Introduction
In women and men, colorectal cancer (CRC) is the
second and third most prevalent type of cancer, respectively
(1). High CRC-associated mortality
rates are attributed to the high levels of metastasis and
recurrence (2). Advances in
technological and genetic analyses have improved CRC diagnosis and
therapy (3); however, the
mortality rate of CRC remains at a high level (4). Further investigations into the
underlying molecular mechanisms involved in CRC progression are
required to identify potential prognostic markers.
The C2H2-type zinc finger protein (ZNF), which is
encoded by the ZNF384 gene, is a transcription factor involved in
the transcription of extracellular matrix genes (5). Coupling of ZNF384 with TET family
genes, such as the TATA box binding protein-associated factor,
transcription factor 3 and Ewing sarcoma breakpoint region 1 gene,
serves vital roles in acute lymphocytic leukemia (6,7).
Moreover, transactivating characteristics of the fusion protein
have been reported in NIH3T3 cells, highlighting the oncogenic
potential of ZNF384 as a fusion protein (8). In melanoma cells, ZNF384
overexpression was shown to promote metastasis (9). Although previous studies have
indicated that ZNF384 may act as an oncogenic factor that promotes
cancer progression and metastasis, the specific expression levels
and functions of ZNF384 in human CRC are yet to be fully
elucidated.
The present study aimed to examine the expression
levels of ZNF384 in CRC tissues and cells, and to determine its
prognostic value in patients with CRC. Additionally, the effects of
ZNF384 on CRC cell invasion and migration, as well as the
underlying mechanism, were assessed.
Materials and methods
Cell culture
The normal colonic epithelial cell line FHC, 293T
cells and six human CRC cell lines (SW480, Caco-2, SW620, HT29,
LoVo and HCT116) were purchased from the American Type Culture
Collection. The 293T and CRC cell lines were seeded in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.), and the FHC cell line was seeded in DMEM:F12
(cat. no. D8437; Sigma-Aldrich; Merck KGaA) with 10% FBS. Cells
were incubated in a humidified atmosphere containing 5%
CO2 at 37°C. For cell culture under 0.5% O2
tension, the cells were incubated for 0–24 h in a humidified
atmosphere at 37°C in a multi-gas CO2-O2
incubator (NuAire) equilibrated with 0.5% O2, 5%
CO2 and 94.5% N2. The cell suppliers stated
that the identity of the cells had been confirmed using STR
profiling and were free of mycoplasma contamination. The HT29 cell
line was also identified by STR profiling in the present study. The
following STR profiling markers were analyzed: Amelogenin, CSF1P0,
D3S1358, D2S1338, D5S818, D8S1179, D7S820, D13S317, D16S539,
D19S433, D18S51, D21S11, Penta D, FGA, Penta E, TPOX, TH01 and
vWA.
Patients and tissue samples
Between September 2008 and December 2009, CRC
tissues and corresponding non-carcinoma samples (≥5 cm away from
the tumor margin) were acquired from a total of 167 patients with
CRC subjected to surgical resection at The Affiliated Hospital of
North Sichuan Medical College (Nanchong, China). In addition, the
metastatic lymph tissues were collected from 50 patients with lymph
node metastasis. Three of the enrolled cases developed distant
metastases and were subsequently excluded from the study. Prior to
surgery, the enrolled patients did not receive any adjuvant
therapies. Resected tissues were immersed in liquid nitrogen and
stored at −80°C for subsequent analyses. Ethics approval was
obtained from the Medical Ethics Committee of North Sichuan Medical
College [approval no. 2021ER(A)007]. Prior to study enrollment,
patients provided written informed consent. In order to evaluate
postoperative survival, all patients with CRC were periodically
followed up for between 11 and 96 months. Table SI displays the clinical
characteristics of the study participants.
Immunohistochemical (IHC) staining and
evaluation
CRC tissues were collected from patients with CRC,
fixed for 12 h in 4% paraformaldehyde at room temperature and
subsequently embedded in paraffin. Samples were sectioned into 4-µm
slices. After incubation for 2 h at 60°C in an incubator, the
sections were deparaffinized using dimethylbenzene and rehydrated
using a descending alcohol series. Sections were subsequently
incubated for 30 min at room temperature in the presence of 0.3%
hydrogen peroxide, and blocked for 1 h using 10% BSA (Sangon
Biotech Co., Ltd.) at room temperature. Sections were incubated
overnight at 4°C with the following antibodies: Rabbit anti-ZNF384
(1:50; cat. no. ab251673; Abcam), rabbit anti-HIF-1α (1:100; cat.
no. ab243860; Abcam) and rabbit anti-MMP2 (1:100; cat. no.
ab235167; Abcam). Subsequently, sections were incubated with an
HRP-conjugated secondary antibody (1:200; Goat Anti-Rabbit IgG
H&L; cat. no. ab205718; Abcam) at 37°C for 2 h. The tissues
were then stained at room temperature for 1 h using DAB (OriGene
Technologies, Inc.) and counterstained for 1 min at room
temperature using hematoxylin. All sections were dehydrated and
sealed. Visualization and imaging were carried out using a light
microscope (Carl Zeiss AG), and sections incubated with Rabbit IgG
(1:50; cat. no. A7016; Beyotime Institute of Biotechnology) acted
as negative controls (NCs). Scoring was based on the ratio of
positively stained cells, namely: 0, 0–5; 1, 6–35; 2, 36–70; and 3,
>70%. Staining intensities were as follows: 3, strong; 2,
moderate; 1, weak and 0, no staining. Final score determination was
carried out by multiplying the scores of the percentage of positive
cells with those of the staining intensities. Final scores were
defined as: -, 0–1; +, 2-3; ++, 4-6; and +++, >6. Low expression
levels were determined by a total score <4, whereas elevated
expression was defined as a total score ≥4. Two pathologists
independently performed the scoring.
Establishment of lentiviruses and
stable cell lines
The pLKO.1-EGFP-PURO (cat. no. FH1717; Hunan Fenghui
Biotechnology Co., Ltd.) vector was used to establish lentiviral
vectors with short hairpin (sh)RNA sequences, which were designated
LV-shZNF384, LV-shHIF-1α, LV-shMMP2 and LV-shcontrol. The
LV-shcontrol contained a non-targeting shRNA control. All shRNA
sequences are displayed in Table
SII. Lentiviral vectors with human ZNF384, MMP2 and HIF-1α gene
sequences were established in PLVX-EGFP-IRES-PURO (cat. no. BR684;
Hunan Fenghui Biotechnology Co., Ltd) and designated as LV-ZNF384,
LV-MMP2 and LV-HIF-1α. An empty vector was used as the NC and
labeled LV-control.
To generate stable cell lines, the pPACKH1 HIV
Lentivector Packaging kit (Systems Bioscience, LLC) was used.
Briefly, the expression plasmids, including control plasmids (1.0
µg) and pPACKH1 packaging plasmid mix [5.0 µg; pPACKH1-GAG (3.0
µg), pPACKH1-REV (1.0 µg) and pVSV-G (1.0 µg) plasmids; 3rd
generation packaging system] were transfected into 293T cells at
37°C using Lipofectamine® 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). After 24 h, the culture medium was
mixed with 20% of the medium volume of PEG-it Reagent (Systems
Biosciences, LLC), and incubated overnight at 4°C. After
centrifugation at 72,000 × g for 120 min at 4°C, pellets were
washed in 1X PBS and aliquots were stored at −80°C. The viruses
were transduced into SW480, Caco-2, SW620 and LoVo cells
(1×105 cells/well) using polybrene (8 mg/ml;
Sigma-Aldrich; Merck KGaA) with infection multiplicities of 30–50.
