Introduction
The treatment options available to assist infertile
couples in having children have progressed immensely during recent
years to include a variety of assisted reproductive technologies
(1). However, for in vitro
fertilization-embryo transfer (IVF-ET), embryo implantation remains
the rate-limiting step (2–4). Couples who fail to achieve a pregnancy
following between two and six IVF cycles, in which >10
high-grade embryos were transferred to the uterus are defined by
various clinicians as having repeated implantation failure (RIF)
(5,6). With the tendency being that only one or
two high-grade embryos are transferred in each cycle, certain
clinicians have recommended that the definition of RIF be changed
to the failure of implantation in at least three consecutive IVF
attempts (6–8). In the present study, patients with
failure of ≥3 consecutive IVF-ET cycles were defined as RIF.
Appropriate immune responses at the time of embryo
introduction are key for successful implantation into the
endometrial wall. Previous studies have reported that the
prevalence of dominant T-helper (Th)1 and Th17 cells may cause
multiple implantation failures in IVF cycles, while the prevalence
of dominant Th2 and regulatory T cells (Tregs) is beneficial for a
successful in vitro treatment outcome (3,8–12). Liang et al (13) identified that the Th1/Th2 ratio in
circulating Th cells was significantly increased in women with RIF
compared to those with a successful implantation. While Persson
et al (9) demonstrated that
unsuccessful IVF outcomes had an abnormality in the quantities of
peripheral Th1 and Th17 cells, and that following embryo transfer
pregnant women had higher numbers of Th2-associated
cytokine-secreting cells. Another study revealed that pregnancy and
live birth rates were significantly improved in women with >0.6%
circling T cells as Tregs (11).
However, these previous studies fail to clarify the enhancement of
humoral immune responses in women with reproductive problems. T
follicular helper (Tfh) cells are involved in humoral immunity
(14,15) and the role they serve in RIF remain
unclear.
Tfh cells are known as one subset of CD4+
T cells settled in the secondary lymph nodes, they have increased
expression of chemokine receptor type 5 (CXCR5) and decreased
expression of chemokine receptor 7 (CCR7), which guides Tfh cell
migration towards B cell follicles (16,17).
Currently, Tfh cells are defined by their expression of
combinations of markers, including programmed death-1 (PD-1),
interleukin (IL)-21 and IL-4, which are directly linked with the
biology of Tfh cells. High expression of PD-1 in combination with
CXCR5 is a reliable way to identify Tfh cells (18). IL-21 and IL-4 are the main cytokines
secreted by Tfh cells, and their cooperation can regulate
immunoglobulin G (IgG)-1 production and the production of multiple
antibody classes (19,20). Transcription factor B-cell lymphoma 6
(Bcl-6) is the master regulator of Tfh cell differentiation
(21–23). IL-6 is helpful in Tfh cell
differentiation while IL-2 negatively regulates Tfh by inducing B
lymphocyte-induced maturation protein 1 expression (24). These molecules are all considered
therapeutic targets to prevent B cells from producing high affinity
autoantibodies directed against self-antigens.
Circulating CD4+ T cells that express
CXCR5 are considered counterparts to Tfh cells (25). They are thought to be primarily
generated in germinal centers (GCs) (26). Very few circulating CD4+ T
cells express high amounts of CXCR5 or PD-1, but a significant
fraction express intermediate amounts of these molecules and are
designated as circulating Tfh (cTfh) cells (18). Human cTfh cells are composed of
distinct subsets with unique phenotypes and function. The
CCR7(lo)PD-1(hi) subset has a partial Tfh effector phenotype that
is indicative of active Tfh differentiation in lymphoid organs and
correlated with clinical indices in autoimmune diseases. This
provides a biomarker to monitor protective antibody responses
during infection or vaccination and pathogenic antibody responses
in autoimmune diseases (26–28). In certain diseases, including
systemic lupus erythematosus (29–31),
rheumatoid arthritis (25,32) and allograft rejection (33), the percentages of cTfh cells are
increased and positively correlated with serum levels of
autoantibodies; however, the underlying mechanisms of cTfh and
associated factors in the development of RIF require further
study.
In the present study, CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells and associated factors
were investigated, and it was identified that their presence was
significantly increased in patients with RIF, suggesting that they
are involved in the development of RIF. These results provide a
novel insight into the development of potential therapies to help
prevent RIF.
Materials and methods
Patients and controls
A total of 60 females were enrolled in the present
study. The patients were admitted to the First Affiliated Hospital
of Xinjiang Medical University (Urumqi, China) between July 2013
and December 2015. They formed two separate groups: The RIF group
and the control group (n=30/group). The inclusion criteria for the
RIF group were females ≤43 years old with a history of ≥3 previous
failed IVF-ET cycles, with a normal uterus, high quality embryos,
menstrual regularity and normal sperm from the partner. The
exclusion criteria were severe endometriosis, adhesiolysis, an
abnormal uterus, having received immune therapy and the presence of
possible causes for the failure of implantation, including
autoimmune diseases, endocrine disorders, couples with chromosomal
abnormalities and sexually transmitted diseases. A detailed history
of the patients was taken, as well as a physical examination and
investigations for infertility. The inclusion criteria for the
control group were healthy females ≤43 years with infertile
partners who got conceived and gave birth during their first cycle
of IVF-ET, the exclusion criteria were the same as those for the
RIF group. Baseline characteristics between the study groups were
not significantly different, as shown in Table I.
 | Table I.Clinicopathological characteristics
of the RIF and control groups. |
Table I.
Clinicopathological characteristics
of the RIF and control groups.
|
| Control (n=30) | RIF (n=30) |
|
|---|
|
|
|
|
|
|---|
| Characteristic | Mean | SD | Mean | SD | P-value |
|---|
| Age (years) | 32.77 | 4.7 | 33.23 | 5.06 | 0.712 |
| BMI
(kg/m2) | 21.69 | 2.5 | 22.96 | 2.74 | 0.066 |
| Length of
infertility (years) | 4.78 | 3.76 | 4.71 | 3.5 | 0.936 |
| Endometrial
thickness (mm) | 10.53 | 1.78 | 10.58 | 1.98 | 0.932 |
| No. of previous
failed IVF-ET cycles | – | – | 3.4 | 0.56 | – |
Ethical approval for the present study was obtained
from the Human Ethics Committee of the First Affiliated Hospital of
Xinjiang Medical University prior to commencement of the study.
Patients who fulfilled the inclusion criteria were recruited
following the attainment of written informed consent.
Specimen collection
Endometrial tissues were collected from the RIF and
control group 1 month prior to the first IVF-ET cycle (34). IVF-ET was performed in the mid luteal
phase of the menstrual cycle when the uterus is most receptive to
the embryo; the specific time period for this is known as the
implantation window. One section of the tissue was fixed with 4%
paraformaldehyde solution for 3 days at room temperature. Sections
were embedded in paraffin and sliced into 4-µm-thick sections to be
utilized for immunohistochemistry, while another section was used
for reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis. Peripheral blood was obtained from the RIF and
control groups in the mid luteal phase of the menstrual cycle; this
was then used to detect the proportion of Tfh cells and the
concentration of cytokines.
Flow cytometric analysis
Peripheral blood mononuclear cells (PBMCs) were
isolated from fresh heparinized peripheral blood samples (35), then suspended in PBS at
1×106 cells/ml. Next, 100 µl of PBMCs in suspension and
anti-human monoclonal antibodies conjugated with different
fluorescent dyes, including 2 µl peridinin chlorophyll-CD4 (1:60;
cat no. 347324), 2 µl phycoerythrin (PE)-cyanin 7-CCR7 (1:60; cat
no. 557648), 2.5 µl fluorescein isothiocyanate-CXCR5 (1:48; cat no.
558112), 2 µl allophycocyanin-CD45RA (1:60; cat no. 561210) and 2
µl PE-PD-1 (1:60; cat no. 12996941) were combined. All antibodies
were purchased from BD Biosciences (San Jose, CA, USA), except
anti-PD-1, which was from eBioscience (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The samples were incubated for 15 min at
4°C in dark conditions. The cells were washed with PBS and then
centrifuged at 300 × g for 5 min at 4°C. After removing the
supernatant, the last step was repeated. Then the cells were
resuspended in 300 µl PBS. After gating for sideward scatter and
CD4 cells positive for CD4 were further analyzed. CD45RA was used
to identify the memory T cells (CD45RA) in the CD4+ T
cell population. The CD45RA− CXCR5+ cells
were then gated and the CCR7(lo)PD-1(hi) subset of
CD45RA− CXCR5+ cells was analyzed. For each
sample >20,000 cells were collected, which were detected using a
BD LSR II flow cytometer (BD Biosciences), and analyzed using
FlowJo software (version 7.6.1; Tree Star, Inc., Ashland, OR,
USA).
