Introduction
Hepatocellular carcinoma (HCC) is the most common
type of liver malignancy, which results in chronic inflammation in
the liver (1,2). Typical HCC progression is a multistep
process involving transformation, survival, proliferation,
invasion, angiogenesis and metastasis (3,4).
Although, a number of clinicopathological factors are important in
the treatment of HCC, there is no marked and effective medical
therapy for patients with advanced HCC (5–8).
Therefore, it is necessary to develop more efficient therapeutic
strategies and targets for patients with HCC.
Hypoxia-inducible factor-1 (HIF-1), a heterodimer
composed of an oxygen-regulated HIF-1α subunit and a HIF-1β
subunit, activates the transcription of genes involved in
proliferation and angiogenesis (9–11).
HIF-1β is stable under hypoxic or normoxic conditions. By contrast,
HIF-1α is an oxygen sensitive subunit and its expression is induced
under hypoxic conditions (12). The
degradation of HIF-1α subunit is facilitated by ubiquitination
following the hydroxylation of proline residues (13,14).
The other post-transnational modifications, such as acetylation and
phosphorylation reactions, can also affect the stability of HIF-1α
(15,16). Activated HIF-1 serves a crucial role
in the regulation of the expression of certain enzymes in the
glycolytic pathway, such as glucose transporters, hexokinase,
phosphofructokinase, pyruvate kinase and pyruvate dehydrogenase
(17–19). Vascular endothelial growth factor
(VEGF), one of the most important angiogenic factors in cancer
progression, is a HIF-1α target gene. VEGF has also been revealed
to regulate cancer cell proliferation and migration (20).
HIF-1α functions by binding hypoxia response
elements (HREs) within its target metabolism-related genes
(21). Fructose-1,6-biphosphatase
(FBP1), which catalyzes the splitting of fructose-1,6-bisphosphate
into fructose 6-phosphate and inorganic phosphate, can inhibit
HIF-1α activity via direct interaction with the HIF-1α C-terminus
(22). The epithelial-mesenchymal
transition (EMT) promotes invasiveness and stem cell-like features
in cancer cells (23,24). The EMT involves the downregulation
of E-cadherin to reinforce the destabilization of adherens
junctions. EMT-related transcription factors, such as Snail, Slug
and Twist, can suppress E-cadherin expression (25,26).
Snail interacts with H3K9 methyltransferase G9a and DNA
methyltransferase Dnmt1 to silence E-cadherin expression.
Similarly, the Snail-G9a-Dnmt1 complex also suppresses FBP1
expression, and thus enhances aerobic glycolysis (27).
LOX-like protein 2 (LOXL2) is a member of the lysyl
oxidase gene family, with both intracellular and extracellular
functions (28,29). Extracellularly, LOXL2 catalyzes the
covalent cross-linkages of collagen and elastin in the
extracellular matrix. Intracellularly, LOXL2 modifies histone tails
and reduces cell polarity, which increases the metastatic potential
of tumors (30–32). Intracellular LOXL2 has also been
revealed to stabilize Snail protein and promote EMT in certain
breast cancer cell lines through interaction with Snail (33). Thus, small molecule LOXL2 inhibitors
were considered as suitable drug candidates for treatment of
numerous types of cancers. LOXL2-IN-1 hydrochloride (LOXL2-IN-1) is
the first published small molecule inhibitor selective for LOXL2
with a strong inhibitory effect (32). The most established LOXL2 inhibitor
is β-aminoproprionitrile (BAPN), which irreversibly inhibits the
enzyme activity of LOXL2 (34).
Recently, a subseries of LOXL2 specific inhibitors containing an
aminomethiophene (AMT) scaffold were revealed to inhibit tumor
growth (35–37).
The present study demonstrated that LOXL2
upregulated HIF-1α/VEGF signaling pathways via the Snail-FBP1 axis,
and this phenomenon could be inhibited by LOXL2 inhibition. LOXL2
represents a potential therapeutic target for HCC. These findings
further support the role of LOXL2 in the progression of HCC and
implicates LOXL2 as a potential therapeutic agent for the treatment
of HCC.
Materials and methods
Cell cultures
293T cells and human HCC cell lines Huh7 and Hep3B
were purchased from the American Type Culture Collection. Huh7,
Hep3B and 293T cells were cultured in Dulbecco's modified Eagles
medium (DMEM; HyClone; GE Healthcare Life Sciences) supplemented
with 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences),
penicillin (100 U/ml) and streptomycin (100 µg/ml) in a humidified
atmosphere of 5% CO2 at 37°C. Human umbilical vein
endothelial cells (HUVECs) were purchased from PromoCell Academy
and cultured in Endothelial Cell Growth Medium 2 (PromoCell
Academy) with Supplement Mix according to the manufacturer's
guidelines.
Reagents
LOXL2-IN-1 hydrochloride (LOXL2-IN-1) was purchased
from MedChemExpress. Anti-human LOXL2 therapeutic antibody
(Simtuzumab; cat. no. HPAB-0160-LSX) was obtained from Creative
Biolabs. Antibodies against LOXL2 (cat. no. ab179810; 1:1,000),
Snail (cat. no. ab53519; 1:1,000), FBP1 (cat. no. ab109020;
1:1,000), VEGF (cat. no. ab69479; 1:1,000), Transferrin (cat. no.
ab88165; 1:2,000) and β-actin (cat. no. ab6276; 1:5,000) were all
obtained from Abcam. Antibody against HIF-1α (cat. no. 14179;
1:500) and GAPDH (cat. no. 5174; dilution 1:2,000) were purchased
from Cell Signaling Technology, Inc.
Quantification of glucose uptake and
lactate generation
Glucose uptake was assessed in the cell lysates with
Glucose Uptake Colorimetric assay kit (BioVision, Inc.). The
extracellular lactate was assessed in the medium with a lactate
assay kit (BioVision, Inc.). Both assays were performed according
to the manufacturer's protocol.
