Introduction
Lung cancer, which originates from the bronchial
mucosa or glands, is a malignant tumor with a high mortality rate.
Approximately 27% of all tumor-related deaths are caused by lung
cancer (1,2). Non-small cell lung cancer (NSCLC)
accounts for the majority (i.e., 85 to 90%) of all lung cancer
cases, where the incidence of lung adenocarcinoma has significantly
increased to a greater extent than that of squamous cell carcinoma.
In addition, it has become the most common type of lung cancer in
many countries (3). With the
development of medical technology, multiple chemotherapy drugs,
such as platinum/pemetrexed (4),
gefitinib (5), paclitaxel (6) and cisplatin (7), have been widely applied to treat lung
cancer including lung adenocarcinoma. As an effective cytotoxic
agent, cisplatin is a common first-line drug that is usually
combined with other agents for the treatment of lung
adenocarcinoma, but its efficacy is largely impaired by increasing
chemoresistance (8). Therefore, it
is necessary to further explore the underlying mechanisms of
cisplatin chemoresistance in lung adenocarcinoma, which in turn
facilitates the designing of treatment strategies for patients with
lung adenocarcinoma.
Surgical resection is still the preferred treatment
for patients with early-stage lung adenocarcinoma, but the 5-year
survival rate of patients with stage I NSCLC is only 45–65%
(9). Local recurrence occurs in
approximately 6–11% of patients with stage I NSCLC, while distant
metastasis accounts for approximately 23–30% of recurrence
(10). Recurrence and metastasis
are important factors in the poor prognosis of patients, and the
presence of circulating tumor cells (CTCs) exacerbates these risks.
CTCs were first reported by Thomas Ashworth (1869), and are defined
as a set of cells that shed from the primary tumor and invade into
the blood stream or surrounding parenchyma to form new tumors
(11). Several studies have
revealed that CTCs present dynamic changes in epithelial and
mesenchymal phenotypes (12,13)
and an increase in the number of CTCs is usually correlated with
tumor relapse and chemotherapy/radiotherapy resistance (14–16).
At present, CTCs have been used as tumor biomarkers in various
tumors including breast cancer, colorectal cancer and prostate
cancer (17). However, the function
and regulatory mechanisms related to CTCs in lung adenocarcinoma
are rarely reported. MicroRNAs (miRNAs) are a class of small
non-coding RNAs, with a length of 20–24 nt. Previous studies have
demonstrated that several miRNAs, including miR-216a/miR-217
(18), miR-497 (19), the miR-200bc/429 cluster (20) and miR-31, are correlated with drug
resistance and recurrence of cancer (18). In addition, miR-206 has been
reported to regulate cisplatin resistance and
epithelial-to-mesenchymal transition (EMT) in human lung
adenocarcinoma cells partly through targeting MET (21). miR-15b was found to regulate
cisplatin resistance and metastasis via targeting PEBP4 in human
lung adenocarcinoma cells (22).
Kitamura et al found that the miR-134/487b/655 cluster could
regulate TGF-β-induced EMT and drug resistance to gefitinib by
targeting MAGI2 in lung adenocarcinoma cells (23). miR-10a is a member of the highly
conserved miR-10 family, and has been confirmed to be involved in
drug resistance in several types of cancers. For example, miR-10a,
miR-195, and miR-455-3p are correlated with temozolomide resistance
in glioblastoma multiforme cells (24). Bao et al reported that serum
miR-10a-5p and miR-196a-5p could serve as non-invasive biomarkers
for NSCLC patients (25). Long
non-coding RNA RP11-838N2.4 was found to increase the effect of
temozolomide in glioblastoma cell lines via suppressing the
function of miR-10a (26).
Recently, Sun et al revealed that miR-10a knockdown could
increase the cisplatin sensitivity of A549 cells via the
TGF-β/Smad/STATA3 signaling pathway (27). However, the function of miR-10a in
the resistance of CTCs in lung adenocarcinoma to cisplatin has not
been elucidated to date.
Therefore, in the present study, CTCs were isolated
from the blood samples of lung adenocarcinoma patients to reveal
the function of miR-10a in cisplatin resistance by when comparing
with this function demonstrated in A549 and H1299 adenocarcinoma
cells. In addition, the downstream target of miR-10a and the
underlying signaling pathway involved in this progression were also
determined. Based on these results, we hope to provide new
information for understanding the cisplatin resistance of lung
adenocarcinoma and new insights in the treatment of lung
adenocarcinoma.
Materials and methods
Patients and sampling
Blood samples from 6 lung adenocarcinoma patients at
the Shenzhen People's Hospital were collected from May to December
2017. The patients enrolled in this study underwent pathological
diagnosis and radiography to confirm the presence of lung
adenocarcinoma, and all underwent chemotherapy. All patients signed
informed consent forms, and this study was authorized by the Ethics
Committee of Shenzhen People's Hospital, The Second Clinical
Medical College of Jinan University (Shenzhen, China).