RNA isolation and RT-qPCR
analysis
Total RNA was isolated from endometrial tissue of
each group with Lipofectamine (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol and
reverse transcribed into complementary (c)DNA using the Roche
Transcriptor cDNA Synthesis kit (Roche Diagnostics, Basel,
Switzerland) according to the manufacturers protocols. The cDNA was
then used as the template for the qPCR reaction. SYBR-Green (Takara
Biotechnology Co., Ltd., Dalian, China) incorporation-based qPCR
was performed to quantify the mRNA levels of Bcl-6, IL-21 and CXCR5
with a CFX96™ Real-Time PCR Detection system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Reactions were carried out through 34
cycles of 30 sec at 94°C and 1 min at 55°C, followed by 30 sec at
72°C. The primers were designed by Sangon Biotech Co., Ltd.
(Shanghai, China) and presented in Table II. The relative expression was
calculated using the 2−∆∆Cq method (36). The results were normalized to the
housekeeping gene GAPDH and represented as lg2−∆∆Cq.
Each sample was conducted with two biological replicates.
| Table II.Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table II.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Direction | Primer sequence
(5′-3′) | Expected product
size (bp) |
|---|
| Bcl-6 | F |
ATGAGGAGTTTCGGGATGTC | 177 |
|
| R |
CCTCTTCTGGGATTGTTTGC |
|
| CXCR5 | F |
GCAAGAAAGAAACCCGACAG | 210 |
|
| R |
TTATGGGAAGGGAGTGAGGA |
|
| IL-21 | F |
CACAGACTAACATGCCCTTCAT | 224 |
|
| R |
GAATCTTCACTTCCGTGTGTTCT |
|
| GAPDH | F |
GAAGGTGAAGGTCGGAGT | 226 |
|
| R |
GAAGATGGTGATGGGATTTC |
|
Pathological morphology and
immunohistochemistry
Hematoxylin and eosin (H&E) staining was used to
observe the endometrial tissue. Slides were deparaffinized with
xylene and rehydrated in gradual dilutions of ethanol. The slides
were then stained with hematoxylin for 3 min at room temperature.
After rinsing under running water for 3 min, slides were
differentiated with 1% HCl in ethanol for 1 sec at room
temperature. Finally, sections were counterstained with eosin for
90 sec at room temperature. Slides were examined and images were
captured using a light microscope at ×200 and ×400
magnification.
Immunohistochemical staining for Bcl-6, IL-21 and
CXCR5 was performed on endometrial slides. Firstly, slides were
deparaffinized with xylene; rehydrated in a graded series of
ethanol, immersed in 3% H2O2 and then placed
in buffered citric acid (pH 6.0) in a microwave oven (800 W) for 13
min. After being washed three times with PBS, the slides were
incubated with 10% goat serum (Beijing Zhongshan Golden Bridge
Biotechnology, Co., Ltd., Beijing, China) for 30 min at 37°C. Then,
the slides were washed three times with PBS and incubated with
rabbit anti-human polyclonal primary antibodies directed against
Bcl-6 (1:100; cat no. bs2734R), IL-21 (1:250 cat no. bs2621R) and
CXCR5 (1:100; cat no. bs3598R) (BIOSS, Beijing, China) overnight at
4°C. After three washes, slides were incubated with the
corresponding peroxidase-conjugated Affinipure goat anti-rabbit IgG
secondary antibodies (1:100; cat no. SPN9001; Beijing Zhongshan
Golden Bridge Biotechnology, Co., Ltd.) for 25 min at 37°C. The
antibody reaction was visualized using the chromogen
diaminobenzidine (Beijing Zhongshan Golden Bridge Biotechnology,
Co., Ltd.). Finally, all the slides were counterstained with
hematoxylin for 2 min at room temperature. Immunoreactivity was
assessed in five randomly selected high power fields (Leica DM3000;
Leica Microsystems Inc., Wetzlar, Germany) and the average optical
density was measured using Image-Pro Plus software (version 6.0;
Media Cybernetics, Inc., Rockville, MD, USA) on the histogram-based
mode.
ELISA
Serum levels of IL-21 from patients with RIF and
healthy females were determined using a human IL-21 ELISA kit (cat
no. bsK00366; BIOSS) according to the manufacturer's protocol.
Firstly, 100 µl/well of diluted serum (1:2) was added to a
microwell plate coated with monoclonal antibodies directed against
human IL-21, this was performed at 37°C for 30 min. The plate was
then washed three times with PBS with 1% Tween (PBST), and then
streptavidin-HRP was added to the plate at 37°C for 40 min. After
washing with PBST four times, 50 µl/well of TMB substrate was added
to react with the enzyme in the dark. Then concentrated
H2SO4 was used to terminate the reaction.
Finally, the optical density of the plates at 450 nm was detected
using a VersaMax™ microplate reader (Molecular Devices, LLC,
Sunnyvale, CA, USA).
Cytometric bead array (CBA)
Serum levels of IL-6, IL-4 and IL-2 were detected
using a CBA kit (cat no. 560484; BD Biosciences) according to the
manufacturer's protocol. In summary, 50 µl of mixed capture beads,
serum sample and PE detection reagents were added to the tubes,
which were then incubated for 3 h in the dark at room temperature.
After washing and centrifuging at 300 × g for 5 min at 4°C, the
supernatant was discarded and 300 µl of wash buffer was added to
each tube. Then, the levels of IL-6, IL-4 and IL-2 were detected
using a BD LSR II flow cytometer, and the data was analyzed using
FlowJo software (version 7.6.1; FlowJo LLC, Ashland, OR, USA).
Statistical analysis
All group data analysis was performed using SPSS
statistical software (version 17.0; SPSS, Inc., Chicago, IL, USA).
Results are expressed as the mean ± standard deviation. An
independent-samples t-test was used to assess differences between
two groups. Pearson's correlation coefficient was used to analyze
the correlation between two variables. P<0.05 (two-tailed) was
considered to indicate a statistically significant difference.
Results
An increased proportion of
CCR7(lo)PD-1(hi) T cells are found in the peripheral blood of women
with RIF
The activated Tfh cell subset was defined as the
CCR7(lo)PD-1(hi) subset. It was hypothesized that the quantities of
CCR7(lo)PD-1(hi) Tfh cells may be different in patients with RIF
compared to the control group. Flow cytometry analysis was
performed to detect the percentage of this subset within the total
CXCR5+ CD4+ T cell population in the
peripheral blood. This could then reflect the systemic contribution
of Tfh cells to implantation failure in IVF. CD4+ T
cells were discriminated from total cells (Fig. 1A) and memory Tfh cells
(CD45RA− CXCR5+ CD4+ T) were
identified in the CD4+ T cell population (Fig. 1B); the CXCR5+
CD45RA− CD4+ cells expressing
CCR7(lo)PD-1(hi) were analyzed (Fig.
1C). The percentage of CXCR5+ CD45RA−
CD4+ T cells in the total number of CD4+ T
cells was not significantly different between the two groups
(Fig. 1D). Whereas, the percentage
of subset CCR7(lo)PD-1(hi) within the CXCR5+
CD4+ T cells in the RIF (16.34±4.69) group was
significantly increased (14.01±4.22) compared with the control
group (P<0.05; Fig. 1E).