Lentiviral vector construction
Wild-type LOXL2 and the point-mutated form of LOXL2
[LOXL2(Y689F)] in pBOBI lentiviral vector were purchased from
Rosetta Stone Biotech Co., Ltd. Wild-type Snail in pBOBI lentiviral
vector was purchased from Rosetta Stone Biotech Co., Ltd. The
sequences of the lentiviral short hairpin RNAs (shRNAs) in
pLKO.1-puro vectors were as follows: sh1LOXL2,
5′-GAAACCCTCCAGTCTATTATA-3′; sh2LOXL2, 5′-GGCAATGAGAAGTCCATTATA-3′;
sh1FBP1, 5′-CCACCATCAAATGCTGTAGAA-3′; and sh2FBP1,
5′-CCACCATCAAATGCTGTAGAA-3′. The sequence of the scrambled hairpin
lentiviral shRNA pLKO.1-puro control (PLK) was
5′-CCTAAGGTTAAGTCGCCCTCGCTCGAGCGAGGGCGACTTAACCTTAGG-3′.
Viral production and infection
293T cells (2×106/cell culture dish) were
plated and transfected with different lentiviral vectors together
with packaging plasmids using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). After 48 h of transfection,
culture supernatants were harvested and filtered with a Millipore
Stericup filter (0.45-µm). Huh7 or Hep3B cells were infected with
different lentivirus for 24 h. After 4 days, the effects on these
cells were examined by western blotting.
RT-qPCR assay
Total RNA was isolated from Huh7 cells by TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). The purity and
quantity of RNA were assessed with a NanoDrop 100 spectrophotometer
(Thermo Fisher Scientific, Inc.). cDNA was synthesized with a
reverse transcriptase kit (Thermo Fisher Scientific, Inc.).
SYBR-Green (Takara Biotechnology Co., Ltd.) was added to quantify
the expression of VEGF, according to the protocol provided. The
sequences of the primers were: VEGF, 5′-TTGCAGATGTGACAAGCCGA-3′ and
5′-GGCCGCGGTGTGTCTA-3′; GAPDH, 5′-GCACCGTCAAGGCTGAGAAC-3′ and
5′-TGGTGAAGACGCCAGTGGA-3′. PCR reaction conditions were performed
as follows: 95°C for 60 sec, and 40 cycles of 95°C for 15 sec, 60°C
for 15 sec, and 72°C for 45 sec. The relative expression levels
were determined using the comparative ΔΔCq method (38).
Colony forming analysis
The different types of HCC cells were seeded at a
density of 300 cells/well in 6-well plates and cultured for 14
days. The culture medium was replaced every 3 days. Colonies were
fixed with 10% buffered formalin at 37°C for 15 min and stained
with 2% crystal violet at 37°C for 30 min. Colonies were counted
under a microscope (Micropublisher 3.3RTV; Olympus Corporation),
and colony forming was determined as the colony formation rate
(number of cell clones experimental group/cell clones in the
control group ×100%).
Western blot analysis
To analyze the expression of proteins, western blot
analysis was performed. Total protein was isolated from the cells
using RIPA buffer containing a protease and phosphatase inhibitor
cocktail (Thermo Fisher Scientific, Inc.). The lysate was
centrifuged at 20,8000 × g for 15 min at 4°C. The supernatants were
collected and mixed with 5X loading buffer, followed by incubation
in a boiling water bath for 5 min. Protein concentration was
determined by Bradford protein assay. Western blot analysis was
performed using 12% SDS-PAGE gels for different proteins (50 µg per
lane). Following transfer to PVDF membranes, 5% non-fat milk in
TBST was used for blocking for 1 h at room temperature, followed by
incubation overnight at 4°C with each of the indicated primary
antibodies. Subsequently, the membranes were washed with PBST three
times for 10 min. The membranes were then probed with
HRP-conjugated secondary antibodies (cat. no. ab205719 and cat. no.
ab205718; Abcam) in TBST for 1 h at room temperature, and washed
with TBST three times. Following washing, enhanced
chemiluminescence was used to incubate the membrane and bands were
visualized using the ECL Plus Western Blotting Detection system™
(GE Healthcare Life Sciences). Signals were detected and documented
with the densitometry system LAS-3000 (Fujifilm).
Tube formation assay
A tube formation assay was used to evaluate vascular
activity of HCC cells on HUVECs tube formation. The upper chamber
was prepared by plating HUVECs onto Matrigel basement membrane
matrix (BD Biosciences) in Transwell filters (0.4-µm pore size;
Costar) followed by incubation at 37°C for 24 h. HUVECs and HCC
cells were co-cultured at 37°C for 48 h. Tubes were observed under
an inverted light microscope.
Immunoprecipitation assays
Huh7 cells were seeded in a 10-cm dish at an initial
concentration of 2×106 cells. After 24 h, the medium was
replaced with serum-free DMEM and cultured for a further 24 h. The
medium was centrifuged at 20,8000 × g for 15 min at 4°C and the
supernatant was then filtered by ultrafiltration using Amicon
Ultra-4 (5K; EMD Millipore, Merck KGaA). Concentrated protein (30
µl) was collected (per dish) and then added to 1 ml RIPA buffer
(Sigma-Aldrich; Merck KGaA). Following incubation with simtuzumab
antibody for 8 h at 4°C, A/G-agarose beads were added with gentle
rocking for 3 h at 4°C. Following centrifugation at 1,008 × g for
30 sec at 4°C, the pellets were washed with RIPA buffer three times
and re-suspended in 2X SDS sample buffer. The samples were then
subjected to 12% SDS gel electrophoresis.
Statistical analysis
All statistical analyses were performed with
GraphPad Prism 6.0 (GraphPad Software, Inc.). The data were
presented as the mean ± standard deviation (SD), and the
experiments were performed in triplicate. For data analyses, a
two-tailed Student's t-test was used to examine the differences
between groups, and comparisons between multiple groups were
determined by one-way analysis of variance (ANOVA) followed by
Tukey's multiple comparison test. In all comparisons, P<0.05 was
considered to indicate a statistically significant difference.
