Introduction
Pseudomonas (P.) aeruginosa is
a ubiquitous and metabolically versatile opportunistic pathogen,
which may colonize in the respiratory tract/gastrointestinal mucosa
of patients as well as medical equipment, including catheters and
intubation tubes. This Gram-negative bacterium may cause a series
of infectious diseases, including keratitis, otitis media and
bacteremia, in immuno-compromised patients and those in intensive
care units with high morbidity and mortality (1). Due to extensive application of
antibiotics, an increasing number of P. aeruginosa isolates
have been identified to be resistant to various traditional
antibacterial agents, including penicillins, fluoroquinolones,
aminoglycosides, polymyxins as well as carbapenems, via intrinsic
as well as acquired resistance mechanisms (1). One of the essential factors
contributing to resistance is associated with efflux pumps, of
which four types, namely MexAB-outer membrane protein (Opr)M,
MexEF-OprN, MexCD-OprJ and MexXY-OprM, have been widely studied
(2). Kiser et al (3) reported that OprM and a group of efflux
regulatory genes or efflux protein (NfxB, MexR and MexZ)
contributed to multidrug resistance in clinical P.
aeruginosa isolates.
Although MexAB-OprM is thought to be mostly
accountable for carbapenem resistance in P. aeruginosa
rather than MexXY-OprM (4,5), increasing evidence indicated that
MexXY-OprM was also inclined to be overexpressed in clinical P.
aeruginosa isolates. In A Brazilian study by Xavier et
al (6) demonstrated that
MexXY-OprM was overexpressed in 50.8% of P. aeruginosa
isolates studied compared with 27.1% for MexAB-OprM. A study from
Thailand by Khuntayaporn et al (7) observed that, increased expression of
MexAB-OprM mRNA accounted for 92.06% followed by 63.49% for
MexXY-OprM mRNA. Obviously, the percentages of MexAB-OprM and
MexXY-OprM overexpression exhibited geographical variations.
Imipenem (IMP), a carbapenem, has an important role
in the treatment of infections caused by multidrug-resistant P.
aeruginosa. However, it has been reported that 20% of P.
aeruginosa isolates were resistant to IMP (8). Due to this IMP resistance in P.
aeruginosa novel antibacterial agents are urgently required.
Berberine (BEB), a natural isoquinoline alkaloid, is prevalent in
numerous medicinal plants and has been demonstrated to have
antibacterial and antifungal activities alone or in combination
with other drugs (9–13). In a previous study, BEB was reported
to be a potential inhibitor of MexXY-mediated resistance in P.
aeruginosa (14). However, the
antipseudomonas activity of BEB in combination with other
traditional antibiotics, including IMP, in the treatment of
clinical P. aeruginosa isolates has remained largely
elusive.
The present study aimed to investigate the efficacy
of BEB alone and in combination with IMP against a clinical P.
aeruginosa isolate and assessed the underlying mechanism from
the viewpoint of the MexXY-OprM efflux pump.
Materials and methods
Bacterial strains
The P. aeruginosa ATCC27853 strain was
obtained from the National Institute for the Control of
Pharmaceutical and Biological Products (Beijing, China). The
clinical isolate, PA012, was isolated from a patient with a urinary
tract infection at and provided by the Clinical Laboratory of
Weifang People's Hospital (Weifang, China). The present study was
approved by the ethics committee of the Weifang People's Hospital.
Written informed consent was obtained from the patient prior to the
collection of PA012. The strains were stored at −80°C and subjected
to cultivation in Luria-Bertani medium (Hope, Qingdao, China) with
constant shaking at 8 × g at 37°C for 18 h until the strains
reached the late logarithmic growth phase.
Susceptibility assay
The minimum inhibitory concentrations (MICs) of BEB
and IMP were assessed using the broth microdilution method
following the criteria of the Clinical and Laboratory Standards
Institute (no. M07-A10) (15). The
bacterial suspension was adjusted to 1×105
colony-forming units (CFU)/ml and incubated with BEB/IMP in a
96-well flat-bottomed microtiter plate for 24 h at 35°C. The
concentrations of BEB/IMP were serially two-fold diluted in a range
of 2–1,014 µg/ml for BEB and 0.25–128 µg/ml for IMP. The MIC was
defined as the lowest concentration of BEB/IMP to cause no visual
growth of bacteria. The combination of BEB and IMP was evaluated by
checkerboard assay. The concentrations of BEB and IMP were set in a
range of 1/64-1 MIC for BEB and 1/64–16 MIC for IMP. The fractional
inhibitory concentration index (FICI) was determined using the
following equation: FICI=(MICBEB in
combination/MICBEB alone) + (MICIMP in
combination/MICIMP alone), in which synergism,
indifference and antagonism were interpreted as FICI ≤0.5, >0.5
and ≤4.0, and >4.0, respectively (16).
