Introduction
Lung cancer is one of the principal causes of
cancer-associated mortality, and contributes to the economic burden
worldwide. In total, ~85% of confirmed lung cancer cases are
classified as non-small cell lung cancer (NSCLC) (1–3).
Traditionally, clinical methods, including surgery, chemotherapy or
a combination are the primary treatment approaches for lung cancer.
Recently, molecular-targeted drugs (4), anti-angiogenic therapy (5) and cancer immunotherapy (6) have been widely applied in patients
with lung cancer. Despite advances in early diagnosis and standard
treatment, the overall 5-year survival rate for NSCLC is <15%
(7), and there is an urgent
requirement for the development of novel therapeutic approaches for
NSCLC.
Phosphatase and tensin homolog (PTEN), located on
chromosome 10q23, is well known as a tumor suppressor, and has
lipid and protein phosphatase activities. It is involved in various
cellular processes and signal transduction pathways in a complex
network system (8–10). Emerging evidence has demonstrated
that PTEN serves an important role in tumorigenesis and tumor
suppression in multiple tumor types, including thyroid cancer
(11), osteosarcoma (12), gastric cancer (13) and myeloma (14). The function of PTEN may be regulated
by genetic mutations, transcriptional regulation and
post-translational modifications (8,10).
Loss of PTEN or a decrease in PTEN expression levels and activity
has been associated with poorer overall survival among patients
with NSCLC. Therefore, regulation of PTEN has emerged as a
promising research topic for cancer therapy (15).
In recent years, natural agents have attracted
increasing attention due to the richness of the resource, easy
accessibility and cost effectiveness for clinical application. As
previously demonstrated, treatment with Osthole and Morinda
citrifolia edible leaf extract (16,17),
regulated PTEN expression, inducing cancer growth and metastasis.
Pectolinarigenin (Pec; C17H14O6;
molecular weight: 314.28; melting point: 204–205°C; storage
conditions: 4°C refrigerated, sealed and protected from light) is a
flavonoid compound widely distributed in a number of medicinal
plants, including Cirsium japonlcum, Eupatorium odoratum and
Trollius chinensis. Under the appearance of a yellow
crystal, it is soluble in dimethyl sulfoxide and hot methanol, and
insoluble in petroleum ether, chlorine and other solvents. As
documented in previous studies, Pec has demonstrated effective
antitumor activities in vivo and in vitro. Cheng
et al (18) observed that
Pec may inhibit cell viability and migration of nasopharyngeal
carcinoma cells, and induce mitochondrial-associated apoptosis
through the accumulation of caspase-3 and caspase-9 in cells. Zhang
et al (19) demonstrated
that Pec was able to disturb signal transducer and activator of
transcription 3 (STAT3) signaling and decrease STAT3 downstream
proteins, including cyclin D1, B-cell lymphoma 2 (BCL-2) B-cell
lymphoma extra-large (BCL-xL), Myeloid cell leukemia 1 (MCL-1),
contributing to the suppression of cell proliferation and apoptosis
in osteosarcoma cells. Additionally, Pec was able to inhibit cell
migration and invasion, and preserved the epithelial-mesenchymal
transition (EMT) phenotype. As uncontrolled cell proliferation and
metastasis are considered hallmarks of malignant tumors, inhibition
of associated signaling pathways is one important aspect of cancer
treatment.
It has been demonstrated that Pec may inhibit growth
and metastasis of nasopharyngeal carcinoma cells and osteosarcoma
cells (18,19); however, the effect of Pec on NSCLC
and its underlying mechanisms have not been reported. In the
present study, the potential effects of Pec on human NSCLCs cells
were investigated to clarify the possible underlying mechanisms. As
a result, it was revealed that Pec may significantly inhibit cell
proliferation, migration, invasion, EMT, and induce apoptosis in
vitro by promoting the expression of PTEN.
