Introduction
Ovarian, endometrial and cervical cancer are the
most common malignant tumors in the female reproductive system, and
they seriously threaten the mental and physical health of women.
Among the three cancers, endometrial cancer makes up 30% of
sarcomas of the female genital tract (1), and its incidence is rising (2). However, the cause and pathogenesis of
endometrial cancer are not very clear. Most endometrial cancers are
estrogen-dependent, such as endometrioid adenocarcinoma; the others
are estrogen-independent, such as clear cell tumor. Ovarian cancer
is less common than endometrial and cervical cancer, but its
mortality is the highest (3). The
clinical symptoms of ovarian cancer are so atypical that its
diagnosis is usually in the terminal stages, leading to the 5-year
survival rate reducing to 15–30% (2). Therefore, it is a hot topic about the
research on the etiology and pathogenesis of ovarian and
endometrial cancer.
Programmed cell death 4 (PDCD4) was initially
associated with cell apoptosis (4). Previously, PDCD4 was regarded as an
important tumor suppressor that was lowly expressed or deleted in
numerous human types of cancer, including ovarian and endometrial
cancer (5), and the rate of
protein downregulation or loss is higher than that of mRNA decrease
or deletion in certain tumors. The low expression of PDCD4 is
related to the differentiation degree of tumor cells and prognosis
of some patients with tumors, suggesting that PDCD4 may be involved
in the process of tumor development. It has been reported that
PDCD4 overexpression could effectively suppress the migration and
invasion, as well as the proliferation of cells in numerous types
of cancer (6), such as breast
(7), ovarian (8–10)
and endometrial cancer (11). The
expression level of PDCD4 is also related to the sensitivity of
tumor cells to chemotherapeutic drugs. Upregulation of PDCD4 may
increase the sensitivity of tumor cells to chemotherapeutic drugs
(12). However, the reason for the
low expression of PDCD4 in human types of cancer remains unclear.
Previous studies reported that the downregulation of PDCD4 at the
mRNA level was relevant to the 5′CpG island methylation (13), and the decreased expression of
PDCD4 at the protein level was related to post-transcriptional
processing, such as the regulation of microRNA, the phosphorylation
and ubiquitination of protein (14–16).
TRIM27 is a member of the tripartite motif (TRIM)
family. Since TRIM27 was originally found as the N-terminal fusion
partner with the RET tyrosine kinase proto-oncogene, it was also
called RET finger protein (RFP) (17). The TRIM family members have three
kinds of characteristic domains, a Ring finger domain, one or two
B-box zinc finger domain and a coiled-coil domain (18). Ring finger domain can mediate the
ubiquitin transfer between the proteins themselves or different
substrates, thus it is a characteristic marker of numerous E3
ubiquitin ligases (19). TRIM27
protein can be localized in the nucleus, cytoplasm and cell
membrane depending on different cells, and its nuclear
translocation is regulated by certain signaling pathways (20). Functionally, TRIM27 is involved in
transcriptional regulation, cell apoptosis, cell differentiation,
inflammatory response and cell cycle. TRIM27 is closely related to
the occurrence and development of tumors, and is highly expressed
in numerous types of cancer such as lung (21), breast (22), ovarian (23) and endometrial cancer (24). It has been found that TRIM27 can
promote the migration and invasion of cancer cells. In some tumors,
TRIM27 knockdown can increase the drug-induced apoptosis (25). In addition, TRIM27, as an E3
ubiquitin ligase, downregulates the expression of other proteins
through the ubiquitination and plays an important role in the
regulation of antiviral natural immune responses and
CD4+ T cell-mediated immune responses (26,27).
Based on the aforementioned and previous studies, it
was found that the levels of PDCD4 and TRIM27 have a negative
relationship in multiple tumors, including ovarian and endometrial
cancer. In function, both PDCD4 and TRIM27 are involved in
tumorigenesis and progression, but the two proteins have opposite
roles. Moreover, PDCD4 can be degraded through the ubiquitination,
while TRIM27 has the E3 ubiquitin ligase activity. Therefore, it
was hypothesized that TRIM27 may regulate the expression of PDCD4
by the ubiquitination effect. In the present study, the expression
of PDCD4 and TRIM27 in ovarian and endometrial cancer cell lines
was detected. The impact of TRIM27 overexpression and knockdown on
PDCD4 expression and the effective mechanism of TRIM27 regulating
PDCD4 expression were also investigated in vitro. The
results confirmed that TRIM27 could increase the migration and
invasion of ovarian and endometrial cancer cells by promoting the
ubiquitination and degradation of PDCD4. The present findings may
provide a new target for the treatment of ovarian and endometrial
cancer.
Materials and methods
Antibodies and reagents
Rabbit monoclonal antibody against PDCD4 (cat. no.
9535) was purchased from Cell Signaling Technology, Inc. Rabbit
polyclonal antibody against TRIM27 (cat. no. 12205-1-AP) was
purchased from ProteinTech Group, Inc. Antibodies against HA (cat.
no. H6908) and FLAG (cat. no. F1804) were purchased from
Sigma-Aldrich; Merck KGaA. The β-actin antibody (cat. no. TA-09)
was purchased from ZSGB-BIO. Protein synthesis inhibitor
(cycloheximide, CHX) was purchased from Beyotime Institute of
Biotechnology. Proteasome inhibitor (MG132) was purchased from
MedChemExpress.
Cell culture
Human endometrial cancer cells (Ishikawa) and
ovarian cancer cells (A2780 and SKOV3) were donated by Qilu
Hospital of Shandong University. Ishikawa cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Hyclone; Cytiva), which
was supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), penicillin (100 U/ml) and streptomycin
(100 µg/ml). A2780 and SKOV3 cells were cultured in RPMI-1640
medium (Hyclone; Cytiva) containing 10% FBS, penicillin (100 U/ml)
and streptomycin (100 µg/ml). Human endometrial cancer cells
(HEC-1-A) were purchased from the China Center for Type Culture
Collection. HEC-1-A cells were cultured in McCoy's 5A medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS,
penicillin (100 U/ml) and streptomycin (100 µg/ml). The cells were
cultured in an incubator containing 5% CO2 at 37°C.
