Introduction
Liver cancer is the fifth most common cancer type
and the second most common cause of cancer-related mortality
worldwide (1). Hepatocellular
carcinoma (HCC) accounts for ~90% of primary liver cancers, and its
incidence has been constantly increasing (2). In China, HCC has a higher incidence
and may account for 50% of new cases globally each year (3). The majority of HCC cases in China and
southeastern Asia are caused by hepatitis B virus (HBV) infection,
while non-alcoholic-associated steatosis may be one of the main
causes of the development of HCC in Western countries (4). In the early stages, liver
transplantation or hepatic resection is the standard treatment
approach, with a high probability of recurrence-free postoperative
course, while in advanced or metastatic stages, sorafenib (a broad
tyrosine kinase inhibitor) is typically used (5). HCC is an immunogenic tumor that
occurs in chronically inflamed livers (6). Therapeutic antibodies blocking the
programmed cell death protein-1 (PD-1)/programmed death ligand-1
(PD-L1) interaction were approved by the Food and Drug
Administration to treat advanced HCC in patients previously treated
with sorafenib as a second line treatment (7). However, the observed objective
response rate was not impressive (~20%) in response to PD-1/PD-L1
antibody treatment (8). Therefore,
the identification of novel immunotherapeutic targets in advanced
HCC is imperative.
The tumor microenvironment (TME) is widely used as a
biomarker for the identification of novel therapeutic targets. The
HCC TME consists of cancer and stromal cells, including hepatic
stellate cells, cancer-associated fibroblasts, immune cells and
endothelial cells, which interact with the tumor and affect its
growth (9). The role of
tumor-infiltrating B cells (B-TILs) remains controversial.
Shalapour et al (10)
indicated that immunoglobulin (Ig) A-expressing B cells could
elicit depletion of liver cytotoxic CD8+ T cells (CTLs)
and that IgA+ cell depletion could enhance the
regression of established non-alcoholic-associated
steatosis-induced HCC. Furthermore, it was also shown that a high
number of T cell Ig and mucin domain-1+ regulatory B
(Breg) cells could infiltrate into the tumors of patients with HCC
(11,12). These infiltrating Breg cells were
found to express IL-10 and demonstrate high suppressive activity
against T cells. Mechanistically, the CD40/CD154 signaling pathway
was found to be involved in HCC progression via the activation of
Breg cells (13,14). By contrast, Zhang et al
(15) demonstrated that B cell
infiltration into tumors was significantly impaired during tumor
progression, whereas high density of B-TILs correlated with
improved clinical outcome. Moreover, the expression levels of CD19
and CD38 were positively correlated with HCC patient survival
(16). Recent studies have shown
that B cells are a vital fraction in the formation of tertiary
lymphoid structures (TLSs). The presence of TLSs in patients is
associated with an improved clinical response to the immune
checkpoint blockade in melanoma, sarcoma and renal cell carcinoma
tumors (17–19). However, to the best of our
knowledge, the clinical significance and functions of B-TILs in
Chinese patients with HCC have not been previously reported. The
investigation of these topics is of high importance for HCC
immunotherapy.
In the present study, clinical specimens were
collected from patients with HCC in Beijing. The clinical data and
the function of B-TILs were assessed. Following immunofluorescence
(IF) staining, it was found that HCC tumors were highly enriched
with CD19/CD20+ B cells, which was associated with
compromised cytotoxic function of T and natural killer (NK) cells.
In addition, PD-L1 expression correlated significantly with CD20
expression. Collectively, these findings indicate that B-TILs may
be a promising candidate for HCC immunotherapy, alone or in
combination with anti-PD-L1/PD-1 antibodies.
Materials and methods
Patients and specimens
Fresh tumor tissues, para-tumor tissues, normal
liver and matched blood samples were collected from patients with
HCC undergoing surgical treatment at the China-Japan Friendship
Hospital (Beijing, China). From May 2018 to July 2020, 50 patients
were enrolled in this study. The ratio of male to female patients
was 42:8. The mean age was 60.8 years, with an age range from 36 to
84 years. All patient data were anonymized prior to study
inclusion.
All procedures followed were in accordance with the
ethical standards of the responsible committee on human
experimentation (The Institutional Review Board at Tsinghua
University (Beijing, China) and with the Helsinki Declaration of
1975, as revised in 2000 (5). All
studies were approved by the Medical Ethics Committee at Tsinghua
University (approval no. 20180017). Informed consent was obtained
from all the subjects in this study.
Isolation of peripheral blood
mononuclear cells (PBMCs) and tumor-infiltrating lymphocytes
(TILs)
Blood samples from patients with HCC were incubated
in heparinized tubes and centrifuged on Ficoll-Hypaque gradients
(Cytiva). Fresh normal liver, para-tumor and tumor samples from
patients with HCC were digested in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 0.5 mg/ml Collagenase
Type IV (Gibco; Thermo Fisher Scientific, Inc.), 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) plus 10 U/ml DNase I for 30 min at
37°C prior to Ficoll-Hypaque (Cytiva) gradient centrifugation.
Flow cytometry
The following antibodies were used: Anti-CD45 (cat.
no. HI30; eBioscience™), anti-CD19 (cat. no. HIB19, eBioscience™),
anti-CD3 (cat. no. OKT3 OKT3), anti-CD56 (cat. no. HCD56;
BioLegend), anti-CD8 (cat. no. SK1; BD Biosciences),
anti-interferon γ (IFN-γ; cat. no. B27; BioLegend), anti-tumor
necrosis factor-α (TNF-α; cat. no. MAb11; Invitrogen™; Thermo
Fisher Scientific, Inc.), anti-Ki67 (cat. no. B56; BD Biosciences),
anti-ranzyme B (cat. no. GRB05; Thermo Fisher Scientific, Inc.) and
anti-perforin (cat. no. dG9; Thermo Fisher Scientific, Inc.). The
cells were stimulated with phorbol 12-myristate 13-acetate (50
ng/ml; Sigma-Aldrich; Merck KGaA) and ionomycin (500 ng/ml;
Sigma-Aldrich; Merck KGaA) for 1 h and subsequently Brefeldin A
(Golgi-plug; BD Biosciences) was added for an additional 4 h.