Isolation of CTCs
Isolation of human CTCs was performed as previously
reported by Gong et al (28). Briefly, 20–50 ml fresh blood was
obtained from lung adenocarcinoma patients and layered over
Ficoll-Paque at a density of 1.077 g, and centrifuged at 400 × g
for 30 min at room temperature. The cells in the interphase,
including lymphocytes, monocytes, and tumor cells, were then
resuspended in 300 µl of solution (consisting of 100 µl each of FcR
blocking reagent, CD45 microbeads, and CD15 microbeads; purchased
from Miltenyi Biotec; Germany) at a density of 5×107
cells. Following this, CD45 and CD15 microbeads were removed using
the magnetic auto MACS™ Pro Separator (Miltenyi Biotec). Then, 100
µl of CD326 (Miltenyi Biotec) per 5×107 cells was added
and incubated at 4°C for 30 min. Subsequently, the magnetic
CD326+ and CD326− cells were eluted as
epithelial cell adhesion molecule (EpCAM)+ and
EpCAM− CTCs. Purification of epithelial cells was
performed by immunofluorescent staining with anti-cytokeratin
antibody (product code ab756; Abcam) to reach up to 95% confluence.
CTCs were identified when they presented positive staining by a
tumor-specific marker cytokeratin and positive scoring based on the
review of a cytopathologist.
Cell culture
Human lung adenocarcinoma cell lines A549 and H1299
were purchased from the Chinese Academy of Sciences Committee on
Type Culture Collection Cell Bank (Shanghai, China) and maintained
in Dulbeccos modified Eagles medium (DMEM, Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS, Gibco;
Thermo Fisher Scientific, Inc.). CTCs were maintained in RPMI-1640
medium supplemented with 20 ng/ml basic FGF (Life Technology;
Thermo Fisher Scientific, Inc.), 20 mg/ml EGF (Life Technology;
Thermo Fisher Scientific, Inc.), 10 ml B27 (Life Technology; Thermo
Fisher Scientific, Inc.), and 1X antibiotic/antimycotic (Life
Technology; Thermo Fisher Scientific, Inc.). All cells were
maintained in a humidified incubator at 37°C with 5% CO2
in atmosphere.
Cell transfection
A549 and H1299 cells were seeded in 6-well plates at
a confluency of 40–60% overnight. Then, 50 nm of miR-10a mimic
(Guangzhou RiboBio Co., Ltd.) was transfected into A549 and H1299
cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturers instructions. In
addition, cells transfected with the negative control (NC) sequence
served as the negative control. After that, the cells were
harvested after transfection for 48 h and used for the following
confirmation and investigation. For pcDNA3.1-PIK3CA, PIK3CA shRNA
and its corresponding scramble sequences (Guangzhou RiboBio Co.,
Ltd.) were transfected into A549 and H1299 cells using
Lipofectamine 2000 according to the manufacturers instructions.
After transfection for 48 h, the cells were harvested for further
validation and investigations.
CCK-8 assay
The half maximal inhibitory concentration
(IC50) of each cell line to cisplatin was evaluated by
CCK-8 assay. Briefly, the cells were seeded into 96-well plates at
a density of 5×103 cells per well for overnight. The
cells were then treated with different concentrations of cisplatin
(0, 0.05, 0.1, 0.5, 1.0, 5.0, 10.0 and 50.0 µg/ml) for 24 h.
Following this, 10 µl of CCK-8 solution (Boster) was added into
each well and incubated for 4 h at 37°C. Subsequently, the
absorbance of each well was determined using a microplate reader
(BioTek ELx800; BioTek Instruments, Inc.) at 450 nm. Each sample
was designed with five repeats and each experiment was performed at
least three times.
Luciferase activity assay
A549 cells were seeded in 6-well plates overnight
and co-transfected with miR-10 mimic/NC and
psiCHECK2-PIK3CA-3′-UTR-WT (WT)/psiCHECK2-PIK3CA-3′-UTR-MUT (MUT)
using Lipofectamine 2000 (Invitrogen) according to the
manufacturers protocol. After 24 h of transfection, luciferase
activity was measured using dual-luciferase reporter assay
(Promega) according to the manufacturers instructions and Firefly
luminescence was normalized to Renilla luminescence.
Plasmids for this assay were purchased from Guangzhou RiboBio Co.,
Ltd.
Quantitative real-time PCR
(RT-qPCR)
After transfection or treatment, total RNA in the
cells was isolated using TRIzol reagent (Takara). Then, RNA was
reverse transcribed into cDNA using Bestar qPCR RT Kit (DBI, China)
according to the manufacturers protocol. Using cDNA as a template,
amplification of genes was performed using DBI Bestar®
Sybr Green qPCR Master Mix (DBI) according to the manufacturers
instructions on an Agilent Stratagene Mx2000P PCR instrument
(Agilent Technologies) under the following conditions: 95°C for 2
min and 40 cycles of 94°C for 20 sec, 58°C for 20 sec, and 72°C for
20 sec. Primers of genes were designed as follows: miR-10a,
forward, 5′-ACACTCCAGCTGGGTACCCTGTAGATCCGAAT-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACAAATTC-3′; U6, forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
Subsequently, the relative gene expression was analyzed using the
2−ΔΔCq method by normalizing to U6 (29).