After 24 h at 37°C, fresh medium was used to replace the viral
medium. Selection of stably transduced cells was carried out by the
addition of puromycin (6 µg/ml) and the maintenance concentration
of puromycin used was 3 µg/ml. Western blotting and reverse
transcription-quantitative (RT-q)PCR were used to confirm
successful transduction.
In vivo metastasis assay
A total of 80 BALB/C nude female mice (age, 6 weeks;
weight ~20 g) were purchased from Shanghai SLAC Laboratory Animal
Co., Ltd. Animals were housed in a 60% humidified atmosphere at
24°C, under a 12-h light/dark cycle with free access to purified
drinking water and food. The Experimental Animal Ethics Committee
of North Sichuan Medical College approved the present study
(approval no. 20200908). For the in vivo tail vein
metastasis assay, mice (n=10/group) were inoculated with
5×106 cells in 100 µl PBS through the tail vein. The
mice were divided in to the following groups: SW480 (LV-Control and
LV-ZNF384), SW620 (LV-shControl and LV-shZNF384), SW480-ZNF384
(LV-shControl and LV-shMMP2) and SW620-shZNF384 (LV-Control and
LV-MMP2). Mice survival was recorded daily for 9 weeks, after
which, animals were sacrificed to remove the lung tissues. Lung
tissues were collected, fixed with 4% paraformaldehyde for 12 h at
room temperature and embedded in paraffin for subsequent
pathological examination. Paraffin-embedded tissues were sectioned
into 4-µm slices. Mice were sacrificed via an intraperitoneal
administration of sodium pentobarbital (200 mg/kg). Finally, a
dissection microscope (SZX7; Olympus Corporation) was used to count
the tumor metastases formed in the lungs.
Hematoxylin and eosin (H&E)
staining
Lung tissues sections were de-paraffinized in two
changes of xylene, followed by rehydration in two changes of
absolute ethanol, and two changes of 95 and 70% ethanol. Tissue was
washed briefly in deionized water and stained with Harris
hematoxylin (Thermo Fisher Scientific, Inc.). Slides were then
processed in 0.25% acid alcohol, blued in lithium carbonate and
counterstained with eosin solution (Thermo Fisher Scientific,
Inc.). Tissues were dehydrated in two changes of 95% and absolute
ethanol, and cleared in xylene. Photomicrographs were captured
using an Olympus BH-2 light microscope with DP70 camera operating
with DPS-BSW v3.1 software (Olympus Corporation).
Chromatin immunoprecipitation (ChIP)
assay
The ChIP assay was conducted using the Magna ChIP G
Assay kit (MilliporeSigma) according to the manufacturer's
protocol. Briefly, crosslinking of the transfected SW480 cells was
carried out for 10 min using 1% formaldehyde at 37°C, followed by
quenching with glycine. Co-immunoprecipitation of bound DNA from
sonicated (VCX750; Sonics & Materials, Inc.; frequency: 20 kHz;
25% power; 4.5S impact; 9S gap; 14 times in total) cell lysates
(six rounds of 15 sec on, 90 sec off) was carried out following
incubation with primary antibodies against HIF-1α (rabbit; 1:100;
cat. no. 36169; Cell Signaling Technology, Inc.), ZNF384 (rabbit;
1:200; cat. no. ab251673; Abcam) and normal IgG (rabbit; 1:100;
cat. no. 3900; Cell Signaling Technology, Inc.) overnight at 4°C.
Amplification of the corresponding promoter binding sites was
carried out using PCR, and the corresponding primer sequences are
displayed in Table SII. PCR was
performed using Taq DNA polymerase (cat. no. EP0405; Thermo Fisher
Scientific, Inc.) and a PCR system (Takara Biotechnology Co.,
Ltd.). The thermocycling conditions were as follows: Initial
denaturation at 94°C for 5 min; followed by 35 cycles at 94°C for
40 sec, 60°C for 30 sec and 72°C for 60 sec, and a final extension
step at 72°C for 7 min. Experiments were carried out for three
biological repeats.
Cell proliferation analysis
An MTT assay was performed to assess the levels of
cell proliferation. Briefly, 100 µl transfected cells (SW480 or
SW620; 5×103 cells/well) were plated in 96-well plates.
Following incubation for 24, 48, 72 or 96 h, 5 mg/ml MTT solution
(20 µl) was added to each well and incubated for a further 4 h at
37°C. Subsequently, the MTT solution was removed and 150 µl DMSO
was added. Absorbance was measured at a wavelength of 490 nm using
a SpectraMax M5 microplate reader (Molecular Devices, LLC). Five
biological repeats were performed.
Transwell migration and invasion
assays
Transwell inserts with polycarbonate membranes (pore
size, 8.0 µm) were placed in 24-well plates. The invasion assay was
performed after precoating the upper chamber with 50 µl Matrigel
(Corning, Inc.) for 30 min at 37°C, followed by overnight drying.
For the migration and invasion assays, 1×104 and
1×105 cells, respectively, were seeded in the top
chamber. Subsequently, complete medium (600 µl) was added to the
lower chamber and incubated for 24 h. Cells on the upper surface
were removed by swabbing. Cells on the lower surface were fixed in
10% formalin at 25°C for 20 min, followed by staining with crystal
violet (0.1%) at 25°C for 5 min and counted using an inverted light
microscope (magnification, ×20; Olympus Corporation). Three
biological repeats were performed.
Wound healing assay
Cells were plated into 6-well plates and cultured
under standard conditions until 100% confluence was reached.
Subsequently, a 1-ml pipette tip was used to scratch the cell
monolayer to generate a linear cell wound and the floating cells
were gently washed twice with DMEM. Cells were cultured at 37°C in
DMEM (containing 1% FBS) for 24 h. The cells migrating into the
wounded areas were captured using a light microscope
(magnification, ×20) at 0 and 48 h. Wound healing was assessed
using MShot Image Analysis System 1.3.10 (Guangzhou Mingmei
Photoelectric Technology Co., Ltd.). Wound closure was assess using
the following equation: Wound closure (%)=(area at T0-area at
T48)/area at T0 ×100. Three biological repeats were performed.
RT-qPCR
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). PrimeScript RT Reagent kit (Takara Bio, Inc.) was used for
cDNA synthesis according to the manufacturer's protocol. To
determine the mRNA expression levels of ZNF384 and MMP2, qPCR was
performed using SYBR Premix Ex Taq II (Takara Bio, Inc.) on the ABI
7500 Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The qPCR thermocycling conditions were as
follows: 95°C for 10 min; followed by 40 cycles at 95°C for 30 sec,
60°C for 30 sec and 72°C for 30 sec; followed by a final extension
step at 72°C for 2 min. Samples without a cDNA template were used
as the NC. Analysis of amplification curves was carried out using
SDS 1.9.1 software (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The expression levels of target genes in the cell lines were
determined using the 2−ΔΔCq method (10) using the following equations:
ΔCq=ΔCqtarget-ΔCqGAPDH and
ΔΔCq=ΔCqexpression vector-ΔCqcontrol vector.