Single cell suspensions were stained with antibodies against
surface molecules at 4°C for 20 min. Intracellular cytokine
staining was performed following fixation of the cells and their
permeabilization with eBioscience Fixation/Permeabilization buffer.
Finally, the cells were stained with antibodies at 4°C for 40 min.
The cells were analyzed on a LSR Fortessa (BD Biosciences) flow
cytometer and the data were analyzed using FlowJo.X 10.8 software
(FlowJo, LLC). The non-viable cells were excluded based on
viability dye staining (Fixable viability dye eF506; eBioscience;
Thermo Fisher Scientific, Inc.).
Histological and immunohistochemical
analyses
All samples were fixed with 4% neutral formaldehyde,
dehydrated conventionally and embedded in paraffin. The slides were
cut at a thickness of 4 µm, stained with hemoxylin and eosin
(H&E) and observed under light microscopy. Immunohistochemical
staining was performed using the streptavidin-peroxidase method.
The primary antibodies [anti-α-fetoprotein (AFP), anti-Ki67 and
anti-CD34] and other kits were purchased from Beijing Zhongshan
Jinqiao Biotechnology Co., Ltd.; OriGene Technologies, Inc. All
operating procedures were carried out in strict accordance with the
instructions provided by the manufacturer. PBS buffer was used
instead of the primary antibody as negative control and the known
positive specimens were used as positive control.
IF and multiplex tissue
fluorescence
All samples were fixed in formalin solution and
embedded in paraffin. The IF activity was performed according to
the method described previously (20). Each slide was incubated with the
primary antibodies against CD45 (cat. no. ab10559; Abcam), CD20
(cat. no. ab9475; Abcam), CD138 (cat. no. ab34164; Abcam) or PD-L1
(cat. no. ab58810, Abcam) at 4°C overnight. AffiniPure F(ab′)2
Fragment donkey anti-rabbit/mouse/goat Ig was selected as the
secondary antibody. The slides were incubated with mounting medium
containing DAPI for 20 min. The images were obtained using a
fluorescence microscope (Carl Zeiss AG). The quantification
analysis was performed using ImageJ 1.51p (National Institutes of
Health) software. Multiplex tissue fluorescence staining was
performed following blocking with the blocking buffer for 15 min
and each slide was incubated with the primary antibodies against
CD20 (cat. no. ab9475; Abcam), CD56 (cat. no. ab200698; Abcam), CD4
(cat. no. ab133616, Abcam), CD8 (cat. no. ab199016; Abcam) and
forkhead box P3 (FOXP3; cat. no. ab20034; Abcam) at 4°C overnight.
Anti-mouse/rabbit HRP was used as the secondary antibody. The
images were acquired by Vectra Polaris 1.0.7 and analyzed by inform
2.4.1 (Perkin Elmer, Inc.).
Single cell sequencing data
analysis
10× Genomics data were downloaded from the Gene
Expression Omnibus (GSE) dataset GSE140228 (21). The immune cells in tumors and PBMCs
were separated for further analysis. Seurat (21) was used to perform clustering
analysis of single-cell data from different tissues. The cells that
exhibited unique feature counts over 6,000 or <200 were
filtered. The cells that exhibited <10% mitochondrial genes, and
genes expressed in >3 cells were selected for further analysis.
The datasets from different tissues were integrated together using
the FindIntegrationAnchors and IntegrateData function.
Non-linear dimensional reduction (UMAP) (22) was selected for clustering analysis.
B cells were selected for this analysis. FindMarkers function was
performed to identify differentially expressed markers across
tissues. FeaturePlot function was used to visualize the gene
expression changes with the split by option.
Statistical analysis
Statistical comparisons were performed using
unpaired Student's t-test for two-group analysis or ordinary
One-way ANOVA followed by Dunnett's multiple comparison test for
three-group analysis. Simple linear regression was used to
calculate the correlations. P<0.05 was considered to indicate a
statistically significant difference.
Results
HCC tumors are enriched by
CD20+ B cells and CD45+CD138+
plasma cells
To explore the roles of B cells derived from HCC TME
in tumor progression, the expression levels of membrane spanning
4-domains A1 (MS4A1; encoding CD20) were initially
investigated using the online database The Cancer Genome Atlas. The
data indicated that MS4A1 expression in tumor tissues was
decreased compared with that in normal control tissues (left panel,
Fig. 1A). This result was further
confirmed by investigation of the HCC tumor samples collected from
the China-Japan Friendship Hospital (right panel, Fig. 1A), n=14 for normal tissue, n=34 for
paratumor and n=31 for tumor. The tumors were stained with H&E
for histological evaluation. Abnormalities in histopathological
morphology were observed in all the cases examined (Fig. 1B). In total, 37.8% (17 of 45 cases)
of the tumors were positive for AFP and 100% (41/41) for CD34. A
high Ki67 index (>15%) was detected in 77.3% (34/44) of the
cases. To note, some of the patients (N=50) were not tested for
AFP, CD34 or Ki67. The clinical and pathological characteristics of
the treatment-naïve HCC patients are summarized in Table I.
 | Table I.Clinicopathological features of the
patients with HCC in this study. |
Table I.