Western blotting
After transfection or treatment, the cells were
lysed using RIPA lysis buffer (Pierce; Thermo Fisher Scientific,
Inc.) supplemented with a Protease Inhibitor Cocktail (Pierce;
Thermo Fisher Scientific, Inc.). Then, the protein concentration of
the lysates was determined using the BCA method (Pierce; Thermo
Fisher Scientific, Inc.). Following this, the protein solution was
boiled with equivalent amounts of loading buffer for 10 min, equal
amounts of protein (30 µg) were subjected to 8–12% SDS-PAGE gel
electrophoresis, and then transferred onto a PVDF membrane
(Millipore). Membranes were blocked with 5% non-fat milk in
Tris-buffered saline and incubated with anti-PIK3CA (product code
ab40776; dilution 1:1,000), PI3K (product code ab32089; dilution
1:1,000), Akt (product code ab179463; dilution 1:1,000),
phosphorylated (p)Akt (product code ab81283; dilution 1:5,000), or
GAPDH (product code ab181602; dilution 1:5,000; Abcam) antibodies
at 4°C overnight followed by horseradish peroxidase-conjugated
secondary antibody (product code ab205718; dilution 1:10,000;
Abcam). Subsequently, the protein bands in the membranes were
visualized using the ECL method (Pierce; Thermo Fisher Scientific,
Inc.) The gel optical processing system (Image-Pro Plus 6.0; Media
Cybernetics) was used to analyze the net optical density of the
bands.
Statistical analyses
All statistical analyses were performed using SPSS
16.0 (SPSS lnc.) and GraphPad Prism 6.0 (GraphPad Software). All
experiments were performed in triplicate and the average values
were calculated and are presented as mean ± standard deviation.
Comparisons between two groups were estimated using unpaired t-test
or Mann-Whitney U test. P<0.05 was assigned to indicate
statistical significance.
Results
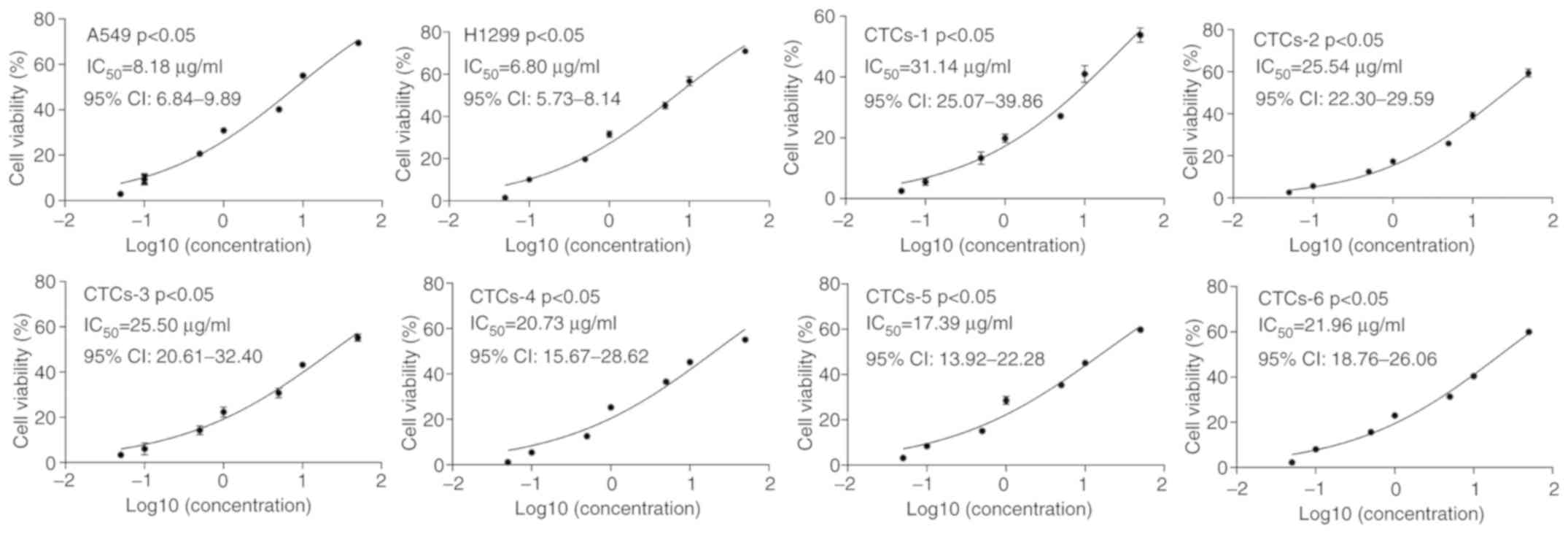
CTCs present a lower sensitivity to
cisplatin than the A549 and H1299 cell lines
A total of 6 lung adenocarcinoma patients were
enrolled in this study, including two males and four females
average aged is 65.3 years. Their clinical stages ranged from stage
II to stage IV. The clinical characteristics of the 6
adenocarcinoma patients are summarized in Table I. CTCs were isolated from 6
adenocarcinoma patients, and A549 and H1299 cells were treated with
different concentrations of cisplatin to determine the
proliferation of each cell line using CCK-8 assay. The results
presented that the IC50 values of the CTCs from 6
patients (IC50=31.14, 25.54, 25.50, 20.73, 17.39, and
21.96, respectively) were significantly higher than that of the
A549 (IC50=8.184) and H1299 cells (IC50=6.80)
(P<0.05) (Fig. 1). These
findings indicated that CTCs may have a lower sensitivity to
cisplatin than that of carcinoma cells in lung cancer tissues.