Materials and methods
Cell lines and reagents
Human NSCLC cell lines A549 and Calu-3 were
purchased from the Shanghai Institute of Biochemistry and Cell
Biology (Shanghai, China). The two cell lines were cultured in
RPMI-1640 medium with 10% fetal bovine serum (FBS), 100 U/ml
penicillin and 100 µg/ml streptomycin (all from Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) in a 5% CO2
incubator at 37°C. Pec reagent was purchased from Abmole Bioscience
Inc. (Houston, TX, USA). Antibodies against Bax (cat. no. 2744),
BCL-xL (cat. no. 2762), PTEN (cat. no. 9188),
phospho-phosphoinositide 3-kinase (p-PI3K; cat. no. 4228),
phospho-protein kinase B (p-AKT; cat. no. 4060), cellular tumor
antigen p53 (p53; cat. no. 9282), Lamin B1 (cat. no. 13435) and
GAPDH (cat. no. 5174) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Fluorescence-tagged secondary
antibodies, IRDye® 680RD goat anti-rabbit IgG (cat. no.
925-68071) and IRDye® 680RD goat anti-mouse IgG (cat.
no. 925-68070) were acquired from LI-COR Biosciences, Inc.
(Lincoln, NE, USA). PTEN inhibitor SF1670 (cat. no. ab141303) and
AKT activator SC79 (cat. no. ab146428) were purchased from Abcam
(Cambridge, UK). SF1670 and/or SC79 (10 µM) were applied to the
culture medium for 1 h at room temperature in the treated group.
NE-PER™ Nuclear and Cytoplasmic Extraction reagents (cat. no.
78835) were purchased from Thermo Fisher Scientific, Inc. The
primary and secondary antibodies were respectively diluted to
1:1,000 and 1:4,000 in 5% BAS (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany).
Cell viability assay
Cells were grown in a 96-well plate overnight at a
density of 4×103 cells/well, and subsequently treated
with different concentrations of Pec for 24, 48 and 72 h. Following
incubation, 10 µl Cell Counting Kit-8 (CCK-8) reagent (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) was added to each
well for 1 h. The optical density of each sample was measured
spectrophotometrically at 450 nm with an automatic microplate
analyzer (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Colony formation assay
Cells were seeded in a 6-well plate at 400
cells/well. After 24 h, the cells were treated with Pec at various
concentrations (0, 10 and 25 µM) and incubated at 37°C with 5%
CO2. The cells were consistently cultured for 10 days
and the culture medium was changed every two days. The cells were
washed twice with PBS and subsequently fixed with 4%
paraformaldehyde for 30 min at room temperature. A crystal violet
stain was used to observe colony formation.
Cell apoptosis assay
A cell apoptosis assay was conducted by flow
cytometry with a Fluorescein Isothiocyanate (FITC) Annexin V
Apoptosis Detection kit (BD Biosciences, Franklin Lakes, NJ, USA),
according to the manufacturer's protocol. After treatment with Pec
for 48 h, the cells were harvested and resuspended in binding
buffer at a concentration of 1×106 cells/ml.
Subsequently, 5 µl FITC Annexin V and 5 µl Propidium Iodide
Staining Solution were added to a 200-µl cell suspension, and
incubated for 30 min at room temperature in the dark. Cell
apoptosis was detected using BD FACSVerse (BD Biosciences) and the
apoptosis rate of cancer cells was analyzed using FlowJo software
version 7.0 (FlowJo LLC, Ashland, OR, USA).
Western blot analysis
Cells were harvested and washed twice with PBS,
subsequently lysed with radioimmunoprecipitation buffer and 1X
protease inhibitors (100:1) at 4°C for 30 min. Subsequent to
centrifuging at 12,000 × g for 10 min at 4°C, the supernatant was
collected and the protein concentration in the supernatant
fractions was quantified using a Bicinchoninic Acid Protein Assay
kit (Thermo Fisher Scientific, Inc.). The remaining supernatant was
mixed with SDS-PAGE loading buffer and boiled at 100°C for 10 min.
The same amount of protein (20 µg) was separated using 12% SDS-PAGE
and subsequently transferred to polyvinylidene difluoride membranes
(EMD Millipore, Billerica, MA, USA). Blocking buffer was used to
block the non-specific binding sites in the membranes for 15 min at
room temperature. Primary antibodies were incubated with the
membranes overnight at 4°C. The membranes were washed three times
with PBS and incubated with fluorescence-tagged secondary
antibodies for 1 h. Images of the membranes were captured with the
Odyssey CLx Infrared Imaging System (LI-COR Biosciences, Lincoln,
NE, USA). GAPDH and Lamin B1 protein intensity was used as internal
controls for cytoplasm and nucleus fractions, respectively.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA of the NSCLC cells treated with or without
Pec were collected and extracted with RNeasy Mini RNA kit (Qiagen
GmbH, Hilden, Germany) according to the manufacturer's protocol.
