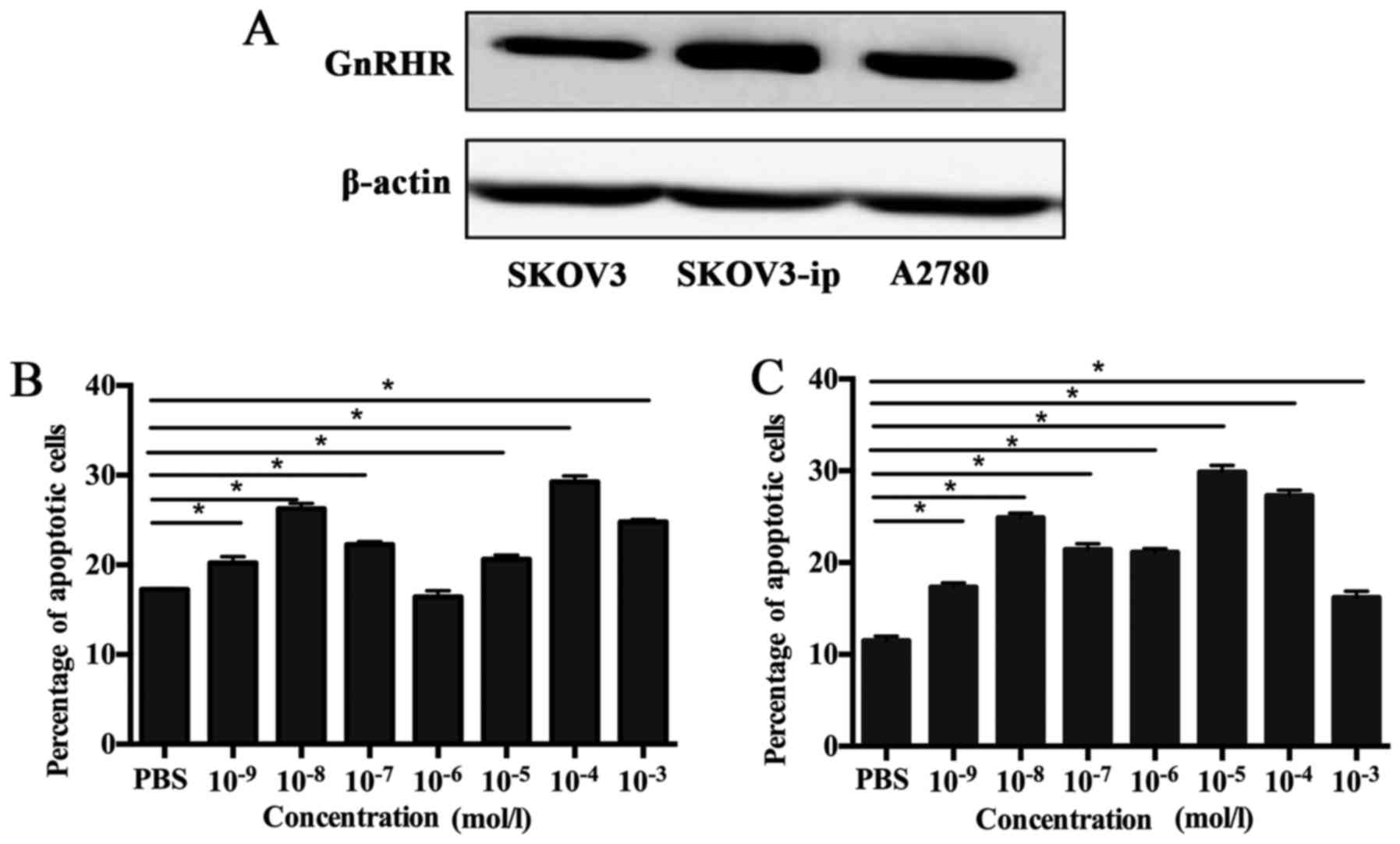
SKOV3, SKOV3-ip and A2780 cells all expressed GnRHR
(Fig. 1A). Apoptosis detected by
flow cytometry showed that different concentrations of goserelin
increased the proportion of apoptotic SKOV3-ip cells compared with
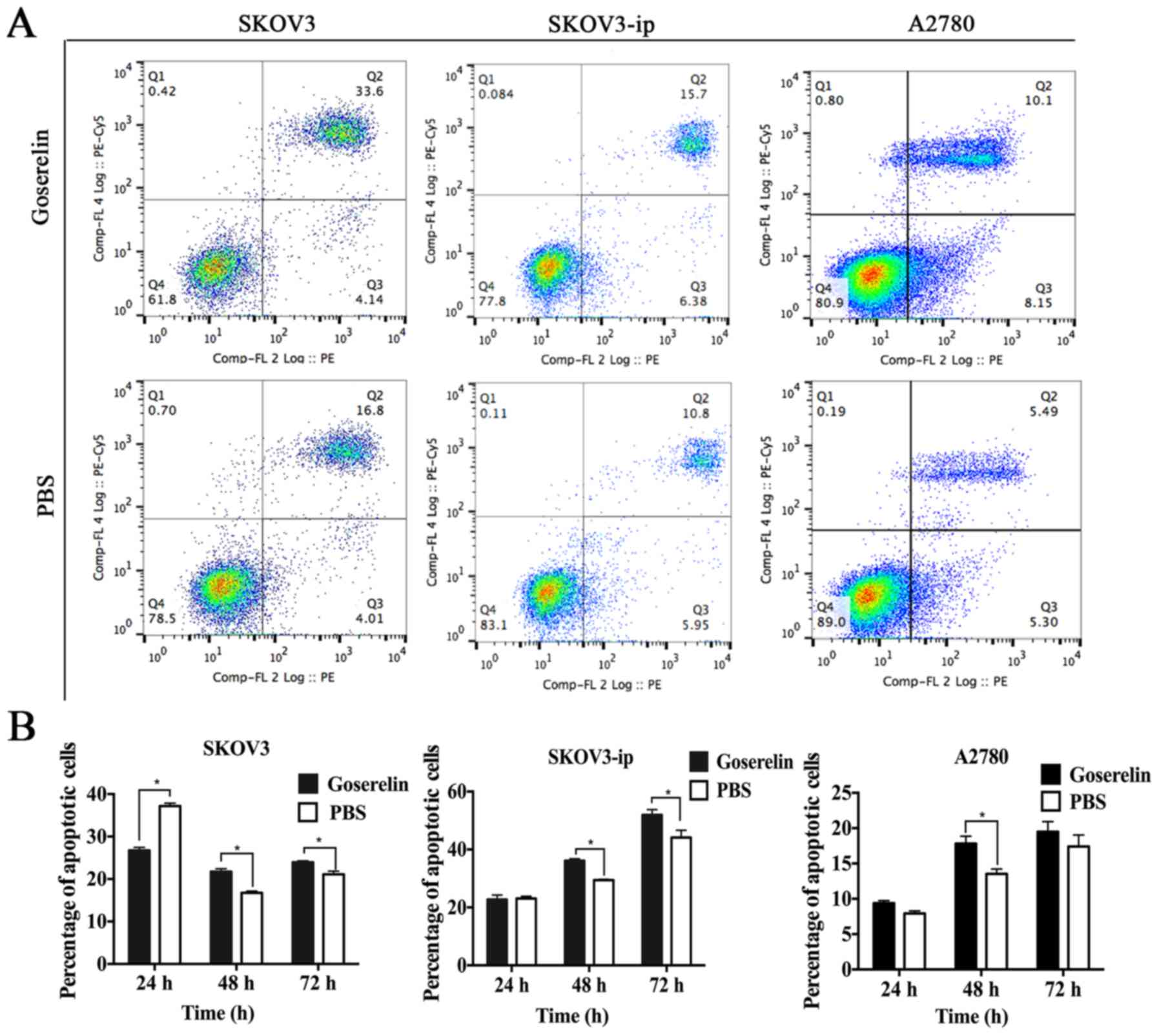
the control group at 48 and 72 h (P<0.05; Fig. 1B and C). A concentration of