 | Table I.General characteristics of the 6
patients with lung adenocarcinoma from whom CTCs were
collected. |
Table I.
General characteristics of the 6
patients with lung adenocarcinoma from whom CTCs were
collected.
|
| Patients |
|---|
|
|
|
|---|
|
Characteristics | #1 | #2 | #3 | #4 | #5 | #6 |
|---|
| Sex | F | M | F | F | M | F |
| Age (years) | 64 | 63 | 66 | 64 | 71 | 64 |
| Tumor size
(mm) | 13 | 8 | 22 | 16 | 24 | 15 |
| Node status | N1 | N1 | N2 | N1 | N2 | N1 |
| Metastasis | M0 | M0 | M1 | M0 | M0 | M0 |
| Clinical stage | II | II | IV | II | III | II |
| CTC counts | 8 | 10 | 55 | 5 | 25 | 15 |
| Date of
surgery | 2017/5/13 | 2017/6/7 | 2017/8/18 | 2017/9/15 | 2017/11/1 | 2017/11/21 |
Expression of miR-10a and
PIK3CA/PI3K/Akt signaling is altered after treatment with
cisplatin
Furthermore, to reveal the underlying mechanism
underlying the cisplatin resistance of CTCs, the expression of
miR-10a and phosphatidylinositol-4,5-bisphosphate 3-kinase
catalytic subunit α (PIK3CA) were detected in CTCs, A549 and H1299
cells after treatment with cisplatin. RT-qPCR revealed that the
expression of miR-10a in CTCs was significantly higher than that in
A549 and H1299 cells after treatment with cisplatin (P<0.01,
Fig. 2A). Moreover, as determined
by western blot analysis, the CTCs exhibited a lower expression of
PIK3CA and phosphoinositide 3-kinase (PI3K) than these levels in
the A549 and H1299 cells after cisplatin treatment (Figs. 2B and S1). Meanwhile, protein kinase B
phosphorylation (pAkt) showed similar results as PIK3CA and PI3K,
but no obvious difference was observed in Akt expression (Fig. 2B). These findings suggest that
modulation of miR-10a expression and PIK3CA in the PI3K/Akt
signaling pathway increases the resistance of CTCs to
cisplatin.
PIK3CA is a target of miR-10a
According to miRWalk database, PIK3CA was
predicted to be a potential target of miR-10a (http://www.targetscan.org/cgibin/targetscan/vert_71/view_gene.cgi?rs=ENST000
00263967.3&taxid=9606&showcnc=0&shownc=0&shownc_nc=&showncf1=&showncf2=&subset=1#miR-10-5p,
Fig. 3A). Then, the regulatory
relationship between miR-10a and PIK3CA was explored using
luciferase reporter assay in A549 cells. The results showed that
the overexpression of miR-10a significantly decreased the activity
of WT-PIK3CA-3UTR (WT+miR-10a), but not the MUT-PIK3CA-3UTR
(MUT+miR-10a) (Fig. 3B), indicating
that PIK3CA is a direct target gene of miR-10a. Finally,
A549 cells were transfected with the miR-10a inhibitor, and
expression of PIK3CA was detected by western blotting and real-time
PCR, respectively. The results showed that miR-10a inhibitor
increased both the protein and mRNA levels of PIK3CA (Fig. 3C and D).
Overexpression of miR-10a or silencing
of PIK3CA decreases the sensitivity to cisplatin
Next, the miR-10a mimic or shPIK3CA was transfected
into A549 and H1299 cells and the sensitivity to cisplatin was
determined using CCK-8 assay. The results showed that
overexpression of miR-10a (IC50=18.56) and knockout of
PIK3CA (shPIK3CA) (IC50=23.77) significantly
increased the IC50 value of A549 cells to cisplatin when
compared with the blank control (IC50=8.184) and NC
(IC50=8.366, Fig. 4A).