The concentrations of total RNA were detected with NanoDrop 2000
and reversed transcribed into cDNA (Takara Biotechnology Co., Ltd.,
Dalian, China). The SYBR kit (Qiagen GmbH) was used for the
detection of E-cadherin, vimentin and GAPDH. The primers sequences
were as follows: E-cadherin forward, 5′-CGAGAGCTACACGTTCACGG-3′ and
reverse, 5′-GGGTGTCGAGGGAAAAATAGG-3′; vimentin forward,
5′-GACGCCATCAACACCGAGTT-3′ and reverse,
5′-CTTTGTCGTTGGTTAGCTGGT-3′; GAPDH forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. The densitometry of relative mRNA
expression was performed using the calculation of 2−ΔΔCq
(20) and GAPDH was used as an
internal control.
Wound scratch assay
Cancer cell migration was assessed using a cell
wound scratch assay. NSCLC cells were seeded in a 6-well plate and
incubated overnight to form a confluent cell monolayer, and a
scratch was made with a 100-µl sterile pipette tip. The cancer
cells were subsequently treated with or without Pec for 24 h and
images of the scratches were obtained with an inverted microscope
(Olympus Corp., Tokyo, Japan).
Transwell assay
For the invasion assay, the upper chambers were
pre-coated with Matrigel (Invitrogen; Thermo Fisher Scientific,
Inc.), the NSCLC cancer cells (1×105 cells/well) were
plated on the upper chamber with Pec (25 µM) or alone; whereas, the
lower chamber contained RPMI-1640 medium with 10% FBS and 1%
penicillin/streptomycin. After 24 h of incubation, the upper
chambers were swabbed out, and the lower chambers were fixed with
paraformaldehyde and stained with crystal violet. Images of the
invaded cells were captured with a Leica fluorescence microscope
(magnification, ×100; Olympus Corp.).
Statistical analysis
All data were analyzed using SPSS 19.0 software (IBM
Corp., Armonk, NY, USA) and comparisons between groups were
determined by Student's t-test. The results are presented as the
mean ± standard deviation. P<0.05 was considered to indicate a
statistically significant difference. All experiments were repeated
at least three times independently.
Results
Pec inhibits cell proliferation in
A549 and Calu-3 cells
It was reported that Pec exhibited an inhibitory
effect on nasopharynx carcinoma (18); thus, in order to investigate whether
Pec had a similar effect on NSCLC cells, a cell proliferation assay
was conducted using a CCK-8 assay. The results revealed that the
cell viability of A549 and Calu-3 cells was decreased following
administration of Pec. The CCK-8 assay demonstrated that Pec was
able to significantly inhibit the cell proliferation in dose-
(Fig. 1A) and time-dependent
(Fig. 1B and C) manners, with half
maximal inhibitory concentration values of 21.49 and 22.63 µM,
respectively. In addition, the colony formation assay demonstrated
that the growth rate was markedly decreased in the two cell lines
treated with Pec (Fig. 1D). These
results indicated that Pec exhibited an inhibitory effect on the
proliferation of NSCLC cells.
 | Figure 1.Pec inhibits proliferation in lung
adenocarcinoma cells. (A) A549 and Calu-3 cells were incubated with
Pec at different concentrations (0, 0.1, 1, 5, 10, 25, 50 and 100
µM) for 72 h. (B and C) A549 and Calu-3 cells were exposed to Pec
(10 and 25 µM) for different time-points (24, 48 and 72 h). Cell
viability and IC50 values were measured using Cell
Counting Kit-8 assays. (D) Clonogenic survival assay of A549 and
Calu-3 cells treated with Pec (10 and 25 µM). Each experiment was
performed three times. The data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01. Pec, pectolinarigenin;
IC50, half maximal inhibitory concentration. |
Pec induces the apoptosis of human
NSCLC cells
To determine the effect of Pec on NSCLC cell
apoptosis, flow cytometry was conducted to detect the apoptosis of
cells treated with Pec. As presented in Fig. 2, the apoptosis rate of the A549 and
Calu-3 cell lines increased gradually with the administration of
different concentrations of Pec (0, 5, 10, 25, 50 and 100 µM) for
48 h, particularly the A549 cell line.
 | Figure 2.Pec induces apoptosis in lung
adenocarcinoma cells. (A and B) A549 cells were treated with Pec at
different concentrations (0, 5, 10, 25, 50 and 100 µM) for 48 h.