10−4 mol/l goserelin significantly increased the total
apoptosis rate of SKOV3-ip, SKOV3 and A2780 cells (P<0.05;
Fig. 2A and B). To further clarify
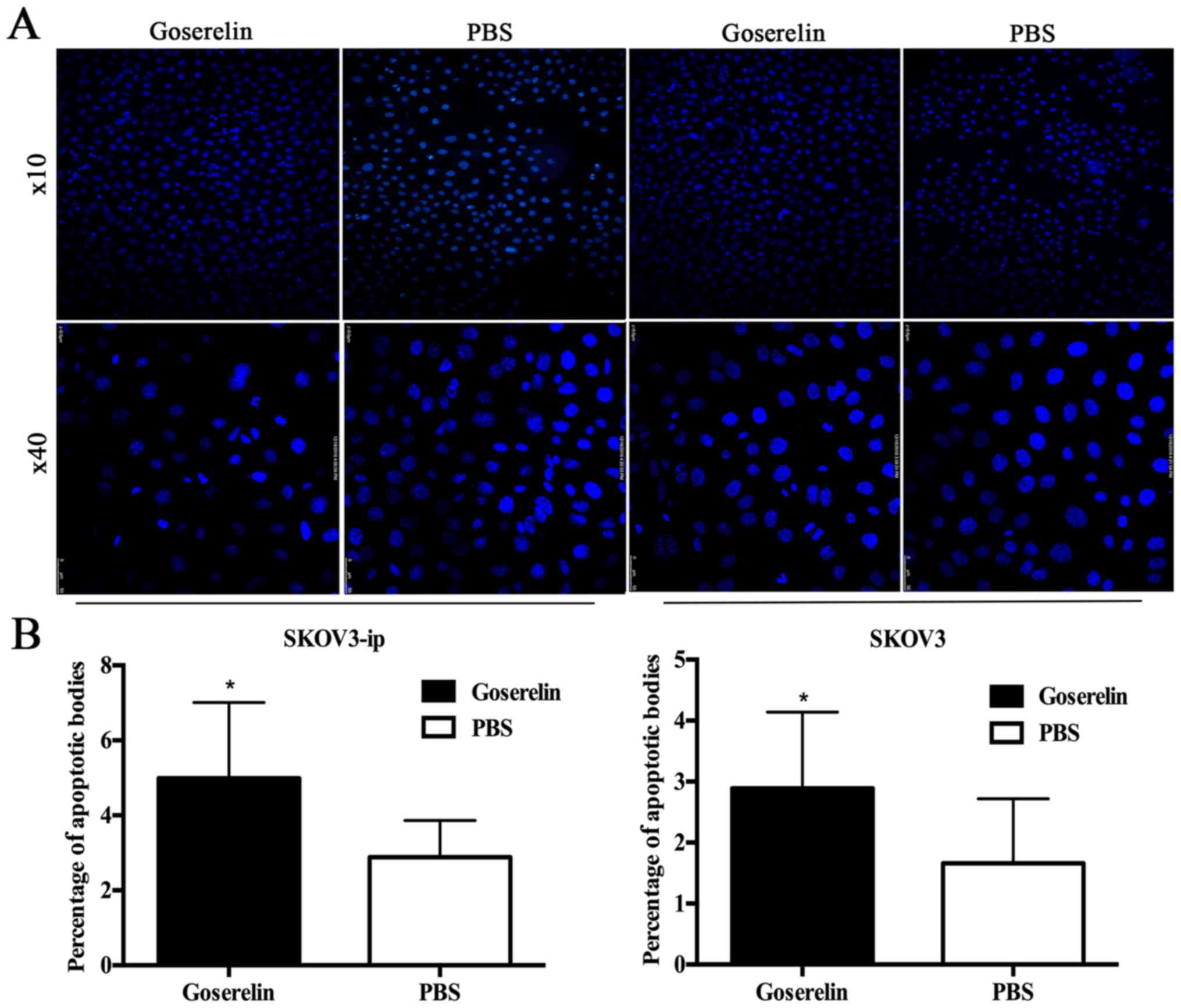
the effect of goserelin on apoptosis, we treated the cells with
10−4 mol/l goserelin for 24, 48 and 72 h. Hoechst
staining showed that the number of apoptotic bodies was