In addition, overexpression of miR-10a (IC50=10.14) and
silencing of PIK3CA (shPIK3CA) (IC50=12.20)
significantly increased the IC50 value of H1299 to
cisplatin compared with the blank control (IC50=6.86)
and NC (IC50=6.56, Fig.
4B). Moreover, western blotting showed that both miR-10a mimic
and shPIK3CA significantly reduced PIK3CA, PI3K and pAkt levels in
both the A549 and H1299 cell lines but not the Akt expression level
(Fig. 4C-4F). These findings
revealed that miR-10a increased the resistance abilities of A549
and H1299 cells to cisplatin by decreasing the expression of PIK3CA
in the PI3K/Akt signaling pathway.
Overexpression of PIK3CA increases the
sensitivities of A549 and H1299 to cisplatin
Moreover, PIK3CA was also co-transfected with
miR-10a mimic into A549 and H1299 cells to explore the effect of
the miR-10a/PIK3CA regulatory relationship on sensitivity of lung
carcinoma to cisplatin. The results showed that overexpression of
miR-10a significantly decreased PIK3CA expression, while
overexpression of PIK3CA obviously reversed this effect in both
A549 and H1299 cells (Fig. 5A and
B). In addition, overexpression of PIK3CA significantly
increased the sensitivities of A549 (IC50: 11.69 vs.
18.56) (Fig. 5C) and H1299
(IC50: 7.71 vs. 10.14) (Fig.
5D) cells to cisplatin after transfecting with miR-10 mimic.
These findings indicated that overexpression of PIK3CA increased
the sensitivities of A549 and H1299 cells to cisplatin.
Discussion
Recurrence and metastasis of lung cancer are
considered as factors of poor prognosis and cancer-related
mortality. The presence of circulating tumor cells (CTCs) increases
the risk of recurrence and metastasis in patients with lung cancer.
Therefore, we isolated CTCs from lung adenocarcinoma blood samples
and treated them with cisplatin to detect the resistance of CTCs to
cisplatin. Consistent with the increased risk of lung cancer
recurrence induced by CTCs, we also revealed that the CTCs
presented a lower sensitivity to cisplatin than A549 and H1299
cells. These results indicated that the cisplatin resistance of
CTCs further lead to the poor prognosis of lung cancer
patients.
miRNAs have been widely acknowledged as important
regulators of human cancer by serving as oncogenes or tumor
suppressors (30). miR-10a is a
member of the miR-10a family, and has been found to be involved in
multiple malignant transformations, including those of breast
cancer (31), glioma (32), liver cancer (33) and pancreatic cancer (34). For instance, Bryant et al
revealed that miR-10a is abnormally overexpressed in nucleophomin
1-mutated acute myeloid leukemia (AML) and is positively correlated
with AML cell survival (35).
Importantly, Yu et al also revealed that miR-10a is elevated
in non-small cell lung cancer (NSCLC) and is positively correlated
with the progression of NSCLC via targeting PTEN (36). In addition, Sun et al
demonstrated that downregulation of miR-10a expression decreased
cisplatin resistance in A549 cells via the TGF-β/smad2/STAT3/STAT5
signaling pathway (27). In the
present study, miR-10a expression was found to be highly expressed
in CTCs when compared to that in A549 and H1299 cells, meaning that
the upregulation of miR-10a expression in CTCs may enhance
cisplatin resistance of CTCs leading to poor patient prognosis.
Phosphatidylinositol-4,5-bisphosphate 3-kinase
catalytic subunit α (PIK3CA) is a p110α subunit of PI3K and often
plays a critical role in regulating the activation of the
PI3K/Akt/mTOR signaling pathway. The PI3K/Akt/mTOR pathway is
closely related with tumorigenesis in breast cancer and
hepatocellular carcinoma by regulating proliferation, migration,
metastasis and apoptosis of tumor cells (37–39).
Dysregulated PIK3CA expression was found to result in the abnormal
activation of the PI3K/Akt pathway and finally in the development
of cancer. Herein, we confirmed that PIK3CA expression in CTCs was
lower than that in A549 and H1299 cells after treatment with
cisplatin, indicating that the malignant biological behavior of
CTCs was more significant than that of lung adenocarcinoma cell
lines. Hu et al documented that miR-10a could modulate the
proliferation of airway smooth muscle cells via directly targeting
PIK3CA in the PI3K pathway (40).
Moreover, miR-10a was found to suppress the progression of breast
cancer via targeting the PIK3CA in the PI3K/Akt/mTOR signaling
pathway (41). In addition, PIK3CA
was confirmed as a direct target of miR-10a and further revealed
that miR-10a overexpression obviously decreased the expression of
PIK3CA and PI3K and the phosphorylation of Akt. Moreover,
upregulation of PIK3CA largely reversed the effect of miR-10a on
the proliferation of A549 and H1299 cells by reducing the
expression of PI3K and phosphorylation of Akt. Taken together, this
evidence suggests that miR-10a directly regulates the PI3K/Akt
signaling pathway via modulating the expression of PIK3CA.