Flow cytometric analysis demonstrated that Pec induced increased
apoptosis with increased doses of Pec. Each experiment was
performed three times. The data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01. Pec, pectolinarigenin.
Pec induces apoptosis in lung adenocarcinoma cells. (C and D)
Calu-3 cells were treated with Pec at different concentrations (0,
5, 10, 25, 50 and 100 µM) for 48 h. Flow cytometric analysis
demonstrated that Pec induced increased apoptosis with increased
doses of Pec. Each experiment was performed three times. The data
are presented as the mean ± standard deviation. *P<0.05,
**P<0.01. Pec, pectolinarigenin. |
Furthermore, western blot analysis demonstrated that
the pro-apoptotic Bax protein expression level was upregulated;
whereas, the anti-apoptotic BCL-xL protein expression level was
downregulated in the A549 cell line (Fig. 3A), and similar results were
additionally observed in the Calu-3 cell line (Fig. 3B). Therefore, it was hypothesized
that Pec exhibited an antitumor effect by inducing NSCLC cancer
cell apoptosis and apoptosis-associated protein expression
levels.
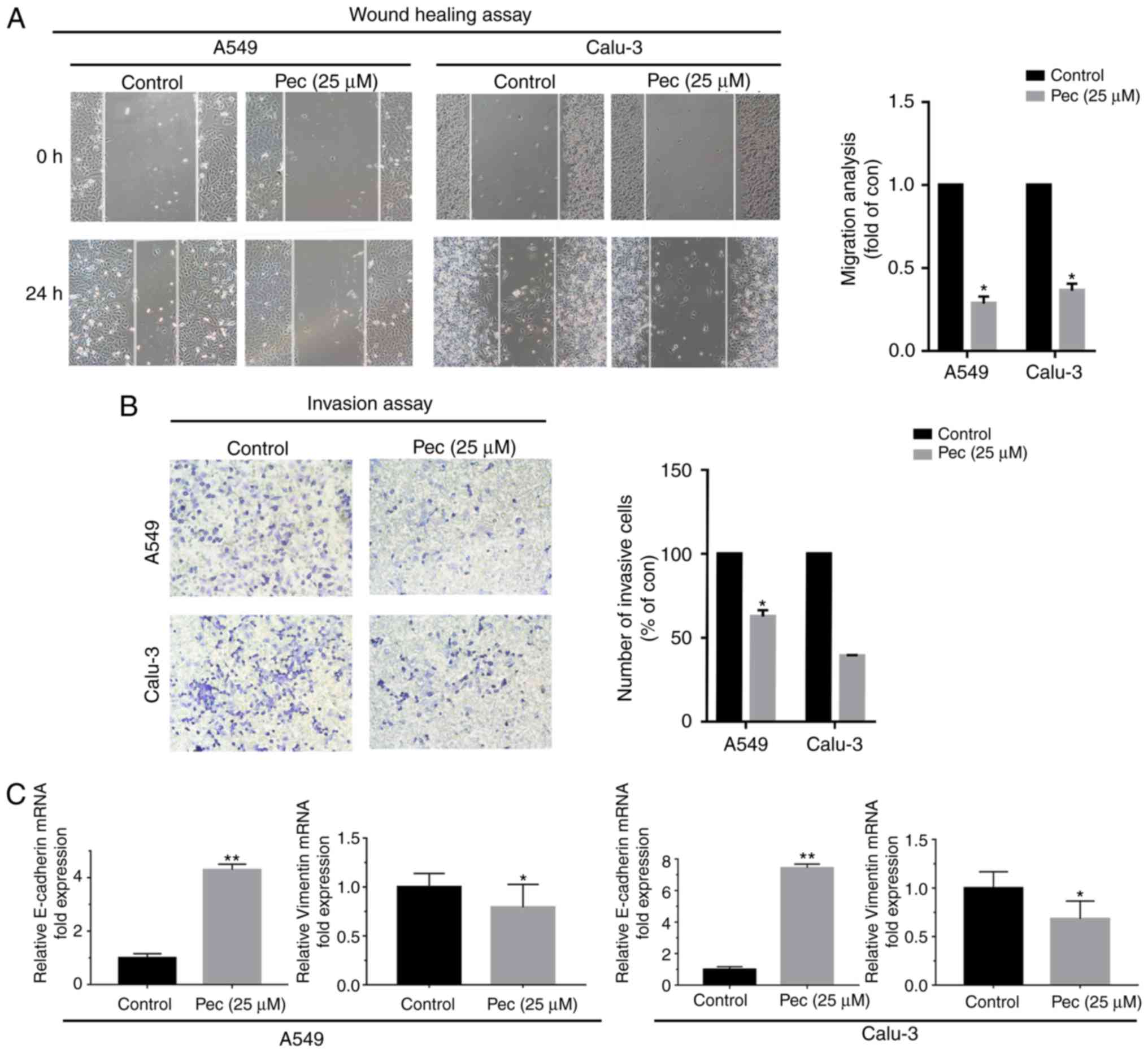
Pec decreases the migration, invasion
and EMT of NSCLC cells
To examine whether Pec was able to inhibit the
migration, invasion and EMT of NSCLC cells, a cell wound scratch
assay was performed and the two cell lines were treated without or
with Pec (25 µM). After 24 h of incubation, Pec significantly
inhibited the migration of NSCLC cells compared with the control
groups (Fig. 4A). A Transwell assay
was conducted to detect the invasion of NSCLC cells; the results
demonstrated that Pec suppressed the invasion of A549 and Calu-3
cells (Fig. 4B). Furthermore, the
expression of epithelial marker E-cadherin and mesenchymal marker
vimentin was detected by RT-qPCR and western blotting. The EMT
process is considered the initial step for cancer metastasis; Pec
markedly upregulated the expression of E-cadherin and downregulated
the vimentin expression at the mRNA (Fig. 4C) and protein expression levels
(Fig. 4D). Collectively, treatment
with Pec suppressed the metastasis and EMT of NSCLC cells.
Effects of Pec on the PTEN/PI3K/AKT
pathway in A549 and Calu-3 cells
A previous study suggested that the PTEN/PI3K/AKT
pathway is involved in cancer progression (21–25),
thus, the effect of Pec on the PTEN/PI3K/AKT pathway in A549 and