significantly increased in the SKOV3 and SKOV3-ip cells compared
with the control group (P<0.05; Fig.
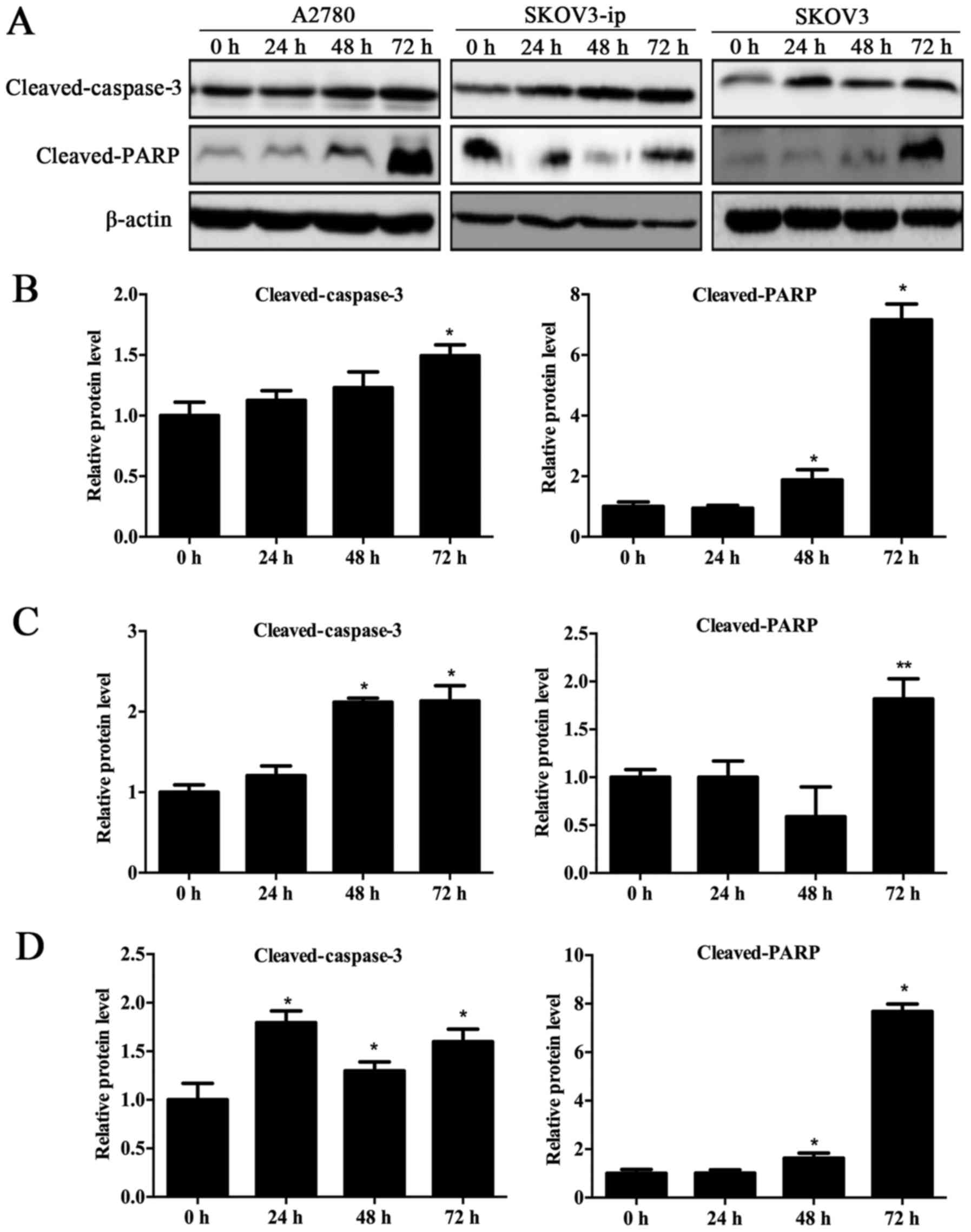
3). Furthermore, the expression of cleaved-caspase-3 and
cleaved-PARP were observably increased after treatment with
10−4 mol/l goserelin (P<0.05; Fig. 4). Taken together, these results
suggested that goserelin promoted EOC cell apoptosis.
