Introduction
Obesity is associated with multiple comorbidities,
including hypertension, sleep apnea, cancers and cardiovascular
disease (1,2). In addition, obesity confers an
increased risk of developing a number of metabolic diseases,
including dyslipidemia, type 2 diabetes and non-alcoholic fatty
liver disease (NAFLD) (3,4). Accumulating evidence has indicated that
elevated fatty acid synthesis and/or impaired fatty acid oxidation
have important roles in the development of obesity and associated
metabolic diseases (5,6). Hence, the modulation of fatty acid
metabolism may be a potential therapeutic target for
obesity-associated metabolic diseases.
Acetyl-CoA carboxylase (ACC) is a biotin-dependent
heterodimeric enzyme responsible for the catalyzation of acetyl-CoA
to form malonyl-CoA, hence regulating fatty acid metabolism and
energy homeostasis (7). Two
tissue-specific isozymes of ACC have been identified, including
ACC1 and ACC2, which have divergent roles and distinct cellular
distributions (4). ACC1, mainly
expressed in the liver and adipose tissue, catalyzes the
ATP-dependent carboxylation of acetyl-CoA to form malonyl-CoA,
which functions as a building block to extend the chain length of
fatty acids in liver and adipose (8,9). By
contrast, ACC2, abundant in heart and muscle tissues and tethered
to the mitochondrial outer membrane, functions as a suppressor of
mitochondrial fatty acid oxidation by inhibiting carnitine
palmitoyl transferase 1 (CPT-1) (10,11).
Located in the mitochondrial outer membrane, CPT-1 assists the
transport of long-chain acyl-CoA into the mitochondrial membrane
for subsequent β-oxidation by converting this to acylcarnitines
(12). Thus, ACC has an important
role in regulating the fatty acid metabolism and energy homeostasis
(13). Other studies have
demonstrated that the genetic knockout of ACC2 lowers malonyl-CoA
levels and enhances fatty acid oxidation in rodent models of
dyslipidemia (14,15). In addition, pharmacological
inhibition of ACC activity benefits fatty acid metabolism and
improves glucose homeostasis in db/db mice (16).
Recently, a novel ACC inhibitor targeting ACC
carboxyl transfer domain, PP-7a, was developed by our group, which
exhibited a potent inhibitory effect on ACC activity in in
vitro studies (17,18). The present study aimed to evaluate
the pharmacological effects of ACC inhibition by PP-7a on metabolic
disorders in mice fed a high-fat diet (HFD).
Materials and methods
Animals and chemical
administration
A total of 35 male C57BL/6J mice (age, 6-7 weeks;
weight, 18-19 g), were obtained from the Branch of National Breeder
Center of Rodents (Shanghai, China). They were housed in an
environment with controlled temperature (23±1˚C) and humidity
(55±5%), and a 12-h light/dark cycle with free access to water and
food. All animal experiments were approved by the Animal Ethics
Committee of Xuzhou Medical University (Xuzhou, China) and
performed in accordance with the National Institutes of Health
Guidelines for the Care and Use of Laboratory Animals. Following
one week of acclimation, the mice were randomly divided into a
control group and an HFD group. According to the protocol of a
previous study (19), the mice in
the control group (n=6) were fed ad libitum a normal chow
diet (purchased from the Animal Experimental Center of Xuzhou
Medical University) composed with 19% kcal protein, 68% kcal
carbohydrates and 13% kcal fat, while the mice of HFD group were
fed a HFD consisting of 15% kcal protein, 43% kcal carbohydrates
and 42% kcal fat ad libitum for 16 weeks to develop obesity.
The most important parameter of the HFD mouse model is the
increased body weight and adipose tissue, as compared with the mice
fed a normal chow diet.