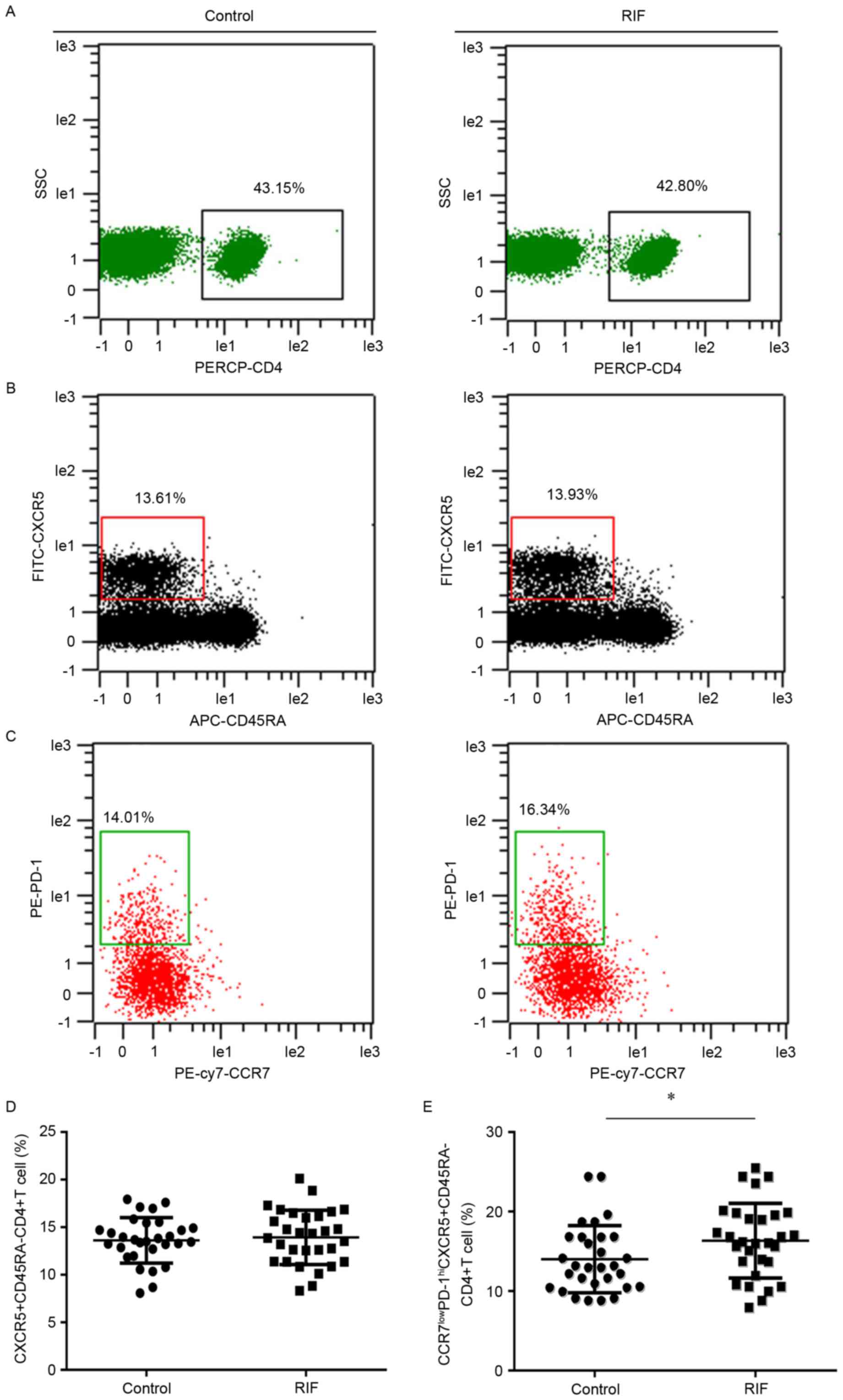 | Figure 1.Expression of CCR7(lo)PD-1(hi) T
follicle helper cells in the peripheral blood. Cytometric plots
indicating how (A) PERCP-CD4 was used to identify CD4+ T
cells. These cells were further analyzed to identify cells
expressing (B) CXCR5 and CD45RA. (C) CXCR5+
CD45RA− cells were analyzed to reveal the presence of
CCR7 and PD-1. The presence of (D) CXCR5+
CD45RA− CD4+ T cells and (E) CCR7(lo)PD-1(hi)
CXCR5+ CD45RA− CD4+ T cells in the
RIF group and control group were compared. *P<0.05 vs. the
control group. CCR7, chemokine receptor 7; PD-1, programmed death
1; Tfh, T follicle helper; CXCR5, chemokine receptor type 5; SSC,
side scatter; RIF, repeated implantation failure; CD, cluster of
differentiation; CD45RA, CD45 RA isotope; PE, phycoerythrin; FITC,
fluorescein isothiocyanate; APC, allophycocyanin; cy7, cyanin
7. |
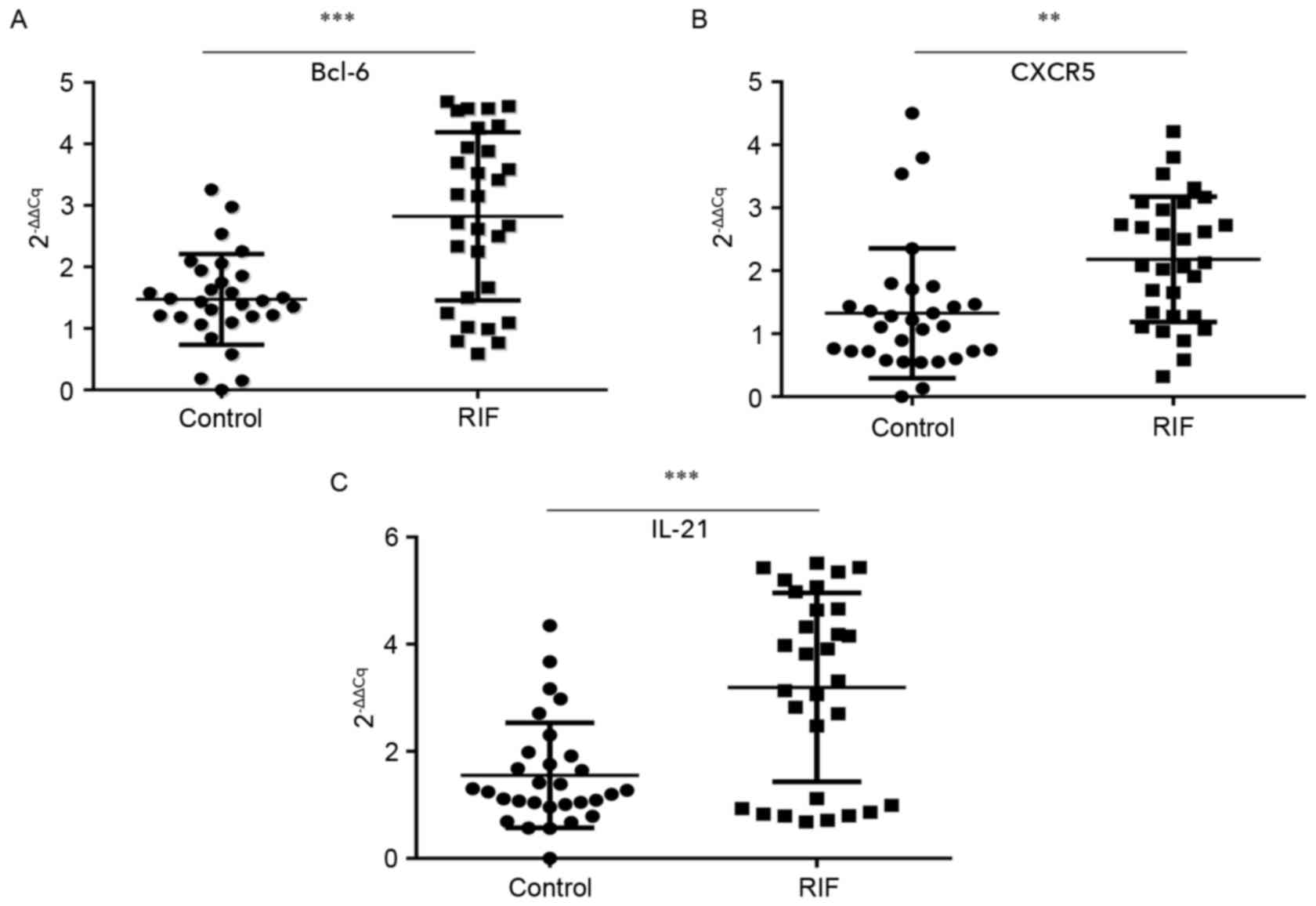
Bcl-6, IL-21 and CXCR5 mRNA expression
is increased within the endometrium of the RIF group in the mid
luteal phase
A successful pregnancy is dependent upon
synchronized, coordinated cross talk between the local and systemic
immune systems. The endometrium is the beginning of the maternal
interface and embryo attachment requires active local endometrial
reactivity on the maternal side. Therefore, the endometrial local
immune microenvironment is important to investigate. It was
proposed that the endometrial immune microenvironment may change as
the proportion of circulating Tfh cells increased in patients with
RIF. The mRNA expression of Bcl-6, IL-21 and CXCR5 in endometrial
tissues during the luteal phase was tested by RT-qPCR. The mean
expression of Bcl-6 (2.82±1.37; Fig.
2A), CXCR5 (2.18±1.00; Fig. 2B)
and IL-21 (3.20±1.76; Fig. 2C) were
all significantly higher in the RIF group than in the control group
(1.47±0.74, P<0.001; 1.33±1.03, P<0.01; and 1.56±0.98,
P<0.001 respectively). These results suggest that the
overexpression of the Tfh-associated markers Blc-6, CXCR5 and IL-21
in the endometrium during the mid luteal phase is associated with
embryo implantation failure.