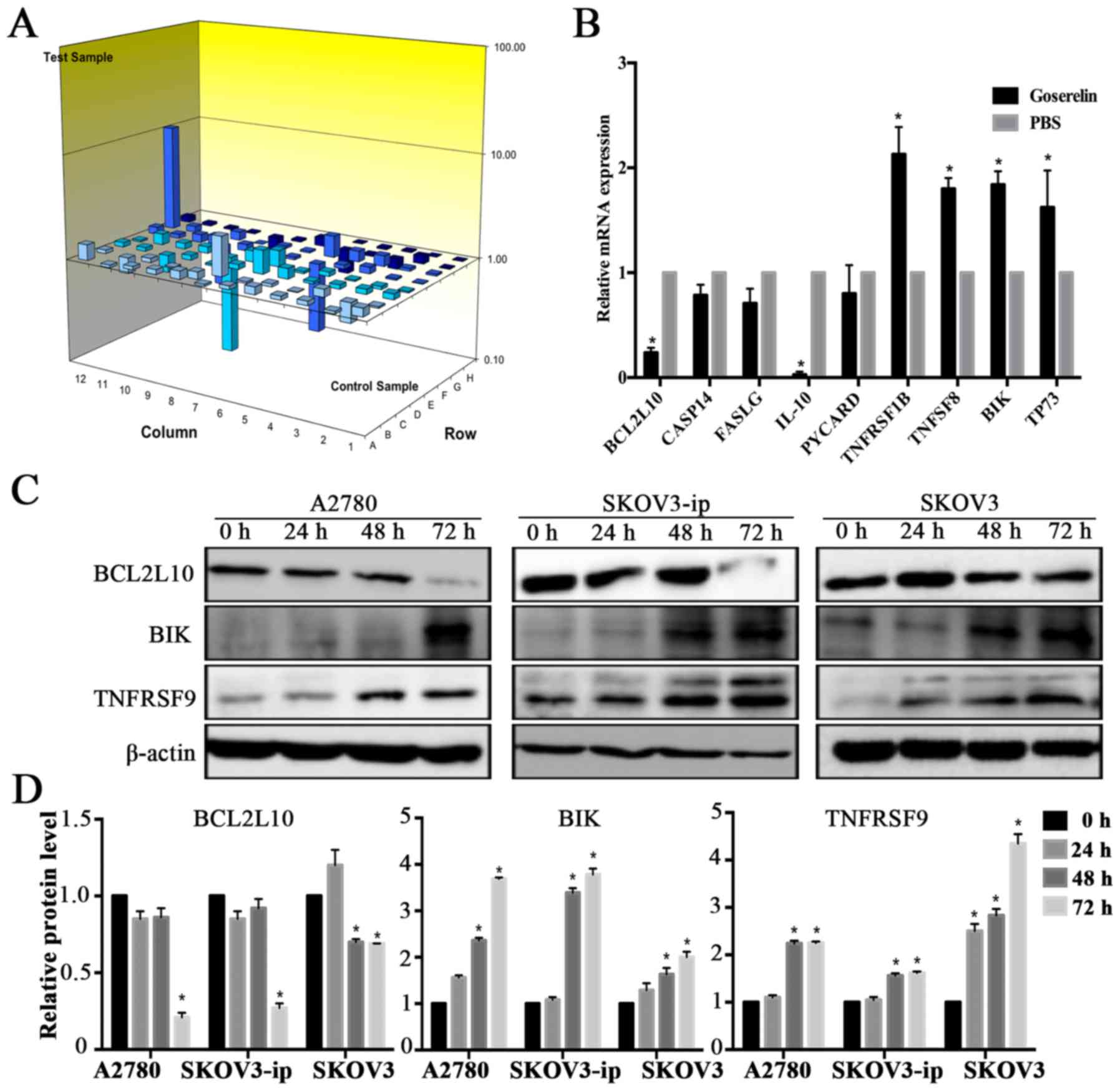
Expression of human apoptosis-related
genes is regulated by goserelin in SKOV3-ip cells
To investigate the downstream molecular process
involving goserelin and EOC apoptosis, human apoptosis gene PCR
arrays were tested in the SKOV3-ip-PBS and SKOV3-ip-Goserelin
group. According to the apoptosis gene PCR array, 9 genes were
obviously expressed (fold change ≥1.5) in the SKOV3-ip-Goserelin
group compared with the SKOV3-ip-PBS group (Table III). qRT-PCR and western blot
analyses were performed to confirm the expression of these genes,
including members of the tumor necrosis factor (TNF) and TNF
receptor superfamilies (such as TNFRSF9, TNFRSF1B and TNFSF8). The
results were mostly consistent with those of the PCR array analysis
(Fig. 5A-D).
| Table III.Nine differentially expressed genes
(fold change ≥1.5) were screened in the SKOV3-ip-Goserelin
group. |
Table III.
