Introduction
Colorectal cancer (CRC) is the most common
gastrointestinal tumor malignancy (1). With the rapid speed of our country's
aging process, the incidence rate of CRC shows an upward trend
(2). At present, the causes of CRC
are considered the result of external environmental factors
combined with internal organism factors. Unhealthy lifestyle,
anti-oncogene inactivation and oncogene mutations can cause cells
to grow uncontrollably, and further lead preexisting diseases such
as ulcerative colitis and colonic adenoma to develop into malignant
tumor (3–6). Research has demonstrated that most
patients die from tumor metastasis and recurrence (7). The essential characteristics of
malignant tumors are excessive proliferation, differentiation
failure and apoptosis disorder (8).
Therefore, it is important to explore the mechanisms of tumor
growth, metastasis and recurrence in CRC. Ribophorin II (RPN2) is a
membrane glycoprotein which is found in rough endoplasmic
reticulum, located at chromosome 20q12-13.1 and has glycosylation
function affecting protein stability and secretion and play a key
role in cell function and signal transduction (9,10).
Research has indicated that RPN2 was highly expressed in tumor stem
cells (11). RPN2 promoted cellular
malignant proliferation in breast cancer by regulating
N-glycosylation of CD36 (12). In
addition, RPN2 interference reduced the glycosylation of
P-glycoprotein to promote docetaxel-dependent apoptosis in
esophageal squamous cell carcinoma (ESCC) (13). In osteosarcoma and gastric
carcinoma, studies have revealed that the expression of RPN2 was
closely associated with patient survival time and tumor stage
(14,15). It was also reported that RPN2 was
highly expressed in CRC (16).
Therefore, we hypothesized that RPN2 plays an important role in the
development and progression of CRC.
Signal transducer and activator of transcription
(STAT)3 belongs to the transcription factor family. STAT3 monomer,
is expressed in the cytoplasm (17). Research has indicated that STAT3 was
persistently activated in 50% of lung cancers (18). In addition, Janus kinase (JAK)2, as
a key factor in the process of STAT3 phosphorylation, can be bound
to the membrane receptor and trigger tyrosine receptor to activate
STAT3 (19). STAT3-mediated target
genes play an important role in the occurrence and development of
the tumor, including migration, invasion and angiogenesis (20,21).
In CRC, the activation of STAT3/JAK2 signaling pathway can promote
epithelial-mesenchymal transition (EMT) and enhance the abilities
of migration and invasion in many types of cancer (22). Therefore, we hypothesized that the
STAT3/JAK2 signaling pathway regulated the expression level of
related proteins to affect the development of CRC with the action
of RPN2.
Materials and methods
Patients and tissue samples
A total of 43 samples of CRC tissues and benign
tissues surgically removed from patients in Huai'an First People's
Hospital were collected from March 2014 to December 2017.
Preoperative clinical and pathological follow-up data were
completed by all patients. Ethical approval for the study was
provided by the Ethics Committee of Huai'an First People's
Hospital. Written informed consent was obtained from all the study
participants.
Immunohistochemistry
Tissue sections were initially treated with
deparaffinization and hydration and then heated in EDTA (pH 8.0)
and antigen-retrieved in 10-mm citrate buffer for 5 min at 100°C.
The reaction of RPN2 antibody (dilution 1:500; cat. no. ab64467;
Abcam, Cambridge, MA, USA) took place for 1 h at room temperature,
and then incubation followed by biotin-labeled secondary antibodies
(1:500; cat. no. PL0306214; PLlabs, Toronto, Canada) and slides
developed using DAB solution and counterstained with hematoxylin
staining (Baso Diagnostic, Inc., Wuhan, China). Immunohistochemical
signals were calculated with the positive staining cells under a
light microscope (Olympus Corporation, Tokyo Japan) at a
magnification of ×200.
Cell culture
Human colon epithelial NCM460 cells were obtained
from the Cell Engineering Research Center of The Fourth Military
Medical University (Xi'an, China) and were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS), 10 ng/ml epidermal growth
factor (EGF) (both from Invitrogen; Thermo Fisher Scientifc, Inc.),
1% insulin (First Biological and Chemical Medication, Co., Ltd.,
Shanghai, China), 5 µg/ml hydrocortisone (The Third Pharmaceutical,
Co., Beijing, China) and 1% penicillin/streptomycin at 37°C in a
humidified atmosphere of 5% CO2. Human colon cancer cell
lines (SW1116, SW480, LoVo, SW403 and SW620) were purchased from
SUER Bio-Technique, Co., Ltd. (Shanghai, China) and cultured in
Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) containing 10% FBS and 1%
penicillin/streptomycin at 37°C in a humidified atmosphere of 5%
CO2.
siRNA transfection
SW1116 and SW480 cells were seeded onto 6-well
culture plates at a density of 3×105 cells/well. The
RPN2 siRNA or control siRNA, were both purchased from Shanghai
GenePharma, Co., Ltd., (Shanghai, China) and were transfected into
cells at 50–60% confluency using Lipofectamine™ 2000 (Invitrogen,
Shanghai, China) following the manufacturer's protocol. After 48 h,
the transfected cells were collected and processed for the
subsequent experiments. The sequence of RPN2 siRNA used was:
forward,
5′-CCTAGAGGTACCGGAATGCGTTAAGCTATACCTGCATTGGCTAGTTAACGTAGAACCG-3′
and reverse,
5′-CCTACGTTAAGCAATTCAATTTTTTGTATAGCTTAACTACCGCATTTCGAGGTAGTAG-3′.