Expression levels were normalized to matched control cells, which
were set to 1.0. In clinical tissue samples, fold changes in
expression levels of target genes were also determined using the
2−ΔΔCq method, as per the following equations:
ΔCq=ΔCqtarget-ΔCqGAPDH, and
ΔΔCq=ΔCqtumor-ΔCqnontumor or
ΔΔCq=ΔCqlymph node metastatic-ΔCqnontumor.
Expression levels were normalized to healthy colorectal tissues,
which were also set to 1.0. Primer sequences are displayed in
Table SII.
Western blotting
Total proteins were extracted from cells using RIPA
lysis buffer (Beijing Solarbio Science & Technology Co., Ltd.)
supplemented with proteinase inhibitors. The BCA method was used to
assess protein concentrations. Proteins (50 µg/lane) were separated
by SDS-PAGE on 10% gels and were subsequently transferred to PVDF
membranes. After blocking for 1.5 h with 5% skim milk at room
temperature, the membranes were incubated overnight at 4°C in the
presence of primary antibodies against ZNF384 (rabbit; 1:1,000;
cat. no. ab251673; Abcam), MMP2 (rabbit; 1:1,000; cat. no.
ab235167; Abcam), HIF-1α (rabbit; 1:2,000; cat. no. ab243860;
Abcam) and the control β-actin (mouse; 1:500; cat. no. ab8226;
Abcam). Subsequently, membranes were incubated with anti-rabbit
(HRP-conjugated; 1:5,000; cat. no. sc-2357; Santa Cruz
Biotechnology, Inc.) and anti-mouse (HRP-conjugated; 1:10,000; cat.
no. sc-2005; Santa Cruz Biotechnology, Inc.) secondary antibodies
for 1 h at 37°C. Visualization of protein bands was carried out
using electrochemiluminescence reagent (cat. no. WBKLS0500;
MilliporeSigma). Semi-quantification of protein expression levels
was carried out using ImageJ software (version 1.8.0; National
Institutes of Health), with β-actin used as the loading control.
Experiments were carried out for three biological repeats.
Plasmid construction
Plasmid vectors were established using standard
procedures. Primers used in the present study are displayed in
Table SII. PCR was used to
amplify the ZNF384 promoter sequence (−1,996/+115) from human
genomic DNA extracted from SW480 cells using a genomic DNA
extraction kit (cat. no. ab156900; Abcam). PCR was performed using
Takara LA Taq polymerase and PCR system (Takara Biotechnology Co.,
Ltd.). The thermocycling conditions were as follows: Initial
denaturation at 94°C for 5 min; followed by 35 cycles at 94°C for
30 sec, 60°C for 30 sec and 72°C for 40 sec, and final extension at
72°C for 5 min. This sequence is localized at the transcriptional
start site position (−1,996/+115) in the 5′-flanking region of the
human ZNF384 gene. Vector construction was carried out by
integrating both forward and reverse primers in the 5′- and 3′-ends
of KpnI and HindIII sites, respectively. Insertion of PCR products
between the digested HindIII and KpnI sites of the pGL3-Basic
vector (Promega Corporation) was performed. Moreover, the
5′-flanking region deletion mutants of the ZNF384 promoter
[(−1,996/+115) ZNF384; (−1,752/+115) ZNF384; and (−317/+115)
ZNF384] were established using the (−1,996/+115) ZNF384 vector as
the template. Mutations in the HIF-1α-binding sites in the ZNF384
promoter were made using the QuikChange II Site-Directed
Mutagenesis kit (Stratagene; Agilent Technologies, Inc.). Vector
construction was verified using first DNA sequencing (Sangon
Biotech Co., Ltd.) and HIF-1α promoter vectors were designed in the
same way.
Transient transfection and luciferase
assay
ZNF384 and HIF-1α expression plasmids were generated
by cloning ZNF384 or HIF-1α DNA into pCMV-tag2A vectors (Agilent
Technologies, Inc.). SW480 or SW620 cells were cultured in a
24-well plate at 1×105 cells/well. After 12–24 h
incubation, cells were co-transfected with expression plasmids [0.6
µg; pCMV-ZNF384, pCMV-HIF-1α or the control (pCMV-Tag)], reporter
plasmids (0.18 µg) and pRL-TK plasmids (0.02 µg) (Promega
Corporation) using Lipofectamine 3000 reagent. A total of 5 h
post-transfection, cells were washed and placed in fresh medium
containing 1% FBS for 48 h to recover. Cells were subsequently
serum-starved for assaying. A Dual-Luciferase Assay kit (Promega
Corporation) was used to detect luciferase activities according to
the manufacturer's protocol. Lysed transfected cells were
centrifuged at 72,000 × g for 120 min at 4°C in Eppendorf
microcentrifuge tubes. A Modulus™ TD20/20 luminometer (Turner
Designs) was used to determine relative luciferase activities.
Luciferase activity was normalized to Renilla luciferase
activity. Three biological repeats were performed.
Bioinformatics analysis
UALCAN (11)
(http://ualcan.path.uab.edu/) is an
online interactive resource, which also provides easy access to
publicly available cancer omics data [The Cancer Genome Atlas
(TCGA), Metastasis 500, Clinical Proteomic Tumor Analysis
Consortium (CPTAC) and Children's Brain Tumor Tissue Consortium].
UALCAN was used to determine the mRNA and protein expression levels
of ZNF384 in primary colon adenocarcinoma cases using data obtained
from TCGA and CPTAC. Gene Expression Profiling Interactive Analysis
(GEPIA; http://gepia2.cancer-pku.cn; version,
2) is an open-access online tool for the interactive evaluation of
RNA sequencing data from 9,736 tumors and 8,587 healthy samples in
TCGA and Genotype-Tissue Expression programs (12). GEPIA2 was also used to evaluate the
association between ZNF384 expression levels, and HIF-1α and MMP2
expression levels in colon or rectum adenocarcinoma. The University
of California Santa Cruz (UCSC) Genome Browser (13) (http://genome.ucsc.edu) is a popular web-based tool
for quickly displaying a requested portion of a genome at any
scale, accompanied by a series of aligned annotation ‘tracks’. UCSC
Genome Browser was used to find the promoter sequence of MMP2 and
ZNF384. JASPAR (14) website
(https://jaspar.genereg.net/) was used to
analyze the MMP2 and ZNF384 promoter (accessed on 10 October 2019).
The relative profile score threshold was set at 85%. Data were
output after calculation through the JASPAR website.
RT-PCR array
Total RNA was extracted from SW480-ZNF384 and
SW480-Control cells using TRIzol according to the manufacturer's
protocol and reverse transcribed to cDNA using PrimeScript RT
Reagent kit (Takara Bio, Inc.) according to the manufacturer's
protocol. Subsequently, RT-PCR was carried out using the Human
Tumor Metastasis RT2 Profiler PCR array (SuperArray Bioscience) in
an ABI PRISM7900 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
The PCR cycling conditions were set as follows: 95°C for 5 min;
followed by 40 cycles at 95°C for 15 sec, 60°C for 15 sec and 72°C
for 20 sec, and a final extension step at 72°C for 5 min. The
results were analyzed using the ∆∆Cq method as aforementioned.