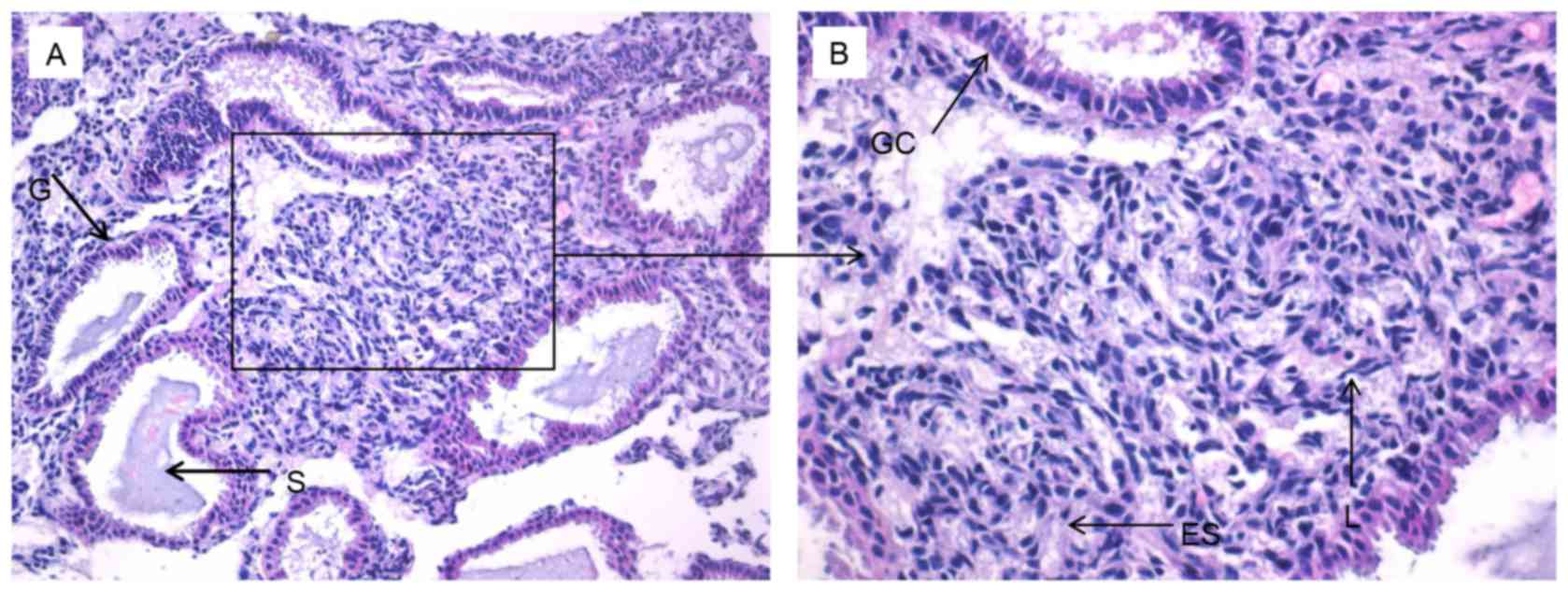
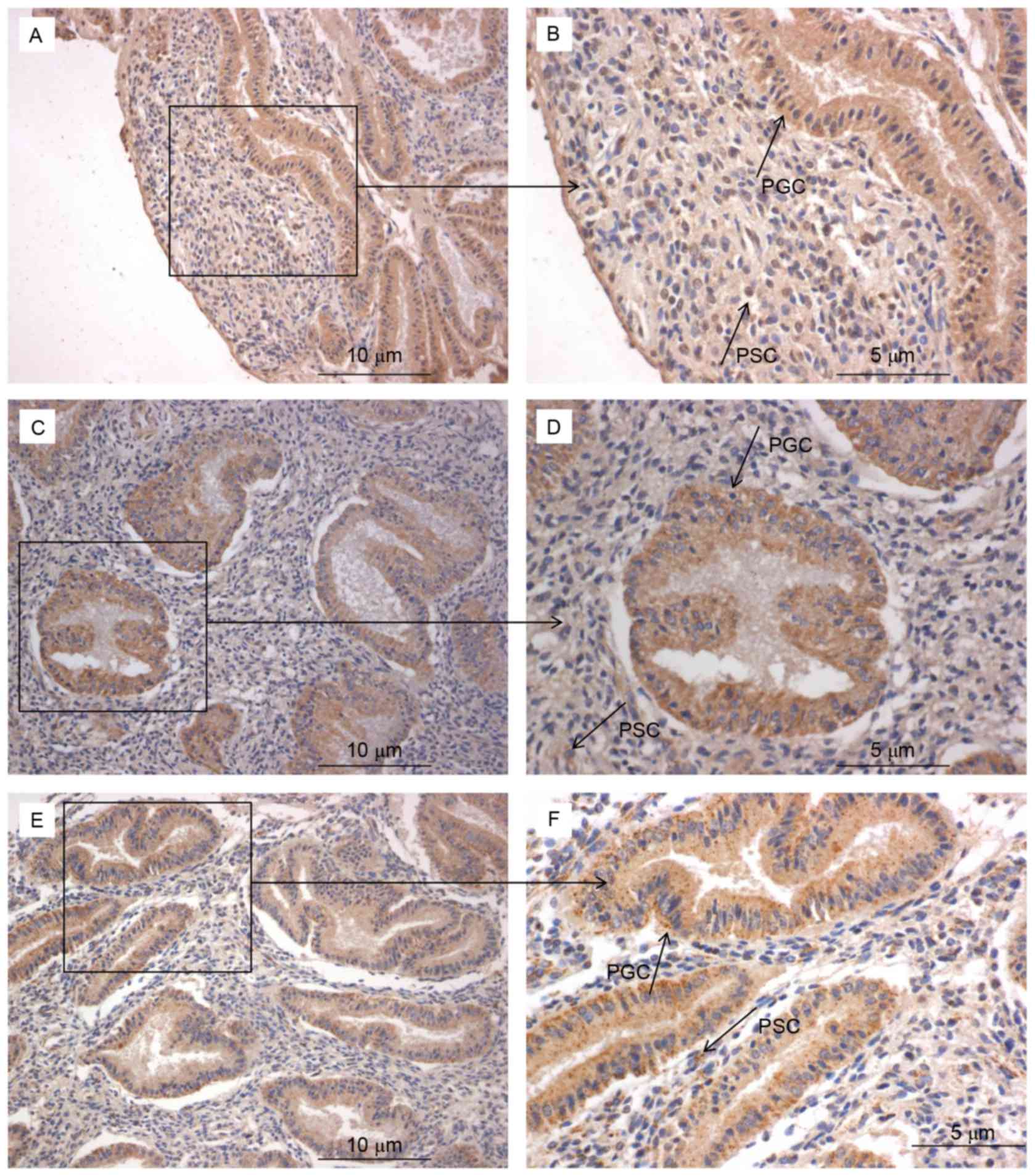
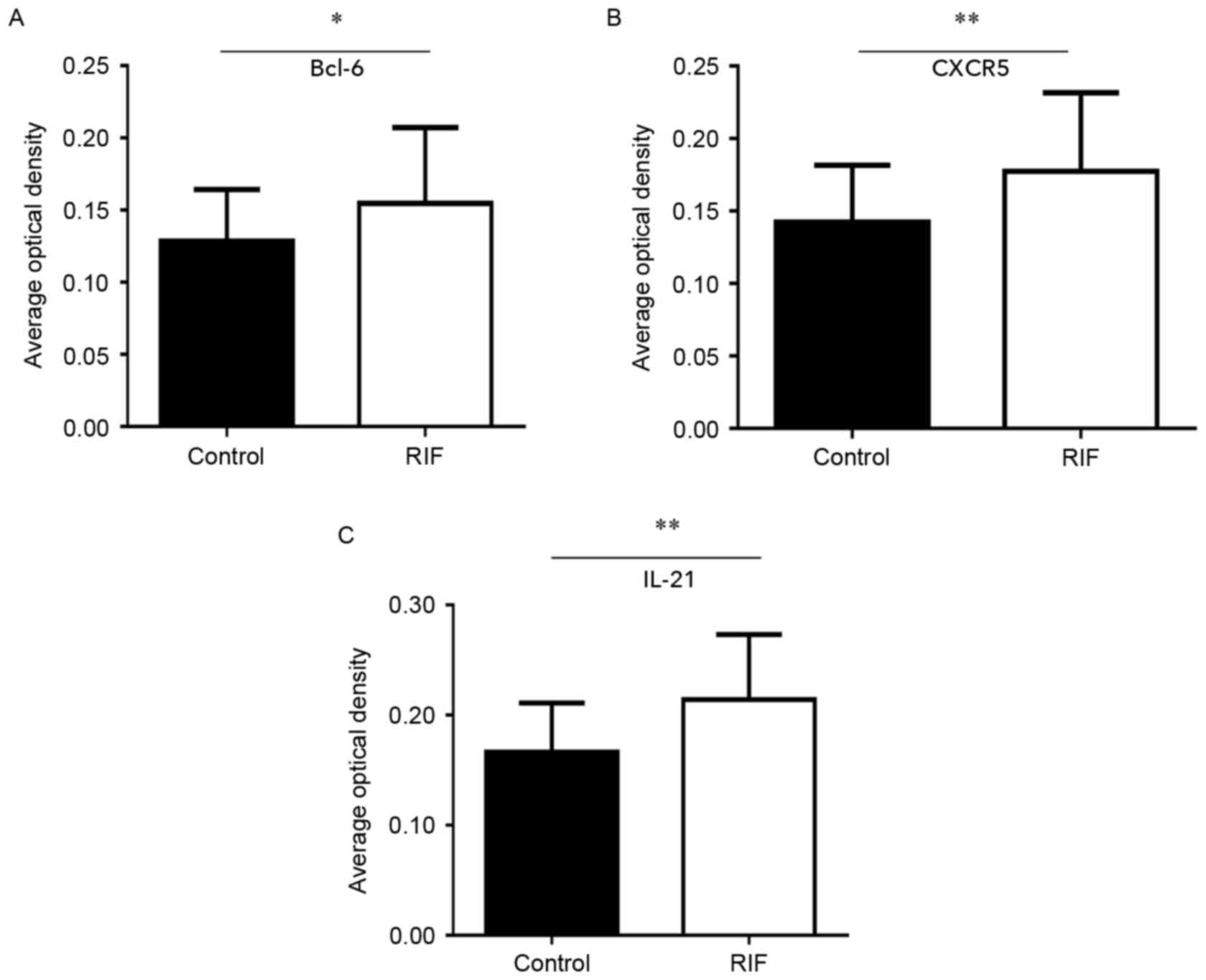
Protein expression of Bcl-6, IL-21 and
CXCR5 is increased within the endometrium during the mid luteal
phase
An improved understanding of the modulation of
Bcl-6, IL-21 and CXCR5 molecules in the endometrium may reveal the
underlying mechanisms of preparation by the endometrium for the
arrival of an embryo. This in turn could provide novel insights
into RIF by exposing deviations from normal activity. Firstly,
H&E staining was performed to observe the pathological
morphology of endometrial tissue (Fig.