Calu-3 cells was evaluated by western blotting to examine the
underlying mechanism of Pec-mediated inhibition of cell
proliferation and metastasis. In the present study, the results of
western blotting demonstrated that the expression of phospho-PI3K
and phospho-AKT was significantly decreased; whereas, the PTEN
protein expression level was increased with increasing
concentrations of Pec (Fig. 5).
Previous studies suggested a complicated association and
interaction between PTEN and p53 (26,27).
To confirm whether p53 is involved in the pathway mediated by PTEN,
cells were pre-treated with Pec and/or an inhibitor of PTEN
(SF1670), and the protein expression level of PTEN and p53 was
detected. The present results indicated that the altered expression
of PTEN was associated with p53 expression. Treatment with SF1670
alone inhibited PTEN and p53 expression; whereas, combined
treatment of Pec and SF1670 in NSCLC cells abolished the
SF1670-induced effect on PTEN and p53 (Fig. 6A). Furthermore, activation of AKT by
SC79, a novel AKT activator, decreased the expression of p53, which
indicated that p53 was negatively regulated by PTEN/PI3K/AKT
(Fig 6B). Generally, the present
results revealed that Pec exerted antitumor activity through the
PTEN/PI3K/AKT signaling pathway and p53 served an important role in
the signaling pathway mediated by PTEN. The specific schematic
diagram of PTEN/PI3K/AKT and the crosstalk between p53 and PTEN is
presented in Fig. 7. Additionally,
it was identified that Pec was able to promote the translocation of
PTEN from the cytoplasm to the nucleus to exert antitumor activity
(data not shown), which is consistent with previous studies
(28).
Discussion
Lung cancer, particularly NSCLC, is responsible for
the majority of cancer-associated mortalities; there were 1.69
million mortalities of patients with lung cancer reported in 2015
worldwide (3,7). Although the development of novel
therapies has greatly ameliorated the conditions of patients with
lung cancer, the 5-year survival rate has not improved much
primarily due to resistance to antitumor agents and toxic
side-effects (29). In clinical
treatments, drug resistance remains one of the principal issues in
developing a successful treatment for cancer. It is important to
develop more effective drugs against lung cancer with fewer
side-effects.