Following 16 weeks on the HFD, the mice were
randomly divided into five groups, including the HFD (n=6),
HFD+PP-7a (15 mg/kg, n=5), HFD+PP-7a (45 mg/kg, n=6), HFD+PP-7a (75
mg/kg, n=6) and HFD+CP-640186 groups (75 mg/kg, n=6). Compound
PP-7a was prepared as detailed in a recent study by our group
(17). CP-640186, a known potent ACC
inhibitor used as the positive control, was supplied by Selleck
Chemicals Co. Ltd (20). PP-7a and
CP-640186 have similar chemical structures and exhibited a
comparable inhibitory effect on ACC1/2 activity and comparable
in vitro cytotoxic activities (17). The doses of PP-7a administered to HFD
mice were selected based on in vivo studies investigating
the pharmacological effects of CP-640186 published previously
(20). PP-7a and CP-640186 were
dissolved in 0.5% carboxymethylcellulose. PP-7a was administered at
the corresponding doses by gavage once daily for 4 weeks. The mice
from the HFD+CP-640186 group received CP-640186 orally each day for
4 weeks. In parallel, the mice in the control group and HFD group
were administered 0.5% carboxymethylcellulose solution. The body
weight was measured once a week. After 4 weeks, the mice were
subjected to a glucose tolerance test and MRI. Under anesthesia
with chloral hydrate (10%, 500 mg/kg, i.p.), the animals were
sacrificed by cervical dislocation and none of them exhibited signs
of peritonitis following the administration of 10% chloral hydrate,
as in previous studies (21,22). All of the experimental animals were
anesthetized with 10% chloral hydrate prior to blood sampling and
were not allowed to survive after blood sampling. Subsequently, the
heart, liver and abdominal adipose tissues were harvested for
biochemical and histological analysis.
Glucose tolerance test
To evaluate the effects of ACC inhibition on
impaired glucose tolerance in obese mice, glucose tolerance tests
were performed after 4 weeks of drug administration. In brief,
following a 16-h fast, the mice were intragastrically administered
glucose (2 g/kg). Blood samples were obtained via the tail vein at
0, 30, 60 and 120 min after glucose loading and the blood glucose
concentrations were measured using the Bayer Contour Glucose Meter
(Bayer AG).
Analysis of serum triglyceride (TG),
total cholesterol (TC) and free fatty acid (FFA) levels
The concentrations of TG, TC and FFA in serum
collected from retro-orbital exsanguination of the mice were
measured with respective assay kits (cat. no. SNM227, SNM226 and
SK125-1, respectively; Beijing Baiaolaibo Technology Co., Ltd.).
Blood samples were collected and centrifuged at 1,200 x g for 10
min at 4˚C to collect serum. The serum samples were then added to
designated ELISA plates and incubated for 30 min at 37˚C. After
discarding the liquid and drying, each well was washed with wash
buffer 5 times. Following the addition of horseradish
peroxidase-conjugated reagent, the ELISA plate incubated for 30 min
at 37˚C. After washing with wash buffer, chromogen solution was
added to each well for chromogenic reaction for 15 min at 37˚C and
placed in the dark. Following the addition of stop solution, the
absorbance was then measured at 450 nm using a microplate reader
(Thermo Fisher Scientific, Inc.).
Measurement of malonyl-CoA levels
The livers (ACC1-rich tissue) and hearts (ACC2-rich
tissue) were harvested and stored at -80˚C for the detection of
malonyl-CoA levels. The liver and heart tissues were homogenized
with PBS buffer and then centrifuged at 12,000 x g at 4˚C for 15
min. Subsequently, the supernatant was subjected to the detection
of malonyl-CoA with a commercial ELISA kit (cat. no. ARB14412;
Beijing Baiaolaibo Technology Co., Ltd.) according to the
manufacturer's protocol.
Histological analysis
Partial liver and abdominal adipose tissues from
individual mice were post-fixed in 4% paraformaldehyde at 4˚C for
48 h, followed by dehydration using graded ethanol and embedding in
paraffin (23). The tissues were cut
to a thickness of 5 µm. Subsequently, the tissue sections were
stained with hematoxylin for 10 min and eosin for 2 min at room
temperature with Hematoxylin-Eosin staining kit (cat. no. D006-1-1;
Nanjing Jiancheng Bio-Engineering Institute Co., Ltd), followed by
histological analysis under a light microscope (magnification,
x400).
Assessment of adipose mass by MRI
To assess the changes in the adipose tissue
compartment, mice were subjected to an MRI scan under anesthesia
using a Philips Achieva 1.5 Tesla MRI (Philips Medical Systems
B.V.). Images were obtained on a ChemiDoc Touch Imaging System and
densitometry was performed using Image Lab Software version 5.2.1
(both Bio-Rad Laboratories, Inc.).