The sequence of the negative (mock) siRNA used was
5′-ACGCCUCCCGAACGUTTUUCUUGUCGUC-3′.
CCK-8 assay
Cell viability was evaluated using cell counting
kit-8 (CCK-8) assay. In brief, after 48 h of transfection, SW1116
and SW480 cells were seeded at a density of 4×103
cells/well in 96-well plates and incubated for 0, 12, 24, 48 and 72
h. Subsequently, 20 µl of CCK-8 was added to each well for another
1-h incubation. The optical density (OD) values were determined at
450 nm using a microplate reader (Thermo Fisher Scientific, Inc.).
All experiment concentrations were assessed in triplicate.
Cell cycle analysis
Flow cytometry was used for the analysis of the cell
cycle. After 48-h transfection, cells were harvested and then fixed
in ice-cold 70% ethanol (stored at −20°C) overnight. Subsequently,
the cells were washed with phosphate-buffered saline (PBS) prior to
resuspending in DNA staining solution [40 µg/ml propidium iodide
(PI), 250 µg/ml RNase in PBS with 2 mM EDTA] for 30 min at 37°C.
Cell cycle distribution was analyzed using flow cytometer
(FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA).
Apoptosis analysis
The apoptosis level of SW1116 and SW480 cell
transfection with RPN2 siRNA or control siRNA was assessed by flow
cytometric assay. The overall cell apoptosis detection procedures
were similar to the previously described procedures with slight
modification on the cell numbers (23). In brief, the cells were washed and
detached in 2 ml PBS with 2 mM ethylenediaminetetraacetic acid
(EDTA) and centrifuged at 15,000 × g for 5 min. The suspended cells
were stained with 250 µl of hypotonic fluorochrome solution
including PBS, 50 µg of PI, 0.1% sodium citrate and 0.1% Triton
X-100 and RNase A (100 U/ml) (BD Biosciences) for 30 min in the
dark at room temperature. Measurements were made using a flow
cytometer (BD Influx; BD Biosciences). B3 quadrant represented
viable cells and B2 and B4 quadrants represented apoptotic
cells.
Cell invasion and migration
assays
The invasion and migration activity of the SW1116
and SW480 cells transfected with RPN2 siRNA or control siRNA were
assessed by a 24-well Transwell chamber coated with or without
Matrigel (BD Biosciences) on the upper surface of the membrane,
with a pore size of 8 µm (Sigma-Aldrich; Merck KGaA). In brief, the
transfected SW1116 and SW480 cells (1×104 cells/well)
were suspended in culture media (100 µl; serum free) and then
placed in the upper Transwell chamber. The lower chamber was filled
with medium containing 10% FBS. After 24-h incubation, the cells
that had invaded or migrated through the membrane to the lower
surface were fixed, stained and counted visually under a microscope
(Olympus IX71-A12FL/PH; Olympus Corporation).
Western blot analysis
Cells were seeded at a density of 5×105
cells/well in 6-well plates after transfection with RPN2 siRNA and
control siRNA for 48 h. Each group of cells was harvested and
washed twice with PBS and protein lysed in ice-cold radio
immunoprecipitation assay buffer (RIPA; Beyotime Institute of
Biotechnology, Shanghai, China) with freshly added 0.01% protease
inhibitor PMSF (Amresco, Shanghai, China) and incubated on ice for
30 min. Cell lysis was centrifuged at 10,000 × g for 5 min at 4°C
and the supernatant (20–30 µg of protein) was run on 10% SDS-PAGE
gel and electrophoretically transferred onto a nitrocellulose (NC)
membrane (Millipore, Shanghai, China), and then detected with RPN2
(1:800; cat. no. ab64467), caspase-3 (1:1,000; cat. no. ab13847),
cyclin D1 (1:800; cat. no. ab16663), E-cadherin (1:800; cat. no.
ab15148), MMP-2 (1:1,200; cat. no. ab37150), TIMP-2 (1:1,000; cat.
no. ab157386), STAT3 (1:1,000; cat. no. ab68153), phosphorylated
(p-) STAT3 (1:800; cat. no. ab32143), JAK2 (1:1,200; cat. no.
ab108596), p-JAK2 (1:1,000; cat. no. ab32101) proteins. Protein
loading was estimated using mouse anti-GAPDH monoclonal antibody
(1:2,500; cat. nos. AG019 and AF006; Beyotime Institute of
Biotechnology). Blots were visualized using enhanced
chemiluminescence (ECL; Thermo Fisher Scientific, Shanghai,
China).