Results
Intracellular LOXL2 inhibits the
expression of FBP1 in HCC cell lines
LOXL2 has been reported to promote EMT by reducing
E-cadherin expression via the upregulation of Snail expression
(33). Snail interacts with the
H3K9 methyltransferase G9a and the DNA methyltransferase Dnmt1 to
induce E-cadherin gene repression. Similarly, the Snail-G9a-Dnmt1
complex also suppresses FBP1 expression, and thus enhances aerobic
glycolysis (27). Therefore, the
present study examined whether knockdown of LOXL2 can affect the
expression level of FBP1 in HCC cells. As presented in Fig. 1A, LOXL2-knockdown markedly inhibited
the expression of Snail. In addition, the expression of FBP1 was
upregulated in LOXL2-knockdown Huh7 and Hep3B HCC cells (Fig. 1A). Furthermore, overexpression of
wild-type LOXL2 increased the expression of Snail and inhibited the
expression of FBP1 (Fig. 1B).
Notably, overexpression of the point-mutated form of LOXL2
[LOXL2(Y689F)], which lacks enzymatic activity, did not affect the
expression of Snail1 or FBP1 (Fig.
1B). LOXL2 is a member of the lysyl oxidase family, with both
intracellular and extracellular functions. To examine whether the
intracellular LOXL2 or extracellular LOXL2 plays a requisite role
in regulation of Snail or FBP1 expression, the anti-human LOXL2
therapeutic antibody simtuzumab was used. Simtuzumab is a humanized
monoclonal antibody designed for the treatment of fibrosis
(39,40). It binds to LOXL2 and acts as an
immunomodulator. The binding between simtuzumab and extracellular
LOXL2 protein was first evaluated with immunoprecipitation assays.
The data demonstrated good binding between extracellular LOXL2
protein of Huh7 cells and simtuzumab (Fig. 1C). Following incubation of Huh7
cells with simtuzumab antibody, the expression of Snail and FBP1
was examined by western blotting. Simtuzumab did not affect the
expression levels of Snail and FBP1 (Fig. 1C). Therefore, intracellular LOXL2
was responsible for the regulation of Snail and FBP1
expression.
LOXL2 inhibits the expression of FBP1
and enhances the glycolysis in a Snail-dependent manner
To further determine the mechanism of
LOXL2-regulated expression of FBP1, Snail was overexpressed in
LOXL2 stable knockdown HCC cells. As presented in Fig. 2A, LOXL2 knockdown did not
significantly affect the expression of FBP1 in Snail-overexpressing
HCC cells. Loss of FBP1 is reported to increase glycolysis,
including cellular glucose uptake and lactate generation (27). To examine whether LOXL2 affects
glycolysis in HCC cells, the present study assessed glucose uptake
and lactate generation, and it was revealed that LOXL2-knockdown
significantly decreased the glucose uptake and lactate generation,
whereas Snail-overexpressing Huh7 and Hep3B cells had normal
glucose uptake and lactate generation compared with their vector
control cells (Fig. 2B). Tumor
cells exhibit high aerobic glycolysis during rapid proliferation
(27,41,42).
LOXL2 has also been reported to upregulate cell proliferation in
clear cell renal cell carcinoma (43). The present study revealed that
LOXL2-knockdown suppressed the proliferation of HCC cells but not
Snail-overexpressing HCC cells (Fig.
2C). These results indicated that LOXL2 inhibited the
expression of FBP1 and enhanced the glycolysis in a Snail-dependent
manner.
 | Figure 2.LOXL2 inhibits the expression of FBP1
and enhances glycolysis. (A) The expression of LOXL2, Snail and
FBP1 were detected using western blot in vector control (PLKO),
LOXL2 knockdown (sh1), and LOXL2 knockdown plus
Snail-overexpressing Huh7 or Hep3B cells. β-actin was used as a
loading control. (B) Glucose uptake and lactate production were
detected in culture medium or the lysates and normalized to total
cellular protein amount in vector control, LOXL2 knockdown, and
LOXL2 knockdown plus Snail-overexpressing Huh7 or Hep3B cells.
*P<0.05 vs. PLKO or the sh1LOXL2-Snail groups. (C) Colony
formation assay was used to estimate the cell proliferation in
vector control, LOXL2 knockdown, and LOXL2 knockdown plus
Snail-overexpressing Huh7 or Hep3B cells. Representative images of
the colony formation assay and quantification of colony formation
efficiency in various types of HCC cells. **P<0.01 vs. PLKO or
the sh1LOXL2-Snail groups. LOXL2, lysyl oxidase-like 2; FBP1,
fructose-1, 6-biphosphatase. |
LOXL2 regulates angiogenesis and
expression of VEGF
A previous study has reported that LOXL2 can promote
bFGF-induced tumor angiogenesis (44). In the present study, HUVECs were
co-cultured along with LOXL2-knockdown or the vector control HCC
cells. As presented in Fig. 3A,
HUVECs co-cultured with LOXL2-knockdown HCC cells reduced the
generation of tubular networks. By contrast, HUVECs co-cultured
with LOXL2-knockdown and Snail-overexpressing HCC cells exhibited a
normal generation of tubular networks (Fig. 3A). It was then revealed that
knockdown of LOXL2 in Huh7 or Hep3B cells inhibited the expression
of VEGF but not in the Snail-overexpressing HCC cells (Fig. 3B). These findings indicated that
LOXL2 may be involved in the regulation of angiogenesis in HCC.
Overexpression of LOXL2 or knockdown
of FBP1 increases the expression of HIF-1α
VEGF is known to be one of the target genes of
HIF-1α (45). The present study
further investigated the role of LOXL2 in the regulation of HIF-1α
expression. As presented in Fig.
4A, overexpression of wild-type LOXL2 resulted in increased
expression levels of HIF-1α in Huh7 and Hep3B cells. However,
overexpression of the point-mutated form LOXL2 (Y689F), which lacks
enzymatic activity, failed to regulate the expression of HIF-1α in
Huh7 or Hep3B cells (Fig. 4A). FBP1
can inhibit HIF-1α activity via direct interaction with the HIF-1α
C-terminus. Furthermore, FBP1 was reported to modulate cell
metabolism of breast cancer cells by inhibiting the expression of
HIF-1α (46). It was also revealed
that knockdown of FBP1 increased the expression of HIF-1α (Fig. 4B). These data indicated that LOXL2
may increase the expression of HIF-1α through inhibition of the
expression of FBP1.