Statistical analysis
Continuous data are displayed as the mean ± standard
deviation, and TCGA and CPTAC data are displayed as median and
interquartile range. The χ2 test was used to analyze
categorical data. Comparison of means between and among groups was
carried out using Student's t-tests and one-way ANOVA followed by
Tukey's or Dunnett's post hoc tests, respectively. Comparison of
means among 50 matched primary CRC, lymph node metastatic and
healthy tissue samples was carried out using repeated measures
ANOVA followed by Tukey's post hoc tests. Based on variables from
univariate analyses, determination of independent factors
influencing survival was carried out using the Cox proportional
hazards model. Kaplan-Meier was used for survival analysis
following surgery, and the log-rank test was performed for
comparisons of survival outcomes. Correlations between ZNF384 and
MMP2 or HIF-1α expression levels in CRC samples were assessed using
Spearman's rank correlation analysis. SPSS software (version, 19;
IBM Corp.) was used for statistical analyses, and GraphPad Prism
(version, 9; GraphPad Software, Inc.) was used for graph
preparation. P<0.05 was considered to indicate a statistically
significant difference.
Results
ZNF384 expression is markedly
increased in human CRC tissues
To explore the functions of ZNF384 in CRC
development, the expression levels of ZNF384 in TCGA and CPTAC
datasets were evaluated. Compared with in healthy tissues, ZNF384
expression levels were significantly elevated in colon
adenocarcinoma tissues (Fig. 1A and
D). TCGA samples were categorized into two groups: i) High
expression (with TPM values above the upper quartile) and ii)
Low/Medium expression (with TPM values below the upper quartile).
The upper quartile of ZNF384 expression in TCGA samples was 36.407.
Based on TCGA datasets, patients with elevated ZNF384 expression
levels exhibited shorter overall survival outcomes compared with
those with lower ZNF384 expression levels, implying that ZNF384 may
be involved in CRC progression (Fig.
1F). The mRNA expression levels of ZNF384 were also measured in
50 matched primary CRC, lymph node metastatic and healthy tissue
samples. The results shown in Fig.
1B demonstrated that compared with in matched healthy tissues,
ZNF384 mRNA expression levels were markedly increased in primary
CRC tissues. Moreover, compared with in primary CRC tissues, ZNF384
mRNA expression levels were markedly elevated in lymph node
metastatic tissues. Consistent with the aforementioned findings,
ZNF384 protein expression levels were markedly elevated in CRC
tissues compared with in adjacent healthy tissues (Figs. 1C, E and S1A).
 | Figure 1.High levels of ZNF384 are predictors
for poor prognostic outcomes in patients with CRC. (A)
Representative data from TCGA datasets displaying the mRNA
expression levels of ZNF384 in colon cancer tissues compared with
in healthy tissues. Box-and-whisker plots represent medians
(horizontal lines), interquartile ranges (boxes), and minimum and
maximum values (whiskers). (B) mRNA expression levels of ZNF384 in
normal, primary CRC and lymph node metastatic tissues. Expression
levels of ZNF384 mRNA were determined by RT-qPCR and normalized to
GAPDH. (C) Western blot analyses were conducted using human CRC and
adjacent normal tissues. (D) ZNF384 protein expression levels in
different tumor types and corresponding healthy tissues from
UALCAN. (E) Immunohistochemical staining for the detection of
ZNF384 in CRC tissues. Scale bars, 100 µm (upper panel) and 50 µm
(lower panel). (F) Results of the Kaplan-Meier analysis
demonstrated the association between ZNF384 mRNA expression levels
and overall survival for patients with COAD in TCGA dataset. (G)
Results of the Kaplan-Meier survival curves demonstrated the
association between ZNF384 expression and overall survival outcomes
in patients with CRC. (H and I) RT-qPCR and western blot analysis
of ZNF384 expression levels in various CRC cell lines. *P<0.05,
**P<0.01 vs. FHC; ##P<0.01 vs. SW480;
&P<0.05, &&P<0.01 vs.
Caco-2; aP<0.05, aaP<0.01 vs. HT29;
bP<0.05 vs. HCT116. ZNF384, zinc finger protein 384;
CRC, colorectal cancer; COAD, colon adenocarcinoma; N, normal; T,
tumor; TCGA, The Cancer Genome Atlas; CPTAC, Clinical Proteomic
Tumor Analysis Consortium; RT-qPCR, reverse
transcription-quantitative PCR. |
Subsequently, ZNF384 expression levels and the
potential clinical significance were evaluated in a cohort of CRC
tissues using IHC staining. Elevated ZNF384 expression levels were
positively associated with poor tumor differentiation, high lymph
node metastasis and an increased American Joint Committee on Cancer
stage (15), and elevated ZNF384
expression levels were an independent risk factor for CRC
progression (Tables I and II). Patients with high expression of
ZNF384 exhibited markedly poor survival outcomes compared with
patients with low expression of ZNF384 (Fig. 1G). In addition, patients with high
expression of HIF-1α and MMP2 exhibited markedly poor survival
outcomes compared with patients with low expression of HIF-1α and
MMP2 (Fig. S2). These results
suggested that ZNF384 expression levels were elevated in CRC and
may be associated with poor prognostic outcomes.
 | Table I.Associations between ZNF384
expression and clinicopathological parameters in 164 patients with
colorectal cancer. |
Table I.
Associations between ZNF384
expression and clinicopathological parameters in 164 patients with
colorectal cancer.
|
|
| ZNF384
expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
variables | Number (n=164) | High (n=66) | Low (n=98) | P-value |
|---|
| Age, years |
|
|
| 0.101 |
|
<60 | 55 | 27 | 28 |
|
|
≥60 | 109 | 39 | 70 |
|
| Sex |
|
|
| 0.655 |
|
Male | 83 | 32 | 51 |
|
|
Female | 81 | 34 | 47 |
|
| T stage |
|
|
| 0.306 |
| T1 and
T2 | 57 | 26 | 31 |
|
| T3 and
T4 | 107 | 40 | 67 |
|
| Lymphatic
invasion |
|
|
|
<0.01a |
|
Absent | 114 | 32 | 82 |
|
|
Present | 50 | 34 | 16 |
|
| AJCC stage |
|
|
|
<0.01a |
| I | 47 | 9 | 38 |
|
| II | 67 | 27 | 40 |
|
|
III | 50 | 30 | 20 |
|
|
Differentiation |
|
|
| 0.016b |
|
Well | 35 | 15 | 20 |
|
|
Moderate | 73 | 21 | 52 |
|
|
Poor | 56 | 30 | 26 |
|
| Tumor location |
|
|
| 0.651 |
|
Colon | 76 | 32 | 44 |
|
|
Rectum | 88 | 34 | 54 |
|
| Table II.Univariate and multivariate analysis
of factors associated with survival in patients with colorectal
cancer. |
Table II.
Univariate and multivariate analysis
of factors associated with survival in patients with colorectal
cancer.
|
Characteristics | Univariate analysis
HR (95% CI) | P-value | Multivariate
analysis HR (95% CI) | P-value |
|---|
| Age, years |
|
|
|
|
| (≥60 vs.