The expression vector and small
interference RNA (siRNA)
The expression plasmids of PDCD4, TRIM27 and
ΔRingTRIM27-flag were constructed by Shanghai Genechem Co., Ltd.
The expression plasmids of ubiquitin (UB)-HA, K63-HA and K48-HA
were donated by Dr Wei Zhao (Department of Immunology, School of
Basic Medical Sciences, Shandong University). A total of 3
different TRIM27-specific siRNAs and PDCD4-specific siRNAs were
synthesized by Shanghai GenePharma Co., Ltd. The TRIM27 lentiviral
expression vector and lentiviral short hairpin RNAs (shRNAs) were
also constructed by Shanghai Genechem Co., Ltd. Specific siRNA
sequences for PDCD4, TRIM27 and control siRNAs sequences are listed
in Table SI. Specific shRNA
sequences for TRIM27 and control short hairpin (sh) RNA sequences
are listed in Table SII.
Transfection of expression plasmids (4 µg) or siRNAs (4 µl) was
performed using 10 µl Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol for 24
or 48 h at 37°C, RT-qPCR and western blotting were used to detect
corresponding mRNA and protein. TRIM27 lentiviral expression vector
(Multiplicity of infection, MOI=5) or lentiviral shRNA specific for
TRIM27 (MOI=20) was respectively transfected to A2780 or Ishikawa
cells using LipofiterTM liposome transfection reagent according to
the manufacturer's protocol (Hanbio Biotechnology Co., Ltd.). The
transfection efficiency was detected by RT-qPCR and western blot
analysis.
RNA extraction and RT-qPCR
Total RNA of all cells was extracted by RNA fast
2000 (Feijie Biological Technology), and then reversely
transcribed to cDNA using Reverse-Transcribe kit according to the
manufacturer's protocol (Tiangen Biotech Co., Ltd.). qPCR was
performed with UltaSYBR Mixture (Kangweishiji Biotech Co., Ltd.)
and different primers. The primer sequences are listed in Table SIII. The thermocycling conditions
were as follows: first, pre-denaturation at 95°C for 10 min,
followed by 39 cycles of amplification at 95°C for 15 sec, 60°C for
1 min and 65°C for 5 sec. GAPDH was used as the reference gene.
Each experiment was repeated at least three times. The data
analysis was according to the 2−∆∆Cq method (28).
Western blot analysis
All cells were washed by PBS and lysed by RIPA
buffer (Beyotime Institute of Biotechnology) for protein
extraction. A bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.) was used to detect the protein concentration. The
same amount (25 µg) of each protein was separated using 10%
SDS-polyacrylamide gel and then transferred to 0.45-µm PDVF
membranes (EMD Millipore). Membranes were blocked with 5% non-fat
milk 2 h at room temperature. Rabbit polyclonal antibody against
TRIM27 or rabbit monoclonal antibody against PDCD4 (both at
1:1,000) were added and incubated with membranes at 4°C overnight.
Following the primary incubation, the membranes were incubated with
a horseradish peroxidase (HRP)-conjugated secondary antibody
(1:5,000; cat. no. 111-035-003; Jackson ImmunoResearch
Laboratories, Inc.) for 50 min at room temperature. The signals
were detected using enhanced chemiluminescence detection system
(Thermo Fisher Scientific, Inc.). ImageJ software (version
1.8.0.172; National Institutes of Health) was used for quantitative
analysis. Each experiment was repeated at least three times.
Immunocytochemistry
The cells were cultured on a coverslip in a 24-well
plate overnight at 37°C. The cells were then washed by PBS and
fixed by 4% paraformaldehyde at room temperature and blocked with
2% bovine serum albumin (Beijing Solarbio Science & Technology
Co., Ltd.) at 37°C for 1 h. Rabbit anti-TRIM27 polyclonal antibody
or mouse anti-PDCD4 monoclonal antibody (1:100; cat. no. sc-376430;
Santa Cruz Biotechnology, Inc.) was added into different wells and
incubated overnight at 4°C. The next day, the cells were washed by
PBS three times and incubated with Alexa Fluor®
488-conjugated anti-human IgG/anti-rabbit IgG (1:200; cat. no.
srbAF488-1) or TRITC-conjugated goat anti-mouse IgG (1:50; cat. no.
SA00007-1) (both from ProteinTech Group, Inc.) for 1 h at 37°C.
Then the cells were washed by PBS and DAPI (5 µg/ml) was added into
each well. The results were observed using a fluorescent
microscope. The experiment was repeated three times. The
specificity of TRIM27 and PDCD4 antibodies was confirmed in the
previous study by establishing negative controls.
Immunohistochemistry (IHC)
Firstly, the paraffin-embedded tissue sections from
the tumor of a xenograft nude mouse model of ovarian cancer used in
a previous study (23) were placed
in an oven at 70°C for 2 h. Then, the sections were dewaxed in
xylene and hydrated in graded alcohol before antigen microwave
retrieval. The sections were washed with 3%
H2O2 for 10 min and blocked with 10% goat
serum (ZSGB-BIO) at 37°C for 15 min, and then incubated with the
primary antibodies specific for TRIM27 (1:200) or PDCD4 (1:200)
overnight at 4°C. The next day, the sections were washed by PBS and
then incubated with HRP-conjugated anti-rabbit IgG (Gene Tech Co.,
Ltd.) at 37°C for 45 min followed by diaminobenzidine (Gene Tech
Co., Ltd.) staining. The nuclei were counterstained with
hematoxylin at room temperature for 5 min. The results were
observed using a light microscope.