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Statistical significance was assessed using Student's
t-test or one-way analysis of variance followed by Tukey's test
using GraphPad version 6.0 software (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
PP-7a suppresses the gain of body
weight in mice fed an HFD
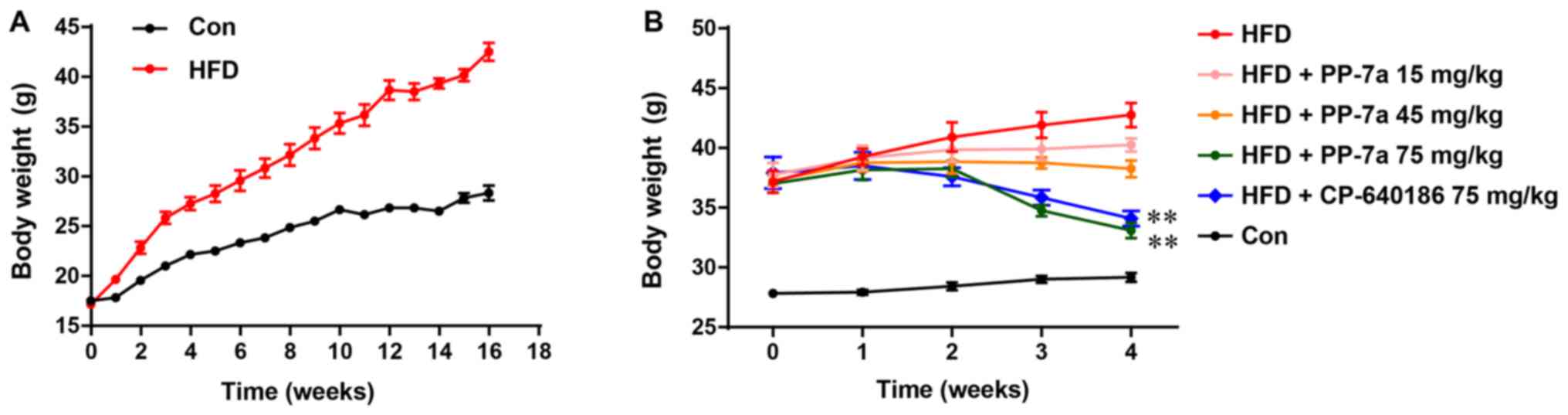
As depicted in Fig.
1A, the body weight of the control mice fed a chow diet or HFD
gradually increased during the period of mouse model generation.
After 16 weeks on the HFD, the average body weight of mice
receiving the HFD increased >10 g compared with that of the mice
fed the normal chow diet (Fig. 1A)
and the changes in body weight were consistent with those reported
in a previous study (19). In
addition, after 4 weeks of treatment with PP-7a or CP-640186, the
weight of the mice in the HFD+PP-7a (75 mg/kg) group and
HFD+CP-640186 (75 mg/kg) group was significantly decreased, while
the weight of the mice in the HFD group without treatment
continually increased. Compared with that of the mice in the HFD
group, the body weight gain was also suppressed by chronic
treatment with PP-7 at doses of 15 and 45 mg/kg (Fig. 1B). In addition, no toxicity was
observed in HFD mice administered PP-7a or CP-640186. In this
experiment, visual observations were made for any lethal reactions
in HFD mice administered with PP-7a or CP-640186. No such adverse
reactions, including a coat with an unhealthy appearance, erythema
on the skin surface, appearance of depression or mortality were
observed in the HFD mice that had been administered drugs.
Treatment with PP-7a improves glucose
tolerance
Glucose tolerance tests were performed to examine
the effects of PP-7a on glucose tolerance in mice fed the HFD.
Following gavage administration of glucose, all mice in the
experimental group displayed elevated blood glucose levels within
120 min. The glucose levels in mice of the HFD group were higher
compared with those of the mice in the control group. Chronic
treatment with PP-7a exerted a suppressive effect on blood glucose
elevation (Fig. 2A and B). PP-7a treatment at 75 mg/kg produced a
glucose-lowering effect comparable to that of the positive control
(Fig. 2A and B).