Time-kill (T-K) test
The initial PA012 suspension (1×105
CFU/ml) was incubated with BEB and/or IMP at their concentrations
at which synergism had been previously determined by using the
checkerboard assay (17). The cells
were counted at the designated time-points (0, 4, 8, 12 and 24 h).
The control group remained untreated. T-K curves were plotted on a
logarithmic scale of CFU/ml vs. time (h). Synergism was defined as
an increase of ≥2log10 CFU/ml in the killing rate for the combined
agents compared with the most active agent used alone. Antagonism
was defined as a decrease of ≥2log10 CFU/ml in the killing rate for
the combined agents compared with the most active agent alone.
Indifference was defined as a decrease of ≤2log10 CFU/ml in the
killing rate for the combined agents compared with any of the two
agents used alone (17).
Scanning electron microscopy
(SEM)
Prior to treatments with BEB (1/4 MIC) and/or IMP
(1/8 MIC), a suspension of PA012 (100 µl; 1×105 CFU/ml)
was added into a 6-well flat-bottomed microtiter plate with sterile
cover slips, followed by incubation for 24 h. The cover slips were
then rinsed with sterile PBS, fixed with 2.5% (v/v) glutaraldehyde
overnight at room temperature, and post-fixed with 0.1% osmium
tetroxide for 1 h. The samples were subsequently dehydrated with a
series of 30, 50, 70, 90 and 100% ethanol for 10 min each. After
critical-point drying, the samples were processed by gold
sputtering (JEOL JFC 1200E Ion sputtering device; JEOL Ltd., Tokyo,
Japan) and observed by SEM (JSM-6700F; JEOL Ltd.).
Adhesion test
PA012 suspension (100 µl; 1×105 CFU/ml)
was incubated with BEB (1/4 MIC) in combination with IMP at 1/16,
1/8, 1/4 and 1/2 MIC for 2 or 4 h. The adhesion test was performed
on the surface of medical-grade polyvinyl chloride (Shengli
Oilfield Central Hospital, Dongying, China). The medium was
discarded and the wells were washed with pH 7.2 sterile PBS. The
bacterial cells in each well were then stained with 150 µl 0.1%
crystal violet for 15 min at room temperature. Following washing
with PBS for several times and drying for 3 h, 150 µl 30% (v/v)
acetic acid was added to the stained cells, followed by incubation
with mixing for 15 min. The optical density at the wavelength of
492 nm (OD492) was measured.
BEB uptake
PA012 cells in the late logarithmic growth phase
were centrifuged at 10,000 × g for 5 min at 35°C, washed twice with
0.01 M sterile PBS (pH 7.2) and resuspended in 1 ml PBS to reach an
OD600 of 0.2. The cells were then treated with BEB (1/4
MIC) or in combination with IMP at 1/16, 1/8, 1/4 and 1/2 of the
MIC at 37°C for 10 min. Cells killed in a 100°C water bath were
incubated with the same concentrations of agents were set as the
control. Subsequently, the cells were centrifuged, washed and
resuspended to reach an OD600 of 0.15. The fluorescence
of BEB was determined with a Synergy™ HT multidetection
microplate reader (BioTek, Winooski, VT, USA) at 360 nm excitation
wavelength and 530 nm emission wavelength (18).