Clinicopathological features of the
patients with HCC in this study.
|
|
|
|
|
|
| Histochemistry |
|---|
|
|
|
|
|
|
|
|
|---|
| Patient no. | Sex | Age (years) | HBV Positivity | BCLC stage | B-TIL density | AFP | Ki67 | CD34 |
|---|
| 1 | Male | 42 | + | B | Low | + | 40% | + |
| 2 | Male | 65 | - | A | High | - | 10%+ | + |
| 3 | Male | 47 | + | A | Low | NA | NA | + |
| 4 |
Female | 73 | - | A | NA | - | 30% | + |
| 5 | Male | 73 | + | A | Low | + | 60% | + |
| 6 | Male | 64 | - | A | High | NA | NA | + |
| 7 | Male | 36 | + | C | Low | + | 50%+ | + |
| 8 | Male | 62 | - | A | High | - | 30%+ | NA |
| 9 |
Female | 59 | + | A | High | + | NA | + |
| 10 | Male | 58 | + | C | High | - | 50%+ | + |
| 11 | Male | 55 | + | B | Low | + | 20%+ | + |
| 12 | Male | 45 | + | C | High | - | 70%+ | + |
| 13 | Male | 51 | + | B | High | - | 60%+ | + |
| 14 | Male | 58 | + | B | Low | + | 90%+ | + |
| 15 | Male | 58 | + | A | Low | - | 40%+ | + |
| 16 | Male | 58 | + | C | High | - | 25%+ | + |
| 17 | Male | 65 | + | C | High | - | 80%+ | + |
| 18 | Male | 64 | + | A | Low | - | 60%+ | + |
| 19 | Male | 84 | + | A | Low | - | 10% | + |
| 20 | Male | 72 | + | A | Low | - | 20%+ | + |
| 21 | Male | 81 | - | C | Low | - | 5%+ | + |
| 22 | Male | 77 | - | B | High | + | 60%+ | + |
| 23 | Male | 77 | + | B | High | - | 50%+ | + |
| 24 |
Female | 60 | + | A | Low | - | 2%+ | + |
| 25 | Male | 60 | + | B | High | - | 5%+ | + |
| 26 | Male | 68 | + | A | High | - | 30%+ | NA |
| 27 | Male | 60 | + | C | Low | - | 30%+ | + |
| 28 |
Female | 76 | + | B | Low | NA | NA | + |
| 29 | Male | 74 | + | A | Low | + | 30%+ | + |
| 30 | Male | 54 | + | A | High | + | 30%+ | + |
| 31 | Male | 61 | + | A | High | - | 30-40% + | + |
| 32 | Male | 40 | + | A | High | - | 20%+ | + |
| 33 | Male | 66 | + | B | Low | + | 50%+ | + |
| 34 | Male | 63 | + | B | High | - | 20%+ | NA |
| 35 | Male | 46 | + | A | High | - | 20%+ | + |
| 36 | Male | 49 | + | B | Low | + | 70%+ | + |
| 37 |
Female | 63 | + | B | Low | + | 40%+ | NA |
| 38 | Male | 63 | + | A | Low | NA | NA | NA |
| 39 | Male | 63 | + | A | NA | - | 15%+ | + |
| 40 | Male | 57 | + | A | High | NA | NA | NA |
| 41 |
Female | 58 | + | B | NA | + | 60%+ | + |
| 42 | Male | 48 | + | B | NA | - | 5%+ | + |
| 43 | Male | 72 | - | A | NA | + | 60%+ | + |
| 44 | Male | 60 | + | B | NA | + | 60%+ | + |
| 45 |
Female | 66 | + | B | NA | + | 10%+ | + |
| 46 | Male | 51 | + | B | NA | + | 60%+ | NA |
| 47 | Male | 76 | + | A | NA | - | 15%+ | NA |
| 48 |
Female | 45 | - | B | NA | - | 5%+ | NA |
| 49 | Male | 47 | + | A | NA | + | 30%+ | + |
| 50 | Male | 70 | + | B | NA | - | 35%+ | + |
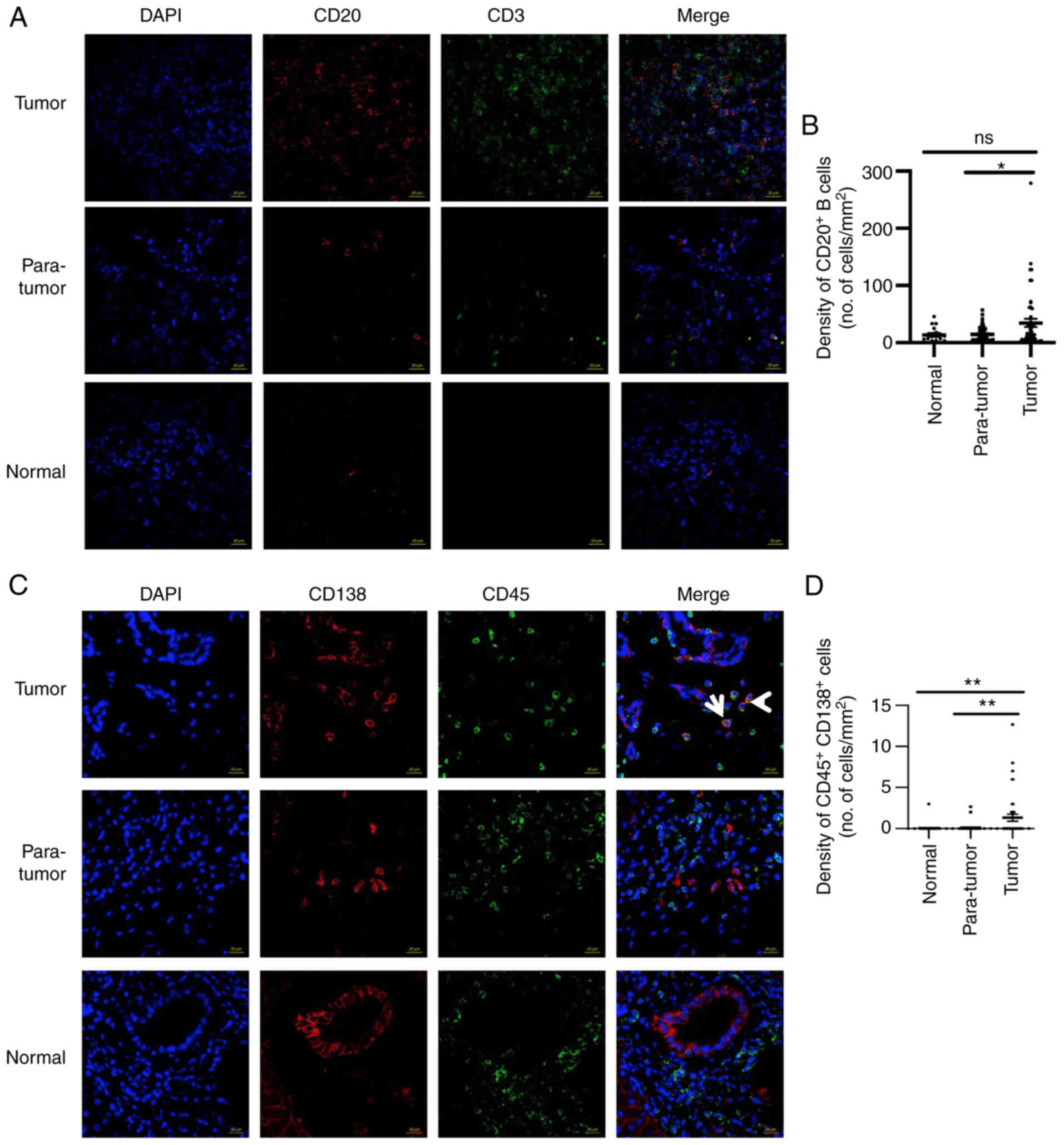
Subsequently, IF was performed to assess CD20
protein levels in HCC tumor and control tissues. The expression
levels of the CD20 protein were detected in normal (n=17),
para-tumor (n=46) and tumor (n=50) tissues (Fig. 2A and B). Non-significant
differences were observed with regards to CD20 expression between
normal and para-tumor tissues. However, in tumor tissues, the
percentage of CD20+ B cells was significantly elevated
compared with that noted in the para-tumor tissues. The difference
between normal controls and tumors was not statistically
significant, which may be due to the smaller size of the normal