3). Nuclear palisading, ferning of the glandular epithelium and
several lymphocytes were observed. Secondly, immunohistochemistry
was used to examine the expression and localization of Bcl-6
(Fig. 4A and B), IL-21 (Fig. 4C and D) and CXCR5 (Fig. 4E and F) in the endometrium. These
factors were expressed in glandular cells, endometrial stroma and
lymphocytes. Finally, quantification of Bcl-6, IL-21 and CXCR5
immunostaining was performed. The expression of Bcl-6 (0.15±0.05;
Fig. 5A), IL-21 (0.21±0.06; Fig. 5B) and CXCR5 (0.18±0.06; Fig. 5C) in RIF endometrium were
significantly increased compared with the control group Bcl-6
(0.13±0.04), IL-21 (0.17±0.04) and CXCR5 (0.14±0.04) levels (all
P<0.05). These results suggest that high expression levels of
Bcl-6, CXCR5 and IL-21 proteins in the endometrium during the
middle luteal phase of menstruation.
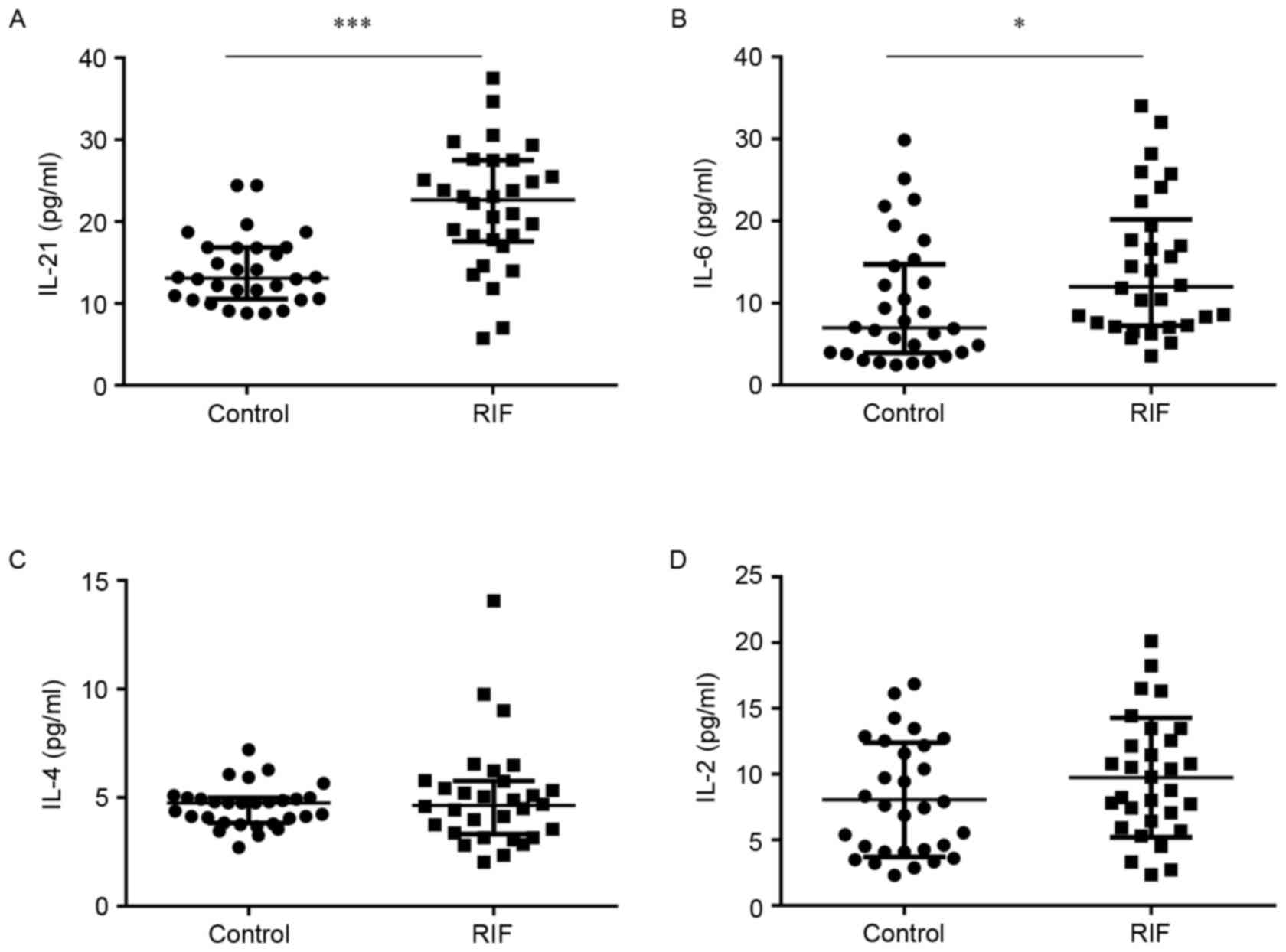
Serum IL-21 and IL-6 concentrations
are increased in females with RIF
Systemic immunity is affected by cytokines; certain
variations in their expression confer an advantage for embryo
implantation, whereas others do not (37,38). In
the current study, ELISAs or CBAs were used to estimate the
concentrations of IL-21, IL-6, IL-4 and IL-2 in the serum. The
concentration of IL-21 was significantly higher in the RIF group
(21.80±7.40) compared to the control group (14.01±4.22) (P<0.05;
Fig. 6A). Similarly, IL-6 levels
were significantly increased in the RIF group (14.45±8.54) compared
with the control group (9.87±7.58) (P<0.05; Fig. 6B). However, the levels of IL-4
(5.03±2.44) and IL-2 (9.74±4.55) in the RIF group did not exhibit a
significant difference compared with the control group (4.60±0.97;
Fig. 6C and 8.05±4.35; Fig. 6D). These results suggest that
increased concentrations of IL-21 and IL-6 in the serum. Whether
overexpression of IL-21 and IL-6 correlates to cTfh levels requires
further research.
Correlation analysis
To find correlations between cTfh cells and
associated factors in patients with RIF, Pearson correlation
analyses were performed. It was revealed that the proportion of
cTfh cells was positively correlated with IL-21 and IL-6
(P<0.01; Table III). No
significant correlations were identified between CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells and IL-4 and IL-2. As
shown in Table IV, IL-21 mRNA
levels were significantly positively correlated with the levels of
Bcl-6 and CXCR5 in the endometrium (P<0.05). These results
suggest that CCR7(lo)PD-1(hi) CXCR5+ CD4+ T
cells and cytokines synergistically regulate local immunity in
women with RIF, and that the occurrence of RIF is not caused by a
single factor.
| Table III.Correlation between CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells and cytokines in the
peripheral blood of patients with repeated implantation
failure. |
Table III.