Co-immunoprecipitation assay
(co-IP)
TRIM27-specific siRNA and UB-HA plasmid were
transfected into A2780 cells and TRIM27 expression plasmid and
UB-HA plasmid were transfected into Ishikawa cells. A total of 24 h
later, the cells were washed by PBS twice, then lysed with 400 µl
of IP buffer [components: NaCl, 4.4 g; NP-40, 5 ml; Tris-HCl (pH
7.4), 50 ml; EDTA, 50 ml; ddH2O, 395 ml]. Then the cells
were lysed at 4°C for 30 min and centrifuged at 14,000 × g for 15
min at 4°C. Protein was divided into two groups, IP group (300 µl)
and Input group (80 µl). IP group protein was incubated with 3 µl
of PDCD4 antibody (1:100) overnight at 4°C in a chromatography
cabinet and Input group protein was stored at −20°C. The next day,
IP group protein was incubated with 40 µl of Protein A/G
Plus-Agarose (Cell Signaling Technology, Inc.) in a chromatography
cabinet at 4°C for 8–12 h. Next, the IP group protein was washed by
IP buffer and centrifuged at 1,000 × g for 5 min at 4°C to collect
the supernatant, which was repeated five times and then the loading
buffer was added. Finally, the IP group protein was placed at 100°C
for 6 min. Input group protein was added the loading buffer and
placed at 100°C for 6 min. The experiment was repeated three
times.
Transwell migration assay
The Ishikawa cells were silenced with the lentiviral
shRNAs targeting TRIM27, and the A2780 cells were overexpressed
with the lentiviral expression vector targeting TRIM27. After
infection, puromycin (3 µg/ml) was used for screening. Western blot
analysis was performed for identification. Then the cells were
overexpressed with PDCD4 plasmid or silenced with PDCD4-specific
siRNA. The cells were cultured at 37°C for 24 h after PDCD4
overexpression and silencing. The cells (5×104/well)
cultured with 1% FBS medium were placed into the upper chamber of a
Transwell membrane with an 8-µm pore size (EMD Millipore). A total
of 650 µl medium with 20% FBS were added into the lower chamber.
After incubation at 37°C for 24 h, the cells were washed with PBS
twice, fixed with 100% methanol at room temperature for 15 min,
then stained with 0.5% crystal violet (Beyotime Institute of
Biotechnology) at room temperature for 30 min. A total of randomly
selected 5 images for each sample were observed using a light
microscope. The experiment was repeated three times.
Matrigel invasion assay
The BD Biocoat Matrigel (BD Biosciences) was used to
detect the effect of TRIM27 and PDCD4 on the invasion ability of
Ishikawa and A2780 cells. A total of 40 µl medium without FBS and
10 µl Matrigel were mixed and added into the chamber. After
incubation at 37°C for 30 min, the cells (8×104/well)
cultured with 1% FBS medium were added into the upper chamber. A
total of 650 µl medium with 20% FBS were added into the lower
chamber. After incubation at 37°C for 48 h, the cells were placed
into methyl alcohol for 15 min, and then stained with 0.5% crystal
violet at room temperature for 30 min. The cells in the upper
chamber were removed by cotton bud. A total of 5 randomly selected
fields of the stained cells in the lower chamber were observed
under a light microscope. The experiment was repeated three
times.
Statistical analysis
GraphPad prism version 6 (GraphPad Software, Inc.)
was used to analyze the data. The results were expressed as the
mean ± SEM. One way ANOVA followed by Tukey's post hoc test was
used for multiple comparisons. The unpaired Student's t-test was
used to evaluate the statistical significance between two groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
PDCD4 is lowly expressed while TRIM27
is highly expressed in endometrial and ovarian cancer cell lines at
the protein level
To investigate the relationship between PDCD4 and
TRIM27 expression, the mRNA and protein expression of PDCD4 and
TRIM27 in endometrial cancer cell lines (Ishikawa and HEC-1-A) and
ovarian cancer cell lines (A2780 and SKOV3) was detected using
RT-qPCR and western blotting. Among the four cell lines, a
relatively low expression of PDCD4 and a relatively high TRIM27
expression was observed in Ishikawa, HEC-1-A and A2780 cells at the
protein level. By contrast, PDCD4 protein expression in SKOV3 was
relatively high, and TRIM27 protein expression was relatively low
(Figs. 1A and S1). However, the results from RT-qPCR
revealed that the expression of TRIM27 and PDCD4 had no obvious
relation at the mRNA level in the aforementioned cell lines
(Fig. 1B and C). The result from
immunofluorescence staining indicated that PDCD4 protein was mainly
distributed around the nucleus in Ishikawa and A2780 cells, and
TRIM27 protein was also in the nucleus and around the nucleus.
Therefore, a co-localization phenomenon of the distribution of
TRIM27 and PDCD4 proteins in endometrial and ovarian cancer cell
lines was identified (Fig. 1D). In
addition, the results from IHC showed that TRIM27 knockdown
increased the expression of PDCD4 in the tumors from a xenograft
nude mouse model of ovarian cancer (Fig. S2). The results indicated that the
protein expression levels of PDCD4 and TRIM27 have a negative
relationship in endometrial and ovarian cancer cell lines.
Silencing or overexpression of TRIM27
upregulates or downregulates the expression of PDCD4 protein
To confirm the negative association between the
expression of TRIM27 and PDCD4, TRIM27 was either overexpressed or
silenced in Ishikawa and A2780 cells and the mRNA and protein
expression of PDCD4 was detected. RT-qPCR results revealed that
PDCD4 mRNA expression had no obvious changes after TRIM27
overexpression in Ishikawa and A2780 cells or TRIM27 downregulation
in Ishikawa cells (Fig. 2A-C).
Compared with the control group, PDCD4 mRNA level was significantly
higher after TRIM27 downregulation in A2780 cells (Fig. 2D). Furthermore, it was found that
TRIM27 overexpression in Ishikawa and A2780 cells significantly
downregulated the PDCD4 protein expression (Fig. 2E and F). By contrast, the level of
PDCD4 protein was significantly upregulated after the
TRIM27-specific siRNA was transfected into Ishikawa and A2780 cells
(Fig. 2G and H). Thus, TRIM27 had
a significant negative impact on the expression of PDCD4
protein.
TRIM27 negatively regulates PDCD4 by
promoting the degradation of PDCD4
To explore whether TRIM27 negatively regulated PDCD4
by promoting the degradation of PDCD4, A2780 cells were transfected
with TRIM27-specific siRNA, or Ishikawa cells were transfected with
a TRIM27 expression plasmid. At 0, 2, 4 and 6 h before the protein
collection, the aforementioned transfected cells were added 2 µl of
CHX (10 mM) and incubated at 37°C for 0, 2, 4 or 6 h. It was found
that the expression of PDCD4 was upregulated after TRIM27 knockdown
in A2780 cells. The degradation rate of PDCD4 in TRIM27 knockdown
groups was slower than that of control groups after adding CHX. By
contrast, TRIM27 overexpression downregulated the levels of PDCD4
protein in Ishikawa cells, and the increasing degradation of PDCD4
protein was observed after adding CHX (Figs. 3A and B and S3). The results indicated that TRIM27
negatively regulates PDCD4 by promoting the degradation of
PDCD4.