RNA isolation and RT-PCR
The expression of genes was evaluated using RT-PCR
and SYBR Green I chemistry (TransStart Top Green qPCR SuperMix;
TransGen Biotech, Co., Ltd., Beijing, China). Cells were seeded in
6-well plates at a density of 5×105 cells/well, cultured
overnight and then treated with siRNA-RPN2 for 48 h. Total RNA (2
µg) was extracted from cells using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol, and was reverse transcribed with the TransCript One-step
gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotech, Co.,
Ltd.). The expression of RPN2, caspase-3, cyclin D1, E-cadherin,
MMP-2 and TIMP-2 mRNA in SW1116 and SW480 cells was detected by
RT-PCR with the cycling parameters defined as follows: an initial
cycling for 5 min at 95°C, followed by 40 cycles at 95°C for 15
sec, 30 sec at 60°C and 30 sec at 72°C. The primers used for the
amplification of the indicated genes were designed using the Primer
Express software (Applied Biosystems; Thermo Fisher Scientific,
Foster City, CA, USA). Primers used were as follows: RPN2, forward,
5′- CAAAGTCACCGGACAAGGTC-3′ and reverse, 5′-TGGTGTTCCGAAGTTGGTCA-3′
(product: 142 bp); caspase-3, forward, 5′-CTATGGCTACGCTGTGGAGA-3′
and reverse, 5′-TGTAGAAGCCAGGGCTCATC-3′ (product: 110 bp); cyclin
D1, forward, 5′-ATAGATGCCAGGCAGAGACC-3′ and reverse,
5′-ACTGCCTGTTCCCACTACTC-3′ (product: 123 bp); E-cadherin, forward,
5′-TGCCAACTGGCTGGAGATTA-3′ and reverse, 5′-AGTGTCCCTGTTCCAGTAGC-3′
(product: 156 bp); MMP-2, forward, 5′-CTACTGAGTGGCCGTGTTTG-3′ and
reverse, 5′-TCCCTGAGGTTCTCTTGCTG-3′ (product: 174 bp); TIMP-2,
forward, 5′-AGCATTTGACCCAGAGTGGA-3′ and reverse,
5′-GGAGAACCAAAGACGGGAGA-3′ (product: 111 bp); GAPDH, forward,
5′-ACCCAGAAGACTGTGGATGG-3′ and reverse, 5′-TCAGCTCAGGGATGACCTTG-3′
(product: 124 bp). The relative expression levels were analyzed
using the 2−ΔΔCt method. GAPDH was used as internal
control to monitor RT-PCR efficiency. All RT reactions were
performed in triplicate.
Statistical analysis
All results are presented as the mean ± SD of three
independent experiments and the data were processed with SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA). Survival analysis was used
in the analysis of the information concerning CRC cancer patients.
Data for multiple comparisons were subjected to one-way ANOVA and
Chi-square test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Overexpression of RPN2 is observed in
CRC specimens and cell lines and associated with poor survival of
CRC patients
In order to determine the biological role of RPN2 in
CRC tumor tissues, RT-PCR and western blot analysis were used to
detect the expression levels of RPN2 in tissues of 43 CRC patients
and their adjacent normal tissues. As displayed in Fig. 1A and B, the RPN2 expression levels
were higher in CRC tissues compared with the adjacent normal
tissues. Immunohistochemical staining results further demonstrated
that RPN2 was highly expressed in CRC tissues (Fig. 1C). Subsequently, a univariate
survival analysis indicated that the survival rate of patients with
high expression of RPN2 and the survival rate of patients with low
expression of RPN2 in 200 days was similar. As time passed, the
survival rate of patients with highly expressed RNP2 was reduced
(Fig. 1D). Subsequently, the mRNA
and protein expression levels of RPN2 in colon cancer cells,
SW1116, SW480, LoVo, SW403 and SW620, as well as normal colon cells
NCM460 were determined. We revealed that RPN2 was positively
identified in colon cancer cell lines, but weakly expressed in
normal colon cells (Fig. 1E-G).
Since the RPN2 expression levels in SW1116 and SW480 cells were
higher than that of LoVo, SW403 and SW620 cells, SW1116 and SW480
cells were used for the following experiment.
siRNA-mediated knockdown of RPN2 in
SW1116 and SW480 cells inhibits cell viability
In order to validate whether RPN2 functions in cell
proliferation, SW1116 and SW480 cells were used for detecting
changes in cell viability after RPN2 knockdown by siRNA
interference. As displayed in Fig.
2A-F, RT-PCR and western blot analysis at 48 h after
transfection demonstrated that RPN2 knockdown in SW1116 and SW480
cells significantly decreased the mRNA and protein levels of RPN2.
The cell proliferation was further assessed in SW1116 and SW480
cells using CCK-8 assay. After 48 h, cell proliferation was reduced
to 53% for SW1116 cell and to 45% for SW480 cells (Fig. 2G and H). These results indicated
that RPN2 siRNA interference significantly inhibited cell
proliferation.