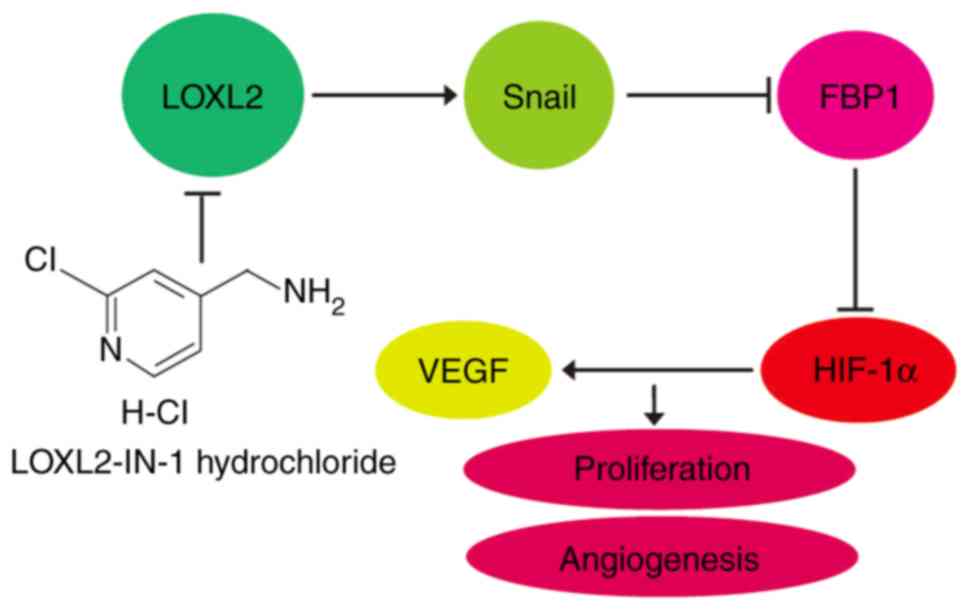
LOXL2 upregulates HIF-1α/VEGF
signaling pathways via the Snail-FBP1 axis
The present study further investigated the mechanism
of LOXL2-induced regulation of HIF-1α expression. It was revealed
that knockdown of FBP1 in Huh7 or Hep3B cells did not alter the
expression levels of Snail (Fig.
5A). This suggests that FBP1 only regulates the expression of
HIF-1α, and Snail is the upstream suppressor of FBP1 (Fig. 2A). LOXL2-IN-1 hydrochloride
(LOXL2-IN-1) is a selective LOXL2 inhibitor. Similar to LOXL2
knockdown, LOXL2-IN-1 treatment resulted in a similar effect on the
expression of Snail, FBP1, HIF-1α and VEGF in Huh7 or Hep3B cells
(Fig. 5B). Inhibition of LOXL2 by
LOXL2-IN-1 increased the expression of FBP1. Therefore, the effect
of LOXL2-IN-1 in FBP1-knockdown HCC cells was examined. As
presented in Fig. 5B, LOXL2-IN-1
did not affect the expression of HIF-1α and VEGF in FBP1-knockdown
HCC cells compared to the DMSO group. HIF-1α/VEGF signaling was
upregulated by LOXL2. The effect of LOXL2 inhibitor in combination
with HIF-1α inhibitor PX-478, the first novel HIF-1α inhibitor in
clinical stage for the treatment of solid tumors, was therefore
assessed (47). A synergistic
inhibitory effect of LOXL2-IN-1 inhibitor with PX-478 inhibitor on
the expression of VEGF was demonstrated in Fig. 5C. In summary, these results
indicated that LOXL2 upregulates HIF-1α/VEGF signaling pathways via
the Snail-FBP1 axis (Fig. 6).
 | Figure 5.LOXL2 upregulates HIF-1α/VEGF
signaling pathways via the Snail-FBP1 axis. (A) The expression
levels of HIF-1α, Snail and FBP1 in vector control and
FBP1-knockdown Huh7 or Hep3B cells were detected using western
blotting. (B) Huh7 or Hep3B cells were treated with DMSO (the
vehicle of LOXL2-IN-1); the vector control (PLKO) or FBP1-knockdown
Huh7 or Hep3B cells were treated with LOXL2-IN-1 hydrochloride
(LOXL2-IN-1, 50 nM). All these HCC cells were treated with DMSO or
LOXL2-IN-1 for 12 h. The expression of LOXL2, Snail, FBP1 and
HIF-1α were detected using western blotting. β-actin was used as a
loading control. (C) Huh7 cells were treated with DMSO, PX-478 (10
µM), LOXL2-IN-1 (50 nM) or PX-478 (10 µM) in combination with
LOXL2-IN-1 (50 nM) for 12 h. VEGF mRNA expression was confirmed by
quantitative RT-PCR assay. *P<0.01 vs. the DMSO treatment group,
**P<0.05 vs. PX-478 or LOXL2-IN-1 treatment group. LOXL2, lysyl
oxidase-like 2; HIF-1α, hypoxia-inducible factor 1α; VEGF, vascular
endolthelial growth factor; FBP1, fructose-1, 6-biphosphatase. |
Discussion
HCC is the most common type of liver malignancy and
its progression is a multistep process involving transformation,
survival, proliferation, invasion, angiogenesis and metastasis
(3,8). Presently, there is no marked and
effective medical therapy for patients with advanced HCC (48). Therefore, it is necessary and
crucial to develop more efficient therapeutic strategies and
targets for patients with HCC.
LOXL2 is a member of the LOX gene family, which
encodes a copper-dependent amine oxidase that catalyzes the first
step in the formation of crosslinks in collagens and elastin
(49,50). It is involved in the progression of
tumor progression and metastasis (31,49,51–53).
Upregulated expression of LOXL2 has been revealed in several types
of cancer, including HCC (54).