PP-7a suppresses the increase in serum
TG, TC and FFA levels in obese mice fed the HFD
Usually, increased serum TG, TC and FFA levels are
associated with obesity (24). Thus,
to determine the effect of PP-7a on lipid metabolism in the mice
fed the HFD, the TG, TC and FFA levels in serum were detected using
ELISA kits. As expected, the levels of TG, TC and FFA in the mice
fed the HFD were significantly higher than those of the control
mice fed a normal chow diet. Chronic administration of PP-7a at the
dose of 75 mg/kg significantly suppressed the increases in the
levels of TG and TC (Fig. 3A and
B). In addition, PP-7a suppressed
the increase in the serum FFA level in a dose-dependent manner
(Fig. 3C). There was no significant
difference in the levels of serum TG, TC and FFA between HFD+PP-7a
(75 mg/kg) group and the positive control group (Fig. 3A-C).
PP-7a reduces the malonyl-CoA levels
in liver and heart tissues of mice fed the HFD
As a key regulator of fatty acid metabolism,
malonyl-CoA is synthesized under the catalysis of ACC (25). Hence, the effects of PP-7a on the
level of malonyl-CoA in liver and heart tissues were assessed in
mice fed the HFD (26). As expected,
chronic administration of PP-7a significantly reduced malonyl-CoA
levels in liver and heart tissues of mice fed the HFD (Fig. 4A and B). There was no significant difference in
the malonyl-CoA levels of liver and heart tissues between HFD+PP-7a
(75 mg/kg) group and the positive control group (Fig. 4A and B).
Hepatic lipogenesis and abdominal
adipose accumulation are alleviated by PP-7a
As presented in Fig.
5, the HFD increased liver fat accumulation, which was
alleviated by chronic treatment with PP-7a or CP-640186 (Fig. 5A). In addition, histological
examination indicated larger adipocytes in the abdominal
subcutaneous adipose tissue of the mice fed the HFD compared with
the mice fed the chow diet. Compared with the mice fed the HFD,
chronic treatment with PP-7a reduced the size of adipocytes
(Fig. 5B).
Treatment with PP-7a suppresses fat
mass accretion
To determine whether the reduction in body weight
was attributed to the decrease of adiposity, quantitative analysis
of the abdominal adipose tissue of mice was performed by MRI
imaging. As observed from the transverse cross-sections and
sagittal sections of MRI images presented in Fig. 6A and B, the abdominal adipose volume in mice from
the HFD group was larger compared with that in the control mice.
Quantitative evaluation indicated that treatment with PP-7a at the
dose of 75 mg/kg significantly suppressed the increase of abdominal
adipose in mice fed the HFD (Fig. 6C
and D). No significant difference in
the abdominal adipose was observed among HFD+PP-7a (75 mg/kg)
group, the positive control group, and the control group (Fig. 6C and D).
Discussion
ACC functions as a critical regulator of fatty acid
biosynthesis and oxidation (27,28).
Inhibition of ACC has been reported to exert a suppressive effect
on fatty acid biosynthesis, while it promotes fatty acid oxidation
(25,29,30). Two
isoforms of acetyl-CoA carboxylases, ACC1 and ACC2, exhibit
divergent expression profiles and biological functions (31). In a previous study by our group, a
novel ACC inhibitor, PP-7a, was developed, which is able to target
the carboxyl transfer domain of ACC (17). The present study investigated the
pharmacological action of PP-7a to reduce HFD-induced metabolic
dysregulation in mice. There is evidence to suggest that HFD is a
major contributor to obesity and associated metabolic syndrome
(32). The results of the present
study demonstrated that a long-term HFD induced obesity and
symptoms of metabolic syndrome, including glucose intolerance, fat
accumulation and hyperlipidemia, in mice. These results are
consistent with those of previous studies (33,34).
However, the weight gain induced by the HFD was suppressed by
chronic treatment with PP-7a in a dose-dependent manner at the
doses of 15, 45 and 75 mg/kg. Furthermore, following treatment with
PP-7a at 75 mg/kg for 4 weeks, the body weight of the HFD-fed mice
was significantly decreased. Abdominal MRI further revealed that
the mice fed the HFD treated with PP-7a displayed less abdominal
fat accumulation compared with the mice treated with the vehicle.