control group (Fig. 2B). The low
protein levels of CD20 in normal tissues were contradictory to the
high mRNA levels noted in the same tissues, which suggested that
translation of CD20 protein in these tissues may be restrictively
regulated at the posttranscriptional or translation levels.
Notably, no correlation was found between the density of
CD20+ B cells and the Ki67 score (data not shown).
In addition, the number of plasma cells in the HCC
TME was measured (Fig. 2C). A
significantly higher number of plasma cells
(CD45+CD138+ cells) were noted in the tumors
than those found in the para-tumors or normal control samples. The
majority of the patients did not exhibit plasma cells in either
normal or para-tumor tissues, whereas these cells were present in
the HCC tumors (Fig. 2C and D).
Taken together, the data indicated that HCC tumors were infiltrated
by CD20+ B and CD45+CD138+ plasma
cells.
Higher density of B-TILs is associated
with compromised cytotoxic function of both T and NK cells
In order to assess the clinical relevance of B-TILs,
infiltrating lymphocytes were isolated from normal, para-tumor and
tumor tissues and the percentage of B cells
(CD3−CD19+) was analyzed via flow cytometry.
The percentages of B cells were similar in the normal and
para-tumor tissues and ranged from 0 to 10% of the total number of
CD45+ lymphocytes (Fig. 3A
and B). In tumor tissues, nearly 50% of the patients indicated
an increase in the percentage of CD19+ B cells
(1.6-38.8% of total CD45+ lymphocytes), which was in
line with the results derived from IF staining.
The patients were divided into two groups based on
the B cell percentages of the total CD45+ lymphocytes
(the cut-off value was 1.5%), with 17 patients in the B
cellhigh group and 19 patients in the B
celllow group. The expression levels of the functional
markers were analyzed in NK and T cells between the two groups.
Although the frequency of CD3+ TILs was significantly
increased in the B cellhigh group, no significant
difference was noted with regard to the percentage of NK cells
(Fig. 3C). Subsequently, the
functional capacities of these TILs were evaluated. Notably, the
CD8+ TILs in the B cellhigh group indicated
reduced cytotoxic activity compared with those in the B
celllow group, based on the decreased expression levels
of granzyme B, perforin and IFN-γ (Fig. 3D) in the B cellhigh
group. However, the cytotoxic activity of CD4+ TILs was
not significantly different between these two groups (data not
shown). Moreover, non-significant differences were noted with
regard to the tumor stage between the two groups (data not
shown).
The concept of innate immunity is increasingly
accepted in tumor immunology. NK cells act as innate effector
lymphocytes that respond to tumor cells. The percentages of NK-TILs
exhibited no significant differences between the B
celllow and B cellhigh groups (Fig. 3E). In addition, the expression
levels of perforin and granzyme B were not different between the
two groups examined (Fig. 3E),
indicating that B cells did not affect NK cell function via the
perforin/granzyme B pathway. However, the B cellhigh
group indicated a significant decrease in the frequency of
IFN-γ+ NK+ cells, compared with that of the B
celllow group (Fig.