Reverse-transcription quantitative
polymerase chain reaction (RT-qPCR) analysis
The PA012 cells (1×105 CFU/ml) were first
treated with BEB (1/4 MIC) and/or IMP (1/8 MIC). The total RNA was
then extracted according to the instructions of the
MagExtractor-RNA-kit (cat. no. NPK-200; Toyobo Life Science, Osaka,
Japan). Complementary DNA was prepared following the instructions
of the ReverTra Ace® qPCR RT Master Mix with gDNA
Remover kit (cat. no. FSQ-301; Toyobo Life Science). Primers with
the following sequences were used: MexZ forward,
5′-CCCTTGTGAGGACGTTCAGT-3′ and reverse, 5′-CCAGCAACAGGTAGGGAGAA-3′;
MexX forward, 5′-CATCAGCGAACGCGAGTACAC-3′ and reverse,
5′-CAATTCGCGATGCGGATTG-3′; MexY forward 5′-CCGCTACAACGGCTATCCCT-3′
and reverse, 5′-AGCGGGATCGACCAGCTTTC-3′; OprM forward
5′-GATCCCCGACTACCAGCGCCCCG-3′ and reverse,
5′-ATGCGGTACTGCGCCCGGAAGGC-3′; OprD forward
5′-ATCTACCGCACAAACGATGAG-3′ and reverse,
5′-GCCGAAGCCGATATAATCAAACG-3′; RpsL forward
5′-GCAAGCGCATGGTCGACAAGA-3′ and reverse,
5′-CGCTGTGCTCTTGCAGGTTGTGA-3′. The 25 µl qPCR mixture consisted of
12.5 µl 2X SYBR® Green Real Time PCR Master mix, 1 µl
PCR forward primer, 1 µl PCR reverse primer, 0.5 µl cDNA and 10 µl
double distilled water. RT-qPCR was performed on an ABI7000
fluorescent qPCR system (Thermo Fisher Scientific, Inc., Waltham,
MA, USA). The thermocycling steps were as follows: 95°C for 10 min,
40 cycles of 95°C for 15 sec, 55°C for 10 sec and 72°C for 10 sec,
and final elongation at 72°C for 5 min. All data were normalized to
reference gene RpsL. The relative target-gene expression was
calculated using the 2−ΔΔCq method (19).
Statistical analysis
Results were obtained from three independent
experiments performed in triplicate. Values are expressed as the
mean ± standard deviation. Statistical analysis was performed using