TRIM27 promotes the degradation of
PDCD4 protein via the ubiquitin-proteasome pathway
Next, it was further confirmed that TRIM27 promoted
the degradation of PDCD4 protein via the ubiquitin-proteasome
pathway. A2780 cells were transfected with TRIM27-specific siRNA,
or Ishikawa cells were transfected with a TRIM27 expression
plasmid. At 0, 4 and 6 h before the protein collection, the
aforementioned transfected cells were added 3 µl of MG132 (10 mM)
and incubated at 37°C for 0, 4 and 6 h. The results revealed that
the expression of PDCD4 was increased after TRIM27 knockdown, and
the expression of PDCD4 was decreased after MG132 stimulation for 4
h compared with the control group. By contrast, TRIM27
overexpression decreased the protein expression of PDCD4. Compared
with the MG132-unstimulated group, the expression of PDCD4 was
increased after MG132 stimulation for 4 and 6 h. These results
demonstrated that the inhibitory effect of TRIM27 on PDCD4
expression disappeared after the degradation pathway of the
proteasome was blocked (Figs. 3C and
D and S4), suggesting that
TRIM27 promotes the degradation of PDCD4 via the
ubiquitin-proteasome pathway.
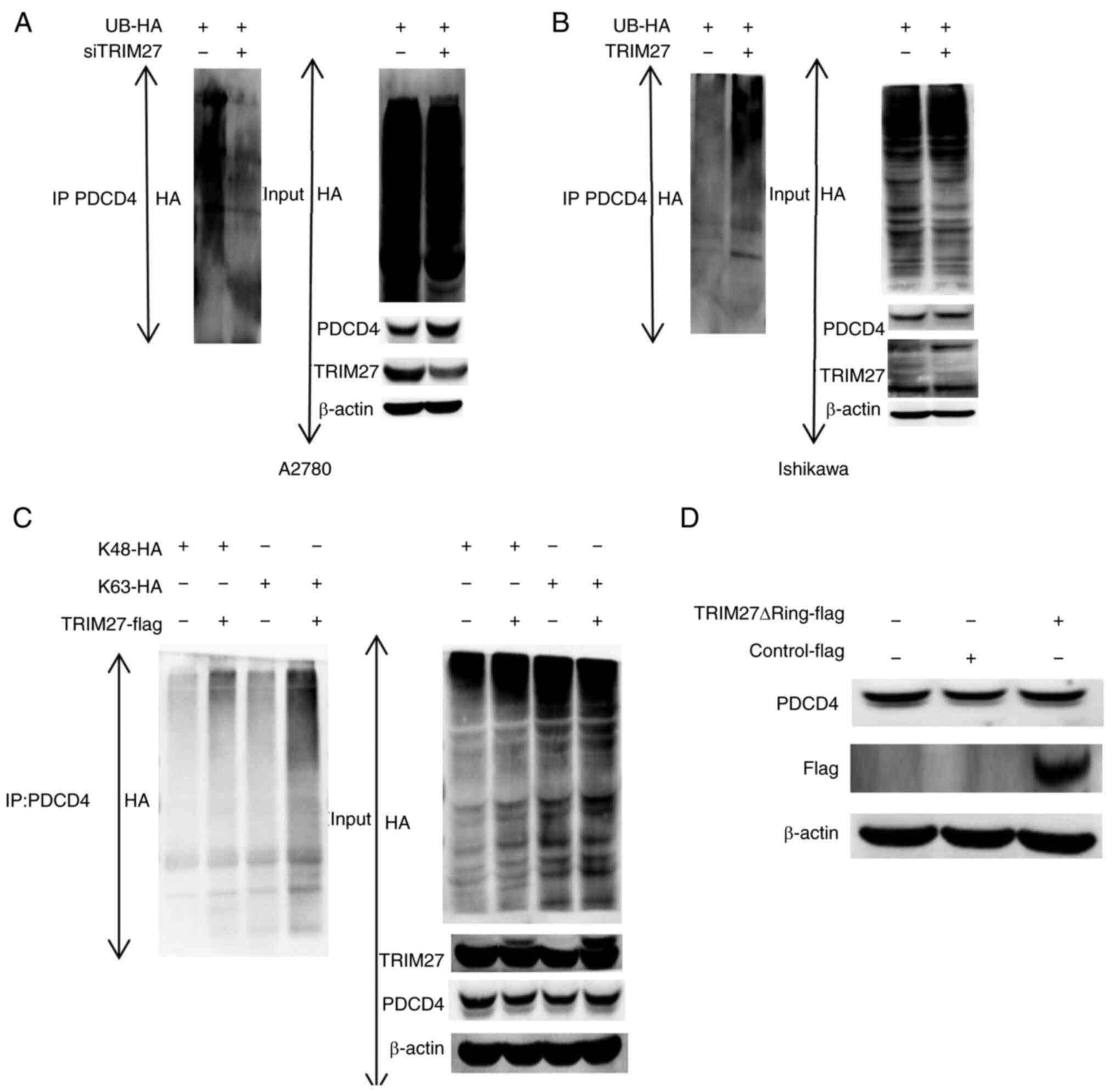
TRIM27 promotes the K48 and K63-linked
ubiquitination of PDCD4
To investigate whether TRIM27 promoted the
ubiquitination of PDCD4, A2780 cells were transfected with
TRIM27-specific siRNA. After 24 h, UB-HA expression plasmid was
transfected into the aforementioned cells. A total of 24 h later,
the results of co-IP showed that the levels of PDCD4 ubiquitination
were decreased. On the contrary, after TRIM27 and UB-HA expression
plasmids were co-transfected into Ishikawa cells, the levels of
PDCD4 ubiquitination increased (Fig.