3E). These results indicated that high accumulation of B cells
in the TME was associated with reduced NK function in the TME.
Intratumoral B cells are positively
associated with infiltration of regulatory T cells (Treg)
cells
To further investigate the profile of immune cells
in HCC tumors, multi-IF staining was used to assess the
colocalization of major lymphocyte subtypes identified by CD20,
CD8, CD57, CD4 and FOXP3 in the tumor tissues from 38 patients with
HCC. CD20+ B cells were co-localized with both
CD4+ and CD8+ T cells, while in some cases
CD20+ B cells were in contact with CD8+ T
cells (Fig. 4A). It was also shown
that CD20+ B cells were surrounded by CD8+ T
cells mostly in the stroma area. This was noted for 11 cases, which
indicated the formation of TLS. It is interesting to note that
FOXP3+ Treg and CD20+ B cells were closely
colocalized within the TLS. In contrast to these observations,
CD57+ NK cells were not colocalized with
CD20+ B cells (Fig.
4A). Additional quantification of the multi-IF staining data
indicated that the number of CD20+ B cells was
positively correlated with FOXP3+ Treg cells (Fig. 4B). In addition, the tumor tissues
were divided into tumor bed and stroma areas. Statistical analysis
indicated that the number of CD20+ B cells was
positively correlated with that of FOXP3+ Treg cells in
the tumor bed but not the stroma area (Fig. 4C and D), which indicated that
B-TILs may contribute to the infiltration of Treg cells, which in
turn leads to tumor immune escape.
High levels of nuclear receptor
subfamily 4 group A member 2 (NR4A2) are expressed in B-TILs
To illustrate the characteristics of B cells in HCC,
the present study used published single cell RNA-sequencing
(RNA-seq) data of CD45+ cells from 16 treatment-naïve
patients with HCC (23). The
single cell RNA-seq data of B cells from tumors and PBMCs were
combined. UMAP was performed to identify three major clusters (C0,
C1 and C2; Fig. 5A). The PBMCs
contained 23% C0, 28% C1 and 49% C2, while the HCC tumors contained
80% C0, 15% C1 and 6% C2 (Fig.
5A). The C0 expressed markers of memory B cells, such as
CCL4, RGS2, AREG, ZFP36, CCL5, CD82 and CD1C, whereas
the C1 expressed markers of plasmablasts, including IGHG1,
IGHG3, CD27, IGHG4, IGHA1, NEAT1, SSPN, S100A10 and
CPNE5, and the C2 was mainly indicative of naïve B cells
(TCL1A, IGHD1, CD72, APLP2, CD200, GCA, CD79A, CD79B and
CD74; Fig. 5B). The
expression levels of 17 genes (FOS, RGS2, RGS1, CD83, CD69,
JUNB, NR4A2, GPR183, HERPUD1, FOSB, YPEL5, SRGN, ANF331, IGHG1,
ZFP36, SLC2A3 and DUSP1) were significantly increased in
intratumoral B cells (fold change cut-off value >2) compared
with those noted in peripheral B cells. Higher expression of
FOS, FOSB and JUNB, which belong to the activator
protein 1 (AP-1) pathway, were noted in HCC B-TILs than those noted
in the peripheral B cells (Fig.
5C). In addition, NR4A2 expression was also enhanced in B-TILs
(Fig. 5C). NR4A2 is a member of an
orphan nuclear hormone receptor family involved in induction of
apoptosis and carcinogenesis (24). It is preferentially recruited to
the binding sites of the transcription factor AP-1, where it
represses effector-gene expression by inhibiting AP-1 function in T
cells (25,26). Whether NR4A2 can impair B cell
function by binding to AP-1 in NR4A2+ B cells requires
further investigation.
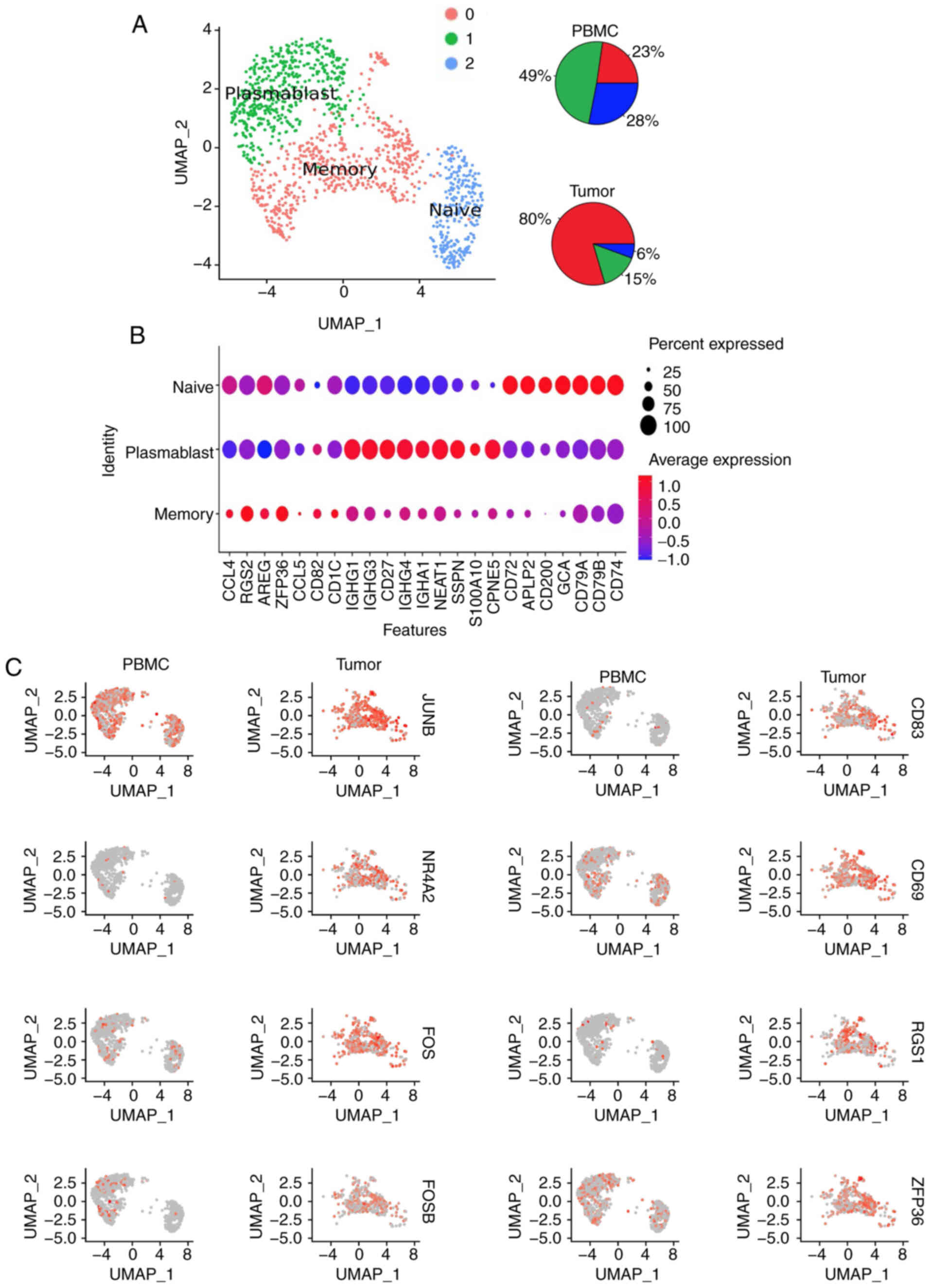 | Figure 5.B-TILs indicate higher expression
levels of NR4A2. (A) UMAP of B cells in tumors and PBMCs
indicating three different clusters. Pie charts indicating
distribution of each cluster in PBMCs and tumors. (B) Expression of
selected genes across the three clusters. (C) UMAP projection split
by tissue of origin (PBMCs or tumor) and colored for expression of