Extracellularly, LOXL2 can catalyze the covalent cross-linkages of
collagen and elastin in the extracellular matrix. Intracellularly,
LOXL2 modifies histone tails and reduces cell polarity, which
increases metastatic potential of tumors (55,56).
Interacting with intracellular Snail protein, intracellular LOXL2
has been reported to stabilize Snail protein and promote EMT in
certain breast cancer cells (33).
The present study revealed that knockdown of LOXL2 inhibited the
expression of Snail. Furthermore, overexpression of wild-type LOXL2
increased the expression of Snail. Notably, overexpression of the
point-mutated form of LOXL2 (Y689F), which lacks enzymatic
activity, did not affect the expression of Snail. Furthermore,
targeting extracellular LOXL2 of HCC cells with a therapeutic
antibody was unable to abolish its regulation to the expression of
Snail. These results indicated that intracellular activity of LOXL2
is responsible for Snail upregulation.
FBP1, a rate-limiting enzyme in gluconeogenesis,
catalyzes the hydrolysis of fructose 1,6-bisphosphate to fructose
6-phosphate and inorganic phosphate (27,57).
Snail interacts with G9a and Dnmt1 to suppress FBP1 expression, and
thus enhances aerobic glycolysis. The present study revealed that
LOXL2-knockdown did not significantly affect the expression of FBP1
in Snail-overexpressing HCC cells. Similar results were obtained in
the analysis of cellular glucose uptake and lactate generation in
Snail-overexpressing HCC cells. Knockdown of FBP1 in HCC cells did
not alter the expression levels of Snail. Thus, Snail is an
upstream suppressor of FBP1. These results indicated that LOXL2
inhibited the expression of FBP1 and enhanced the glycolysis in a
Snail-dependent manner. There have been numerous studies that have
revealed the expression of LOXL2, Snail and FBP1 in human HCC
tissues. For example, LOXL2 was revealed to be frequently
overexpressed in human HCC and displayed poor prognosis (54,58–60).
High expression of Snail or decreased expression of FBP1 was
correlated with poor outcomes in HCC patients (61–66).
Notably, HCC patients with high Snail but low FBP1 expression were
identified with the worst prognosis. Conversely, patients with low
Snail but high FBP1 expression were identified with the best
prognosis (67).
HIF-1α is a subunit of the heterodimeric
transcription factor HIF-1, which is considered to be a master
transcriptional regulator of cellular and developmental responses
to hypoxia (68). In accordance
with its dynamic biological role, HIF-1α forms a heterodimer with
HIF-1β and thus associates with HREs of promoters of
hypoxia-responsive genes to induce transcription (69). VEGF is known to be a target gene of
HIF-1α. VEGF is one of the most important angiogenic factors in
cancer progression and is important to regulate cancer cell
proliferation and migration (70).
The present study indicated that LOXL2 could induce cell
proliferation and angiogenesis of HCC cells. Accordingly, it also
indicated that LOXL2 was responsible for the upregulation of HIF-1α
and VEGF.
FBP1 has been reported to inhibit HIF-1α activity or
expression (27,46). The present study also revealed that
knockdown of FBP1 increased the expression of HIF-1α in HCC cells.
Additionally, knockdown of FBP1 in HCC cells did not alter the
expression levels of Snail. This indicated that FBP1 only regulated
the expression of HIF-1α, and Snail was an upstream suppressor of
FBP1. Inhibition of the activity of LOXL2 by LOXL2-IN-1 also
demonstrated a similar effect in LOXL2-knockdown HCC cells to the
expression of Snail, FBP1, HIF-1α and VEGF. By contrast, LOXL2-IN-1
did not alter the expression of Snail, HIF-1α and VEGF in
FBP1-knockdown HCC cells. Moreover, it was revealed that LOXL2-IN-1
inhibitor could exert a synergistic inhibitory effect with HIF-1α
inhibitor PX-478 on the expression of VEGF. HCC is a highly
angiogenic cancer (71,72), and therefore antiangiogenic therapy
is an effective strategy in the treatment of HCC. LOX-L2 inhibitors
could be potent combination partners for the antiangiogenic drugs
approved for the treatment of HCC such as Cabometyx, Cyramza,
Lenvatinib Mesylate, Ramucirumab and Sorafenib Tosylate (73,74).
The present study revealed a potential role of LOXL2 in HCC, which
may provide new insights into the treatment of HCC. However, a
limitation of the present study was that the effect of LOXL2
therapeutic antibody or LOXL2 inhibitor was not verified in an
in vivo study. We will further investigate the effect of
LOXL2 therapeutic antibody or LOXL2 inhibitor in animal models of
HCC.
In conclusion, the present study demonstrated that
LOXL2 upregulated HIF-1α/VEGF signaling pathways via the Snail-FBP1
axis, and this phenomenon could be inhibited by LOXL2 inhibition.
Therefore, these findings further support that LOXL2 serves an
important role in the progression of HCC. Considering the high
expression level of LOXL2 in HCC observed in several studies, LOXL2
may be considered a promising candidate for novel treatment
strategies against HCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Nature Science Foundation of China (grant no. 81502694).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All the authors participated in data collection and
analysis. ZF, QJ and QL participated in the design of the study. ZF
and QL participated in the writing of the manuscript and data
interpretation. All authors read and approved the final manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hatzaras I, Bischof DA, Fahy B, Cosgrove D
and Pawlik TM: Treatment options and surveillance strategies after
therapy for hepatocellular carcinoma. Ann Surg Oncol. 21:758–766.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Morise Z, Kawabe N, Tomishige H, Nagata H,
Kawase J, Arakawa S, Yoshida R and Isetani M: Recent advances in
liver resection for hepatocellular carcinoma. Front Surg. 1:212014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chan TH, Lin CH, Qi L, Fei J, Li Y, Yong
KJ, Liu M, Song Y, Chow RK, Ng VH, et al: A disrupted RNA editing
balance mediated by ADARs (Adenosine DeAminases that act on RNA) in
human hepatocellular carcinoma. Gut. 63:832–843. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raveh E, Matouk IJ, Gilon M and Hochberg
A: The H19 Long non-coding RNA in cancer initiation, progression
and metastasis-a proposed unifying theory. Mol Cancer. 14:1842015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu AX, Kang YK, Rosmorduc O, Evans TR,
Santoro A, Ross P, Gane E, Vogel A, Jeffers M, Meinhardt G and Peña
CE: Biomarker analyses of clinical outcomes in patients with
advanced hepatocellular carcinoma treated with sorafenib with or
without erlotinib in the SEARCH trial. Clin Cancer Res.