Correlation between CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells and cytokines in the
peripheral blood of patients with repeated implantation
failure.
|
|
| Cytokine
(pg/ml) |
|---|
|
|
|
|
|---|
| Cell type | Correlation | IL-21 | IL-6 | IL-4 | IL-2 |
|---|
|
CCR7(lo)PD-1(hi) | Correlation
coefficient |
0.649 | 0.497 | 0.279 | 0.358 |
| CXCR5+
CD4+ T | P-value |
<0.001b |
0.005a | 0.135 | 0.052 |
| Table IV.Correlation between IL-21 and CXCR5
and Bcl-6 mRNA in the endometrium of patients with repeated
implantation failure. |
Table IV.
Correlation between IL-21 and CXCR5
and Bcl-6 mRNA in the endometrium of patients with repeated
implantation failure.
|
|
| mRNA |
|---|
|
|
|
|
|---|
| Factors | Correlation | CXCR5 | Bcl-6 |
|---|
| IL-21 | Correlation
coefficient | 0.544 | 0.569 |
|
| P-value | 0.002a |
0.001a |
Discussion
In the present study, CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells and associated factors
were detected in increased concentrations in patients with RIF
compared to the control group during the mid-luteal phase in the
endometrium and the peripheral blood. This suggests that
CCR7(lo)PD-1(hi) CXCR5+ CD4+ T cells are
involved in the pathogenesis and pathophysiology of RIF. Increased
PD-1 on the surface of Tfh cells represents an activated status. By
co-culturing with autologous memory B cells, a previous study
identified that CCR7(lo)PD-1(hi) CXCR5+ but not naive
CD4+ T cells potently induced CD27 (hi) CD38 (hi)
plasmablast or plasma cell differentiation and total IgG production
(26). Similar results were
identified in the studies of Zhang et al (27) and Akiyama et al (39). This indicates that CCR7(lo)PD-1(hi)
CXCR5+ CD4+ T cells can induce RIF by
promoting antibody production.
Autoantibodies and alloantibodies have been
identified to be associated with infertility, miscarriage and
embryo implantation failure (40,41). The
embryo, as a semi-allograft, can trigger antigenic responses and
induce the production of maternal antibodies (41,42).
These alloantibodies have no pathogenic effect on the embryo; on
the contrary, a lack of these antibodies can cause recurrent
miscarriage (42). Women with
recurrent miscarriage and RIF also have higher levels of
autoantibodies, including antinuclear autoantibodies,
antiphospholipid antibodies, anti-thyroperoxidase and
anti-thyroglobulin antibodies (43–45).
These autoantibodies can damage the endometrium and hinder the
implantation of the embryo, resulting in infertility and recurrent
spontaneous abortion (43,46). A lack of alloantibodies and
overexpression of autoantibodies can lead to abnormalities in
humoral immunity (42,44). Tfh cells prime B cells to initiate
antibody responses and maintain humoral immunity (22,23).
Realizing the role of Tfh cells in embryo implantation is crucial
for the development of potential novel therapies against RIF.
Successful implantation of an embryo requires
synchronous development of the embryo and the endometrium (47,48).
Endometrial receptivity is generally at its highest during the
mid-luteal phase of the menstrual cycle, which makes it the best
time for implantation. Reduced receptivity can result in failed or
inadequate implantation (49). This
means that the microenvironment provided by uterine fluid,
particularly glandular secretions, is essential for implantation.
Analysis of endometrial fluid has identified cytokines, chemokines,
proteases, antiproteases and other factors that modulate blastocyst
functions relevant to implantation (50). The present study discovered that
Bcl-6, CXCR5 and IL-21 were highly expressed in endometrial glands
and the stroma of the RIF group. These results suggest that
Tfh-associated factors may be involved in regulating the
implantation microenvironment.
During early pregnancy, immune cells are recruited
extensively to the endometrium. Chemokines and their receptors
mediate immune cell recruitment and subsequent chronic activation
of the immune system. They serve an important role in embryonic
attachment and placental development (51). Franasiak et al (52) revealed that the chemokine CXCL13
reached optimal levels in the endometrium during the mid-luteal
phase. Notably, the present study identified that the level of its
receptor CXCR5 was increased in the endometrium during the same
phase. CXCR5 is critical for the migration of Tfh and B cells into
B cell follicles (26). Therefore,
it has been suggested that the increased CXCR5 was involved in Tfh
cells. In vivo, CXCR5 is closely associated with Bcl-6
expression (53). Bcl-6 is involved
in migration control, differentiation and products of Tfh cells,
and is also required for the formation of GCs. Overexpression of
Bcl-6 results in the repression of CCR7 and the upregulation of
PD-1 and CXCR5 (54). Bcl-6 is
usually upregulated in GC Tfh cells and B cells, but hardly
expressed in non-Tfh and cTfh cells (16,17).
Normally, Bcl-6 acts mainly as a transcriptional repressor,
interfering with the differentiation of non-Tfh cells; for
instance, by antagonizing the expression of transcription factors
T-bet, trans-acting T-cell-specific transcription factor GATA-3 and
nuclear receptor ROR γ inhibit polarization of Th1, Th2 and Th17
cells (53,55,56).
Previous studies have demonstrated that Bcl-6 is the master
regulator of Tfh cell differentiation (16,54).
Bcl-6 is upregulated during Tfh cell differentiation and
Bcl6− T cells fail to differentiate into Tfh cells. In
the present study, the expression of Bcl-6 was clearly increased in
the RIF endometrium, and there was a significant positive
correlation between Bcl-6 and IL-21 levels.
IL-21 is the main cytokine secreted by Tfh cells, it
can enhance the differentiation of Tfh cells, regulate B cell
differentiation and proliferation, stimulate plasma cell
differentiation and immunoglobulin production, and induce the
expression of Bcl-6 (57). IL-21 is
the most potent inducer of plasma-cell differentiation in
vitro (58,59). Altered IL-21 production induces
miscarriage by promoting an inflammatory state. In the present
study, the expression of IL-21 was similar to that of Bcl-6 and
CXCR5 in the RIF endometrium. Wang et al (60) observed that interferon regulatory
factor 4 mediated Th17 cell activation and upregulated the
expression of IL-17A and IL-21, which resulted in pregnancy loss.
Messaoudi et al (61)
revealed that polymorphisms of IL-21 were associated with recurrent
spontaneous miscarriage through pro-inflammatory pathways. The
results of the present study demonstrated that the mRNA and protein
levels of Blc-6, CXCR5 and IL-21 were increased in the RIF
endometrium at the mid luteal phase. Furthermore, Blc-6 and CXCR5
levels had a positive correlation with IL-21 levels. From the
results of the current study and the previous studies discussed, it
was proposed that increased Bcl-6, CXCR5 and IL-21 collaborate to
cause an imbalance of the endometrial immune microenvironment in
women with RIF. The activation of these factors may be involved in
Tfh and B cell differentiation and antibody secretion.
Cytokines are critical for embryo implantation and
pregnancy maintenance (12,13). Several cytokines are involved in the
regulation of Tfh cell differentiation. IL-21, IL-6 and IL-4 are
considered to support Tfh differentiation, while IL-2 is a potent
inhibitor of this. IL-6 is a pro-inflammatory cytokine secreted by
numerous cell types. IL-6 derived from follicular dendritic cells
is essential for Tfh cell maintenance (62). Plasmablast-derived IL-6 can also
increase the number of human cTfh cells (63). IL-6 induces an early wave of Bcl-6
expression and is important for the initiation of Tfh cell
production. A lack of IL-6 results in a severe reduction in
CXCR5+ Bcl6+ early Tfh cells in vivo
(64). Previously, a significantly
higher expression of IL-6 in women with unexplained recurrent
spontaneous abortion has been reported (12). IL-21 and IL-4 are two cytokines
expressed by Tfh cells (19,65,66).
Production of IL-4 and IL-21 is coupled in Tfh cells and required
for optimal antibody responses (19,20).
IL-21 is a novel susceptibility gene for recurrent spontaneous
miscarriage (61). Upregulating the
expression of IL-21 may result in pregnancy loss (60) and infertility (31). In the current study, IL-21 and IL-6
were increased in the serum of the RIF group, but IL-4 and IL-2
levels did not demonstrate a statistically significant difference
compared to the control group. There is a significant positive
correlation between the number of cTfh cells and IL-21 and IL-6
levels. These results suggest that IL-21 may co-operate with IL-6,
resulting in RIF by promoting Tfh cell differentiation.