In recent years, there have been an increasing
number of previous studies investigating the antitumor activity of
medicinal plant extracts, which have successfully demonstrated
their potential use in treatments. Previous modern pharmacological
studies demonstrated the excellent antitumor effects of various
medicinal plant extracts, including oxymatrine (30) and deguelin (31). Pec is a natural extract from a
diverse range of herbal medicine and was previously demonstrated to
have an antitumor effect on nasopharyngeal carcinoma (18) and osteosarcoma (19). However, there has been no study, to
the best of our knowledge, on the biological activity of Pec in
human NSCLC cells. With regard to the identification of the
anticancer effects of Pec on NSCLC cells, in the present study,
experiments were conducted to demonstrate the inhibitory effect of
Pec on NSCLC cell proliferation and metastasis.
In the present study, it was observed that Pec was
able to significantly decrease the cell viabilities of A549 and
Calu-3 cancer cells in dose- and time-dependent manners with the
administration of Pec. Pec-mediated cell apoptosis was augmented
with increasing Pec concentrations, demonstrating a dose-dependent
effect. Furthermore, it was observed that Pec significantly
inhibited the migration and invasion of cancer cells, and
upregulated E-cadherin expression; whereas it downregulated
vimentin expression. Collectively, these results indicated that Pec
exerted an effective anticancer activity against lung cancer.
Furthermore, an aim of the present study was to
elucidate the underlying mechanisms involved in Pec-induced
antitumor progression. It was previously observed that PTEN, a
tumor suppressor phosphatase, may negatively regulate the
activation of the PI3K/AKT signaling pathway (22). The PI3K/AKT pathway is essential for
maintaining cell growth, survival, death and metabolism, and is
commonly activated in cancer initiation and progression (32). Activation of the PTEN/PI3K/AKT
pathway may be a therapeutic molecular target for lung cancer.
(26). In the past decade, it was
demonstrated that various natural compounds positively regulated
PTEN gene expression, consequently exerting an inhibitory effect
against tumors. Similarly, in the present study, it was
demonstrated that Pec activated PTEN and downregulated the activity
of PI3K/AKT, which indicated that the PTEN/PI3K/AKT signaling
pathway was involved in the Pec-induced antitumor effect. It was
additionally revealed that Pec was able to promote the nuclear
translocation of PTEN. Furthermore, p53 is a well-documented tumor
suppressor protein and is involved in apoptosis, autophagy, cell
cycle regulation, and DNA replication and repair. Previous studies
have indicated that PTEN could regulate p53 protein stability by
antagonizing the PI3K/AKT-mediated MDM2 (murine double minute
2)-p53 pathway or by interacting with p53 directly. The present
results indicated that p53, as a complementary signal, was involved
in the PTEN-mediated PI3K/AKT pathway. However, cancer cell growth
and metastasis is a complicated process in which multiple signaling
pathways are involved (33–35). Therefore, whether other signaling
pathways are involved in the Pec-induced anticancer effect and the
PTEN-mediated p53 signaling pathway through a phosphatase-dependent
manner or phosphatase-independent manner requires further
investigation.
In conclusion, the present study demonstrated that
Pec exhibited an antitumor effect on human NSCLC cancer cell lines,
which was able to inhibit NSCLC cell proliferation, metastasis and
EMT, and promote apoptosis through the PTEN/PI3K/AKT signaling
pathway. Pec has potential for further development and may provide
additional strategies for mono-therapy or combination treatments
for NSCLC.
Acknowledgements
The authors thank Mr. D. Zhao for the technical
support.
Funding
The present study was supported by grants from the
Public Welfare Project of Zhejiang Province (grant no. 2012C23074)
and the Natural Science Found of Zhejiang Province (grant no.
Y2110167).
Availability of data and materials
The datasets used during the current study are
available from the corresponding author upon reasonable
request.
Authors' contributions
FX and HP designed and conceived the study. FX and
XG performed the experiments, analyzed the data and wrote the
manuscript. HP reviewed and edited the manuscript, as well as
provided funds. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EMT
|
epithelial-mesenchymal transition
|
|
NSCLC
|
non-small cell lung cancer
|
|
Pec
|
pectolinarigenin
|
|
PTEN
|
phosphatase and tensin homolog
|
References
|
1
|
Smith RA, Andrews KS, Brooks D, Fedewa SA,
Manassaram-Baptiste D, Saslow D, Brawley OW and Wender RC: Cancer
screening in the United States, 2017: A review of current american
cancer society guidelines and current issues in cancer screening.