<60) | 2.605
(0.9748-6.96) | 0.056 |
|
|
| Sex |
|
|
|
|
| (female vs.
male) | 0.986
(0.4466-2.179) | 0.973 |
|
|
| T stage |
|
|
|
|
| (T3 and T4 vs. T1
and T2) | 1.114
(0.4691-2.647) | 0.806 |
|
|
| Lymphatic
invasion |
|
|
|
|
| (present vs.
absent) | 5.931
(2.034-17.29) | 0.001 | 3.849
(1.201-12.302) | 0.022 |
|
Differentiation |
|
|
|
|
| (poor vs.
well/moderate) | 1.817
(0.8395-3.934) | 0.130 |
|
|
| AJCC stage |
|
|
|
|
| (III vs. I and
II) | 3.088
(1.29-7.395) | 0.010 | 0.998
(0.360-2.740) | 0.897 |
| Tumor location |
|
|
|
|
| (colon vs.
rectum) | 0.995
(0.986-1.018) | 0.549 |
|
|
| ZNF384
expression |
|
|
|
|
| (high vs. low) | 2.478
(1.102-5.574) | 0.028 | 1.864
(1.450-2.380) | 0.012 |
| HIF-1α
expression |
|
|
|
|
| (high vs. low) | 1.910
(1.020-3.575) | 0.043 | 2.146
(0.881-5.230) | 0.093 |
| MMP2
expression |
|
|
|
|
| (high vs. low) | 3.946
(1.052-14.804) | 0.042 | 3.472
(1.134-10.631) | 0.029 |
ZNF384 enhances CRC cell invasion and
metastasis
The results of the present study demonstrated that
ZNF384 expression was closely associated with tumor metastasis.
SW480 and SW620 represented cells with low and high metastatic
ability, respectively, and ZNF384 was differentially expressed in
these two cell lines (Fig. 1H and
I). Therefore, the present study selected these two cell lines
to study the effect of ZNF384 on the proliferation of colorectal
cancer cells. Notably, ZNF384 expression exerted no significant
effect on the proliferation of SW480 and SW620 CRC cells (Fig. S3). Thus, the effect of ZNF384
expression on the migratory and invasive capacity of CRC cells was
investigated. Both the mRNA and protein expression levels of ZNF384
were measured in various CRC cell lines, including a normal colonic
cell line (FHC), and cell sublines with weak metastatic ability
(SW480, Caco-2, HCT116 and HT29) or strong metastatic ability
(SW620 and LoVo). Consistent with the clinical findings, the
expression levels of ZNF384 were significantly elevated in the six
CRC cell lines compared with in the healthy colonic cell line
(Figs. 1H, I and S1B). In the six CRC cell lines, ZNF384
expression was significantly elevated in metastatic CRC cells (LoVo
and SW620) compared with in non-metastatic cell lines (SW480,
Caco-2, HT29 and HCT116). In addition, the trend in HIF-1α
expression in CRC cells under conditions of normoxia were
inconsistent with the expression levels of ZNF384 and MMP2; under
normoxic conditions, HIF-1α was highly expressed in HT29 and HCT116
cells, whereas ZNF384 and MMP2 were highly expressed in SW620 and
LoVo cells (Figs. S4, S1O, S1P
and 1I). These findings indicated that ZNF384 may have a vital role
in CRC metastasis (Fig. 1H and I).
Subsequently, four stable cell lines were used to investigate the
role of ZNF384 in CRC progression (Figs. 2A-C, S1C and D). The levels of in vitro
CRC cell migration and invasion were investigated using Transwell
assays. The results displayed in Fig.
2D demonstrated that ZNF384 overexpression was associated with
a high number of migrating and invading cells; however, ZNF384
knockdown led to a notable decrease in the levels of cell migration
and invasion. These findings were consistent with those of the
wound healing assays (Fig. S5).
Therefore, the results of the present study highlighted that ZNF384
promoted CRC cell migration and invasion.
To further establish the role of ZNF384 in
metastasis, nude mice were injected with either SW480-ZNF384 or
SW620-shZNF384-1 cells, or the corresponding control cells. The
results of the present study demonstrated that mice that had been
injected with cells overexpressing ZNF384 exhibited elevated CRC
metastasis levels, whereas mice injected with cells in which ZNF384
was knocked down exhibited reduced CRC metastasis levels (Fig. 2E). In addition, the SW620-shControl
group exhibited higher metastatic ability, compared with
SW480-Control group. This is due to the difference in the
metastatic ability of the two colorectal cancer cells themselves.
Consistent with the aforementioned elevated lung metastasis levels,
significantly reduced survival periods were observed in
SW480-ZNF384 cell-injected mice compared with those injected with
SW480-control cells (Fig. 2F). The
survival period for SW620-shZNF384 cell-injected mice was
significantly increased compared with the survival period of
control cell-injected mice (Fig.
2F). Further histopathological evaluation of the lungs revealed
a markedly high number of metastatic nodules in
SW480-ZNF384-injected mice, compared with SW480-control-injected
mice. Moreover, SW620-shZNF384 cell-injected mice exhibited
significantly fewer metastatic nodules compared with those injected
with SW620-sh control cells (Fig. 2G
and H). In the tail vein metastasis assay, the nude mice
quickly suffered casualties due to lung metastases, and colon
metastases were rarely found. These results indicated that ZNF384
increased CRC cell invasion and metastasis.
MMP2 is a transcriptional target of
ZNF384
To establish the mechanisms through which ZNF384
enhances CRC progression, a Metastasis RT2 Profiler PCR Array
analysis was carried out to detect ZNF384 overexpression-associated
transcriptomic variations in SW480 cells. By using a fold change of
two as the cut off, 23 of the 89 metastasis-associated genes were
elevated upon overexpression of ZNF384 in SW480 cells, 17 genes
were suppressed and 49 genes did not exhibit any significant
variations (Table SIII). Among
the elevated genes, MMP2 had the strongest induction response to
ZNF384 overexpression (Table
SIII). Moreover, ZNF384, a transcriptional factor that exhibits
a C2H2 ZNF structure, regulates MMP1, collagen α-1 chain precursor
and MMP3 gene transcription levels (16–18).
The transcriptional recognition site has an adenine
enriched-sequence (15). By
querying the UCSC Genome Browser, it was revealed that the promoter
region of MMP2 is rich in adenine. Therefore, it was hypothesized
that MMP2 was a target gene of ZNF384. To confirm whether ZNF384
activated MMP2 expression, the MMP2 mRNA and protein expression
levels were evaluated in cells following either ZNF384 silencing or
overexpression. ZNF384 overexpression significantly elevated the
MMP2 mRNA expression levels, whereas the mRNA expression levels of
MMP2 were markedly suppressed following ZNF384 knockdown (Fig. 3A and B). The changes in MMP2
protein expression levels were consistent with the aforementioned
findings (Figs. 3C, S1E and F). To establish whether ZNF384
mediated MMP2 transcription activation, constructs expressing the
MMP2 promoter were co-transfected with pCMV-ZNF384, and the
relative luciferase activity was determined. The results of the
present study demonstrated that luciferase activity was markedly
elevated in MMP2 promoter-transfected cells (Fig. 3D), indicating that ZNF384 activated
MMP2 transcription. Moreover, sequence analysis via JASPAR revealed
the presence of three putative ZNF384-binding sites in the MMP2
promoter. Site-directed mutagenesis and serial deletion analyses
revealed that binding site 3 is vital for ZNF384-induced MMP2
transactivation (Fig. 3E).
Furthermore, direct binding of ZNF384 to the MMP2 promoter was
further verified using ChIP (Fig.
3F). These results indicated that ZNF384 directly binds to
specific sites on the MMP2 promoter to transcriptionally activate
subsequent gene expression.