In addition to the regulation of the occurrence and
development of multiple cancers, the PI3K/Akt signaling pathway is
also closely related to cisplatin resistance, whereby this process
includes the participation of many miRNAs. miR-221 has been
reported to induce cell survival and cisplatin resistance to human
osteosarcoma via the PI3K/Akt signaling pathway (42). Downregulation of miR-497 was found
to contribute to the cell growth and cisplatin resistance via the
PI3K/Akt signaling pathway (43).
Moreover, Klotho was found to increase lung cancer sensitivity to
cisplatin through the PI3K/Akt signaling pathway (44). In the present study, PI3K and
phosphorylation of Akt was obviously downregulated, and miR-10a was
highly expressed in CTCs when compared with A549 and H1299 cells,
indicating that CTCs present a lower sensitivity to cisplatin than
A549 and H1299 cells through downregulation of the PI3K/Akt pathway
under cisplatin treatment.
There were some limitations to the present study.
First, due to the limited sample size, there may have been some
bias between this study and the actual situation. Second, although
we confirmed that the miR-10a/PIK3CA axis is involved in the
pathogenesis of cisplatin resistance of lung cancer, the exact
upstream of miR-10a remains unclear. Despite these limitations, our
findings provide new information regarding the mechanism of lung
cancer resistance to cisplatin.
In conclusion, miR-10a was significantly upregulated
in CTCs, exhibiting higher cisplatin resistance than A549 and H1299
cells. Overexpression of miR-10a enhanced the cisplatin resistance
of A549 and H1299 and overexpression of PIK3CA largely reversed
this effect via the PI3K/Akt signaling pathway. These findings
suggest that miR-10a promotes the cisplatin resistance of CTCs by
downregulating PIK3CA expression in the PI3K/Akt signaling
pathway.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the Shenzhen Healthcare
Research Project (no. 201501011) and Guangdong Medical Research
Foundation (no. A2016475).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
Conception and design of the study were carried out
by TH and KR. Administrative and experimental support were carried
out by TH and GD. Provision of study materials and recruitment of
patients and samples were achieved by KR and LY. Collection and
assembly of data were carried out by LY and YW. Data analysis and
interpretation were performed by BP, GW and ZW. Writing of the
manuscript was conducted by all of the authors. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
All patients signed informed consent forms, and this
study was authorized by the Ethics Committee of Shenzhen People's
Hospital, The Second Clinical Medical College of Jinan University
(Shenzhen, Guangdong, China). This study was conducted in
accordance with the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sorber L, Zwaenepoel K, Deschoolmeester V,
Van Schil PE, Van Meerbeeck J, Lardon F, Rolfo C and Pauwels P:
Circulating cell-free nucleic acids and platelets as a liquid
biopsy in the provision of personalized therapy for lung cancer
patients. Lung Cancer. 107:100–107. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gan Y, Zhou P and Li W: Correlations of
Tumor Stage and Serum Tumor Markers with Age in Nonsmoking Females
with Lung Adenocarcinoma. D62 types, Genotypes, and Phenotypes: The
Three Ts of thoracic oncology. American Thoracic Society; pp.
ppA73342018
|
|
4
|
Zhai X, Zheng Q, Yang L, Zhu Y, Li J, Liu
Y and Wang Y: Impact of platinum/pemetrexed combination versus
other platinum-based regimens on adjuvant chemotherapy in resected
lung adenocarcinoma. Sci Rep. 7:14532017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jin B, Niu Y, Zhang Y, Chu T, Gu A, Wu J,
Pei J, Zhu L and Han B: 1281Pcombination of chemotherapy and
gefitinib as first-line treatment of patients with advanced lung
adenocarcinoma and sensitive egfr mutations: A randomised
controlled trial. Ann Oncol. 25 (4 Suppl):iv4512014. View Article : Google Scholar
|
|
6
|
Xu R, Mao Y, Chen K, He W, Shi W and Han
Y: The long noncoding RNA ANRIL acts as an oncogene and contributes
to paclitaxel resistance of lung adenocarcinoma A549 cells.
Oncotarget. 8:39177–39184. 2017.PubMed/NCBI
|
|
7
|
Sato C, Okuda K, Tamiya H, Yamamoto K,
Hoshina K, Narumoto O, Urushiyama H, Noguchi S, Amano Y, Watanabe
K, et al: Acute arterial thrombosis during postoperative adjuvant
cisplatin-based chemotherapy for completely resected lung
adenocarcinoma. Intern Med. 57:557–561. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Q, Cheng N, Li X, Pan H, Li C, Ren S,
Su C, Cai W, Zhao C, Zhang L and Zhou C: Correlation of long
non-coding RNA H19 expression with cisplatin-resistance and
clinical outcome in lung adenocarcinoma. Oncotarget. 8:2558–2567.