22:4870–4879. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ikeda M, Shimizu S, Sato T, Morimoto M,
Kojima Y, Inaba Y, Hagihara A, Kudo M, Nakamori S, Kaneko S, et al:
Sorafenib plus hepatic arterial infusion chemotherapy with
cisplatin versus sorafenib for advanced hepatocellular carcinoma:
Randomized phase II trial. Ann Oncol. 27:2090–2096. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lencioni R, Chen XP, Dagher L and Venook
AP: Treatment of intermediate/advanced hepatocellular carcinoma in
the clinic: How can outcomes be improved? Oncologist. 15 (Suppl
4):S42–S52. 2010. View Article : Google Scholar
|
|
8
|
Finn RS, Zhu AX, Farah W, Almasri J, Zaiem
F, Prokop LJ, Murad MH and Mohammed K: Therapies for advanced stage
hepatocellular carcinoma with macrovascular invasion or metastatic
disease: A systematic review and meta-analysis. Hepatology.
67:422–435. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carmeliet P, Dor Y, Herbert JM, Fukumura
D, Brusselmans K, Dewerchin M, Neeman M, Bono F, Abramovitch R,
Maxwell P, et al: Role of HIF-1alpha in hypoxia-mediated apoptosis,
cell proliferation and tumour angiogenesis. Nature. 394:485–490.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eyries M, Siegfried G, Ciumas M, Montagne
K, Agrapart M, Lebrin F and Soubrier F: Hypoxia-induced apelin
expression regulates endothelial cell proliferation and
regenerative angiogenesis. Circ Res. 103:432–440. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Semenza GL: Hypoxia-inducible factor 1
(HIF-1) pathway. Sci STKE. 2007:cm82007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Lu H, Xiang L, Bullen JW, Zhang
C, Samanta D, Gilkes DM, He J and Semenza GL: HIF-1 regulates CD47
expression in breast cancer cells to promote evasion of
phagocytosis and maintenance of cancer stem cells. Proc Natl Acad
Sci USA. 112:E6215–E6223. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Botusan IR, Sunkari VG, Savu O, Catrina
AI, Grünler J, Lindberg S, Pereira T, Ylä-Herttuala S, Poellinger
L, Brismar K and Catrina SB: Stabilization of HIF-1alpha is
critical to improve wound healing in diabetic mice. Proc Natl Acad
Sci USA. 105:19426–19431. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim DH, Sung B, Kim JA, Kang YJ, Hwang SY,
Hwang NL, Suh H, Choi YH, Im E, Chung HY and Kim ND: HS-1793, a
resveratrol analogue, downregulates the expression of
hypoxia-induced HIF-1 and VEGF and inhibits tumor growth of human
breast cancer cells in a nude mouse xenograft model. Int J Oncol.
51:715–723. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Masoud GN and Li W: HIF-1α pathway: Role,
regulation and intervention for cancer therapy. Acta Pharm Sin B.
5:378–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Choudhry H and Harris AL: Advances in
hypoxia-inducible factor biology. Cell Metab. 27:281–298. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Semenza GL: HIF-1: Upstream and downstream
of cancer metabolism. Curr Opin Genet Dev. 20:51–56. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Soga T: Cancer metabolism: Key players in
metabolic reprogramming. Cancer Sci. 104:275–281. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meijer TW, Kaanders JH, Span PN and
Bussink J: Targeting hypoxia, HIF-1, and tumor glucose metabolism
to improve radiotherapy efficacy. Clin Cancer Res. 18:5585–5594.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fan C, Tang Y, Wang J, Xiong F, Guo C,
Wang Y, Zhang S, Gong Z, Wei F, Yang L, et al: Role of long
non-coding RNAs in glucose metabolism in cancer. Mol Cancer.
16:1302017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang S, Wang X, Weng Z, Zhang S, Ning H
and Li B: Expression and role of microRNA 18b and hypoxia inducible
factor-1α in placental tissues of preeclampsia patients. Exp Ther
Med. 14:4554–4560. 2017.PubMed/NCBI
|
|
22
|
Li B, Qiu B, Lee DS, Walton ZE, Ochocki
JD, Mathew LK, Mancuso A, Gade TP, Keith B, Nissim I and Simon MC:
Fructose-1,6-bisphosphatase opposes renal carcinoma progression.
Nature. 513:251–255. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sato R, Semba T, Saya H and Arima Y:
Concise review: Stem cells and epithelial-mesenchymal transition in
cancer: Biological implications and therapeutic targets. Stem
Cells. 34:1997–2007. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
May CD, Sphyris N, Evans KW, Werden SJ,
Guo W and Mani SA: Epithelial-mesenchymal transition and cancer
stem cells: A dangerously dynamic duo in breast cancer progression.
Breast Cancer Res. 13:2022011. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi S, Zhao J, Wang J, Mi D and Ma Z: HPIP
silencing inhibits TGF-β1-induced EMT in lung cancer cells. Int J
Mol Med. 39:479–483. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dong C, Yuan T, Wu Y, Wang Y, Fan TW,
Miriyala S, Lin Y, Yao J, Shi J, Kang T, et al: Loss of FBP1 by
Snail-mediated repression provides metabolic advantages in
basal-like breast cancer. Cancer Cell. 23:316–331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jourdan-Le Saux C, Tronecker H, Bogic L,
Bryant-Greenwood GD, Boyd CD and Csiszar K: The LOXL2 gene encodes
a new lysyl oxidase-like protein and is expressed at high levels in
reproductive tissues. J Biol Chem. 274:12939–12944. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi SE, Jeon N, Choi HY, Shin JI, Jeong
HJ and Lim BJ: Lysyl oxidase-like 2 is expressed in kidney tissue
and is associated with the progression of tubulointerstitial
fibrosis. Mol Med Rep. 16:2477–2482. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu L and Zhu Y: The function and
mechanisms of action of LOXL2 in cancer (Review). Int J Mol Med.