In conclusion, the results of the present study
indicate that CCR7(lo)PD-1(hi) CXCR5+ CD4+ T
cells, CXCR5, IL-21, Bcl-6 and IL-6 are involved in the
pathogenesis of RIF. Whether CCR7(lo)PD-1(hi) CXCR5+
CD4+ T cells and associated factors contribute to
abnormal humoral immunity in patients with RIF requires additional
research. CCR7(lo)D-1(hi) CXCR5+ CD4+ T cells
and associated factors could be a potential therapeutic target to
help reduce the risk of implantation failure.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Xinjiang Autonomous Region (grant no.
2013211A087) and the National Natural Science Foundation of China
(grant nos. 81660343, 8160202 and 81460307).
Glossary
Abbreviations
Abbreviations:
|
RIF
|
repeated implantation failure
|
|
Bcl-6
|
B-cell lymphoma 6
|
|
CXCR5
|
chemokine receptor type 5
|
|
Tfh
|
follicular helper T
|
|
cTfh
|
circulating Tfh
|
|
IVF-ET
|
in vitro fertilization-embryo
transfer
|
|
CCR7
|
chemokine receptor 7
|
|
PD-1
|
programmed death-1
|
References
|
1
|
Valenzuela OA, Couturier-Tarrade A, Choi
YH, Aubrière MC, Ritthaler J, Chavatte-Palmer P and Hinrichs K:
Impact of equine assisted reproductive technologies (standard
embryo transfer or intracytoplasmic sperm injection (ICSI) with in
vitro culture and embryo transfer) on placenta and foal morphometry
and placental gene expression. Reprod Fertil Dev. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dekel N, Gnainsky Y, Granot I, Racicot K
and Mor G: The role of inflammation for a successful implantation.
Am J Reprod Immunol. 72:141–147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ledee N, Petitbarat M, Chevrier L, Vitoux
D, Vezmar K, Rahmati M, Dubanchet S, Gahéry H, Bensussan A and
Chaouat G: The uterine immune profile may help women with repeated
unexplained embryo implantation failure after in vitro
fertilization. Am J Reprod Immunol. 75:388–401. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crawford G, Ray A, Gudi A, Shah A and
Homburg R: The role of seminal plasma for improved outcomes during
in vitro fertilization treatment: Review of the literature and
meta-analysis. Hum Reprod Update. 21:275–284. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tan BK, Vandekerckhove P, Kennedy R and
Keay SD: Investigation and current management of recurrent IVF
treatment failure in the UK. BJOG. 112:773–780. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Margalioth EJ, Ben-Chetrit A, Gal M and
Eldar-Geva T: Investigation and treatment of repeated implantation
failure following IVF-ET. Hum Reprod. 21:3036–3043. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Simon A and Laufer N: Repeated
implantation failure: Clinical approach. Fertil Steril.
97:1039–1043. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakagawa K, Kwak-Kim J, Ota K, Kuroda K,
Hisano M, Sugiyama R and Yamaguchi K: Immunosuppression with
tacrolimus improved reproductive outcome of women with repeated
implantation failure and elevated peripheral blood TH1/TH2 cell
ratios. Am J Reprod Immunol. 73:353–361. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Persson M, Ekerfelt C, Jablonowska B,
Jonsson Y, Ernerudh J, Jenmalm MC and Berg G: Immunological status
in patients undergoing in vitro fertilisation: Responses to hormone
treatment and relationship to outcome. J Reprod Immunol. 96:58–67.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schlossberger V, Schober L, Rehnitz J,
Schaier M, Zeier M, Meuer S, Schmitt E, Toth B, Strowitzki T and
Steinborn A: The success of assisted reproduction technologies in
relation to composition of the total regulatory T cell (Treg) pool
and different Treg subsets. Hum Reprod. 28:3062–3073. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou J, Wang Z, Zhao X, Wang J, Sun H and
Hu Y: An increase of Treg cells in the peripheral blood is
associated with a better in vitro fertilization treatment outcome.
Am J Reprod Immunol. 68:100–106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saifi B, Rezaee SA, Tajik N, Ahmadpour ME,
Ashrafi M, Vakili R, SoleimaniAsl S, Aflatoonian R and Mehdizadeh
M: Th17 cells and related cytokines in unexplained recurrent
spontaneous miscarriage at the implantation window. Reprod Biomed
Online. 29:481–489. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang PY, Diao LH, Huang CY, Lian RC, Chen
X, Li GG, Zhao J, Li YY, He XB and Zeng Y: The pro-inflammatory and
anti-inflammatory cytokine profile in peripheral blood of women
with recurrent implantation failure. Reprod Biomed Online.
31:823–826. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Figueiredo MM, Costa PAC, Diniz SQ,
Henriques PM, Kano FS, Tada MS, Pereira DB, Soares IS,
Martins-Filho OA, Jankovic D, et al: T follicular helper cells
regulate the activation of B lymphocytes and antibody production
during Plasmodium vivax infection. PLoS Pathog.
13:e10064842017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Monteiro C, Kasahara TM, Castro JR,
Sacramento PM, Hygino J, Centurião N, Cassano T, Lopes LMF, Leite
S, Silva VG, et al: Pregnancy favors the expansion of circulating
functional follicular helper T Cells. J Reprod Immunol. 121:1–10.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tangye SG, Ma CS, Brink R and Deenick EK:
The good, the bad and the ugly-TFH cells in human health and
disease. Nat Rev Immunol. 13:412–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ueno H: Human circulating T follicular
helper cell subsets in health and disease. J Clin Immunol. 36 Suppl
1:S34–S39. 2016. View Article : Google Scholar
|
|
18
|
Vinuesa CG, Linterman MA, Yu D and
MacLennan IC: Follicular helper T Cells. Annu Rev Immunol.
34:335–368. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weinstein JS, Herman EI, Lainez B,
Licona-Limón P, Esplugues E, Flavell R and Craft J: TFH cells
progressively differentiate to regulate the germinal center
response. Nat Immunol. 17:1197–1205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McGuire HM, Vogelzang A, Warren J, Loetsch
C, Natividad KD, Chan TD, Brink R, Batten M and King C: IL-21 and
IL-4 collaborate to shape T-dependent antibody responses. J
Immunol. 195:5123–5135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schmitt N, Bentebibel SE and Ueno H:
Phenotype and functions of memory Tfh cells in human blood. Trends
Immunol. 35:436–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Crotty S: T follicular helper cell
differentiation, function and roles in disease. Immunity.
41:529–542. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Badell IR and Ford ML: T follicular helper
cells in the generation of alloantibody and graft rejection. Curr
Opin Organ Transplant. 21:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nurieva RI, Podd A, Chen Y, Alekseev AM,
Yu M, Qi X, Huang H, Wen R, Wang J, Li HS, et al: STAT5 protein
negatively regulates T follicular helper (Tfh) cell generation and
function. J Biol Chem. 287:11234–11239. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arroyo-Villa I, Bautista-Caro MB, Balsa A,
Aguado-Acín P, Bonilla-Hernán MG, Plasencia C, Villalba A, Nuño L,
Puig-Kröger A, Martín-Mola E and Miranda-Carús ME: Constitutively
altered frequencies of circulating follicullar helper T cell
counterparts and their subsets in rheumatoid arthritis. Arthritis
Res Ther. 16:5002014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He J, Tsai LM, Leong YA, Hu X, Ma CS,
Chevalier N, Sun X, Vandenberg K, Rockman S, Ding Y, et al:
Circulating precursor CCR7(lo)PD-1(hi) CXCR5+
CD4+ T cells indicate Tfh cell activity and promote
antibody responses upon antigen reexposure. Immunity. 39:770–781.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang F, Pang N, Zhu Y, Zhou D, Zhao H, Hu
J, Ma X, Li J, Wen H, Samten B, et al: CCR7(lo)PD-1(hi) CXCR5(+)
CD4(+) T cells are positively correlated with levels of IL-21 in
active and transitional cystic echinococcosis patients. BMC Infect
Dis. 15:4572015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu R, Li X, Zhang Z, Zhou M, Sun Y, Su D,
Feng X, Gao X, Shi S, Chen W and Sun L: Allogeneic mesenchymal stem
cells inhibited T follicular helper cell generation in rheumatoid
arthritis. Sci Rep. 5:127772015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Blanco P, Ueno H and Schmitt N: T
follicular helper (Tfh) cells in lupus: Activation and involvement
in SLE pathogenesis. Eur J Immunol. 46:281–290. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Le Coz C, Joublin A, Pasquali JL, Korganow
AS, Dumortier H and Monneaux F: Circulating TFH subset distribution
is strongly affected in lupus patients with an active disease. PLoS
One. 8:e753192013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu H, Liu J, Cui X, Zuo Y, Zhang Z, Li Y,
Tao R, Li Y and Pang J: Increased frequency of circulating
follicular helper T cells in lupus patients is associated with
autoantibody production in a CD40L-dependent manner. Cell Immunol.