MMP2 is crucial for ZNF384-mediated
CRC cell migration and metastasis
Both MMP2 knockdown and overexpression were verified
using RT-qPCR and western blotting (Figs. 4A-C, S1G-J, Q, R and S6). MMP2 knockdown significantly
inhibited ZNF384-induced cell migration and invasion (Figs. 4D and S7); however, MMP2 overexpression
reversed the ZNF384 knockdown-mediated levels of migration and
invasion (Fig. 4E and S7). In vivo, the SW480-ZNF384 +
LV-shMMP2-1 group exhibited poor metastasis, limited lung
metastasis and scarce metastatic nodules, compared with the
SW480-ZNF384 + LV-shControl group (Fig. 4F, I and J). However, MMP2
overexpression reversed the ZNF384 knockdown-induced decrease in
SW620 cell metastasis (Fig. 4F, I and
J). Compared with in mice injected with control cells, those
that were injected with MMP2-overexpressing SW620-shZNF384 cells
exhibited significantly reduced survival periods, whereas those
injected with MMP2-knockdown SW480-ZNF384 cells exhibited
significantly increased survival periods (Fig. 4G and H). These results indicated
that ZNF384 may increase CRC migration and metastasis by promoting
MMP2 expression.
HIF-1α transactivates ZNF384
Hypoxia is common in a number of malignancies,
including CRC. Notably, the expression levels of HIF-1α and −2α
have been reported to be elevated in numerous types of human
cancer, and the expression levels of these proteins are associated
with poor prognostic outcomes (19). The role of HIF-1α in cancer
progression and the corresponding implications in CRC metastasis
have previously been reported (20,21).
During hypoxia, HIF-1α activates downstream genes involved in
various cancer biological processes, such as angiogenesis, glucose
metabolism, cell survival and invasion, to adapt to the hypoxic
environment (22,23). Moreover, MMPs play important roles
in this process, and a previous study indicated that HIF-1α
initiated the expression of MMPs (24). Osinsky et al (25) demonstrated the positive correlation
between hypoxia levels and MMP2, as well as MMP9, in Lewis lung
carcinoma (25). Exposure of
neonatal mice to chronic hypoxia (10% O2), an inducer
for lung development arrest, also led to elevated MMP2 levels with
corresponding suppression of MMP inhibitor 2 precursor (26). Under hypoxic conditions (0.5%
O2), the ZNF384 expression levels in Caco-2 and SW480
cells were markedly elevated in a time-dependent manner (Figs. 5A, B, S1K and L). To investigate whether
hypoxia-mediated ZNF384 expression was involved in the
transactivation of its promoter, a reporter plasmid with the ZNF384
gene promoter was transfected into CRC cell lines and subjected to
hypoxia. Under conditions of hypoxia, ZNF384 transcription levels
were markedly elevated, demonstrated by enhanced luciferase
activities (Fig. 5C). HIF-1α
overexpression was associated with high luciferase promoter
activities, and elevated ZNF384 mRNA and protein expression levels,
whereas HIF-1α knockdown exerted the opposite effects (Figs. 5D, S1M and N). Moreover, the ZNF384 promoter
sequences were explored to establish potential cis-regulatory
elements, and two putative HIF-1α-binding sites were identified
through the JASPAR website. Site-directed mutagenesis and serial
deletion assays revealed that both ZNF384-binding sites were
crucial for HIF-1α-initiated ZNF384 transactivation (Fig. 5E). Results of the ChIP assay
further verified the direct binding of HIF-1α to the ZNF384
promoter in CRC cells (Fig. 5F).
The aforementioned findings demonstrated that ZNF384 is a target
gene for HIF-1α.
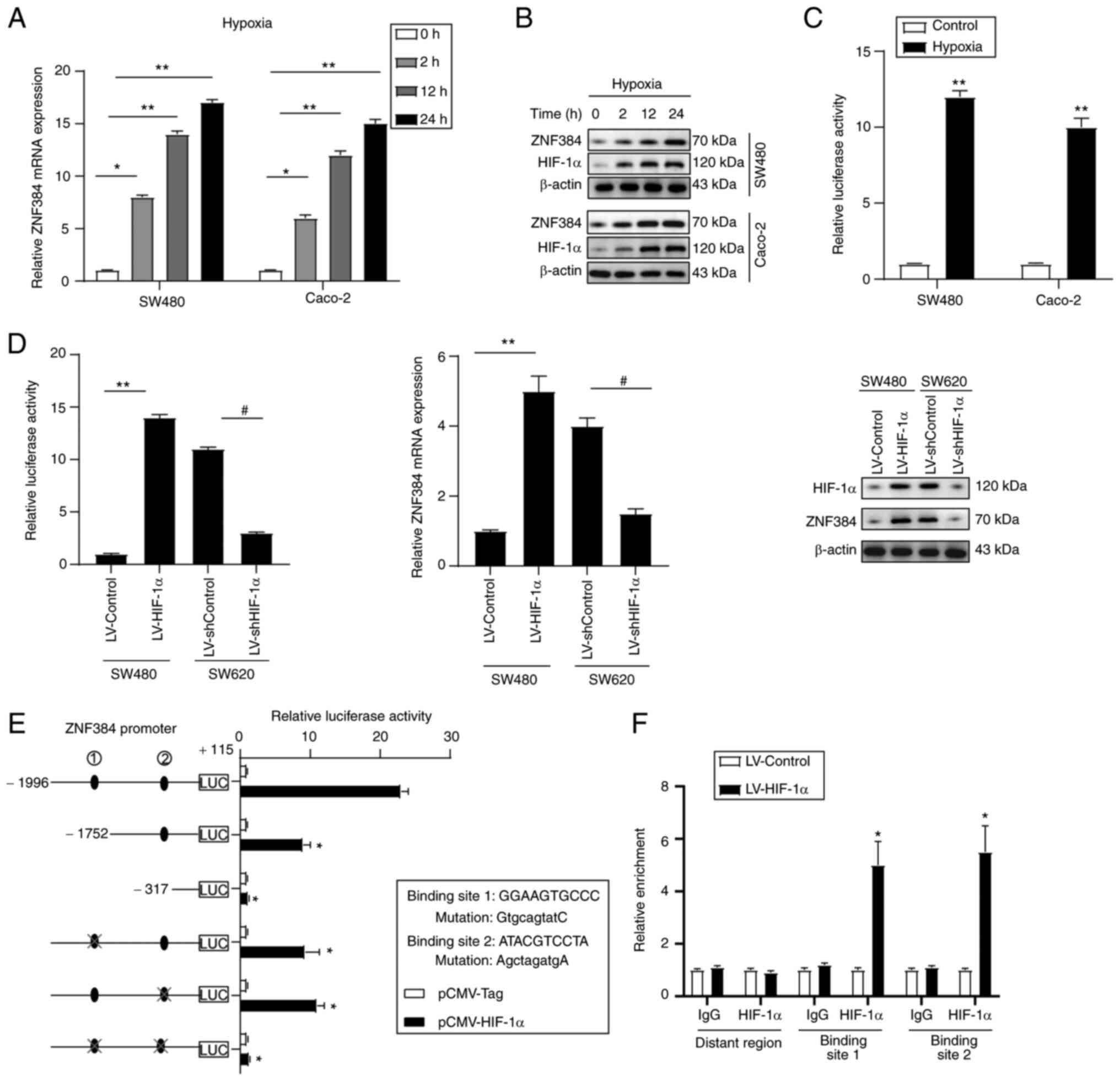 | Figure 5.ZNF384 is a direct target gene for
HIF-1α. CRC cells were cultured in a hypoxic atmosphere (0.5%
O2) for the indicated time intervals, after which ZNF384
expression was evaluated by (A) RT-qPCR and (B) western blot
analysis. *P<0.05, **P<0.01 vs. 0 h. (C) Luciferase reporter
construct with the (−1,996/+115) ZNF384 promoter was transfected in
CRC cells, and luciferase activity was measured after 24 h.