36:1200–1204. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cuevas EP, Eraso P, Mazón MJ, Santos V,
Moreno-Bueno G, Cano A and Portillo F: LOXL2 drives
epithelial-mesenchymal transition via activation of IRE1-XBP1
signalling pathway. Sci Rep. 7:449882017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hutchinson JH, Rowbottom MW, Lonergan D,
Darlington J, Prodanovich P, King CD, Evans JF and Bain G: Small
molecule lysyl Oxidase-like 2 (LOXL2) inhibitors: The
identification of an inhibitor selective for LOXL2 over LOX. ACS
Med Chem Lett. 8:423–427. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peinado H, Del Carmen Iglesias-de la Cruz
M, Olmeda D, Csiszar K, Fong KS, Vega S, Nieto MA, Cano A and
Portillo F: A molecular role for lysyl oxidase-like 2 enzyme in
snail regulation and tumor progression. EMBO J. 24:3446–3458. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chitty JL, Setargew YFI and Cox TR:
Targeting the lysyl oxidases in tumour desmoplasia. Biochem Soc
Trans. 47:1661–1678. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Leung L, Niculescu-Duvaz D, Smithen D,
Lopes F, Callens C, McLeary R, Saturno G, Davies L, Aljarah M,
Brown M, et al: Anti-metastatic Inhibitors of Lysyl Oxidase (LOX):
Design and Structure-activity relationships. J Med Chem.
62:5863–5884. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tang H, Leung L, Saturno G, Viros A, Smith
D, Di Leva G, Morrison E, Niculescu-Duvaz D, Lopes F, Johnson L, et
al: Lysyl oxidase drives tumour progression by trapping EGF
receptors at the cell surface. Nat Commun. 8:149092017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Smithen DA, Leung LMH, Challinor M,
Lawrence R, Tang H, Niculescu-Duvaz D, Pearce SP, Mcleary R, Lopes
F, Aljarah M, et al: 2-Aminomethylene-5-sulfonylthiazole inhibitors
of Lysyl Oxidase (LOX) and LOXL2 show significant efficacy in
delaying tumor growth. J Med Chem. Sep 4–2019.(Epub ahead of
print). doi: 10.1021/acs.jmedchem.9b01112.
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Harrison SA, Abdelmalek MF, Caldwell S,
Shiffman ML, Diehl AM, Ghalib R, Lawitz EJ, Rockey DC, Schall RA,
Jia C, et al: Simtuzumab is ineffective for patients with bridging
fibrosis or compensated cirrhosis caused by nonalcoholic
steatohepatitis. Gastroenterology. 155:1140–1153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Altinbas A: A quick overview to the early
phase clinical trials of Simtuzumab®: Are we loosing the
most promising anti-fibrotic product? Med Hypotheses. 108:159–160.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lunt SY and Vander Heiden MG: Aerobic
glycolysis: Meeting the metabolic requirements of cell
proliferation. Annu Rev Cell Dev Biol. 27:441–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Brault C, Zerbib Y, Delette C, Marc J,
Gruson B, Marolleau JP and Maizel J: The warburg effect as a type B
lactic acidosis in a patient with acute myeloid leukemia: A
diagnostic challenge for clinicians. Front Oncol. 8:2322018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qu Y, Xiao H, Xiao W, Xiong Z, Hu W, Gao
Y, Ru Z, Wang C, Bao L, Wang K, et al: Upregulation of MIAT
regulates LOXL2 expression by competitively binding MiR-29c in
clear cell renal cell carcinoma. Cell Physiol Biochem.
48:1075–1087. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zaffryar-Eilot S, Marshall D, Voloshin T,
Bar-Zion A, Spangler R, Kessler O, Ghermazien H, Brekhman V,
Suss-Toby E, Adam D, et al: Lysyl oxidase-like-2 promotes tumour
angiogenesis and is a potential therapeutic target in angiogenic
tumours. Carcinogenesis. 34:2370–2379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cheng L, Yu H, Yan N, Lai K and Xiang M:
Hypoxia-inducible factor-1α target genes contribute to retinal
neuroprotection. Front Cell Neurosci. 11:202017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shi L, He C, Li Z, Wang Z and Zhang Q:
FBP1 modulates cell metabolism of breast cancer cells by inhibiting
the expression of HIF-1α. Neoplasma. 64:535–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee K and Kim HM: A novel approach to
cancer therapy using PX-478 as a HIF-1α inhibitor. Arch Pharm Res.
34:1583–1585. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Barone C, Koeberle D, Metselaar H, Parisi
G, Sansonno D and Spinzi G: Multidisciplinary approach for HCC
patients: Hepatology for the oncologists. Ann Oncol. 24 (Suppl
2):ii15–ii23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ahn SG, Dong SM, Oshima A, Kim WH, Lee HM,
Lee SA, Kwon SH, Lee JH, Lee JM, Jeong J, et al: LOXL2 expression
is associated with invasiveness and negatively influences survival
in breast cancer patients. Breast Cancer Res Treat. 141:89–99.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bais MV and Goldring MB: LOXL2 as a
protective in osteoarthritis cartilage. Aging (Albany NY).
9:2024–2025. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Atala A: Re: Tumour-suppressive
MicroRNA-29s directly regulate LOXL2 expression and inhibit cancer
cell migration and invasion in renal cell carcinoma. J Urol.