2017.PubMed/NCBI
|
|
9
|
Raz DJ, Zell JA, Ou SH, Gandara DR,
Anton-Culver H and Jablons DM: Natural history of stage Inon-small
cell lung cancer:Implications for early detection. Chest.
132:193–199. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Roselli M, Mariotti S, Ferroni P, Laudisi
A, Mineo D, Pompeo E, Ambrogi V and Mineo TC: Postsurgical
chemotherapy in stage IB non-small cell lung cancer: Long-term
survival in a randomized study. Int J Cancer. 119:955–960. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
García SA, Weitz J and Schölch S:
Circulating tumor cells. Cancer stem cells: Methods and protocols.
Papaccio G and Desiderio V: Springer; New York, NY: pp. 213–219.
2018, View Article : Google Scholar
|
|
12
|
Yu M, Bardia A, Wittner BS, Stott SL, Smas
ME, Ting DT, Isakoff SJ, Ciciliano JC, Wells MN, Shah AM, et al:
Circulating breast tumor cells exhibit dynamic changes in
epithelial and mesenchymal composition. Science. 339:580–584. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kallergi G, Papadaki MA, Politaki E,
Mavroudis D, Georgoulias V and Agelaki S: Epithelial to mesenchymal
transition markers expressed in circulating tumour cells of early
and metastatic breast cancer patients. Breast Cancer Res.
13:R592011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giuliano M, Giordano A, Jackson S, Hess
KR, De Giorgi U, Mego M, Handy BC, Ueno NT, Alvarez RH and De
Laurentiis M: Circulating tumor cells as prognostic and predictive
markers in metastatic breast cancer patients receiving first-line
systemic treatment. Breast Cancer Res. 13:R672011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pierga JY, Hajage D, Bachelot T, Delaloge
S, Brain E, Campone M, Diéras V, Rolland E, Mignot L, Mathiot C and
Bidard FC: High independent prognostic and predictive value of
circulating tumor cells compared with serum tumor markers in a
large prospective trial in first-line chemotherapy for metastatic
breast cancer patients. Ann Oncol. 23:618–624. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yokobori T, Iinuma H, Shimamura T, Imoto
S, Sugimachi K, Ishii H, Iwatsuki M, Ota D, Ohkuma M, Iwaya T, et
al: Plastin3 is a novel marker for circulating tumor cells
undergoing the epithelial-mesenchymal transition and is associated
with colorectal cancer prognosis. Cancer Res. 73:2059–2069. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lianidou ES, Markou A and Strati A: The
role of CTCs as tumor biomarkers. Adv Exp Med Biol. 867:341–367.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xia H, Ooi LL and Hui KM:
MicroRNA-216a/217-induced epithelial-mesenchymal transition targets
PTEN and SMAD7 to promote drug resistance and recurrence of liver
cancer. Hepatology. 58:629–641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu W, Zhu D, Lu S, Wang T, Wang J, Jiang
B, Shu Y and Liu P: miR-497 modulates multidrug resistance of human
cancer cell lines by targeting BCL2. Med Oncol. 29:384–391. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu W, Xu H, Zhu D, Zhi H, Wang T, Wang J,
Jiang B, Shu Y and Liu P: miR-200bc/429 cluster modulates multidrug
resistance of human cancer cell lines by targeting BCL2 and XIAP.
Cancer Chemother Pharmacol. 69:723–731. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen QY JD, Wang J, Hu H, Tang X, Chen J,
Mou H and Lu W: miR-206 regulates cisplatin resistance and EMT in
human lung adenocarcinoma cells partly by targeting MET.
Oncotarget. 7:24510–24526. 2016.PubMed/NCBI
|
|
22
|
Zhao Z, Zhang L, Yao Q and Tao Z: miR-15b
regulates cisplatin resistance and metastasis by targeting PEBP4 in
human lung adenocarcinoma cells. Cancer Gene Ther. 22:108–114.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kitamura K, Seike M, Okano T, Matsuda K,
Miyanaga A, Mizutani H, Noro R, Minegishi Y, Kubota K and Gemma A:
MiR-134/487b/655 cluster regulates TGF-β-induced
epithelial-mesenchymal transition and drug resistance to gefitinib
by targeting MAGI2 in lung adenocarcinoma cells. Mol Cancer Ther.
13:444–453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ujifuku K, Mitsutake N, Takakura S,
Matsuse M, Saenko V, Suzuki K, Hayashi K, Matsuo T, Kamada K,
Nagata I and Yamashita S: miR-195, miR-455-3p and miR-10a(*) are
implicated in acquired temozolomide resistance in glioblastoma
multiforme cells. Cancer Lett. 296:241–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bao M, Pan S, Yang W, Chen S, Shan Y and
Shi H: Serum mir-10a-5p and mir-196a-5p as non-invasive biomarkers
in non-small cell lung cancer. Int J Clin Exp Pathol. 11:773–780.