Nine differentially expressed genes
(fold change ≥1.5) were screened in the SKOV3-ip-Goserelin
group.
| Gene name | GeneBank ID | Description | Fold change |
|---|
| BCL2L10 | NM_020396 | BCL2-like 10
(apoptosis facilitator) | −1.53 |
| BIK | NM_001197 | BCL2-interacting
killer (apoptosis-inducing) | 2.38 |
| CASP14 | NM_012114 | Caspase-14,
apoptosis-related cysteine peptidase | 1.67 |
| FASLG | NM_000639 | Fas ligand (TNF
superfamily, member 6) | −4.49 |
| IL10 | NM_000572 | Interleukin 10 | −2.39 |
| PYCARD | NM_013258 | PYD and CARD domain
containing | 1.6 |
| TNFRSF1B | TNFRSF1B | Tumor necrosis
factor receptor superfamily, member 1B | 10.57 |
| TNFSF8 | NM_001244 | Tumor necrosis
factor (ligand) superfamily, member 8 | 1.68 |
| TP73 | NM_005427 | Tumor protein
p73 | −1.93 |
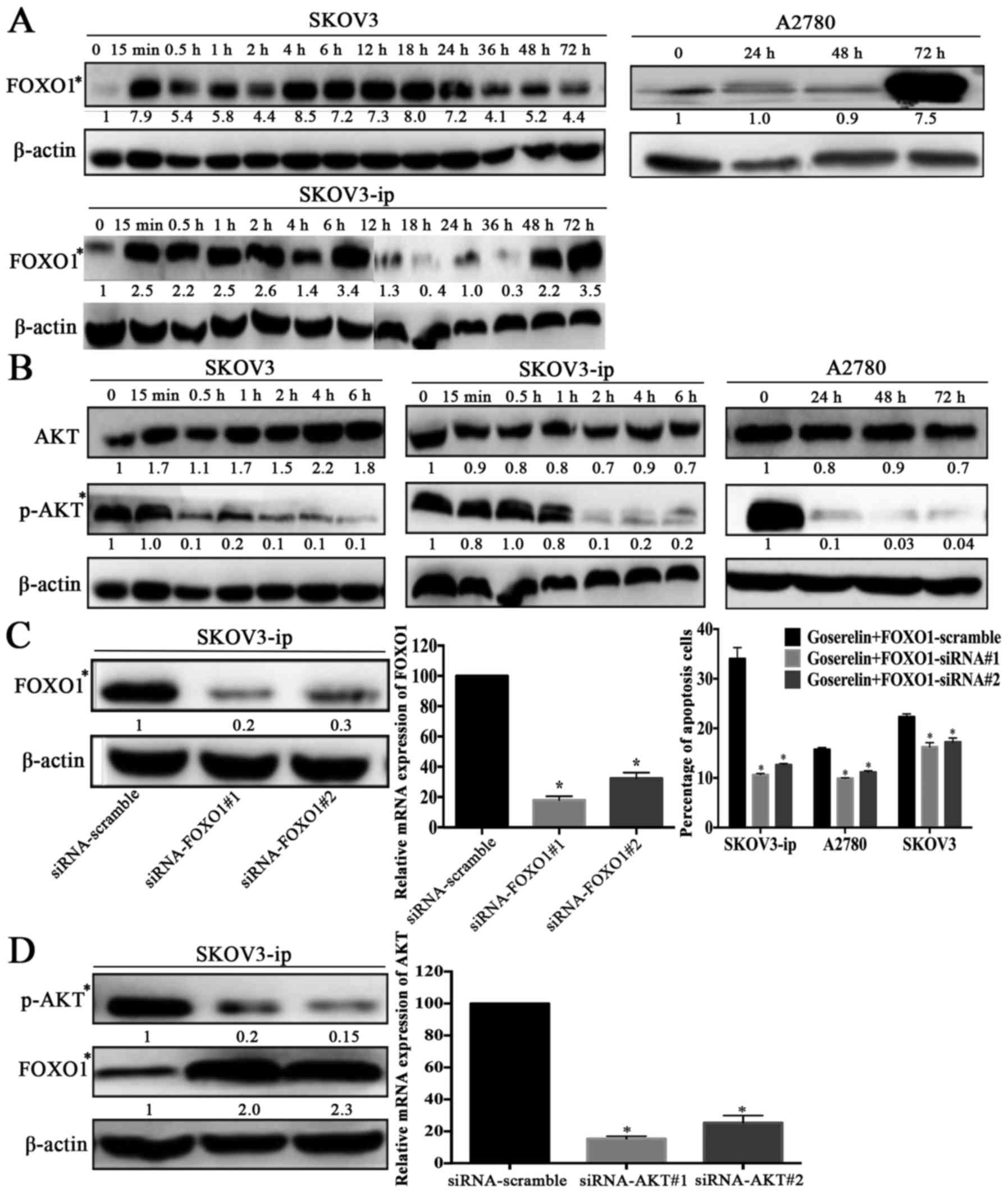
Goserelin influences EOC cell
apoptosis by upregulating FOXO1 through the PI3K/AKT signaling
pathway
TNF and TNF receptors are reportedly regulated by
FOXO1 (19). We then detected the
expression of FOXO1 by goserelin at 10−4 mol/l. As shown
in Fig. 6A, FOXO1 expression was
significantly increased (P<0.05) and the goserelin-mediated
promotion of apoptosis was abrogated by FOXO1-siRNA (P<0.05;
Fig. 6C).