These results suggested that decreased fat storage contributed to
the weight loss induced by PP-7a.
In addition, the regulation of fatty acid metabolism
affects body weight gain and fat accumulation (7,35). The
present study indicated that ACC inhibition by PP-7a suppressed
HFD-induced fat accumulation. Malonyl-CoA, generating ACC, mediates
the regulatory effect of ACC on fatty acid metabolism (36,37).
Hence, the malonyl-CoA levels in the liver and heart were detected
to confirm the ACC inhibition in the present study. As expected, it
was indicated that chronic treatment with PP-7a reduced the
malonyl-CoA levels in liver and heart tissues, suggesting a
simultaneous inhibitory effect on ACC1 and ACC2 activities.
Previous studies have demonstrated that ACC2 deletion enhances
ex vivo skeletal muscle fatty acid oxidation and inhibition
of ACC2 promotes skeletal muscle fatty acid oxidation in db/db mice
accompanied by a decrease in the malonyl-CoA level (16,30).
However, similar effects were not observed in another study
(38). Although the reasons for this
discrepancy remains elusive, it may be due to the compensatory
increases in fatty acid synthesis mitigated by the ACC2
deletion-mediated efficacy (39).
The results of the present study demonstrate that simultaneous
inhibition of ACC1 and ACC2 by PP-7a resulted in a reduction of
malonyl-CoA levels, fat accumulation and weight gain induced by the
HFD. In Fig. 7, the hypothesis that
PP-7a exerts the beneficial effects on metabolic dysregulation
through the inhibition of ACC was rasied, thereby promoting the
fatty acid oxidation and inhibiting fatty acid synthesis. The
potential mechanisms underlying the pharmacological effects of
PP-7a warrant further investigation in the future.
Obesity is associated with hyperlipidemia and
glucose intolerance (40,41). In addition, the modulation of fatty
acid metabolism affects the plasma lipid levels (42). Hence, in the present study, the
effects of chronic treatment with PP-7a on hyperlipidemia in mice
with HFD-induced obesity were assessed. As expected,
pharmacological inhibition of ACC reduced the levels of TG, TC and
FFA in the serum of mice fed the HFD. These results demonstrated
that the targeting of ACC by PP-7a has the potential to suppress
the increased lipid levels caused by the HFD. Excessive TG levels
are the cause of a fatty liver, which contributes to dysregulated
glucose metabolism (43). The
results of the present study further revealed that PP-7a treatment
decreased lipid deposition in the livers of mice fed the HFD. PP-7a
administration improved glucose tolerance in HFD-fed mice,
suggesting an enhanced effect on glucose homeostasis. Thus, these
data support the hypothesis that inhibition of ACC activity
suppresses HFD-induced hyperlipidemia and dysregulates glucose
homeostasis in mice. However, a limitation of the present study was
the lack of ACC expression analysis. The assessment of ACC
expression may be helpful to clarify the underlying molecular
mechanisms of the pharmacological effect of PP-7a and the effect of
PP-7a on the expression of ACC in liver and heart tissues should be
examined in a further study.
It should be noted that increased fatty acid
oxidation is not always desirable and innocent; hence, the
potential adverse effects of ACC inhibition should be considered.
Fatty acid synthesis is necessary for T-cell clonal expansion in
the immune system and fatty acid oxidation regulates the
differentiation of regulatory T cells, a major regulatory component
of the immune system (44,45). Loss of ACC impairs T-cell homeostasis
in the periphery and homeostatic proliferation (46,47). In
addition, high glucose triggers endothelial injury through
increased fatty acid oxidation (48). Other studies have demonstrated that
increased free fatty acid levels mediate endothelial dysfunction in
subjects with obesity and type 2 diabetes (49-51).
Therefore, the present study has a limitation due to the lack of
in vivo toxicity tests on PP-7a. More attention should be
paid to the side effects of ACC inhibition by ACC inhibitors in
future studies. More importantly, the pharmacokinetics of PP-7a
should be further explored.