**P<0.01 vs. Control. (D) SW480 and SW620 cells were infected
with LV-HIF-1α and LV-shHIF-1α. After 24 h, ZNF384 transcription
and expression levels were assessed using luciferase assays (left
panel), western blotting (right panel) and RT-qPCR (middle panel).
**P<0.01 vs. LV-Control; #P<0.05 vs. LV-shControl.
(E) Truncated and mutated ZNF384 promoter constructs were
co-transfected with pCMV-HIF-1α in SW480 cells, after which
relative luciferase activities were confirmed. *P<0.05 vs.
ZNF384 promoter sequence (−1,996/+115). (F) Chromatin
immunoprecipitation assays confirmed the binding of HIF-1α to the
ZNF384 promoter in SW480 cells. *P<0.05 vs. anti-IgG. ZNF384,
zinc finger protein 384; CRC, colorectal cancer; RT-qPCR, reverse
transcription-quantitative PCR; HIF-1α, hypoxia-inducible factor
1α; sh, short hairpin RNA. |
ZNF384 expression is positively
associated with HIF-1α and MMP2 expression levels in CRC
The potential association between ZNF384, MMP2 or
HIF-1α expression in CRC tissues was evaluated. Results of the
present study demonstrated that there was a positive association
between ZNF384 and HIF-1α expression, and between ZNF384 and MMP2
expression (Fig. 6A and B), which
was further verified using GEPIA (Fig.
6C).
Discussion
In numerous types of cancer, ZNF384 promotes various
malignant processes, including cell proliferation, migration and
invasion (9,27,28).
Results of a previous study demonstrated that ZNF384 overexpression
promoted melanoma cell progression (9). In addition, ZNF384 has been shown to
bind the APOBEC3B (A3B) promoter to modulate A3B expression in
cervical cancer (27). ZNF384 may
also promote hepatocellular carcinoma (HCC) cell proliferation by
promoting cyclin D1 expression and is considered a prognostic
factor for patients with HCC (28).
The results of the present study demonstrated that
ZNF384 was markedly elevated in metastatic CRC tissues, and
increased ZNF384 expression was associated with lymph node
metastasis and advanced clinical stage. ZNF384 expression was also
revealed to be an independent risk factor for low patient survival
outcomes following curative resection. Furthermore, the results of
the present study demonstrated that ZNF384 overexpression markedly
enhanced CRC metastasis, whereas ZNF384 knockdown exerted the
opposite effects. However, the results of the present study
revealed that ZNF384 expression did not affect CRC cell
proliferation. The discrepancies in the function of ZNF384 in
different types of cancer may be associated with differences in
cellular context or differences in targeted genes. Therefore, in
CRC, ZNF384 may act as a potential metastatic promoter and
prognostic marker.
MMPs are involved in cancer invasion and metastases
(29). Among the MMPs, MMP2, also
referred to as gelatinase A, plays critical roles in malignant cell
migration, which is attributed to its ability to degrade type IV
collagen (30). MMP2 is a
potential prognostic biomarker (31). Elevated MMP2 expression in cancer
cells has been reported to be a significant predictive factor for
poor survival outcomes in CRC (31,32).
Kostova et al (33)
demonstrated a significant positive correlation between MMP2 tissue
expression and the presence of nodal metastasis in CRC. Increased
expression levels of MMP2 have also been shown to be correlated
with poor overall and progression-free survival in patients with
CRC (34). In CRC, increased
levels of MMP2 have been revealed to be associated with
microvascular angiogenesis and apoptotic resistance, and increased
levels of MMP2 could elevate the levels of cell adhesion, and
promote invasion and metastasis (35). The results of intrasplenic
injection assays carried out in a previous study demonstrated that,
in nude mice, the metastatic potential of CRC cell lines was
associated with the levels of secreted MMP2 (36). These findings highlighted that MMP2
may be a crucial oncogene in CRC; however, the mechanisms through
which MMP2 is dysregulated in CRC are yet to be fully elucidated.
The results of the present study demonstrated that MMP2 was a
direct functional target of ZNF384, which transactivates MMP2
expression by binding its promoter. Suppression of MMP2 markedly
inhibited ZNF384-mediated CRC migration, invasion and lung
metastasis, whereas MMP2 overexpression reversed the ZNF384
knockdown-induced suppression of CRC cell malignancy. Moreover,
ZNF384 expression exhibited a positive correlation with MMP2
expression. In the present study, in addition to using serial
deletion, site-directed mutagenesis was used as a control. Compared
with the mutant plasmid, the wild-type plasmid had almost the same
structure except the mutant base, which can also exclude the
influence of nucleosomes. Therefore, these findings indicated that
ZNF384 increased CRC metastasis by transactivating MMP2.
The mechanisms through which ZNF384 and MMP2 were
dysregulated in CRC were also investigated. The association between
hypoxia and MMPs, such as MMP2, has previously been verified
(24). However, the role of
hypoxia in ZNF384-associated MMP2 dysregulation has yet to be fully
elucidated in human CRC. The results of the present study
demonstrated two potential HIF-1α-binding sites in the ZNF384
promoter. Further analyses revealed that HIF-1α transactivated
ZNF384 by binding both HIF-1α-binding sites within the ZNF384
promoter. Moreover, HIF-1α expression was correlated with ZNF384
expression levels. However, the results of the present study
demonstrated that the trend in HIF-1α expression in CRC cells under
conditions of normoxia were inconsistent with the expression levels
of ZNF384 and MMP2. Thus, the ZNF384/MMP2 axis may not be regulated
by HIF-1α under conditions of normoxia. Results of previous studies
have demonstrated that hypoxia induces the Warburg effect, which is
key in the development of cancer and may alter glucose metabolism
(22,23). In the present study, the role of
the HIF-1α/ZNF384/MMP2 axis in CRC progression has yet to be fully
elucidated; thus, further investigations are required. The
aforementioned results of the present study highlighted that the
hypoxia-associated protein, HIF-1α, is a transcriptional regulator
of ZNF384, which may enhance both MMP2 expression and CRC cell
metastasis. These results highlight that under hypoxic conditions,
human CRC progression may be promoted by dysregulated MMP2.
In conclusion, ZNF384, a direct HIF-1α target, was
revealed to be markedly elevated in CRC and to be associated with
poor prognostic outcomes. Furthermore, ZNF384 overexpression
enhanced CRC cell metastasis by transactivating MMP2 expression.
Therefore, these findings indicated that ZNF384 may act as a
potential prognostic factor and therapeutic target for CRC.
Supplementary Material
Supporting Data
Acknowledgments
Not applicable.
Funding
The present study was supported by the Sichuan Youth Science and
Technology Foundation (grant no. 2017JQ0039).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZY, YZ and YY designed the study, analyzed and
interpreted the data, and wrote the manuscript. CZ analyzed and
interpreted the data. PL and HT analyzed and interpreted the data,
and wrote the manuscript. XT and GZ interpreted the data and
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of North Sichuan Medical College [approval no.