4A and B). The results indicated TRIM27 promotes the
ubiquitination of PDCD4. Next, the sites of ubiquitination were
further detected. Ishikawa cells were respectively transfected with
TRIM27 expression plasmid, K48-HA and K63-HA expression plasmids,
and the proteins were collected after 24 h. TRIM27-mediated K48 and
K63-linked ubiquitination of PDCD4 was detected by co-IP. The
results showed that the total amount of K48 and K63-linked
ubiquitination of PDCD4 was upregulated after TRIM27
overexpression, which indicated that TRIM27 promoted the K48 and
K63-linked ubiquitination of PDCD4 (Fig. 4C).
TIRM27 regulates PDCD4 expression via
the Ring finger domain
The Ring finger domain of TRIM27 can mediate the
ubiquitin transfer between the proteins themselves or different
substrates. To confirm whether TRIM27 promoted the ubiquitination
of PDCD4 via the Ring finger domain, the Ring finger domain-deleted
expression plasmid (TRIM27∆Ring-flag) was constructed and
transfected into Ishikawa cells. The results showed that the
expression of PDCD4 had no change after TRIM27∆Ring-flag was
transfected into Ishikawa cells, which indicated that TRIM27
regulated PDCD4 expression via the Ring finger domain (Fig. 4D).
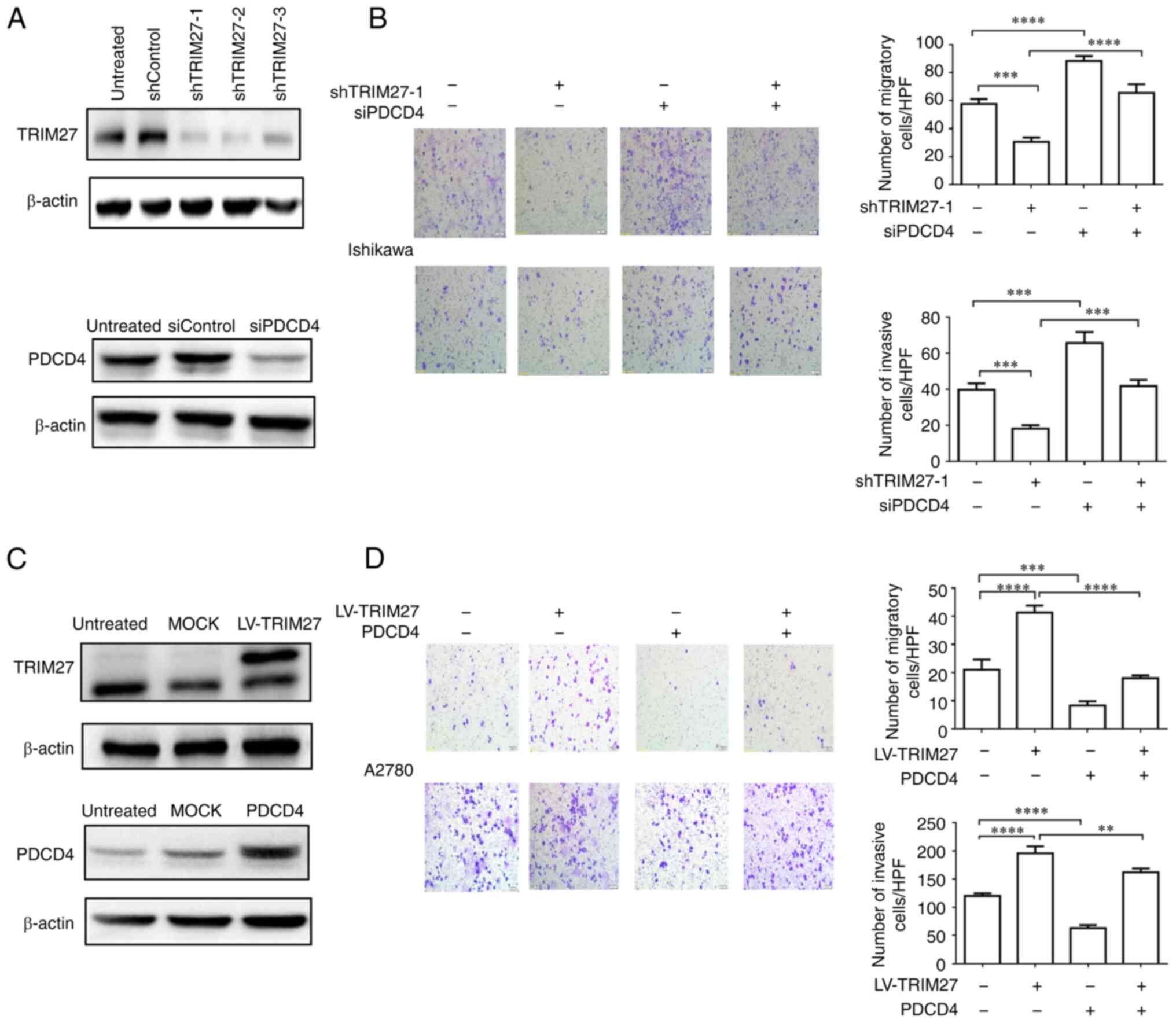
TRIM27 promotes the migration and
invasion of endometrial and ovarian cancer cell lines by affecting
PDCD4
To confirm whether TRIM27 could enhance the
migration and invasion of endometrial and ovarian cancer cells by
regulating PDCD4, a lentiviral expression vector and lentiviral
shRNAs specific for TRIM27 were constructed and
TRIM27-overexpressed A2780 cells or TRIM27-silenced Ishikawa cells
were set up. The results showed that TRIM27 overexpression
significantly increased the migration and invasion ability of A2780
cells, while TRIM27 knockdown significantly decreased the migration
and invasion ability of Ishikawa cells. By contrast, PDCD4
overexpression or knockdown significantly downregulated or
upregulated, respectively, the migration and invasion ability of
A2780 or Ishikawa cells. Furthermore, in TRIM27-overexpressing
A2780 cells, PDCD4 overexpression significantly decreased the
migration and invasion ability of cells. Similarly, in
TRIM27-silenced Ishikawa cells, PDCD4 knockdown obviously increased
the migration and invasion ability of cells (Figs. 5A-D; Figs. S5 and S6). The aforementioned results suggested
that TRIM27 could promote the migration and invasion of endometrial
and ovarian cancer cells by affecting PDCD4.