As FOXO1 is a downstream target of the PI3K/AKT
signaling pathway (20,21), the expression of AKT/p-AKT was
analyzed. As shown in Fig. 6B,
goserelin downregulated p-AKT (Ser473) expression. Meanwhile, the
expression of FOXO1 was increased by AKT-siRNA (P<0.05; Fig. 6D). Taken together, these data
suggest that goserelin may promote EOC cell apoptosis by
upregulating FOXO1 through the PI3K/AKT signaling pathway.
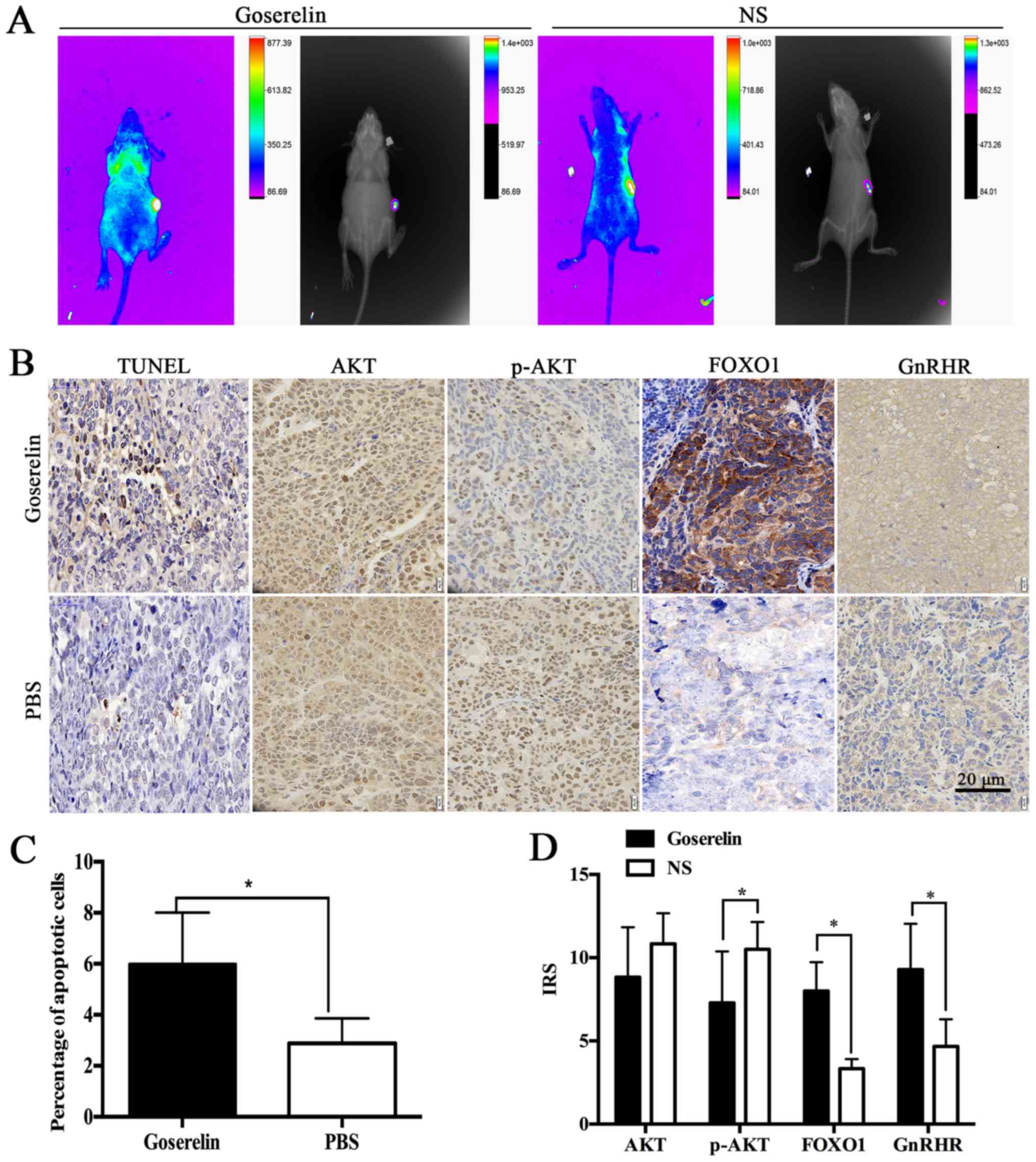
Effect of goserelin on subcutaneous
xenograft tumors in nude mice
In vivo images of subcutaneous xenograft
tumors in nude mice revealed that the subcutaneous xenograft tumor
model was constructed successfully (Fig. 7A). TUNEL staining showed that
goserelin significantly increased the proportion of apoptotic cells
in the subcutaneous xenograft tumors (P<0.05; Fig. 7B and C). Immunohistochemistry
results showed that the difference in AKT expression between the
two groups was not significant. In the goserelin group, p-AKT
expression was significantly lower, while the expression of FOXO1
and GnRHR were significantly increased compared with the NS group
(P<0.05; Fig. 7B and D).