SPSS 19.0 software (IBM Corp., Armonk, NY, USA). Differences
between groups were determined using one-way analysis of variance
with a Student-Newman-Keuls method post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Synergistic effect of BEB and IMP
against PA012
The MIC of BEB was 512 µg/ml in PA ATCC27853 as well
as in PA012 cells, displaying weak antibacterial activity. Although
the MIC of BEB in PA ATCC27853 cells was decreased 2-fold if used
in combination with IMP at its MIC of 1 µg/ml, it was not
synergistic with IMP, as the FICI was 1.5, which was interpreted as
being indifferent. In the clinical isolate, PA012, the MIC of IMP
was 265 µg/ml, and a synergism of BEB and IMP was identified with
an FICI of 0.375 (<0.5). The MIC of BEB and IMP declined 4-and
8-fold, respectively, in combination compared with each alone
(Table I). The synergism of BEB and
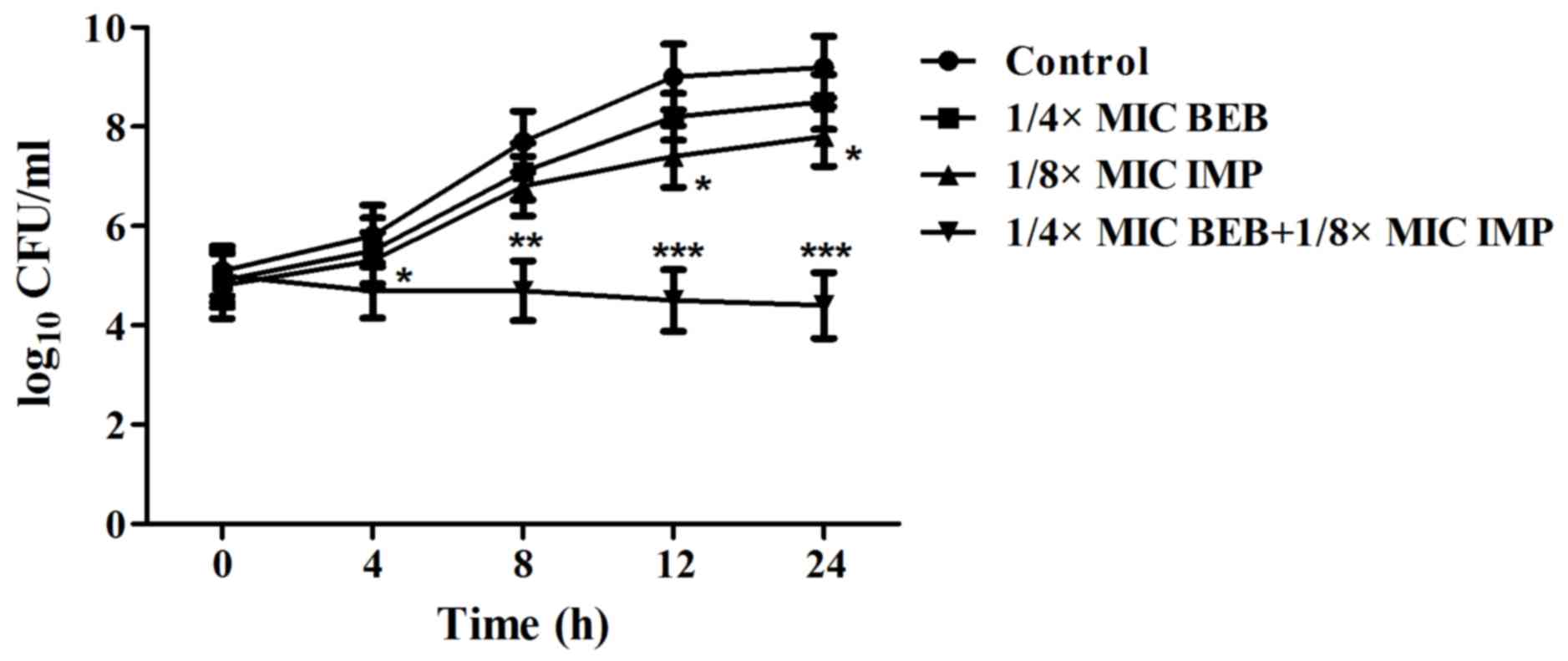
IMP in PA012 cells was also evaluated by a T-K test. The results
indicated that BEB (1/4 MIC) plus IMP (1/8 MIC) or IMP (1/8 MIC)
alone significantly reduced the quantities of PA012 (P<0.05 vs.
the control group), but the growth continued. The combination of
BEB and IMP completely inhibited the bacterial growth, with a
decrease of 3.4 log10 CFU/ml compared with IMP, the most
effective agent, alone (Fig. 1).
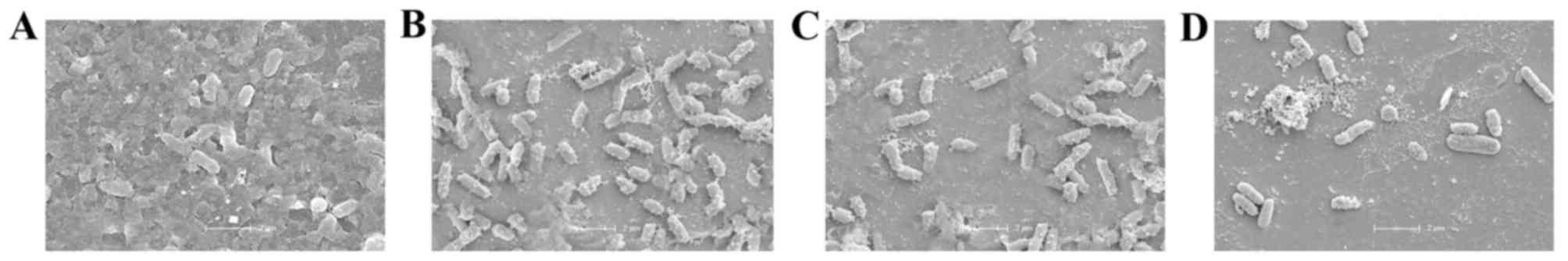
Furthermore, the synergistic activity of BEB with IMP was confirmed
by SEM evaluation, displaying a marked decrease in the density of
PA012 cells compared with that in the control group (Fig. 2).
 | Table I.Interaction of BEB and IMP alone and
in combination against Pseudomonas aeruginosa. |
Table I.
Interaction of BEB and IMP alone and
in combination against Pseudomonas aeruginosa.