Discussion
Previous studies showed that TRIM27 was highly
expressed in ovarian serous carcinoma and endometrial cancer
(23,24). Higher TRIM27 expression was
significantly correlated with the metastasis and FIGO stage in
patients with ovarian serous carcinoma, and an unfavorable clinical
outcome in patients with endometrial cancer. PDCD4 was lowly
expressed or deleted in ovarian serous cystadenocarcinomas and
endometrioid endometrial carcinomas (EEC) (10,11).
The loss or reduction of PDCD4 expression was significantly
associated with higher pathological grade of ovarian serous
cystadenocarcinomas and EEC, as well as the prognosis of patients
with ovarian serous cystadenocarcinomas. The aforementioned results
suggested that TRIM27 and PDCD4 have a negative relationship. The
present study aimed to detect whether TRIM27 could take effect by
regulating the expression of PDCD4 in endometrial and ovarian
cancer cells. It was found that TRIM27 could degrade PDCD4 by the
ubiquitination effect and promote the migration and invasion of
endometrial and ovarian cancer cells by regulating PDCD4.
The expression of TRIM27 and PDCD4 in two kinds of
endometrial cancer cell lines (Ishikawa and HEC-1-A) and two types
of ovarian cancer cell lines (A2780 and SKOV3) was first detected.
The four kinds of cancer cells are all epithelial-derived cells and
share similar tumor biology. The results showed that TRIM27 and
PDCD4 had a negative association at protein levels while they had
no distinct association with each other at mRNA levels. The
differences in the expression of PDCD4 and TRIM27 between normal
and ovarian/endometrial cancer cells will be investigated in future
studies. The immunofluorescence double staining results revealed
that the protein distribution of TRIM27 and PDCD4 existed in
co-localization and were mainly distributed around the nucleus in
Ishikawa and A2780 cells. It has been reported that PDCD4 and
TRIM27 express in the cytoplasm and nuclei in a different
situation, and contain a nuclear export sequence and shuttle
between the cytoplasm and the nucleus (20,29).
These results suggested that TRIM27 and PDCD4 may combine to
interact.
Gene expression is regulated at multiple levels,
including gene, transcriptional, post-transcriptional,
translational and post-translational level. It has been reported
that PDCD4 expression could be regulated at different levels
(30). Gao et al (13) reported that the methylation of
PDCD4 5′CpG islands was significantly correlated with the loss of
PDCD4 mRNA expression in glioma tissue. DNA methyltransferase
inhibitor 5-aza-2 deoxycytidine could block the methylation in
glioma cells, restore PDCD4 expression and inhibit the
proliferation of cells (13). The
translation of PDCD4 mRNA is hampered by numerous miRNAs, including
miR21, miR182, miR16, miR150 and miR499. The miR21
post-transcriptionally downregulates PDCD4 expression by targeting
PDCD4 mRNA, inducing the increase in transformation, invasion and
metastasis of cancer cells (31).
PDCD4 is recognized and ubiquitinated by a ubiquitin E3 ligase,
β-TRCP, consequently degraded by the proteasome after it is
phosphorylated at Ser67 by p70S6K and Akt (32). The results of the present study
revealed that TRIM27 overexpression downregulated the expression of
PDCD4 protein, while TRIM27 knockdown had the opposite effect on
PDCD4 protein expression. However, enhanced or silenced TRIM27
expression had no obvious effect on the mRNA levels of PDCD4. These
results suggested that the regulation of PDCD4 expression by TRIM27
mainly exists after transcription. TRIM27 is a newly identified E3
ubiquitin ligase and degrades other proteins through the
ubiquitin-proteasome pathway (26,27).
In the present study, it was mainly investigated whether TRIM27
regulated the expression of PDCD4 protein via the
ubiquitin-proteasome pathway.
Ubiquitination is a kind of post-translational
modification. A small protein containing 76 amino acids is
covalently combined with lysine residues in substrate proteins
through a three-step E1, E2, and E3 enzymatic cascade.
Ubiquitination has different effects on its substrates depending on
the length and type of ubiquitin chains. Among them, K48-linked or
K63-linked ubiquitin chains are the most common, and their
functions are different. K48-linked ubiquitination mainly promotes
the target proteins to be degraded by the 26S proteasome. However,
K63-linked ubiquitination mostly plays an important role in
altering protein-protein interactions, protein conformations, or
targeting proteins for lysosomal delivery (33,34).
In the present study, it was found that the PDCD4
degradation rate was slower after stimulation with protein
synthesis inhibitor CHX. Furthermore, the expression of PDCD4 was
increased after proteasome inhibitor MG132 stimulation compared
with the MG132-unstimulated group. These results revealed that
PDCD4 could be degraded via the proteasome pathway. The present
results also demonstrated that ubiquitination levels of PDCD4 were
upregulated after TRIM27 overexpression, while TRIM27 knockdown
inhibited the ubiquitination level of PDCD4, indicating that TRIM27
can play a role in promoting the ubiquitination of PDCD4. It has
been reported that RFP (TRIM27) physically interacts with Pax7 and
serves as an E3 ligase to induce ubiquitin-proteasome-dependent
degradation of MyoD in muscle atrophy, which results in resistance
of muscles to denervation injury (35). Cai et al (26) reported that TRIM27 functioned as an
E3 ligase and mediated lysine 48 polyubiquitination of PI3KC2β,
leading to a decrease in PI3K enzyme activity.
Similarly, Zurek et al (36) reported that TRIM27 mediated
K48-linked ubiquitination and subsequent proteasomal degradation of
NOD2 negatively influencing NOD2-mediated NF-kB activation.