JUNB, NR4A2, FOS, FOSB, CD83, CD69, RGS1, ZFP36. Each spot
represents one B cell. B-TILs, tumor-infiltrating B cells; NR4A2,
nuclear receptor subfamily 4 group A member 2; UMAP, uniform
manifold approximation and projection; PBMCs, peripheral blood
mononuclear cells. |
PD-L1 expression significantly
correlates with CD20+ B cell density
The correlation between PD-L1 and CD20 was examined
in order to determine the rationale of combined therapy. The HCC
tumor slides were analyzed via IF staining (Fig. 6A) and the data indicated that
patients with high PD-L1 expression exhibited high density of B
cells as measured by CD20 expression (Fig. 6B). Subsequently, the HCC tumor
samples were divided into two groups, namely CD20high
PD-L1 high and CD20low PD-L1low
(the thresholds of CD20 and PD-L1 were 6 and 20
cells/mm2, respectively). The data indicated that the
group with the CD20 high PD-L1high samples
exhibited a significant decrease in the percentage of granzyme
B+ CD8+ TILs compared with the
CD20low PD-L1low group (Fig. 6C). Moreover, the
CD20high PD-L1high group exhibited reduced
percentages of both perforin+ and granzyme B+
CD4+ TILs (Fig. 6D).
These data indicated that the combined presence of CD20 and PD-L1
could be used as an indicator of compromised antitumor effects in
the context of HCC.
Discussion
The present study demonstrated that the density of B
cells was increased in the hepatocellular carcinoma (HCC) tumor
microenvironment (TME). Patients with high B cell infiltration had
impaired antitumor activity, featured by both CD8+ T
cells and NK cells with compromised ability to express granzyme B
and IFN-γ. B cells positively were found to be correlated with
FOXP3+ Treg cells. Moreover, PD-L1 expression
significantly correlated with the expression of the B cell marker
CD20.
Immune tolerance can result in weak responses
towards tumor antigens. The understanding of the underlying
mechanisms responsible for immune tolerance can provide strategies
to restore an antitumor immune response (27). The identification of the dominant
immunosuppressive cells in patients with cancer could serve as an
alternative mechanism to overcome immune suppression. It is highly
possible that each TME is controlled by a distinct profile of a
suppressive cellular compartment (28). The present study indicated that B
cells may be a subpopulation of lymphocytes that act as
immunosuppressive cells in patients with HCC. In a mouse model of
skin carcinogenesis, B cells were required to maintain a chronic
inflammation status, which in turn promoted de novo
carcinogenesis. Therefore, deficiency of B cells reduced
infiltration of immune cells in premalignant lesions, while leading
to inhibition of carcinoma development (29,30).
Furthermore, B cell deficiency was also found to be associated with
enhanced antitumor T cell responses in mice with increased IFN-γ
secretion (31), which was
consistent with the findings reported in the present study. In
addition, regulatory B (Breg) cells were identified as a leading
cause of HCC progression (32).
Breg cells were shown to promote HCC progression by increasing
interleukin (IL)-10 and tumor growth factor (TGF)-β production via
activation of the CD40/CD40 ligand signaling pathway, which
resulted in decreased tumor necrosis factor (TNF)-α levels and
reduced antitumor immune response (14). Notably, Breg cells were also
proposed to suppress T cell antitumor immune response by converting
naive CD4+ T cells to Treg cells in the TME (33). In the present study, a positive
correlation between the density of B-TILs and intratumoral Treg
cells was noted. However, whether B cells can recruit Treg cells or
directly interact with each other requires further
investigation.