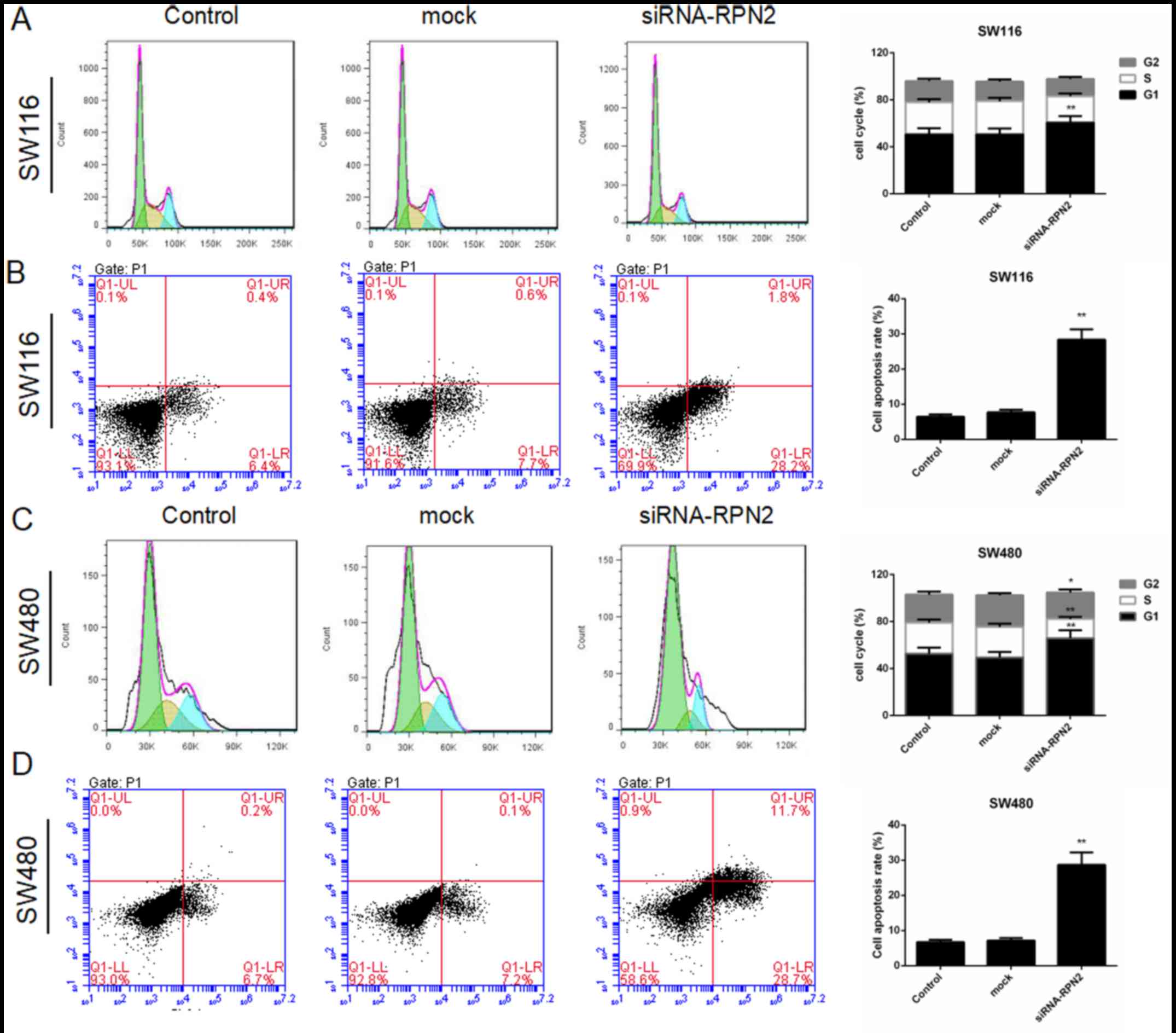
siRNA-mediated knockdown of RPN2
induces G1-phase cellcycle arrest and apoptosis in SW1116 and SW480
cells
In order to further confirm the effect of RPN2 on
cell proliferation, flow cytometry was used to detect the cell
cycle and apoptosis. After SW1116 and SW480 cell transfection with
RPN2 siRNA or control siRNA, there was no significant difference in
cell cycle and apoptosis between the control and the mock groups in
SW1116 and SW480 cells (Fig. 3).
The results demonstrated that the knockdown of RPN2 significantly
increased G1-phase cell population and increased the apoptosis rate
(27.9±1.9%) in SW1116 cells (Fig. 3A
and B). Then, the knockdown of RPN2 significantly increased
G1-phase cell population and decreased S-phase cell population in
SW480 cell (Fig. 3C). RPN2
downregulation in SW480 cells indicated a significant increase in
apoptosis rate by 28.8±3.2% (Fig.
3D). Collectively, these results demonstrated that the
knockdown of RPN2 inhibited cell proliferation and promoted
apoptosis of CRC cells.
siRNA-mediated knockdown of RPN2
suppresses the abilities of migration and invasion in SW1116 and
SW480 cells
The abilities of migration and invasion play a vital
role in tumor metastasis. Therefore, we further investigated the
effect of siRNA-mediated knockdown of RPN2 on the abilities of
migration and invasion in colon cancer cell lines (SW1116 and SW480
cells). Consistently, as displayed in Fig. 4, the downregulation RPN2 in SW1116
cells significantly reduced cell migration by 61.7% and the cell
invasion by 59.4% compared with control cells (Fig. 4A-C). In addition, we demonstrated
that RPN2 downregulation in SW480 cells significantly suppressed
cell migration by 49.5% and the cell invasion by 59.7% compared
with the control cells (Fig. 4D-F).