CA Cancer J Clin. 67:100–121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
By the numbers: Cancer mortality,
2011–2015. Cancer Discov. 8:9072018.
|
|
3
|
de Groot PM, Wu CC, Carter BW and Munden
RF: The epidemiology of lung cancer. Transl Lung Cancer Res.
7:220–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rybarczyk-Kasiuchnicz A and Ramlau R:
Current views on molecularly targeted therapy for lung cancer-a
review of literature from the last five years. Kardiochir
Torakochirurgia Pol. 15:119–124. 2018.PubMed/NCBI
|
|
5
|
Varol M, Koparal AT, Benkli K and
Bostancioglu RB: Anti-lung cancer and anti-angiogenic activities of
new designed boronated phenylalanine metal complexes. Curr Drug
Deliv. Jul 27–2018.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miura Y and Sunaga N: Role of
immunotherapy for oncogene-driven non-small cell lung cancer.
Cancers (Basel). 10:E2452018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CY, Chen J, He L and Stiles BL: PTEN:
Tumor suppressor and metabolic regulator. Front Endocrinol
(Lausanne). 9:3382010. View Article : Google Scholar
|
|
9
|
Jamaspishvili T, Berman DM, Ross AE, Scher
HI, De Marzo AM, Squire JA and Lotan TL: Clinical implications of
PTEN loss in prostate cancer. Nat Rev Urol. 15:222–234. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee YR, Chen M and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor: New modes
and prospects. Nat Rev Mol Cell Biol. 19:547–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kong Y, Yin J, Fu Y, Chen Y, Zhou Y and
Geng X: Suppression of Elk1 inhibits thyroid cancer progression by
mediating PTEN expression. Oncol Rep. 40:1769–1776. 2018.PubMed/NCBI
|
|
12
|
Tian DD, Zhang RX, Wu N, Yuan W, Luo SH,
Chen HQ, Liu Y, Wang Y, He BC and Deng ZL: Tetrandrine inhibits the
proliferation of human osteosarcoma cells by upregulating the PTEN
pathway. Oncol Rep. 37:2795–2802. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiong J, Li Z, Zhang Y, Li D, Zhang G, Luo
X, Jie Z, Liu Y, Cao Y, Le Z, et al: PRL-3 promotes the peritoneal
metastasis of gastric cancer through the PI3K/Akt signaling pathway
by regulating PTEN. Oncol Rep. 36:1819–1828. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Yang C, Zhou F and Chen X: PDK1
inhibitor GSK2334470 synergizes with proteasome inhibitor MG132 in
multiple myeloma cells by inhibiting full AKT activity and
increasing nuclear accumulation of the PTEN protein. Oncol Rep.
39:2951–2959. 2018.PubMed/NCBI
|
|
15
|
Zhao Y, Zheng R, Li J, Lin F and Liu L:
Loss of phosphatase and tensin homolog expression correlates with
clinicopathological features of non-small cell lung cancer patients
and its impact on survival: A systematic review and meta-analysis.
Thorac Cancer. 8:203–213. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu X, Li Z, Li T, Long F, Lv Y, Liu L,
Liu X and Zhan Q: Osthole inhibits the PI3K/AKT signaling pathway
via activation of PTEN and induces cell cycle arrest and apoptosis
in esophageal squamous cell carcinoma. Biomed Pharmacother.
102:502–509. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lim SL, Goh YM, Noordin MM, Rahman HS,
Othman HH, Bakar Abu NA and Mohamed S: Morinda citrifolia edible
leaf extract enhanced immune response against lung cancer. Food
Funct. 7:741–751. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang C, Cheng Y, Liu H, Xu Y, Peng H, Lang
J, Liao J, Liu H, Liu H and Fan J: Pectolinarigenin suppresses the
tumor growth in nasopharyngeal carcinoma. Cell Physiol Biochem.
39:1795–1803. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang T, Li S, Li J, Yin F, Hua Y, Wang Z,
Lin B, Wang H, Zou D, Zhou Z, et al: Natural product
pectolinarigenin inhibits osteosarcoma growth and metastasis via
SHP-1-mediated STAT3 signaling inhibition. Cell Death Dis.