295:46–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park HJ, Kim DH, Lim SH, Kim WJ, Youn J,
Choi YS and Choi JM: Insights into the role of follicular helper T
cells in autoimmunity. Immune Netw. 14:21–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shi J, Luo F, Shi Q, Xu X, He X and Xia Y:
Increased circulating follicular helper T cells with decreased
programmed death-1 in chronic renal allograft rejection. BMC
Nephrol. 16:1822015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lu KH, Loose DS, Yates MS,
Nogueras-Gonzalez GM, Munsell MF, Chen LM, Lynch H, Cornelison T,
Boyd-Rogers S, Rubin M, et al: Prospective multicenter randomized
intermediate biomarker study of oral contraceptive versus
depo-provera for prevention of endometrial cancer in women with
Lynch syndrome. Cancer Prev Res (Phila). 6:774–781. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jia L, Wang Y, Li J, Li S, Zhang Y, Shen
J, Tan W and Wu C: Detection of IL-9 producing T cells in the PBMCs
of allergic asthmatic patients. BMC immunology. 18:382017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Franasiak JM and Scott RT: Contribution of
immunology to implantation failure of euploid embryos. Fertil
Steril. 107:1279–1283. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang WJ, Liu FJ, Zhang X, Liu XM, Qu QL,
Li FH, Zhuang LL, Li XX and Hao CF: Periodic elevation of
regulatory T cells on the day of embryo transfer is associated with
better in vitro fertilization outcome. J Reprod Immunol. 119:49–53.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akiyama M, Yasuoka H, Yamaoka K, Suzuki K,
Kaneko Y, Kondo H, Kassai Y, Koga K, Miyazaki T, Morita R, et al:
Enhanced IgG4 production by follicular helper 2 T cells and the
involvement of follicular helper 1 T cells in the pathogenesis of
IgG4-related disease. Arthritis Res Ther. 18:1672016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
An LF, Zhang XH, Sun XT, Zhao LH, Li S and
Wang WH: Unexplained infertility patients have increased serum
IL-2, IL-4, IL-6, IL-8, IL-21, TNFα, IFNgamma and increased Tfh/CD4
T cell ratio: Increased Tfh and IL-21 strongly correlate with
presence of autoantibodies. Immunol Invest. 44:164–173. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Perricone C, de Carolis C and Perricone R:
Pregnancy and autoimmunity: A common problem. Best Pract Res Clin
Rheumatol. 26:47–60. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen JL, Yang JM, Huang YZ and Li Y:
Clinical observation of lymphocyte active immunotherapy in 380
patients with unexplained recurrent spontaneous abortion. Int
Immunopharmacol. 40:347–350. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ticconi C, Pietropolli A, Borelli B, Bruno
V, Piccione E, Bernardini S and Di Simone N: Antinuclear
autoantibodies and pregnancy outcome in women with unexplained
recurrent miscarriage. Am J Reprod Immunol. 76:396–399. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Breen KA, Sanchez K, Kirkman N, Seed PT,
Parmar K, Moore GW and Hunt BJ: Endothelial and platelet
microparticles in patients with antiphospholipid antibodies. Thromb
Res. 135:368–374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kwak-Kim J, Skariah A, Wu L, Salazar D,
Sung N and Ota K: Humoral and cellular autoimmunity in women with
recurrent pregnancy losses and repeated implantation failures: A
possible role of vitamin D. Autoimmun Rev. 15:943–947. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Unuane D, Velkeniers B, Deridder S,
Bravenboer B, Tournaye H and De Brucker M: Impact of thyroid
autoimmunity on cumulative delivery rates in in vitro
fertilization/intracytoplasmic sperm injection patients. Fertil
Steril. 106:144–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Valdes CT, Schutt A and Simon C:
Implantation failure of endometrial origin: It is not pathology,
but our failure to synchronize the developing embryo with a
receptive endometrium. Fertil Steril. 108:15–18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Teh WT, McBain J and Rogers P: What is the
contribution of embryo-endometrial asynchrony to implantation
failure? J Assist Reprod Genet. 33:1419–1430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kumar V, Soni UK, Maurya VK, Singh K and
Jha RK: Integrin beta8 (ITGB8) activates VAV-RAC1 signaling via FAK
in the acquisition of endometrial epithelial cell receptivity for
blastocyst implantation. Sci Rep. 7:18852017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Salamonsen LA, Evans J, Nguyen HP and
Edgell TA: The microenvironment of human implantation: Determinant
of reproductive success. Am J Reprod Immunol. 75:218–225. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bidarimath M, Khalaj K, Kridli RT, Wessels
JM, Koti M and Tayade C: Altered expression of chemokines and their
receptors at porcine maternal-fetal interface during early and
mid-gestational fetal loss. Cell Tissue Res. 366:747–761. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Franasiak JM, Burns KA, Slayden O, Yuan L,
Fritz MA, Korach KS, Lessey BA and Young SL: Endometrial CXCL13
expression is cycle regulated in humans and aberrantly expressed in
humans and Rhesus macaques with endometriosis. Reprod Sci.
22:442–451. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Choi YS, Yang JA, Yusuf I, Johnston RJ,
Greenbaum J, Peters B and Crotty S: Bcl6 expressing follicular
helper CD4 T cells are fate committed early and have the capacity
to form memory. J Immunol. 190:4014–4026. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Nurieva RI, Chung Y, Martinez GJ, Yang XO,
Tanaka S, Matskevitch TD, Wang YH and Dong C: Bcl6 mediates the
development of T follicular helper cells. Science. 325:1001–1005.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hatzi K, Nance JP, Kroenke MA, Bothwell M,
Haddad EK, Melnick A and Crotty S: BCL6 orchestrates Tfh cell
differentiation via multiple distinct mechanisms. J Exp Med.
212:539–553. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Vaeth M, Eckstein M, Shaw PJ, Kozhaya L,
Yang J, Berberich-Siebelt F, Clancy R, Unutmaz D and Feske S:
Store-operated Ca(2+) entry in follicular T cells controls humoral
immune responses and autoimmunity. Immunity. 44:1350–1364. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Belanger S and Crotty S: Dances with
cytokines, featuring TFH cells, IL-21, IL-4 and B cells. Nat
Immunol. 17:1135–1136. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Spolski R and Leonard WJ: Interleukin-21:
A double-edged sword with therapeutic potential. Nat Rev Drug
Discov. 13:379–395. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Tangye SG: Advances in IL-21
biology-enhancing our understanding of human disease. Curr Opin
Immunol. 34:107–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang J, Yin T, Wen Y, Tian F, He X, Zhou
D, Lin Y and Yang J: Potential effects of interferon regulatory
factor 4 in a murine model of polyinosinic-polycytidylic
acid-induced embryo resorption. Reprod Fertil Dev. 2015.
|
|
61
|
Messaoudi S, Al-Khateeb GM, Dendana M,
Sater MS, Jazia KB, Nouira M, Almawi WY and Mahjoub T: Genetic
variations in the interleukin-21 gene and the risk of recurrent
idiopathic spontaneous miscarriage. Eur Cytokine Netw. 22:123–126.
2011.PubMed/NCBI
|
|
62
|
Harker JA, Lewis GM, Mack L and Zuniga EI:
Late interleukin-6 escalates T follicular helper cell responses and
controls a chronic viral infection. Science. 334:825–829. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chavele KM, Merry E and Ehrenstein MR:
Cutting edge: Circulating plasmablasts induce the differentiation
of human T follicular helper cells via IL-6 production. J Immunol.
194:2482–2485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Choi YS, Eto D, Yang JA, Lao C and Crotty
S: Cutting edge: STAT1 is required for IL-6-mediated Bcl6 induction
for early follicular helper cell differentiation. J Immunol.
190:3049–3053. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sahoo A, Wali S and Nurieva R: T helper 2
and T follicular helper cells: Regulation and function of
interleukin-4. Cytokine Growth Factor Rev. 30:29–37. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Fairfax KC, Everts B, Amiel E, Smith AM,
Schramm G, Haas H, Randolph GJ, Taylor JJ and Pearce EJ:
IL-4-secreting secondary T follicular helper (Tfh) cells arise from
memory T cells, not persisting Tfh cells, through a B
cell-dependent mechanism. J Immunol. 194:2999–3010. 2015.
View Article : Google Scholar : PubMed/NCBI
|