Therefore, the expression of the key factors in the PI3K/AKT-FOXO1
pathway was consistent with that observed in vitro.
Discussion
According to the gonadotropin theory (5), gonadotropins, including LH and FSH,
are conducive to the growth of ovarian cancer, which can be
downregulated by GnRH agonists. Theoretically, GnRH agonists
inhibit the growth and proliferation of ovarian cancer. Several
studies, including ours, have revealed that GnRH agonists can
directly inhibit the growth and proliferation of various human
cancer cell lines, including ovarian cancer cell lines, through the
GnRHR in a dose- and time-dependent manner (8–10,12–14).
In the present study, we demonstrated that SKOV3, SKOV3-ip and
A2780 cells all expressed GnRHR, which may be required for
goserelin to act on these three cell lines. We found that goserelin
promoted EOC cell apoptosis, which was verified by different
methods in three EOC cell lines. In vivo, goserelin also
promoted the apoptosis of xenograft tumors. Considering its less
adverse side-effects compared with chemotherapeutic drugs,
goserelin may be a safe and effective antitumor strategy in the
future.
It has been reported that the apoptosis of human
ovarian cancer cells may be regulated by goserelin (13,22).
We used human apoptosis gene PCR array to explore the downstream
molecular events involving goserelin and EOC apoptosis. Goserelin
regulated several apoptosis-related factors, especially members of
the TNF and TNF receptor superfamilies, which are regulated by
FOXO1 (19). The FOXO transcription
factors are considered to be tumor suppressors that inhibit cell
proliferation and induce apoptosis (21). Indeed, FOXO1 plays multiple roles in
various types of human cancers, including skin (23), liver (24) and breast cancer (25). An increasing number of studies have
linked the tumor suppressor activity of FOXOs to the regulation of
genes involved in cell death (e.g., Fasl) (26) and cell cycle arrest (e.g.,
p27KIP1) (27). In
agreement with these reports, we found that goserelin increased
FOXO1 expression. Meanwhile, the pro-apoptotic effect of goserelin
was abrogated by FOXO1-siRNA. These results indicate that goserelin
induces EOC cell apoptosis by modulating FOXO1 activity.
It has been reported that AKT phosphorylation
following growth factor stimulation can lead to FOXO inactivation
(28). In breast cancer cell lines,
the targeted depletion of PI3K using siRNA reactivates FOXO1, 3 and
4, which then induces cell cycle arrest and apoptosis (29). The same anti-proliferation and
pro-apoptotic effects were observed when FOXO1 was reactivated in
cervical cancer treated with the PI3K inhibitor LY294002 (30). We hypothesized that goserelin may
promote EOC cell apoptosis by upregulating FOXO1 through the
PI3K/AKT signaling pathway. As expected, goserelin down-regulated
p-AKT expression, and FOXO1 upregulation was dependent on PI3K/AKT.
The mechanism by which goserelin activates the AKT signaling
pathway requires further exploration. In vivo, the
expression of key factors in the PI3K/AKT-FOXO1 pathway was
consistent with that observed in vitro. Importantly, GnRHR
expression in the goserelin group increased significantly,
indicating that goserelin may exert its biological effects through
the GnRHR.
In summary, our results demonstrated the
pro-apoptotic effects of goserelin on EOC cells and showed for the
first time that goserelin may promote EOC cell apoptosis by
upregulating FOXO1 through the PI3K/AKT signaling pathway.
PI3K/AKT-FOXO1 pathway may be the novel therapeutic target for the
treatment of EOC. GnRH agonists may also be potential antitumor
agents.
Acknowledgements
The present study was supported by the Youth
National Natural Science Foundation of China (NSFC no.
81402152).
References
|
1
|
Hemminki K, Sundquist J and Brandt A:
Incidence and mortality in epithelial ovarian cancer by family
history of any cancer. Cancer. 117:3972–3980. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rustin G, van der Burg M, Griffin C, Qian
W and Swart AM: Early versus delayed treatment of relapsed ovarian
cancer. Lancet. 377:380–381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Casagrande JT, Louie EW, Pike MC, Roy S,
Ross RK and Henderson BE: ‘Incessant ovulation’ and ovarian cancer.
Lancet. 2:170–173. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Millar RP: GnRHs and GnRH receptors. Anim
Reprod Sci. 88:5–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leung PC and Choi JH: Endocrine signaling
in ovarian surface epithelium and cancer. Hum Reprod Update.
13:143–162. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brekelmans CT: Risk factors and risk
reduction of breast and ovarian cancer. Curr Opin Obstet Gynecol.