|
| MIC alone
(µg/ml) | MIC in combination
(µg/ml) |
|
|---|
|
|
|
|
|
|---|
| Strain | BEB | IMP | BEB | IMP | FICI
(interpretation) |
|---|
| PA ATCC27853 | 512 |
1 | 256 | 1 | 1.5
(indifference) |
| PA012 | 512 | 256 | 128 | 32 | 0.375
(synergism) |
BEB and IMP have synergistic effects
via enhancement of intracellular BEB accumulation and inhibition of
cell adherence
In the subsequent experiments, the concentration of
BEB was set at 1/4 of the MIC to investigate the effects of its
combination with a series of IMP at concentrations of 1/16, 1/8,
1/4 and 1/2 of the MIC on bacterial adhesion and BEB uptake into
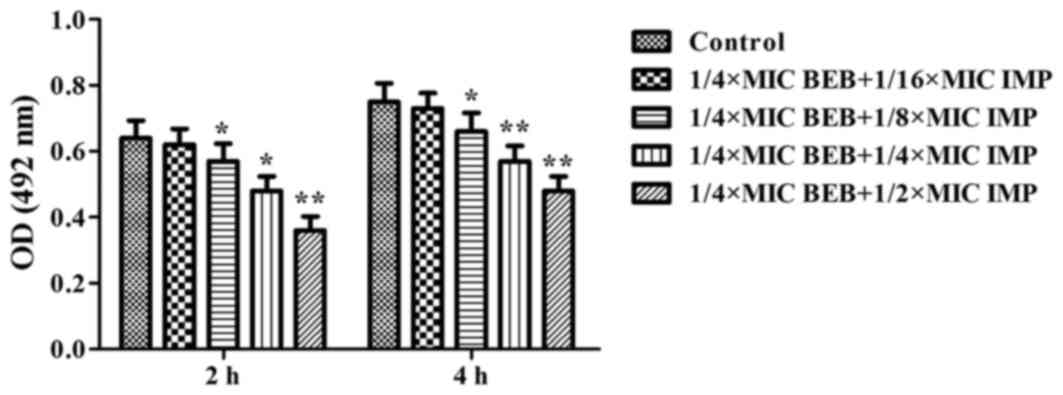
PA012 cells. It was observed that BEB (1/4 MIC) plus IMP (1/16 MIC)
did not produce any evident inhibition of the bacterial adhesion at
neither 2 nor 4 h, while application of BEB (1/4 MIC) plus IMP at
1/8, 1/4 and 1/2 of the MIC resulted in a marked decrease of
bacterial adhesion at 2 h (P<0.05, P<0.05 and P<0.01,
respectively) and at 4 h (P<0.05, P<0.01 and P<0.01,
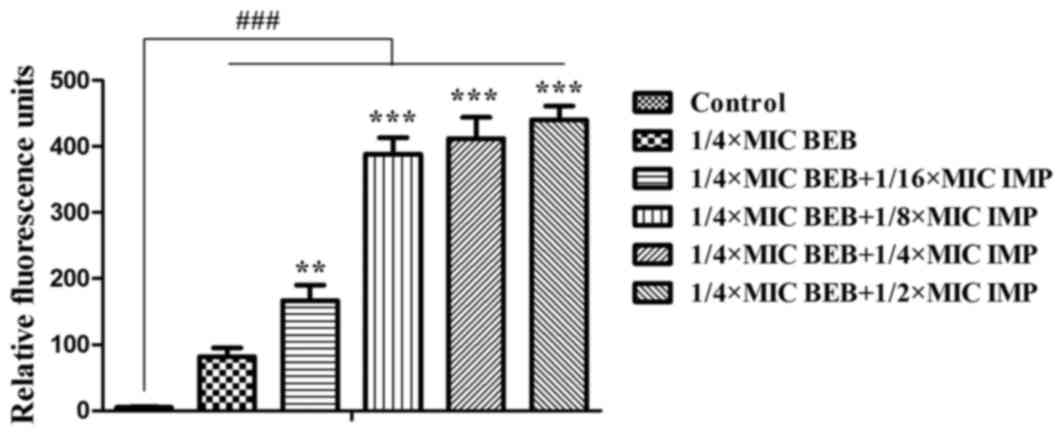
respectively; Fig. 3). BEB has
fluorescent properties that may utilized for quantifying its
intracellular accumulation. While significant fluorescence was
detected in the BEB only group compared with the control, IMP
significantly increased the fluorescence of BEB (P<0.01 or
P<0.001; Fig. 4).