2021ER(A)007]. All patients provided written informed consent for
participation in the present study. The animal experiments were
approved by the Experimental Animal Ethics Committee of North
Sichuan Medical College (approval no. 20200908).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fakih MG: Metastatic colorectal cancer:
Current state and future directions. J Clin Oncol. 33:1809–1824.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Punt CJ, Koopman M and Vermeulen L: From
tumour heterogeneity to advances in precision treatment of
colorectal cancer. Nat Rev Clin Oncol. 14:235–246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sato J, Nakamura M, Watanabe O, Yamamura
T, Funasaka K, Ohno E, Miyahara R, Kawashima H, Goto H and Hirooka
Y: Prospective study of factors important to achieve observation of
the entire colon on colon capsule endoscopy. Therap Adv
Gastroenterol. 10:20–31. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamoto T, Yamagata T, Sakai R, Ogawa S,
Honda H, Ueno H, Hirano N, Yazaki Y and Hirai H: CIZ, a zinc finger
protein that interacts with p130(cas) and activates the expression
of matrix metalloproteinases. Mol Cell Biol. 20:1649–1658. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhong CH, Prima V, Liang X, Frye C,
McGavran L, Meltesen L, Wei Q, Boomer T, Varella-Garcia M, Gump J
and Hunger SP: E2A-ZNF384 and NOL1-E2A fusion created by a cryptic
t(12;19)(p13.3; p13.3) in acute leukemia. Leukemia. 22:723–729.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Do Amaral A: Complexities of nomenclature
in biology. Gender of generic names ending in ops z, n, (s) 1572.
Mem Inst Butantan. 39:27–36. 1975.(In Portuguese). PubMed/NCBI
|
|
8
|
Lilljebjörn H and Fioretos T: New
oncogenic subtypes in pediatric B-cell precursor acute
lymphoblastic leukemia. Blood. 130:1395–1401. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sakuma T, Nakamoto T, Hemmi H, Kitazawa S,
Kitazawa R, Notomi T, Hayata T, Ezura Y, Amagasa T and Noda M:
CIZ/NMP4 is expressed in B16 melanoma and forms a positive feedback
loop with RANKL to promote migration of the melanoma cells. J Cell
Physiol. 227:2807–2812. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47:W556–W560. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kent WJ, Sugnet CW, Furey TS, Roskin KM,
Pringle TH, Zahler AM and Haussler D: The human genome browser at
UCSC. Genome Res. 12:996–1006. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Castro-Mondragon JA, Riudavets-Puig R,
Rauluseviciute I, Berhanu Lemma R, Turchi L, Blanc-Mathieu R, Lucas
J, Boddie P, Khan A, Manosalva Pérez N, et al: JASPAR 2022: The 9th
release of the open-access database of transcription factor binding
profiles. Nucleic Acids Res. 30:gkab11132021.
|
|
15
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Khemlina G, Ikeda S and Kurzrock R: The
biology of Hepatocellular carcinoma: Implications for genomic and
immune therapies. Mol Cancer. 16:1492017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fan Z, Tardif G, Hum D, Duval N, Pelletier
JP and Martel-Pelletier J: Hsp90{beta} and p130(cas): Novel
regulatory factors of MMP-13 expression in human osteoarthritic
chondrocytes. Ann Rheum Dis. 68:976–982. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Torrungruang K, Alvarez M, Shah R, Onyia
JE, Rhodes SJ and Bidwell JP: DNA binding and gene activation
properties of the Nmp4 nuclear matrix transcription factors. J Biol
Chem. 277:16153–16159. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keith B, Johnson RS and Simon MC: HIF1α
and HIF2α: Sibling rivalry in hypoxic tumour growth and
progression. Nat Rev Cancer. 12:9–22. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Finger EC and Giaccia AJ: Hypoxia,
inflammation, and the tumor microenvironment in metastatic disease.
Cancer Metastasis Rev. 29:285–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoshimura H, Dhar DK, Kohno H, Kubota H,
Fujii T, Ueda S, Kinugasa S, Tachibana M and Nagasue N: Prognostic
impact of hypoxia-inducible factors 1alpha and 2alpha in colorectal
cancer patients: Correlation with tumor angiogenesis and
cyclooxygenase-2 expression. Clin Cancer Res. 10:8554–8560. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Renga G, Oikonomou V, Moretti S,
Stincardini C, Bellet MM, Pariano M, Bartoli A, Brancorsini S,
Mosci P, Finocchi A, et al: Thymosin β4 promotes autophagy and
repair via HIF-1α stabilization in chronic granulomatous disease.
Life Sci Alliance. 2:e2019004322019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu WL, Wang SH, Sun WB, Gao J, Ding XM,
Kong J, Xu L and Ke S: Insufficient radiofrequency ablationinduced
autophagy contributes to the rapid progression of residual
hepatocellular carcinoma through the HIF-1α/BNIP3 signaling
pathway. BMB Rep. 52:277–282. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Ni Z, Duan Z, Wang G and Li F:
Altered expression of hypoxia-inducible factor-1α (HIF-1α) and its
regulatory genes in gastric cancer tissues. PLoS One. 9:e998352014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Osinsky SP, Ganusevich II, Bubnovskaya LN,
Valkovskaya NV, Kovelskaya AV, Sergienko TK and Zimina SV: Hypoxia
level and matrix metalloproteinases-2 and −9 activity in Lewis lung
carcinoma: Correlation with metastasis. Exp Oncol. 27:202–205.
2005.PubMed/NCBI
|
|
26
|
Ryu J, Vicencio AG, Yeager ME, Kashgarian
M, Haddad GG and Eickelberg O: Differential expression of matrix
metalloproteinases and their inhibitors in human and mouse lung
development. Thromb Haemost. 94:175–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mori S, Takeuchi T, Ishii Y and Kukimoto
I: Identification of APOBEC3B promoter elements responsible for
activation by human papillomavirus type 16 E6. Biochem Biophys Res
Commun. 460:555–560. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He L, Fan X, Li Y, Chen M, Cui B, Chen G,
Dai Y, Zhou D, Hu X and Lin H: Overexpression of zinc finger
protein 384 (ZNF 384), a poor prognostic predictor, promotes cell
growth by upregulating the expression of Cyclin D1 in
Hepatocellular carcinoma. Cell Death Dis. 10:4442019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shay G, Lynch CC and Fingleton B: Moving
targets: Emerging roles for MMPs in cancer progression and
metastasis. Matrix Biol. 44–46. 200–206. 2015.PubMed/NCBI
|
|
30
|
Nagase H and Woessner JF Jr: Matrix
metalloproteinases. J Biol Chem. 274:21491–21494. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zucker S and Vacirca J: Role of matrix
metalloproteinases (MMPs) in colorectal cancer. Cancer Metastasis
Rev. 23:101–117. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Leeman MF, Curran S and Murray GI: New
insights into the roles of matrix metalloproteinases in colorectal
cancer development and progression. J Pathol. 201:528–534. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kostova E, Slaninka-Miceska M, Labacevski
N, Jakovski K, Trojachanec J, Atanasovska E, Janevski V, Jovanovik
R and Janevska V: Expression of matrix metalloproteinases 2, 7 and
9 in patients with colorectal cancer. Vojnosanit Pregl. 71:52–59.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shi M, Yu B, Gao H, Mu J and Ji C: Matrix
metalloproteinase 2 overexpression and prognosis in colorectal
cancer: A meta- analysis. Mol Biol Rep. 40:617–623. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shah V, Kumar S and Zirvi KA: Metastasis
of human colon tumor cells in vivo: Correlation with the
overexpression of plasminogen activators and 72 kDa gelatinase. In
Vivo. 8:321–326. 1994.PubMed/NCBI
|


