In conclusion, the present study demonstrated that
the pharmacological inhibition of ACC by PP-7a, a novel acetyl-CoA
carboxylase inhibitor, exerted a beneficial effect on metabolic
dysregulation induced by an HFD in mice. In addition, the present
study provided further evidence that ACC represents a therapeutic
target for the treatment of obesity-associated metabolic
diseases.
Acknowledgements
Not applicable.
Funding
This study was funded by grants from China
Postdoctoral Science Foundation-funded projects (grant nos.
2017M611916 and 2019M661943), the Natural Science Foundation of
Jiangsu Province (grant nos. BK20171184 and BK20170258), Jiangsu
Planned Projects for Postdoctoral Research Funds (grant no.
1701132C) and National Natural Science Foundation of China (grant
no. 81703493).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
THH and TYL designed the current study and wrote the
first draft of the manuscript. TYL, LSG and SRY performed the
experiments. LSG, SRY and THH collected, analyzed and interpreted
the data. TYL, LSG prepared the figures and participated in
substantive revisions of the important content of the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in compliance
with the Chinese legislation on the use and care of laboratory
animals and the study was approved by the Ethics Committee on
Animal Care and Use of Xuzhou University Medical College (Xuzhou,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Basen-Engquist K and Chang M: Obesity and
cancer risk: Recent review and evidence. Curr Oncol Rep. 13:71–76.
2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Gou L, Zhao L, Song W, Wang L, Liu J,
Zhang H, Huang Y, Lau CW, Yao X, Tian XY, et al: Inhibition of
miR-92a suppresses oxidative stress and improves endothelial
function by upregulating heme oxygenase-1 in db/db mice. Antioxid
Redox Signal. 28:358–370. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Mohammed MS, Sendra S, Lloret J and Bosch
I: Systems and WBANs for controlling obesity. J Healthc Eng.
2018(1564748)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Tong L and Harwood HJ Jr: Acetyl-coenzyme
A carboxylases: Versatile targets for drug discovery. J Cell
Biochem. 99:1476–1488. 2006.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Harriman G, Greenwood J, Bhat S, Huang X,
Wang R, Paul D, Tong L, Saha AK, Westlin WF, Kapeller R and Harwood
HJ Jr: Acetyl-CoA carboxylase inhibition by ND-630 reduces hepatic
steatosis, improves insulin sensitivity, and modulates dyslipidemia
in rats. Proc Natl Acad Sci USA. 113:E1796–E1805. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kim CW, Addy C, Kusunoki J, Anderson NN,
Deja S, Fu X, Burgess SC, Li C, Ruddy M, Chakravarthy M, et al:
Acetyl CoA carboxylase inhibition reduces Hepatic steatosis but
elevates plasma triglycerides in mice and humans: A bedside to
bench investigation. Cell Metabol. 26(576)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Wakil SJ and Abu-Elheiga LA: Fatty acid
metabolism: Target for metabolic syndrome. J Lipid Res. 50
(Suppl):S138–S143. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Harwood HJ Jr: Treating the metabolic
syndrome: Acetyl-CoA carboxylase inhibition. Expert Opin Ther
Targets. 9:267–281. 2005.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Goedeke L, Bates J, Vatner DF, Perry RJ,
Wang T, Ramirez R, Li L, Ellis MW, Zhang D, Wong KE, et al:
Acetyl-CoA carboxylase inhibition reverses NAFLD and hepatic
insulin resistance but promotes hypertriglyceridemia in rodents.
Hepatology. 68:2197–2211. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Abu-Elheiga L, Brinkley WR, Zhong L,
Chirala SS, Woldegiorgis G and Wakil SJ: The subcellular
localization of acetyl-CoA carboxylase 2. Proc Natl Acad Sci USA.
97:1444–1449. 2000.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Marin-Garcia J and Goldenthal MJ: Fatty
acid metabolism in cardiac failure: Biochemical, genetic and
cellular analysis. Cardiovasc Res. 54:516–527. 2002.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Qu Q, Zeng F, Liu X, Wang QJ and Deng F:
Fatty acid oxidation and carnitine palmitoyltransferase I: Emerging
therapeutic targets in cancer. Cell Death Dis.