In the present study, the numbers of both
CD8+ T and Treg cells were positively correlated with B
cell density. Garnelo et al (16) also reported that the number of B
cells was also found to correlate with the number of
CD8+ T cells in HCC using IF staining. Although the
number of CD8+ T cells was positively correlated with
that of B cells, their cytotoxic CD8+ T cell (CTL)
function was decreased in patients with high percentage of B cells.
One possible explanation could be the increased percentage of Treg
cells. Treg cells play an indispensable role in immune suppression,
by suppressing the effector activities of CD4+ and
CD8+ T cells as well as the function of nature killer
(NK) cells (34,35). It has been reported that Treg cells
allow CD8+ T cells to proliferate, while compromising
their IFN-γ production and cell-mediated cytotoxicity (36). It is interesting to note that in
the tertiary lymphoid structure (TLS), CD8+ T cells were
localized close to B cells, and Treg cells were present within the
same area. B cells may interact with Treg cells, which in turn
inhibit CTL function. The mechanism and pathway through which B
cells orchestrate the TME requires further extensive research.
Taken together, the data indicate that B-TILs in HCC
are associated with dampened antitumor immunity and may serve as a
promising target for immunotherapy.
Acknowledgements
Not applicable.
Funding
This research study was supported by grants from Natural Science
Foundation of China (31991173 and 31991170), Tsinghua University
Spring Breeze Fund (2020Z99CFG008), Tsinghua University-Xiamen
Chang Gung Hospital Joint Research Center for Anaphylactic Disease,
Youth Program of National Natural Science Foundation of China
(81702822) and Bristol-Myers Squibb.
Availability of data and materials
The RNA-seq datasets analyzed in this study are
available in GEPIA2 (http://gepia2.cancer-pku.cn/#index). 10× Genomics data
were downloaded from GEO dataset (GSE140228, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi) in
the published article (21). Data
sharing is not applicable to this article as no genomics were
generated in this study.
Authors' contributions
YF performed the majority of the experiments; JL,
YS, SX, JHX, LM and WZ performed some of the experiments. LL, JH
and HT collected and analyzed the clinical specimens; LL and ZY
supervised and analyzed the clinical specimens. XZ performed the
single cell data analysis. LN and YF wrote the manuscript; LN
designed and supervised the study. All authors reviewed and
approved the final manuscript. All authors confirm the accuracy of
the data, read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All procedures followed were in accordance with the
ethical standards of the responsible committee on human
experimentation (The Institutional Review Board at Tsinghua
University (Beijing, China) and with the Helsinki Declaration of
1975, as revised in 2000 (5). All
studies were approved by the Medical Ethics Committee at Tsinghua
University (approval no. 20180017). Informed consent was obtained
from all the subjects in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests. JHX and LM are employees of Bristol Myers Squibb.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
AFP
|
α-fetoprotein
|
|
TME
|
tumor microenvironment
|
|
TCGA
|
The Cancer Genome Atlas
|
|
B-TILs
|
tumor-infiltrating B cells
|
|
CTLs
|
cytotoxic CD8+ T cells
|
|
Breg cells
|
regulatory B cells
|
|
TLS
|
tertiary lymphoid structure
|
|
ICB
|
immune checkpoint blockade
|
|
PBMCs
|
peripheral blood mononuclear cells
|
|
UMAP
|
non-linear dimensional reduction
|
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
European Association for the Study of the
Liver. Electronic address, . simpleeasloffice@easloffice.eu;
European Association for the Study of the Liver: EASL clinical
practice guidelines: Management of hepatocellular carcinoma. J
Hepatol. 69:182–236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu RX, Seto WK, Lai CL and Yuen MF:
Epidemiology of hepatocellular carcinoma in the asia-pacific
region. Gut Liver. 10:332–339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang JD and Roberts LR: Hepatocellular
carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 7:448–458.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
et al: Sorafenib in advanced hepatocellular carcinoma. N Engl J
Med. 359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Buonaguro L, Mauriello A, Cavalluzzo B,
Petrizzo A and Tagliamonte M: Immunotherapy in hepatocellular
carcinoma. Ann Hepatol. 18:291–297. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
El-Khoueiry AB, Sangro B, Yau T, Crocenzi
TS, Kudo M, Hsu C, Kim TY, Choo SP, Trojan J, Rd THW, et al:
Nivolumab in patients with advanced hepatocellular carcinoma
(CheckMate 040): An open-label, non-comparative, phase 1/2 dose
escalation and expansion trial. Lancet. 389:2492–2502. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yarchoan M, Hopkins A and Jaffee EM: Tumor
mutational burden and response rate to PD-1 inhibition. N Engl J
Med. 377:2500–2501. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leonardi GC, Candido S, Cervello M,
Nicolosi D, Raiti F, Travali S, Spandidos DA and Libra M: The tumor
microenvironment in hepatocellular carcinoma (review). Int J Oncol.