Collectively, these results demonstrated that the knockdown of RPN2
inhibited the migration and invasion abilities of CRC cells.
siRNA-mediated knockdown of RPN2
regulates the expression levels of caspase-3, cyclin D1,
E-cadherin, MMP-2 and TIMP-2 in SW1116 and SW480 cells
In order to further investigate the related
mechanisms involved in RPN2-mediated cell proliferation, migration
and invasion in CRC cells, the expression levels of caspase-3,
cyclin D1, E-cadherin, MMP-2 and TIMP-2 were detected by means of
RT-PCR and western blot analysis. As displayed in Fig. 5, the apoptosis-related protein
(caspase-3) expression was increased and the cell cycle-related
protein (cyclin D1) expression was decreased in SW1116 cells
(Fig. 5A and B). In addition,
migration- and invasion-related proteins including E-cadherin,
MMP-2 and TIMP-2 were detected by RT-PCR and western blot analysis.
The results indicated that E-cadherin and TIMP-2 expression levels
were increased and MMP-2 expression level was decreased after
SW1116 cell transfection with RPN2 siRNA (Fig. 5A and B). Concerning SW480 cells, the
knockdown of RPN2 increased the expression levels of caspase-3,
E-cadherin and TIMP-2 and decreased the expression levels of cyclin
D1 and MMP-2 after SW1116 cell transfection with RPN2 siRNA
(Fig. 5C and D).
 | Figure 5.Changes in the expression levels of
caspase-3, cyclin D1, E-cadherin, MMP-2 and TIMP-2 on RPN2 siRNA
transfection of SW1116 and SW480 cells. (A) The mRNA expression
levels of caspase-3, cyclin D1, E-cadherin, MMP-2 and TIMP-2 were
detected after RPN2 siRNA transfection of SW1116 cells for 48 h by
RT-PCR. (B) The protein expression levels of caspase-3, cyclin D1,
E-cadherin, MMP-2 and TIMP-2 were detected after RPN2 siRNA
transfection of SW1116 cells for 48 h by western blot analysis. (C)
The mRNA expression levels of caspase-3, cyclin D1, E-cadherin,
MMP-2 and TIMP-2 were detected after RPN2 siRNA transfection of
SW480 cells for 48 h by RT-PCR. (D) The protein expression levels
of caspase-3, cyclin D1, E-cadherin, MMP-2 and TIMP-2 were detected
after RPN2 siRNA transfection of SW480 cells for 48 h by western
blot analysis. GAPDH was also detected as the control of sample
loading. Data are expressed as the mean ± SD for three independent
experiments. **P<0.01 vs. control and mock. |
siRNA-mediated knockdown of RPN2
blocks the STAT3/JAK2 signaling in SW1116 and SW480 cells
After 48 h of RPN2 siRNA transfection, the
phosphorylation protein levels of STAT3 and JAK2 in SW1116 and
SW480 cells were determined by western blot analysis. The results
demonstrated that there was no change in the total protein
expression levels of JAK2 and STAT3 in SW1116 and SW480 cells
transfected with RNP2 siRNA. RPN2 downregulation suppressed the
phosphorylation levels of JAK2 and STAT3 (Fig. 6). Namely, the relative expression
levels of p-JAK2 and p-STAT3 were significantly decreased after
RPN2 siRNA interference (Fig. 6).
Total JAK2 (t-JAK2) and STAT3 (t-STAT3) protein expression level
had not any change. It was demonstrated that knockdown of RPN2 by
siRNA interference inhibited the activation of STAT3/JAK2 signaling
pathway in CRC cells.
Discussion
Tumor cells have the characteristics of malignant
proliferation, migration and invasion (24) which significantly influence the
development of cancer. Therefore, identifying specific genetic
markers associated with tumor growth and metastasis may be useful
to study cancer progression. According to the literature, an
intensive expression of RPN2 was found in CRC (16). But the related molecular mechanism
remains unknown. In the present study, we collected 43 samples of
CRC tissues and benign CRC tissues surgically removed in our
hospital from 2014 to 2017. Subsequently, the experiment results
indicated that RPN2 was highly expressed in CRC tissues and
associated with poor survival of CRC patients (Fig. 1A-D). It was identical with those
reported in literature. In order to further verify this result, the
expression levels of RPN2 in colon cancer cell lines (SW1116,
SW480, LoVo, SW403 and SW620) was determined by means of RT-PCR and
western blot analysis. We found that RPN2 was highly expressed in
CRC cell lines, especially SW1116 and SW480 cells (Fig. 1D and E). These results clearly
indicated that RPN2 is an important gene in the development of CRC.
Therefore, the function and related mechanisms of RPN2 should be
further investigated.
Concerning the role of RPN2 in CRC, it was reported
that protein glycation plays an important role in the regulation of
cell function by affecting protein stability, localization and
secretion (12). RPN2, as a kind of
glycoprotein, exists in eukaryotic organisms and was involved in
the development of breast, gastric and esophageal squamous cell
cancer (12,13,15).
Judging from the above-mentioned results, we hypothesized that RPN2
may regulate the proliferation, apoptosis, migration and invasion
in CRC. Subsequently, the knockdown of RPN2 by siRNA interference
was transfected into SW1116 and SW480 cells. In addition, our
results indicated that the siRNA interference vectors targeting the
RPN2 gene were successfully transfected into CRC cells with high
expression of RPN2 and inhibited cell viability (Fig. 2). Low expression of RPN2 suppressed
the process of cell cycle, restraining cells in the G1-phase,
inhibiting cell proliferation and promoting cell apoptosis
(Fig. 3). We further determined
that the migration and invasion abilities in cells transfected with
RPN2 siRNA were significantly reduced (Fig. 4). The above-mentioned observations
indicated that RPN2 regulated cell proliferation, apoptosis,
migration and invasion. Yet, the related mechanisms need to be
further researched.