15:63–68. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim HJ, Yoon TI, Chae HD, Kim JE, Chae EY,
Yu JH, Sohn G, Ko BS, Lee JW, Son BH, et al: Concurrent
gonadotropin-releasing hormone agonist administration with
chemotherapy improves neoadjuvant chemotherapy responses in young
premenopausal breast cancer patients. J Breast Cancer. 18:365–370.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hoda MR, Kramer MW, Merseburger AS and
Cronauer MV: Androgen deprivation therapy with Leuprolide acetate
for treatment of advanced prostate cancer. Expert Opin
Pharmacother. 18:105–113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Jia L, Feng Y and Zheng W:
Overexpression of follicle-stimulating hormone receptor facilitates
the development of ovarian epithelial cancer. Cancer Lett.
278:56–64. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Z, Liao H, Chen X, Zheng Y, Liu Y,
Tao X, Gu C, Dong L, Duan T, Yang Y, et al: Luteinizing hormone
upregulates survivin and inhibits apoptosis in ovarian epithelial
tumors. Eur J Obstet Gynecol Reprod Biol. 155:69–74. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
So WK, Cheng JC, Poon SL and Leung PC:
Gonadotropin-releasing hormone and ovarian cancer: A functional and
mechanistic overview. FEBS J. 275:5496–5511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meyer C, Sims AH, Morgan K, Harrison B,
Muir M, Bai J, Faratian D, Millar RP and Langdon SP: Transcript and
protein profiling identifies signaling, growth arrest, apoptosis,
and NF-κB survival signatures following GNRH receptor activation.
Endocr Relat Cancer. 20:123–136. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Ding JX, Tao X, Lu ZY, Wang JJ,
Feng WW and Hua KQ: Goserelin can inhibit ovarian cancer
proliferation and simultaneously protect ovarian function from
cisplatin: An in vitro and in vivo study. J Chemother. 25:96–103.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aguilar-Rojas A, Pérez-Solis MA and
Maya-Núñez G: The gonadotropin-releasing hormone system:
Perspectives from reproduction to cancer (Review). Int J Oncol.
48:861–868. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yano T, Pinski J, Halmos G, Szepeshazi K,
Groot K and Schally AV: Inhibition of growth of OV-1063 human
epithelial ovarian cancer xenografts in nude mice by treatment with
luteinizing hormone-releasing hormone antagonist SB-75. Proc Natl
Acad Sci USA. 91:pp. 7090–7094. 1994; View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dondi D, Moretti RM, Montagnani Marelli M,
Pratesi G, Polizzi D, Milani M, Motta M and Limonta P:
Growth-inhibitory effects of luteinizing hormone-releasing hormone
(LHRH) agonists on xenografts of the DU 145 human
androgen-independent prostate cancer cell line in nude mice. Int J
Cancer. 76:506–511. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Friedrichs K, Gluba S, Eidtmann H and
Jonat W: Overexpression of p53 and prognosis in breast cancer.
Cancer. 72:3641–3647. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van der Vos KE and Coffer PJ: The
extending network of FOXO transcriptional target genes. Antioxid
Redox Signal. 14:579–592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cardozo T and Pagano M: The SCF ubiquitin
ligase: Insights into a molecular machine. Nat Rev Mol Cell Biol.
5:739–751. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Coomans de Brachène A and Demoulin JB:
FOXO transcription factors in cancer development and therapy. Cell
Mol Life Sci. 73:1159–1172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gründker C, Schulz K, Günthert AR and
Emons G: Luteinizing hormone-releasing hormone induces nuclear
factor kappaB-activation and inhibits apoptosis in ovarian cancer
cells. J Clin Endocrinol Metab. 85:3815–3820. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsitsipatis D, Klotz LO and Steinbrenner
H: Multifaceted functions of the forkhead box transcription factors
FoxO1 and FoxO3 in skin. Biochim Biophys Acta. 1861:1057–1064.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carbajo-Pescador S, Mauriz JL,
García-Palomo A and González-Gallego J: FoxO proteins: Regulation
and molecular targets in liver cancer. Curr Med Chem. 21:1231–1246.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Procaccia S, Ordan M, Cohen I,
Bendetz-Nezer S and Seger R: Direct binding of MEK1 and MEK2 to AKT
induces Foxo1 phosphorylation, cellular migration and metastasis.
Sci Rep. 7:430782017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fu Z and Tindall DJ: FOXOs, cancer and
regulation of apoptosis. Oncogene. 27:2312–2319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ho KK, Myatt SS and Lam EW: Many forks in
the path: Cycling with FoxO. Oncogene. 27:2300–2311. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo
P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Reagan-Shaw S and Ahmad N: RNA
interference-mediated depletion of phosphoinositide 3-kinase
activates forkhead box class O transcription factors and induces
cell cycle arrest and apoptosis in breast carcinoma cells. Cancer
Res. 66:1062–1069. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Prasad SB, Yadav SS, Das M, Govardhan HB,
Pandey LK, Singh S, Pradhan S and Narayan G: Down regulation of
FOXO1 promotes cell proliferation in cervical cancer. J Cancer.
5:655–662. 2014. View
Article : Google Scholar : PubMed/NCBI
|