However, Hao et al (37)
found that the E3 Ring ubiquitin ligase, MAGE-L2-TRIM27,
facilitated K63-linked ubiquitination of WASH K220. The disruption
of WASH ubiquitination impaired endosomal F-actin nucleation and
retromer-dependent transport (37). The aforementioned results confirmed
that TRIM27 could mediate K48 or K63-linked ubiquitination of
different target proteins. Our present study found that the total
amount of K48 and K63-linked ubiquitination of PDCD4 was increased
after TRIM27 overexpression. Given different K48 and K63-linked
ubiquitination functions, it was hypothesized that TRIM27 promotes
the proteasome-dependent degradation of PDCD4 by mediating
K48-linked ubiquitination of PDCD4. The role of K63-linked
ubiquitination of PDCD4 needs to be further explored in the future.
As aforementioned, the Ring finger is a zinc-binding domain that
plays an important role in ubiquitin E3 ligase binding to
UB-conjugating enzymes (E2) (38).
Therefore, the Ishikawa cells were transfected with TRIM27 mutant
plasmid without the Ring finger domain. The result showed that
PDCD4 was not degraded, confirming that TRIM27 could degrade PDCD4
by ubiquitination via the Ring finger domain.
PDCD4 is an important tumor suppressor, which
inhibits the proliferation, migration and invasion of various
cancer cells. The present results demonstrated that TRIM27 promotes
the ubiquitination and degradation of PDCD4. However, whether
TRIM27 could enhance the migration and invasion of endometrial and
ovarian cancer cells by regulating PDCD4 remains unclear. In the
present study, it was found that PDCD4 overexpression decreased the
migration and invasion ability of cancer cells in
TRIM27-overexpressing cells, while PDCD4 knockdown increased the
migration and invasion ability of cells in TRIM27-silenced cells.
The aforementioned results suggested that TRIM27 could promote the
migration and invasion of endometrial and ovarian cancer cell lines
by affecting PDCD4.
In conclusion, the present study was, to the best of
our knowledge, the first to demonstrate the negative association of
TRIM27 with PDCD4. TRIM27 could degrade the PDCD4 protein level
through the ubiquitin-proteasome pathway and promote the migration
and invasion of endometrial and ovarian cancer cell lines by
regulating PDCD4. The present results will provide important
insights into the effective mechanism of TRIM27 and the reason for
PDCD4 downregulation in cancers.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 81771554 and 81471437) and the
Natural Science Foundation of Shandong (grant no. ZR2018MH013).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZW and XiaW designed the project and conducted the
experimental study. HY performed the experiments and wrote the
manuscript. LW, ZT, CY, DZ, MJ and CW participated in performing
certain experiments. YL, CX and XisW were involved in data
collection and statistical analysis. YS and LZ participated in
designing the experiments and reviewing the manuscript. All authors
read and approved the final manuscript. ZW and HY confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Visser NCM, Reijnen C, Massuger LFAG,
Nagtegaal ID, Bulten J and Pijnenborg JMA: Accuracy of endometrial
sampling in endometrial carcinoma: A Systematic review and
meta-analysis. Obstet Gynecol. 130:803–813. 2017. View Article : Google Scholar
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, ParkinD M, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hinchcliff EM, Bednar EM, Lu KH and
Rauh-Hain JA: Disparities in gynecologic cancer genetics
evaluation. Gynecol Oncol. 153:184–191. 2019. View Article : Google Scholar
|
|
4
|
Schlichter U, Burk O, Worpenberg S and
Klempnauer KH: The chicken Pdcd4 gene is regulated by v-Myb.
Oncogene. 20:231–239. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jansen AP, Camalier CE, Stark C and
Colburn NH: Characterization of programmed cell death 4 in multiple
human cancers reveals a novel enhancer of drug sensitivity. Mol
Cancer Ther. 3:103–110. 2004.PubMed/NCBI
|
|
6
|
Wang Q and Yang HS: The role of Pdcd4 in
tumour suppression and protein translation. Biol Cell. May
28–2018.(Epub ahead of print). View Article : Google Scholar
|
|
7
|
Nieves-Alicea R, Colburn NH, Simeone AM
and Tari AM: Programmed cell death 4 inhibits breast cancer cell
invasion by increasing tissue inhibitor of metalloproteinases-2
expression. Breast Cancer Res Treat. 114:203–209. 2009. View Article : Google Scholar
|
|
8
|
Wei N, Liu SS, Chan KK and Ngan HY: Tumour
suppressive function and modulation of programmed cell death 4
(PDCD4) in ovarian cancer. PLoS One. 7:e303112012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wei ZT, Zhang X, Wang XY, Gao F, Zhou CJ,
Zhu FL, Wang Q, Gao Q, Ma CH, Sun WS, et al: PDCD4 inhibits the
malignant phenotype of ovarian cancer cells. Cancer Sci.
100:1408–1413. 2009. View Article : Google Scholar
|
|
10
|
Wang X, Wei Z, Gao F, Zhang X, Zhou C, Zhu
F, Wang Q, Gao Q, Ma C, Sun W, et al: Expression and prognostic
significance of PDCD4 in human epithelial ovarian carcinoma. Anti
Res. 28:2991–2996. 2008.
|
|
11
|
Liu Y, Sun H, Mao H, Gao M, Tan X, Li Y,
Li Y, Muloye GM, Zhang L, Wang X and Wei Z: Expression of tumor
suppressor programmed cell death 4 in endometrioid endometrial
carcinomas and clinicopathological significance. Oncol Lett.
15:9369–9376. 2018.
|
|
12
|
Zhang X, Wang X, Song X, Liu C, Shi Y,
Wang Y, Afonja O, Ma C, Chen YH and Zhang L: Programmed cell death
4 enhances chemosensitivity of ovarian cancer cells by activating
death receptor pathway in vitro and in vivo. Cancer Sci.
101:2163–2170. 2010. View Article : Google Scholar
|
|
13
|
Gao F, Wang X, Zhu F, Wang Q, Zhang X, Guo
C, Zhou C, Ma C, Sun W, Zhang Y, et al: PDCD4 gene silencing in
gliomas is associated with 5′CpG island methylation and
unfavourable prognosis. J Cell Mol Med. 13:4257–4267. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun J, Yong J and Zhang H: microRNA-93,
upregulated in serum of nasopharyngeal carcinoma patients, promotes
tumor cell proliferation by targeting PDCD4. Exp Ther Med.