Cell proliferation and apoptosis is a process of
dynamic equilibration (25).
Apoptosis suppression drive cells to escape death and leads to cell
abnormal proliferation, further causing the occurrence and
development of tumors (26,27). The members of the caspase family
were activated in the whole process of apoptosis. Caspase-3 is
downstream gene in the apoptosis signaling pathway. In general, the
activation of caspase-8 leads to the release of caspase-9 to induce
the activation of caspase-3, and ultimately to cause apoptosis
(28). It is the most common
apoptosis pathway in the human organism. Therefore, we detected the
expression of caspase-3 after cells were transfected with RPN2
siRNA and control siRNA. In addition, our results revealed that
caspase-3 expression was significantly increased in cells
transfected with RPN2 siRNA (Fig.
5). Cell cycle is a complex process. Once the body damage
induced by internal and external causes cell cycle arrest, and the
apoptosis could be induced when the damage could not be repaired on
time (29). Therefore, the
developments of tumor are linked with progressive dysregulation of
cell cycle and cell cycle is closely associated with cell apoptosis
(30). Cyclin D1 is a key protein
of cell proliferation signaling in G1-phase and overexpression of
cyclin D1 promotes the G1-S-phase transition of the cell cycle to
cause abnormal cell proliferation (31). It has been reported that cyclin D1
was highly expressed in CRC (32).
In our results, the expression levels of cyclin D1 were decreased
in cell transfection with RPN2 siRNA (Fig. 5). Migration and invasion are the
most important biological characteristics in tumors. Low expression
of E-cadherin would reduce the adhesion ability to further lead to
EMT (33). MMP-2 degraded cell
matrix to promote the ability to break through cell basement
membrane. TIMP-2, as natural inhibitors of MMP, can affect the
combination of MMP-2 and substrate (34). Therefore, E-cadherin, MMP-2 and
TIMP-2 were considered as marked proteins in the process of
migration and invasion in many types of cancer. Our results
indicated that E-cadherin and TIMP-2 expression was increased and
MMP-2 expression was decreased after cells transfected with siRNA
interference vectors targeting the RPN2 gene (Fig. 5). Collectively, RPN2 siRNA affected
the expression levels of caspase-3, cyclin D1, E-cadherin, MMP-2
and TIMP-2 to inhibit cell proliferation, induce cell apoptosis,
suppress cell migration and invasion.
JAK2/STAT3 signal pathway is a key signal
transduction pathway and plays an important role in maintaining a
constant internal environment (35). The activation of JAK affects cell
proliferation, apoptosis, migration and invasion. STAT3 is closely
associated with signaling pathway in many tumors (17,19).
In addition, the activation of STAT3 can upregulate anti-apoptosis
genes to inhibit cell apoptosis (20,21).
It was reported that some extracts or genes promoted cell apoptosis
and inhibited cell migration and invasion and caused cycle arrest
in kidney, breast, ovarian and colon cancer by inhibiting the
activation of STAT3 (16,36–39).
Therefore, the phosphorylation levels of JAK2 and STAT3 were
further determined in cells transfected with RPN2 siRNA. We
observed that the phosphorylation levels of JAK2 and STAT3 were
significantly decreased after the cells were transfected with RPN2
siRNA (Fig. 6). It was revealed
that RPN2 siRNA inhibited the activation of JAK2/STAT3. It was
further demonstrated that RPN2 may be a target gene of JAK2/STAT3
signal pathway.
According to the above-mentioned results,
downregulation of RPN2 inhibited cell proliferation, caused
cell-cycle arrest, promoted cell apoptosis and suppressed the
abilities of cell migration and invasion, regulated the expression
levels of apoptosis- and cycle-related genes (caspase-3 and cyclin
D1) and invasion- and metastatic-related genes (E-cadherin, MMP-2
and TIMP-2) and inhibited the activation of JAK2/STAT3 signaling
pathway. Therefore, the present study indicated that implicate
signaling through JAK2/STAT3 as a critical mechanism by which RPN2
siRNA may affect cell proliferation, apoptosis, migration and
invasion in CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by a fund of Shanxi
Province Science and Technology Development Research project (no.
2010R034-2).
Availability of data and materials
The datasets used and/or analyzed during the current
study areavailable from the corresponding author on reasonable
request.
Authors' contributions
CB and BJ designed and performed the experiments; BJ
contributed to the data analysis; CB enrolled patients and measured
the RNA levels in the clinical samples; CB initiated the work and
wrote the manuscript. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The use of human tissues was approved by the Ethics
Committee of Huai'an First People's Hospital.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Niknami Z, Eslamifar A, Emamirazavi A,
Ebrahimi A and Shirkoohi R: The association of vimentin and
fibronectin gene expression with epithelial-mesenchymal transition
and tumor malignancy in colorectal carcinoma. EXCLI J.
16:1009–1017. 2017.PubMed/NCBI
|
|
2
|
Xu RH, Shen L, Li J, Xu JM, Bi F, Ba Y,
Bai L, Shu YQ, Liu TS, Li YH, et al: Expert consensus on
maintenance treatment for metastatic colorectal cancer in China.