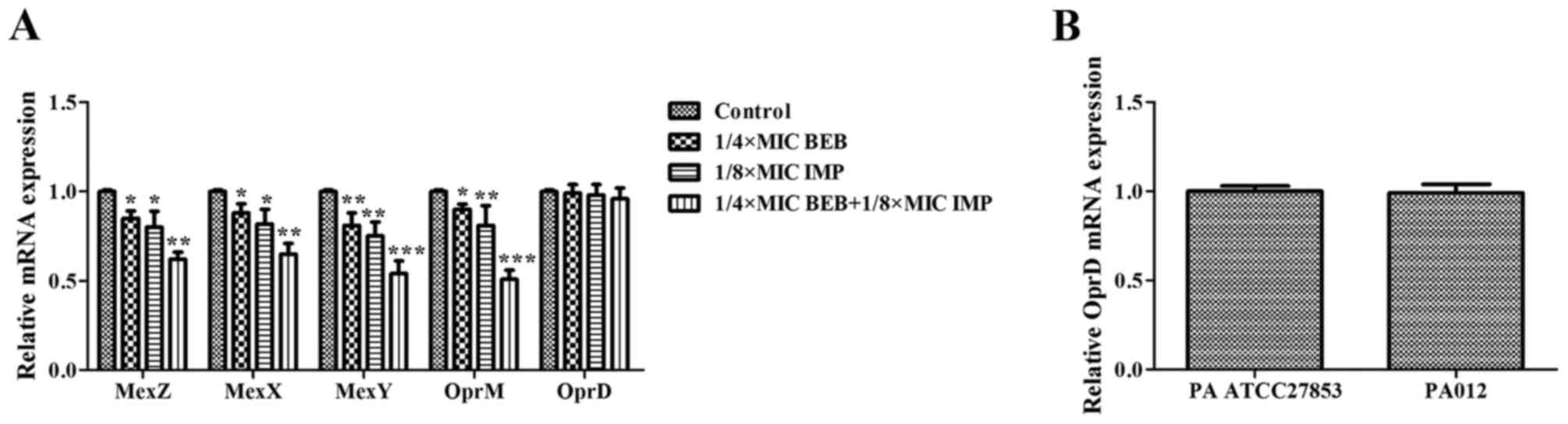
BEB and IMP synergistically inhibit
the expression of MexXY-associated genes
As demonstrated above, increasing concentrations of
IMP promoted the intracellular accumulation of BEB in PA012.
RT-qPCR was then used to analyze the antibacterial effects of BEB
and IMP alone and in combination on the genes associated with the
MexXY-OprM efflux pump in PA012. Compared with the control, BEB at
1/4 of the MIC and IMP at 1/8 of the MIC alone inhibited the
expression of MexZ (P<0.05), MexX (P<0.05), MexY (P<0.01)
and OprM (P<0.05, P<0.01). Of note, BEB plus IMP at the
abovementioned concentrations resulted in decreases of 38% for MexZ
(P<0.01), 35% for MexX (P<0.01), 46% for MexY (P<0.001)
and 49% for OprM (P<0.001). However, OprD expression was not
significantly affected by treatment with BEB and/or IMP (Fig. 5A). It was also demonstrated that the
expression of OprD in PA012 was identical to that in PA ATCC27853,
thereby excluding the possibility of OprD loss in response to the
treatments (Fig. 5B).
Discussion
Increasing carbapenem resistance in clinical P.
aeruginosa isolates caused by IMP is responsible for the
occurrence of life-threatening nosocomial diseases. The present
study demonstrated for the first time, to the best of our
knowledge, that BEB was able to enhance the antibacterial effect of
IMP in a clinical P. aeruginosa isolate by using a
susceptibility test, T-K assay, adhesion measurement and SEM
evaluation. Furthermore, it was demonstrated that the reversion of
IMP resistance by BEB may be due to inhibition of the MexXY-OprM
efflux pump.
Among the four efflux pumps associated with
carbapenem resistance, MexAB-OprM and MexXY-OprM are constitutively
expressed and their overexpression mediates intrinsic resistance in
P. aeruginosa, while MexCD-OprJ and MexEF-OprN usually
remain quiescent in wild-type P. aeruginosa and their
overexpression results in acquired multi-drug resistant phenotypes
in mutant isolates (20–22). As described previously, MexAB-OprM
and MexXY-OprM are usually overexpressed in clinical P.
aeruginosa isolates (4,5), and the latter was also demonstrated to
be a target of BEB (14). Therefore,
MexXY-OprM was extensively investigated in the present study.
The antipseudomonas activity of BEB has been
revealed to be rather weak (MIC≥512 µg/ml) (14,17). In
accordance with this, the present study also found a relatively
high MIC of BEB in P. aeruginosa. However, BEB combined with
IMP exhibited strong synergism, inferring the potential of BEB to
reverse IMP resistance or enhance the susceptibility of P.
aeruginosa to IMP. PCR analysis also demonstrated that BEP plus
IMP significantly decreased the mRNA expression of MexX, MexY and
MexZ, which was responsible for MexXY-OprM repression, while the
inhibitory effect of BEB alone on the expression of the three genes
was less pronounced. Sub-MIC IMP has been reported to possess
antipseudomonas activity (23). As
is known, the major mechanism of resistance to IMP may be
attributed to the loss or mutation of OprD, which is primarily in
charge of basic amino acid uptake and transport of carbapenems
across the outer membrane (4). Of
note, the present study did not observe any significant changes in
OprD mRNA in the clinical P. aeruginosa isolates following
incubation with BEB and/or IMP. Compared with the standard P.