7:e24212016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu C, Wang H, Chen S, Yang R, Li H and
Zhang G: Baicalein inhibits cell growth and increases cisplatin
sensitivity of A549 and H460 cells via miR-424-3p and targeting
PTEN/PI3K/Akt pathway. J Cell Mol Med. 22:2478–2487. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu C, Zhang B, Li YL and Yu XR: SIX1
reduces the expression of PTEN via activating PI3K/AKT signal to
promote cell proliferation and tumorigenesis in osteosarcoma.
Biomed Pharmacother. 105:10–17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Z, Hong S and Liu Z: LncRNA LINC00641
predicts prognosis and inhibits bladder cancer progression through
miR-197-3p/KLF10/PTEN/PI3K/AKT cascade. Biochem Biophys Res Commun.
503:1825–1829. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu J, Xing Y and Rong L: miR-181
regulates cisplatin-resistant non-small cell lung cancer via
downregulation of autophagy through the PTEN/PI3K/AKT pathway.
Oncol Rep. 39:1631–1639. 2018.PubMed/NCBI
|
|
25
|
Jing X, Cheng W, Wang S, Li P and He L:
Resveratrol induces cell cycle arrest in human gastric cancer
MGC803 cells via the PTEN-regulated PI3K/Akt signaling pathway.
Oncol Rep. 35:472–478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsuda S, Nakagawa Y, Kitagishi Y,
Nakanishi A and Murai T: Reactive oxygen species, superoxide
dimutases, and PTEN-p53-AKT-MDM2 signaling loop network in
mesenchymal stem/stromal cells regulation. Cells. 7:E362018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
de Assis LV and Isoldi MC: The function,
mechanisms, and role of the genes PTEN and TP53 and the effects of
asbestos in the development of malignant mesothelioma: A review
focused on the genes' molecular mechanisms. Tumour Biol.
35:889–901. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He M, Jiang L, Li B, Wang G, Wang J and Fu
Y: Oxymatrine suppresses the growth and invasion of MG63 cells by
up-regulating PTEN and promoting its nuclear translocation.
Oncotarget. 8:65100–65110. 2017.PubMed/NCBI
|
|
29
|
Oxnard GR, Hu Y, Mileham KF, Husain H,
Costa DB, Tracy P, Feeney N, Sholl LM, Dahlberg SE, Redig AJ, et
al: Assessment of resistance mechanisms and clinical implications
in patients with EGFR T790m-positive lung cancer and acquired
resistance to osimertinib. JAMA Oncol. Aug 2–2018.(Epub ahead of
print). View Article : Google Scholar
|
|
30
|
Dai Z, Wang L, Wang X, Zhao B, Zhao W,
Bhardwaj SS, Ye J, Yin Z, Zhang J and Zhao S: Oxymatrine induces
cell cycle arrest and apoptosis and suppresses the invasion of
human glioblastoma cells through the EGFR/PI3K/Akt/mTOR signaling
pathway and STAT3. Oncol Rep. 40:867–876. 2018.PubMed/NCBI
|
|
31
|
Li W, Yu X, Ma X, Xie L, Xia Z, Liu L, Yu
X, Wang J, Zhou H, Zhou X, et al: Deguelin attenuates non-small
cell lung cancer cell metastasis through inhibiting the CtsZ/FAK
signaling pathway. Cell Signal. 50:131–141. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Koundouros N and Poulogiannis G:
Phosphoinositide 3-kinase/Akt signaling and redox metabolism in
cancer. Front Oncol. 8:1602018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liao K, Yong CW and Hua K: SB431542
inhibited cigarette smoke extract induced invasiveness of A549
cells via the TGF-β1/Smad2/MMP3 pathway. Oncol Lett. 15:9681–9686.
2018.PubMed/NCBI
|
|
34
|
Wu C, Zhu X, Liu W, Ruan T, Wan W and Tao
K: NFIB promotes cell growth, aggressiveness, metastasis and EMT of
gastric cancer through the Akt/Stat3 signaling pathway. Oncol Rep.
40:1565–1573. 2018.PubMed/NCBI
|
|
35
|
Xing Y, Liu Y, Liu T, Meng Q, Lu H, Liu W,
Hu J, Li C, Cao M, Yan S, et al: TNFAIP8 promotes the proliferation
and cisplatin chemoresistance of non-small cell lung cancer through
MDM2/p53 pathway. Cell Commun Signal. 16:432018. View Article : Google Scholar : PubMed/NCBI
|