195:16222016. View Article : Google Scholar
|
|
52
|
Ye Y, Song YN, He SF, Zhuang JH, Wang GY
and Xia W: GINS2 promotes cell proliferation and inhibits cell
apoptosis in thyroid cancer by regulating CITED2 and LOXL2. Cancer
Gene Ther. 26:103–113. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Park JS, Lee JH, Lee YS, Kim JK, Dong SM
and Yoon DS: Emerging role of LOXL2 in the promotion of pancreas
cancer metastasis. Oncotarget. 7:42539–42552. 2016.PubMed/NCBI
|
|
54
|
Wu L, Zhang Y, Zhu Y, Cong Q, Xiang Y and
Fu L: The effect of LOXL2 in hepatocellular carcinoma. Mol Med Rep.
14:1923–1932. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Barker HE, Chang J, Cox TR, Lang G, Bird
D, Nicolau M, Evans HR, Gartland A and Erler JT: LOXL2-mediated
matrix remodeling in metastasis and mammary gland involution.
Cancer Res. 71:1561–1572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cuevas EP, Moreno-Bueno G, Canesin G,
Santos V, Portillo F and Cano A: LOXL2 catalytically inactive
mutants mediate epithelial-to-mesenchymal transition. Biol Open.
3:129–137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu X, Wang X, Zhang J, Lam EK, Shin VY,
Cheng AS, Yu J, Chan FK, Sung JJ and Jin HC: Warburg effect
revisited: An epigenetic link between glycolysis and gastric
carcinogenesis. Oncogene. 29:442–450. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wong CC, Tse AP, Huang YP, Zhu YT, Chiu
DK, Lai RK, Au SL, Kai AK, Lee JM, Wei LL, et al: Lysyl
oxidase-like 2 is critical to tumor microenvironment and metastatic
niche formation in hepatocellular carcinoma. Hepatology.
60:1645–1658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wang M, Zhao X, Zhu D, Liu T, Liang X, Liu
F, Zhang Y, Dong X and Sun B: HIF-1α promoted vasculogenic mimicry
formation in hepatocellular carcinoma through LOXL2 up-regulation
in hypoxic tumor microenvironment. J Exp Clin Cancer Res.
36:602017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ninomiya G, Yamada S, Hayashi M, Takeda S,
Suenaga M, Takami H, Kanda M, Iwata N, Niwa Y, Tanaka C, et al:
Significance of Lysyl oxidaselike 2 gene expression on the
epithelial-mesenchymal status of hepatocellular carcinoma. Oncol
Rep. 39:2664–2672. 2018.PubMed/NCBI
|
|
61
|
Yang MH, Chen CL, Chau GY, Chiou SH, Su
CW, Chou TY, Peng WL and Wu JC: Comprehensive analysis of the
independent effect of twist and snail in promoting metastasis of
hepatocellular carcinoma. Hepatology. 50:1464–1474. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wu TJ, Chang SS, Li CW, Hsu YH, Chen TC,
Lee WC, Yeh CT and Hung MC: Severe hepatitis promotes
hepatocellular carcinoma recurrence via NF-κB pathway-mediated
epithelial-mesenchymal transition after resection. Clin Cancer Res.
22:1800–1812. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hirata H, Sugimachi K, Komatsu H, Ueda M,
Masuda T, Uchi R, Sakimura S, Nambara S, Saito T, Shinden Y, et al:
Decreased expression of Fructose-1,6-bisphosphatase associates with
glucose metabolism and tumor progression in hepatocellular
carcinoma. Cancer Res. 76:3265–3276. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yang J, Jin X, Yan Y, Shao Y, Pan Y,
Roberts LR, Zhang J, Huang H and Jiang J: Inhibiting histone
deacetylases suppresses glucose metabolism and hepatocellular
carcinoma growth by restoring FBP1 expression. Sci Rep.
7:438642017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang J, Wang C, Zhao F, Luo X, Qin M,
Arunachalam E, Ge Z, Wang N, Deng X, Jin G, et al: Loss of FBP1
facilitates aggressive features of hepatocellular carcinoma cells
through the Warburg effect. Carcinogenesis. 38:134–143.
2017.PubMed/NCBI
|
|
66
|
Jiang H, Zhou Z, Jin S, Xu K, Zhang H and
Xu J, Sun Q, Wang J and Xu J: PRMT9 promotes hepatocellular
carcinoma invasion and metastasis via activating
PI3K/Akt/GSK-3β/Snail signaling. Cancer Sci. 109:1414–1427. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu GM, Li Q, Zhang PF, Shen SL, Xie WX,
Chen B, Wu J, Hu WJ, Huang XY and Peng BG: Restoration of FBP1
suppressed Snail-induced epithelial to mesenchymal transition in
hepatocellular carcinoma. Cell Death Dis. 9:11322018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen Z, Zuo X, Zhang Y, Han G, Zhang L, Wu
J and Wang X: MiR-3662 suppresses hepatocellular carcinoma growth
through inhibition of HIF-1α-mediated Warburg effect. Cell Death
Dis. 9:5492018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Courtnay R, Ngo DC, Malik N, Ververis K,
Tortorella SM and Karagiannis TC: Cancer metabolism and the Warburg
effect: The role of HIF-1 and PI3K. Mol Biol Rep. 42:841–851. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Takano S, Ishikawa E, Matsuda M, Sakamoto
N, Akutsu H, Yamamoto T and Matsumura A: The anti-angiogenic role
of soluble-form VEGF receptor in malignant gliomas. Int J Oncol.
50:515–524. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhu AX, Duda DG, Sahani DV and Jain RK:
HCC and angiogenesis: Possible targets and future directions. Nat
Rev Clin Oncol. 8:292–301. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yang ZF and Poon RT: Vascular changes in
hepatocellular carcinoma. Anat Rec (Hoboken). 291:721–734. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Berretta M, Rinaldi L, Di Benedetto F,
Lleshi A, De Re V, Facchini G, De Paoli P and Di Francia R:
Angiogenesis inhibitors for the treatment of hepatocellular
carcinoma. Front Pharmacol. 7:4282016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mossenta M, Busato D, Baboci L, Cintio FD,
Toffoli G and Bo MD: New insight into therapies targeting
angiogenesis in hepatocellular carcinoma. Cancers (Basel). 11(pii):
E10862019. View Article : Google Scholar : PubMed/NCBI
|