aeruginosa strain PA ATCC27853, an equivalent expression of
OprD mRNA was noted in the clinical isolate PA012, indicating no
loss of the OprD gene. Due to experimental restrictions, it was not
determined whether the OprD gene was mutated in the clinical P.
aeruginosa isolate. The present results implied that BEB has a
synergistic effect with IMP by inhibiting the MexXY-OprM efflux
pump system; however, the roles of OprD in the effects of the drug
combination require further evaluation. Distinct from the
conventional concept, El Amin et al (24) reported that alterations of OprD and
efflux pumps may in part explain for carbapenem resistance patterns
in clinical P. aeruginosa isolates. The present study did
not assess all efflux pump systems, so that the possible
participation of efflux pumps other than MexXY-OprM cannot be ruled
out.
Several studies indicated that the weak
antibacterial effect of BBR was mainly due to the low accumulation
of intracellular BBR, and drugs that increased the intracellular
BBR concentration were able to promote the antibacterial effect of
BBR (25–28). The present study observed that IMP
significantly increased the intracellular BBR fluorescence. BER has
also been reported to exert its effects via targeting the
cytoplasmic membrane and intra-nuclear DNA (18,29).
Collectively, based on the present results, it may be speculated
that IMP had synergistic effects with BEB via increasing the
accumulation of BEB in the intracellular space of P.
aeruginosa cells. However, whether the ingressive BEB finally
caused cell cycle arrest and DNA damage similar to its effects in
Candida albicans still requires to be elucidated (13).
In conclusion, the present study suggested that BEB
has a synergistic effect with IMP to re-sensitize IMP-resistant
P. aeruginosa via inhibiting the MexXY-OprM efflux pump
system. In future studies, more clinical P. aeruginosa
isolates will be collected to confirm the synergistic activity of
BEB and IMP, and simultaneously, nucleotide sequences of OprD
should be analyzed. The antibacterial effects of BEB and/or IMP
will also be evaluated in in vivo studies and relevant
experiments are underway.
References
|
1
|
Morita Y, Tomida J and Kawamura Y:
Responses of Pseudomonas aeruginosa to antimicrobials. Front
Microbiol. 4:4222014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li XZ, Plésiat P and Nikaido H: The
challenge of efflux-mediated antibiotic resistance in Gram-negative
bacteria. Clin Microbiol Rev. 28:337–418. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kiser TH, Obritsch MD, Jung R, MacLaren R
and Fish DN: Efflux pump contribution to multidrug resistance in
clinical isolates of Pseudomonas aeruginosa. Pharmacotherapy.
30:632–638. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Livermore DM: Of Pseudomonas, porins,
pumps and carbapenems. J Antimicrob Chemother. 47:247–250. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Okamoto K, Gotoh N and Nishino T:
Extrusion of penem antibiotics by multicomponent efflux systems
MexAB-OprM, MexCD-OprJ and MexXY-OprM of Pseudomonas aeruginosa.
Antimicrob Agents Chemother. 46:2696–2699. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xavier DE, Picão RC, Girardello R,
Fehlberg LC and Gales AC: Efflux pumps expression and its
association with porin down-regulation and beta-lactamase
production among Pseudomonas aeruginosa causing bloodstream
infections in Brazil. BMC Microbiol. 10:2172010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Khuntayaporn P, Montakantikul P,
Santanirand P, Kiratisin P and Chomnawang MT: Molecular
investigation of carbapenem resistance among multidrug-resistant
Pseudomonas aeruginosa isolated clinically in Thailand. Microbiol
Immunol. 57:170–178. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
American Thoracic Society, ; Infectious
Diseases Society of America, . Guidelines for the management of
adults with hospital-acquired, ventilator-associated and
healthcare-associated pneumonia. Am J Respir Crit Care Med.
171:388–416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guo N, Zhao X, Li W, Shi C, Meng R, Liu Z
and Yu L: The synergy of berberine chloride and totarol against
Staphylococcus aureus grown in planktonic and biofilm cultures. J
Med Microbiol. 64:891–900. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dziedzic A, Wojtyczka RD and Kubina R:
Inhibition of oral Streptococci growth induced by the complementary
action of berberine chloride and antibacterial compounds.