2018.PubMed/NCBI
|
|
26
|
Liu Y, Xu N, Liu B, Huang Y, Zeng H, Yang
Z, He Z and Guo H: Long noncoding RNA RP11-838N2.4 enhances the
cytotoxic effects of temozolomide by inhibiting the functions of
miR-10a in glioblastoma cell lines. Oncotarget. 7:43835–43851.
2016.PubMed/NCBI
|
|
27
|
Sun W, Ma Y, Chen P and Wang D:
MicroRNA-10a silencing reverses cisplatin resistance in the
A549/cisplatin human lung cancer cell line via the transforming
growth factor-β/Smad2/STAT3/STAT5 pathway. Mol Med Rep.
11:3854–3859. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gong C, Liu B, Yao Y, Qu S, Luo W, Tan W,
Liu Q, Yao H, Zou L, Su F and Song E: Potentiated DNA damage
response in circulating breast tumor cells confers resistance to
chemotherapy. J Biol Chem. 290:14811–14825. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) methods. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Damavandi Z, Torkashvand S, Vasei M,
Soltani BM, Tavallaei M and Mowla SJ: Aberrant expression of breast
development-related MicroRNAs, miR-22, miR-132, and miR-212, in
breast tumor tissues. J Breast Cancer. 19:148–155. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yan Y, Wang Q, Yan XL, Zhang Y, Li W, Tang
F, Li X and Yang P: miR-10a controls glioma migration and invasion
through regulating epithelial-mesenchymal transition via EphA8.
FEBS Lett. 589:756–765. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Liu Z, Yao B, Dou C, Xu M, Xue Y,
Ding L, Jia Y, Zhang H and Li Q: Long non-coding RNA TUSC7 acts a
molecular sponge for miR-10a and suppresses EMT in hepatocellular
carcinoma. Tumour Biol. 37:11429–11441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weiss FU, Marques IJ, Woltering JM,
Vlecken DH, Aghdassi A, Partecke LI, Heidecke CD, Lerch MM and
Bagowski CP: Retinoic acid receptor antagonists inhibit miR-10a
expression and block metastatic behavior of pancreatic cancer.
Gastroenterology. 137:2136–2145.e1-7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bryant A, Palma CA, Jayaswal V, Yang YW,
Lutherborrow M and Ma DD: miR-10a is aberrantly overexpressed in
Nucleophosmin1 mutated acute myeloid leukaemia and its suppression
induces cell death. Mol Cancer. 11:82012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu T, Liu L, Li J, Yan M, Lin H, Liu Y,
Chu D, Tu H, Gu A and Yao M: MiRNA-10a is upregulated in NSCLC and
may promote cancer by targeting PTEN. Oncotarget. 6:30239–30250.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Janku F, Wheler JJ, Westin SN, Moulder SL,
Naing A, Tsimberidou AM, Fu S, Falchook GS, Hong DS, Garrido-Laguna
I, et al: PI3K/AKT/mTOR inhibitors in patients with breast and
gynecologic malignancies harboring PIK3CA mutations. J Clin Oncol.
30:777–782. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Woo SU, Sangai T, Akcakanat A, Chen H, Wei
C and Meric-Bernstam F: Vertical inhibition of the PI3K/Akt/mTOR
pathway is synergistic in breast cancer. Oncogenesis. 6:e3852017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chai R, Fu H, Zheng Z, Liu T, Ji S and Li
G: Resveratrol inhibits proliferation and migration through SIRT1
mediated post-translational modification of PI3K/AKT signaling in
hepatocellular carcinoma cells. Mol Med Rep. 16:8037–8044. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hu R, Pan W, Fedulov AV, Jester W, Jones
MR, Weiss ST, Panettieri RA Jr, Tantisira K and Lu Q: MicroRNA-10a
controls airway smooth muscle cell proliferation via direct
targeting of the PI3 kinase pathway. FASEB J. 28:2347–2357. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ke K and Lou T: MicroRNA-10a suppresses
breast cancer progression via PI3K/Akt/mTOR pathway. Oncol Lett.
14:5994–6000. 2017.PubMed/NCBI
|
|
42
|
Zhao G, Cai C, Yang T, Qiu X, Liao B, Li
W, Ji Z, Zhao J, Zhao H, Guo M, et al: MicroRNA-221 induces cell
survival and cisplatin resistance through PI3K/Akt pathway in human
osteosarcoma. PLoS One. 8:e539062013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shao XJ, Miao MH, Xue J, Xue J, Ji XQ and
Zhu H: The Down-regulation of MicroRNA-497 contributes to cell
growth and cisplatin resistance through PI3K/Akt pathway in
osteosarcoma. Cell Physiol Biochem. 36:2051–2062. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang Y, Chen L, Huang G, He D, He J, Xu W,
Zou C, Zong F, Li Y, Chen B, et al: Klotho sensitizes human lung
cancer cell line to cisplatin via PI3k/Akt pathway. PLoS One.
8:e573912013. View Article : Google Scholar : PubMed/NCBI
|