Chin J Cancer. 35:132016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shin CM, Han K, Lee DH, Choi YJ, Kim N,
Park YS and Yoon H: Association among obesity, metabolic health,
and the risk for colorectal cancer in the general population in
Korea using the National Health Insurance Service-National Sample
Cohort. Dis Colon Rectum. 60:1192–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lopes-Ramos CM, Barros BP, Koyama FC,
Carpinetti PA, Pezuk J, Doimo NT, Habr-Gama A, Perez RO and
Parmigiani RB: E2F1 somatic mutation within miRNA target site
impairs gene regulation in colorectal cancer. PLoS One.
12:e01811532017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jayasekara H, English DR, Haydon A, Hodge
AM, Lynch BM, Rosty C, Williamson EJ, Clendenning M, Southey MC1,
Jenkins MA, et al: Associations of alcohol intake, smoking,
physical activity and obesity with survival following colorectal
cancer diagnosis by stage, anatomic site and tumor molecular
subtype. Int J Cancer. 142:238–250. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Altobelli E, Latella G, Morroni M, Licini
C, Tossetta G, Mazzucchelli R, Profeta VF, Coletti G, Leocata P,
Castellucci M, et al: Low HtrA1 expression in patients with long
standing ulcerative colitis and colorectal cancer. Oncol Rep.
38:418–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nandra R, Parry M, Forsberg J and Grimer
R: Can a bayesian belief network be used to estimate 1-year
survival in patients with bone sarcomas? Clin Orthop Relat Res.
475:1681–1689. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yogarajah M and Tefferi A: Leukemic
transformation in myeloproliferative neoplasms: A literature review
on risk, characteristics, and outcome. Mayo Clin Proc.
92:1118–1128. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fujieda Y, Amengual O, Matsumoto M, Kuroki
K, Takahashi H, Kono M, Kurita T, Otomo K, Kato M, Oku K, et al:
Ribophorin II is involved in the tissue factor expression mediated
by phosphatidylserine-dependent antiprothrombin antibody on
monocytes. Rheumatology (Oxford). 55:1117–1126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ono M, Tsuda H, Kobayashi T, Takeshita F,
Takahashi RU, Tamura K, Akashi-Tanaka S, Moriya T, Yamasaki T,
Kinoshita T, et al: The expression and clinical significance of
ribophorin II (RPN2) in human breast cancer. Pathol Int.
65:301–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Honma K, Iwao-Koizumi K, Takeshita F,
Yamamoto Y, Yoshida T, Nishio K, Nagahara S, Kato K and Ochiya T:
RPN2 gene confers docetaxel resistance in breast cancer. Nat Med.
14:939–948. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tominaga N, Hagiwara K, Kosaka N, Honma K,
Nakagama H and Ochiya T: RPN2-mediated glycosylation of tetraspanin
CD63 regulates breast cancer cell malignancy. Mol Cancer.
13:1342014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kurashige J, Watanabe M, Iwatsuki M,
Kinoshita K, Saito S, Nagai Y, Ishimoto T, Baba Y, Mimori K and
Baba H: RPN2 expression predicts response to docetaxel in
oesophageal squamous cell carcinoma. Br J Cancer. 107:1233–1238.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fujiwara T, Takahashi RU, Kosaka N, Nezu
Y, Kawai A, Ozaki T and Ochiya T: RPN2 gene confers osteosarcoma
cell malignant phenotypes and determines clinical prognosis. Mol
Ther Nucleic Acids. 3:e1892014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan TM, Liang RY, Chueh PJ and Chuang SM:
Role of ribophorin II in the response to anticancer drugs in
gastric cancer cell lines. Oncol Lett. 9:1861–1868. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loo LW, Tiirikainen M, Cheng I, Lum-Jones
A, Seifried A, Church JM, Gryfe R, Weisenberger DJ, Lindor NM,
Gallinger S, et al: Integrated analysis of genome-wide copy number
alterations and gene expression in microsatellite stable, CpG
island methylator phenotype-negative colon cancer. Genes
Chromosomes Cancer. 52:450–466. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ju KD, Lim JW and Kim H: Peroxisome
proliferator-activated receptor-gamma inhibits the activation of
STAT3 in cerulein-stimulated pancreatic acinar cells. J Cancer
Prev. 22:189–194. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim EY, Jung JY, Kim A, Chang YS and Kim
SK: ABT-737 synergizes with cisplatin bypassing aberration of
apoptotic pathway in non-small cell lung cancer. Neoplasia.
19:354–363. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang SN, Ruan HZ, Chen MY, Zhou G and
Qian ZM: Aspirin increases ferroportin 1 expression by inhibiting
hepcidin via the JAK/STAT3 pathway in interleukin 6-treated PC-12
cells. Neurosci Lett. 662:1–5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y, Zhou P, Chen S, Hu C, Bai Q, Wu H,
Chen Y, Zhou P, Zeng X, Liu Z, et al: The JAK/STAT3 signaling
pathway mediates inhibition of host cell apoptosis by Chlamydia
psittaci infection. Pathog Dis. Aug 3–2017.(Epub ahead of
print). doi: 10.1093/femsle/ftx088. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Xu J and Xing G: Lycorine inhibits
the growth and metastasis of breast cancer through the blockage of
STAT3 signaling pathway. Acta Biochim Biophys Sin (Shanghai).