Molecules. 20:13705–13724. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gokgoz N Budeyri, Avci FG, Yoneten KK,
Alaybeyoglu B, Ozkirimli E, Sayar NA, Kazan D and Akbulut B
Sariyar: Response of Escherichia coli to prolonged berberine
exposure. Microb Drug Resist. 23:531–544. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shao J, Shi G, Wang T, Wu D and Wang C:
Antiproliferation of berberine in combination with fluconazole from
the perspectives of reactive oxygen species, ergosterol and drug
efflux in a fluconazole-resistant Candida tropicalis isolate. Front
Microbiol. 7:15162016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li D, Xu Y, Zhang D, Quan H, Mylonakis E,
Hu D, Li M, Zhao L, Zhu L, Wang Y and Jiang YY: Fluconazole assists
berberine to kill fluconazole-resistant Candida albicans.
Antimicrob Agents Chemother. 57:6016–6027. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Morita Y, Nakashima K, Nishino K, Kotani
K, Tomida J, Inoue M and Kawamura Y: Berberine is a novel type
efflux inhibitor which attenuates the MexXY-mediated aminoglycoside
resistance in Pseudomonas aeruginosa. Front Microbiol. 7:12232016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Clinical and Laboratory Standards
Institute (CLSI), . Methods for dilution antimicrobial
susceptibility tests for bacteria that grow aerobically; Approved
StandardCLSI Document M07-A10. 10th. CLSI; Wayne, PA, USA: 2015
|
|
16
|
Odds F: Synergy, antagonism and what the
chequerboard puts between them. J Antimicrob Chemother. 52:12003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jayaraman P, Sakharkar MK, Lim CS, Tang TH
and Sakharkar KR: Activity and interactions of antibiotic and
phytochemical combinations against Pseudomonas aeruginosa in vitro.
Int J Biol Sci. 6:556–568. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Yao X, Zhu Z, Tang T, Dai K,
Sadovskaya I, Flahaut S and Jabbouri S: Effect of berberine on
Staphylococcus epidermidis biofilm formation. Int J Antimicrob
Agents. 34:60–66. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Poole K and Srikumar R: Multidrug efflux
in Pseudomonas aeruginosa: Components, mechanisms and clinical
significance. Curr Top Med Chem. 1:59–71. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sobel ML, Hocquet D, Cao L, Plesiat P and
Poole K: Mutations in PA3574 (nalD) lead to increased MexAB-OprM
expression and multidrug resistance in laboratory and clinical
isolates of Pseudomonas aeruginosa. Antimicrob Agents Chemother.
49:1782–1786. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao L, Srikumar R and Poole K: MexAB-OprM
hyperexpression in NalC-type multidrug-resistant Pseudomonas
aeruginosa: Identification and characterization of the nalC gene
encoding a repressor of PA3720-PA3719. Mol Microbiol. 53:1423–1436.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fonseca AP and Sousa JC: Effect of
antibiotic-induced morphological changes on surface properties,
motility and adhesion of nosocomial Pseudomonas aeruginosa strains
under different physiological states. J Appl Microbiol.
103:1828–1837. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
El Amin N, Giske CG, Jalal S, Keijser B,
Kronvall G and Wretlind B: Carbapenem resistance mechanisms in
Pseudomonas aeruginosa: Alterations of porin OprD and efflux
proteins do not fully explain resistance patterns observed in
clinical isolates. APMIS. 113:187–196. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stermitz FR, Lorenz P, Tawara JN, Zenewicz
LA and Lewis K: Synergy in a medicinal plant: Antimicrobial action
of berberine potentiated by 5′-methoxyhydnocarpin, a multidrug pump
inhibitor. Proc Natl Acad Sci USA. 97:pp. 1433–1437. 2000;
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ettefagh KA, Burns JT, Junio HA, Kaatz GW
and Cech NB: Goldenseal (Hydrastis canadensis L.) Extracts
synergistically enhance the antibacterial activity of berberine via
efflux pump inhibition. Planta Med. 77:835–840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ball AR, Casadei G, Samosorn S, Bremner
JB, Ausubel FM, Moy TI and Lewis K: Conjugating berberine to a
multidrug efflux pump inhibitor creates an effective antimicrobial.
ACS Chem Biol. 1:594–600. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Samosorn S, Tanwirat B, Muhamad N, Casadei
G, Tomkiewicz D, Lewis K, Suksamrarn A, Prammananan T, Gornall KC,
Beck JL and Bremner JB: Antibacterial activity of berberine-NorA
pump inhibitor hybrids with a methylene ether linking group. Bioorg
Med Chem. 17:3866–3872. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Severina II, Muntyan MS, Lewis K and
Skulachev VP: Transfer of cationic antibacterial agents berberine,
palmatine and benzalkonium through bimolecular planar phospholipid
film and Staphylococcus aureus membrane. IUBMB Life. 52:321–324.
2001. View Article : Google Scholar : PubMed/NCBI
|