19:2579–2587. 2020.PubMed/NCBI
|
|
15
|
Zhou XH, Chai HX, Bai M and Zhang Z:
LncRNA-GAS5 regulates PDCD4 expression and mediates myocardial
infarction-induced cardiomyocytes apoptosis via targeting MiR-21.
Cell Cycle. 19:1363–1377. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cui H, Wang Q, Lei Z, Feng M, Zhao Z, Wang
Y and Wei G: DTL promotes cancer progression by PDCD4
ubiquitin-dependent degradation. J Exp Clin Cancer Res. 38:3502019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Townson SM, Kang K, Lee AV and Oesterreich
S: Novel role of the RET finger protein in estrogen
receptor-mediated transcription in MCF-7 cells. Biochem Biophys Res
Commun. 349:540–548. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Micale L, Chaignat E, Fusco C, Reymond A
and Merla G: The tripartite motif: Structure and function. Adv Exp
Med Biol. 770:11–25. 2012. View Article : Google Scholar
|
|
19
|
Joazeiro CA and Weissman AM: RING finger
proteins: Mediators of ubiquitin ligase activity. Cell.
102:549–552. 2000. View Article : Google Scholar
|
|
20
|
Harbers M, Nomura T, Ohno S and Ishii S:
Intracellular localization of the Ret finger protein depends on a
functional nuclear export signal and protein kinase C activation. J
Biol Chem. 276:48596–48607. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iwakoshi A, Murakumo Y, Kato T, Kitamura
A, Mii S, Saito S, Yatabe Y and Takahashi M: RET finger protein
expression is associated with prognosis in lung cancer with
epidermal growth factor receptor mutations. Pathol Int. 62:324–330.
2012. View Article : Google Scholar
|
|
22
|
Tezel GG, Uner A, Yildiz I, Guler G and
Takahashi M: RET finger protein expression in invasive breast
carcinoma: Relationship between RFP and ErbB2 expression. Pathol
Res Pract. 205:403–408. 2009. View Article : Google Scholar
|
|
23
|
Ma Y, Wei Z, Bast RC Jr, Wang Z, Li Y, Gao
M, Liu Y and Wang X, Guo C, Zhang L and Wang X: Downregulation of
TRIM27 expression inhibits the proliferation of ovarian cancer
cells in vitro and in vivo. Lab Invest. 96:37–48. 2016. View Article : Google Scholar
|
|
24
|
Tsukamoto H, Kato T, Enomoto A, Nakamura
N, Shimono Y, Jijiwa M, Asai N, Murakumo Y, Shibata K, Kikkawa F
and Takahashi M: Expression of Ret finger protein correlates with
outcomes in endometrial cancer. Cancer Sci. 100:1895–1901. 2009.
View Article : Google Scholar
|
|
25
|
Horio M, Kato T, Mii S, Enomoto A, Asai M,
Asai N, Murakumo Y, Shibata K, Kikkawa F and Takahashi M:
Expression of RET finger protein predicts chemoresistance in
epithelial ovarian cancer. Cancer Med. 1:218–229. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai X, Srivastava S, Sun Y, Li Z, Wu H,
Zuvela-Jelaska L, Li J, Salamon RS, Backer JM and Skolnik EY:
Tripartite motif containing protein 27 negatively regulates CD4 T
cells by ubiquitinating and inhibiting the class II PI3K-C2β. Proc
Natl Acad Sci USA. 108:20072–20077. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng Q, Hou J, Zhou Y, Yang Y, Xie B and
Cao X: Siglec1 suppresses antiviral innate immune response by
inducing TBK1 degradation via the ubiquitin ligase TRIM27. Cell
Res. 25:1121–1136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Böhm M Sawicka K, Siebrasse JP,
Brehmer-Fastnacht A, Peters R and Klempnauer KH: The transformation
suppressor protein Pdcd4 shuttles between nucleus and cytoplasm and
binds RNA. Oncogene. 22:4905–4910. 2003. View Article : Google Scholar
|
|
30
|
Matsuhashi S, Manirujjaman M, Hamajima H
and Ozaki I: Control mechanisms of the tumor suppressor PDCD4:
Expression and functions. Int J Mol Sci. 20:23042019. View Article : Google Scholar
|
|
31
|
Frankel LB, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dorrello NV, Peschiaroli A, Guardavaccaro
D, Colburn NH, Sherman NE and Pagano M: S6K1- and betaTRCP-mediated
degradation of PDCD4 promotes protein translation and cell growth.
Science. 314:467–4671. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mallette FA and Richard S: K48-linked
ubiquitination and protein degradation regulate 53BP1 recruitment
at DNA damage sites. Cell Res. 22:1221–1223. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Metcalf JL, Bradshaw PS, Komosa M, Greer
SN, Stephen Meyn M and Ohh M: K63-Ubiquitylation of VHL by SOCS1
mediates DNA double-strand break repair. Oncogene. 33:1055–1065.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Joung H, Eom GH, Choe N, Lee HM, Ko JH,
Kwon DH, Nam YS, Min H, Shin S, Kook J, et al: Ret finger protein
mediates Pax7-induced ubiquitination of MyoD in skeletal muscle
atrophy. Cell Signal. 26:2240–2248. 2014. View Article : Google Scholar
|
|
36
|
Zurek B, Schoultz I, Neerincx A,
Napolitano LM, Birkner K, Bennek E, Sellge G, Lerm M, Meroni G,
Söderholm JD and Kufer TA: TRIM27 negatively regulates NOD2 by
ubiquitination and proteasomal degradation. PLoS One. 7:e412552012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hao YH, Doyle JM, Ramanathan S, Gomez TS,
Jia D, Xu M, Chen ZJ, Billadeau DD, Rosen MK and Potts PR:
Regulation of WASH-dependent actin polymerization and protein
trafficking by ubiquitination. Cell. 152:1051–1064. 2013.
View Article : Google Scholar
|
|
38
|
Lipkowitz S and Weissman AM: RINGs of good
and evil: RING finger ubiquitin ligases at the crossroads of tumour
suppression and oncogenesis. Nat Rev Cancer. 11:629–643. 2011.
View Article : Google Scholar : PubMed/NCBI
|