49:771–779. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Berthenet K, Bokhari A, Lagrange A,
Marcion G, Boudesco C, Causse S, De Thonel A, Svrcek M, Goloudina
AR, Dumont S, et al: HSP110 promotes colorectal cancer growth
through STAT3 activation. Oncogene. 36:2328–2336. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan Y, Su C, Hang M, Huang H, Zhao Y, Shao
X and Bu X: Recombinant Newcastle disease virus rL-RVG enhances the
apoptosis and inhibits the migration of A549 lung adenocarcinoma
cells via regulating alpha 7 nicotinic acetylcholine receptors in
vitro. Virol J. 14:1902017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo JM, Cao FL, Meng C, Lin LJ, Ma SQ,
Peng SH, Gao HL, Javidiparsijani S, Wang GR, Zhang ML, et al:
Clinicopathological and molecular characteristics of synchronous
gastric adenocarcinoma and gastrointestinal stromal tumors. Sci
Rep. 7:128902017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Naine SJ, Devi CS, Mohanasrinivasan V,
Doss CG and Kumar DT: Binding and molecular dynamic studies of
sesquiterpenes
(2R-acetoxymethyl-1,3,3-trimethyl-4t-(3-methyl-2-buten-1-yl)-1t-cyclohexanol)
derived from marine Streptomyces sp. VITJS8 as potential
anticancer agent. Appl Microbiol Biotechnol. 100:2869–2882. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li T, Ding ZL, Zheng YL and Wang W:
MiR-484 promotes non-small-cell lung cancer (NSCLC) progression
through inhibiting Apaf-1 associated with the suppression of
apoptosis. Biomed Pharmacother. 96:153–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Taira J, Sonamoto M and Uehara M: Dual
biological functions of a cytoprotective effect and apoptosis
induction by bioavailable marine carotenoid fucoxanthinol through
modulation of the Nrf2 activation in RAW264.7 macrophage cells. Mar
Drugs. 15:E3052017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao Q, Li M, Chen M, Zhou L, Zhao L, Hu
R, Yan R and Dai K: Lovastatin induces platelet apoptosis. Environ
Toxicol Pharmacol. 42:69–75. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hazama Y, Moriya T, Sugihara M, Sano R,
Shiota M, Nakamura T and Shimoya K: The analysis of cell
cycle-related proteins in ovarian clear cell carcinoma versus
high-grade serous carcinoma. Int J Gynecol Pathol. Oct
10–2017.(Epub ahead of print). doi: 10.1097/PGP.0000000000000461.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang X, Li Y, Lin M, Jin J and Huang Z:
Rhodium (II) complex with 2-benzoylpyridine, a novel potential
chemotherapeutic drug, induces cell cycle arrest and apoptosis in
HepG2 cells. Biometals. 30:903–915. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He S, Liao B, Deng Y, Su C, Tuo J, Liu J,
Yao S and Xu L: MiR-216b inhibits cell proliferation by targeting
FOXM1 in cervical cancer cells and is associated with better
prognosis. BMC Cancer. 17:6732017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu L, Zhang C, Li X, Sun W, Qin S, Qin L
and Wang X: miR-223 promotes colon cancer by directly targeting
p120 catenin. Oncotarget. 8:63764–63779. 2017.PubMed/NCBI
|
|
33
|
Liu H, Zhen Q and Fan Y: LncRNA GHET1
promotes esophageal squamous cell carcinoma cells proliferation and
invasion via induction of EMT. Int J Biol Markers. 32:e403–e408.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pisamai S, Rungsipipat A, Kunnasut N and
Suriyaphol G: Immunohistochemical expression profiles of cell
adhesion molecules, matrix metalloproteinases and their tissue
inhibitors in central and peripheral neoplastic foci of feline
mammary carcinoma. J Comp Pathol. 157:150–162. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ohba S, Lanigan TM and Roessler BJ: Leptin
receptor JAK2/STAT3 signaling modulates expression of Frizzled
receptors in articular chondrocytes. Osteoarthritis Cartilage.
18:1620–1629. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li W, Zhang H, Nie M, Tian Y, Chen X, Chen
C, Chen H and Liu R: Ursolic acid derivative FZU-03,010 inhibits
STAT3 and induces cell cycle arrest and apoptosis in renal and
breast cancer cells. Acta Biochim Biophys Sin (Shanghai).
49:367–373. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou W, Bi X, Gao G and Sun L: miRNA-133b
and miRNA-135a induce apoptosis via the JAK2/STAT3 signaling
pathway in human renal carcinoma cells. Biomed Pharmacother.
84:722–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu L and Ding X: Molecular design of
Stat3-derived peptide selectivity between BET proteins Brd2 and
Brd4 in ovarian cancer. J Mol Recognit. Oct 6–2017.(Epub ahead of
print). doi: 10.1002/jmr.2679.
|
|
39
|
Liu X, Wei W, Li X, Shen P, Ju D, Wang Z,
Zhang R, Yang F, Chen C, Cao K, et al: BMI1 and MEL18 promote
colitis-associated cancer in mice via REG3B and STAT3.
Gastroenterology. 153:1607–1620. 2017. View Article : Google Scholar : PubMed/NCBI
|