40:1733–1747. 2012.PubMed/NCBI
|
|
10
|
Shalapour S, Lin XJ, Bastian IN, Brain J,
Burt AD, Aksenov AA, Vrbanac AF, Li W, Perkins A, Matsutani T, et
al: Inflammation-induced IgA+ cells dismantle anti-liver
cancer immunity. Nature. 551:340–345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye L, Zhang Q, Cheng Y, Chen X, Wang G,
Shi M, Zhang T, Cao Y, Pan H, Zhang L, et al: Tumor-derived
exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by
promoting TIM-1(+) regulatory B cell expansion. J Immunother
Cancer. 6:1452018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xue H, Lin F, Tan H, Zhu ZQ, Zhang ZY and
Zhao L: Overrepresentation of IL-10-expressing B cells suppresses
cytotoxic CD4+ T cell activity in HBV-induced
hepatocellular carcinoma. PLoS One. 11:e01548152016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schwartz M, Zhang Y and Rosenblatt JD: B
cell regulation of the anti-tumor response and role in
carcinogenesis. J Immunother Cancer. 4:402016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shaso Y, Lo CM, Ling CC, Liu XB, Ng KTP,
Chu AC, Ma YY, Li CX, Fan ST and Man K: Regulatory B cells
accelerate hepatocellular carcinoma progression via CD40/CD154
signaling pathway. Cancer Lett. 355:264–272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Z, Ma L, Goswami S, Ma J, Zheng B,
Duan M, Liu L, Zhang L, Shi J, Dong L, et al: Landscape of
infiltrating B cells and their clinical significance in human
hepatocellular carcinoma. Oncoimmunology. 8:e15713882019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garnelo M, Tan A, Her Z, Yeong J, Lim CJ,
Chen J, Lim KH, Weber A, Chow P, Chung A, et al: Interaction
between tumour-infiltrating B cells and T cells controls the
progression of hepatocellular carcinoma. Gut. 66:342–351. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cabrita R, Lauss M, Sanna A, Donia M,
Larsen MS, Mitra S, Johansson I, Phung B, Harbst K,
Vallon-Christersson J, et al: Tertiary lymphoid structures improve
immunotherapy and survival in melanoma. Nature. 577:561–565. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Helmink BA, Reddy SM, Gao J, Zhang S,
Basar R, Thakur R, Yizhak K, Sade-Feldman M, Blando J, Han G, et
al: B cells and tertiary lymphoid structures promote immunotherapy
response. Nature. 577:549–555. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Petitprez F, de Reynies A, Keung EZ, Chen
TWW, Sun CM, Calderaro J, Jeng YM, Hsiao LP, Lacroix L, Bougoüin A,
et al: B cells are associated with survival and immunotherapy
response in sarcoma. Nature. 577:556–560. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie S, Huang J, Qiao Q, Zang W, Hong S,
Tan H, Dong C, Yang Z and Ni L: Expression of the inhibitory B7
family molecule VISTA in human colorectal carcinoma tumors. Cancer
Immunol Immunother. 67:1685–1694. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stuart T, Butler A, Hoffman P, Hafemeister
C, Papalexi E, Mauck WM III, Hao Y, Stoeckius M, Smibert P and
Satija R: Comprehensive integration of single-cell data. Cell.
177:1888–1902.e21. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McInne L and Healy J: UMAP: Uniform
Manifold Approximation and Projection for Dimension Reduction.
ArXiv e-prints 1802.03426. 2018.https://arxiv.org/abs/1802.03426
|
|
23
|
Zhang Q, He Y, Luo N, Patel SJ, Han Y, Gao
R, Modak M, Carotta S, Haslinger C, Kind D, et al: Landscape and
dynamics of single immune cells in hepatocellular carcinoma. Cell.
179:829–845, e820. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li QX, Ke N, Sundaram R and Wong-Staal F:
NR4A1, 2, 3-an orphan nuclear hormone receptor family involved in
cell apoptosis and carcinogenesis. Histol Histopathol. 21:533–540.
2006.PubMed/NCBI
|
|
25
|
Liu X, Wang Y, Lu H, Li J, Yan X, Xiao M,
Hao J, Alekseev A, Khong H, Chen T, et al: Genome-wide analysis
identifies NR4A1 as a key mediator of T cell dysfunction. Nature.
567:525–529. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen J, Lopez-Moyado IF, Seo H, Lio CWJ,
Hempleman LJ, Sekiya T, Yoshimura A, Scott-Browne JP and Rao A:
NR4A transcription factors limit CAR T cell function in solid
tumours. Nature. 567:530–534. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ren Z, Peng H and Fu YX: PD-1 shapes B
cells as evildoers in the tumor microenvironment. Cancer Discov.
6:477–478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
de Visser KE, Korets LV and Coussens LM:
De novo carcinogenesis promoted by chronic inflammation is B
lymphocyte dependent. Cancer Cell. 7:411–423. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Andreu P, Johansson M, Affara NI, Pucci F,
Tan T, Junankar S, Korets L, Lam J, Tawfik D, DeNardo DG, et al:
FcRγ activation regulates inflammation-associated squamous
carcinogenesis. Cancer Cell. 17:121–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qin Z, Richter G, Schüler T, Ibe S, Cao X
and Blankenstein T: B cells inhibit induction of T cell-dependent
tumor immunity. Nat Med. 4:627–630. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hetta HF: Regulatory B cells: Key players
in hepatocellular carcinoma progression. Gastroenterology &
Hepatology. Open Access 52016. https://medcraveonline.com/GHOA/GHOA-05-00136.pdf
|
|
33
|
DiLillo DJ, Matsushita T and Tedder TF:
B10 cells and regulatory B cells balance immune responses during
inflammation, autoimmunity, and cancer. Ann NY Acad Sci.
1183:38–57. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shevach EM: From vanilla to 28 flavors:
Multiple varieties of T regulatory cells. Immunity. 25:195–201.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
von Boehmer H: Mechanisms of suppression
by suppressor T cells. Nat Immunol. 6:338–344. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin CY, Graca L, Cobbold SP and Waldmann
H: Dominant transplantation tolerance impairs CD8+ T
cell function but not expansion. Nat Immunol. 3:1208–1213. 2002.
View Article : Google Scholar : PubMed/NCBI
|