7(e2226)2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ussher JR and Lopaschuk GD: The malonyl
CoA axis as a potential target for treating ischaemic heart
disease. Cardiovasc Res. 79:259–268. 2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Baig NA, Herrine SK and Rubin R: Liver
disease and diabetes mellitus. Clin Lab Med. 21:193–207.
2001.PubMed/NCBI
|
|
15
|
Abu-Elheiga L, Matzuk MM, Abo-Hashema KA
and Wakil SJ: Continuous fatty acid oxidation and reduced fat
storage in mice lacking acetyl-CoA carboxylase 2. Science.
291:2613–2616. 2001.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Glund S, Schoelch C, Thomas L, Niessen HG,
Stiller D, Roth GJ and Neubauer H: Inhibition of acetyl-CoA
carboxylase 2 enhances skeletal muscle fatty acid oxidation and
improves whole-body glucose homeostasis in db/db mice.
Diabetologia. 55:2044–2053. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Huang T, Sun J, Wang Q, Gao J and Liu Y:
Synthesis, biological evaluation and molecular docking studies of
piperidinylpiperidines and spirochromanones Possessing quinoline
moieties as Acetyl-CoA carboxylase inhibitors. Molecules.
20:16221–16234. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Huang TH, Sun J, Xu M and Liu Y:
Biological evaluation studies of Acetyl-Coa carboxylase inhibitor
in high fat diet induced obese mice. Basic Clin Pharmacol.
119:44–45. 2016.
|
|
19
|
Liu Y, Fu X, Lan N, Li S, Zhang J, Wang S,
Li C, Shang Y, Huang T and Zhang L: Luteolin protects against high
fat diet-induced cognitive deficits in obesity mice. Behav Brain
Res. 267:178–188. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Harwood HJ Jr, Petras SF, Shelly LD,
Zaccaro LM, Perry DA, Makowski MR, Hargrove DM, Martin KA, Tracey
WR, Chapman JG, et al: Isozyme-nonselective N-substituted
bipiperidylcarboxamide acetyl-CoA carboxylase inhibitors reduce
tissue malonyl-CoA concentrations, inhibit fatty acid synthesis,
and increase fatty acid oxidation in cultured cells and in
experimental animals. J Biol Chem. 278:37099–37111. 2003.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Matsushita H, Johnston MV, Lange MS and
Wilson MA: Protective effect of erythropoietin in neonatal hypoxic
ischemia in mice. Neuroreport. 14:1757–1761. 2003.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen B, Ma Y, Xue X, Wei J, Hu G and Lin
Y: Tetramethylpyrazine reduces inflammation in the livers of mice
fed a high fat diet. Mol Med Rep. 19:2561–2568. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Feng Y, Yu YH, Wang ST, Ren J, Camer D,
Hua YZ, Zhang Q, Huang J, Xue DL, Zhang XF, et al: Chlorogenic acid
protects D-galactose-induced liver and kidney injury via
antioxidation and anti-inflammation effects in mice. Pharm Biol.
54:1027–1034. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lu JC, Jing J, Yao Q, Fan K, Wang GH, Feng
RX, Liang YJ, Chen L, Ge YF and Yao B: Relationship between lipids
levels of serum and seminal plasma and semen parameters in 631
Chinese subfertile men. PLoS One. 11(e0146304)2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cuthbert KD and Dyck JR: Malonyl-CoA
decarboxylase is a major regulator of myocardial fatty acid
oxidation. Curr Hypertens Rep. 7:407–411. 2005.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Awan MM and Saggerson ED: Malonyl-CoA
metabolism in cardiac myocytes and its relevance to the control of
fatty acid oxidation. Biochem J. 295:61–66. 1993.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Munday MR: Regulation of mammalian
acetyl-CoA carboxylase. Biochem Soc Trans. 30:1059–1064.
2002.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Saddik M, Gamble J, Witters LA and
Lopaschuk GD: Acetyl-CoA carboxylase regulation of fatty acid
oxidation in the heart. J Biol Chem. 268:25836–25845.
1993.PubMed/NCBI
|
|
29
|
Eleftheriadis T, Pissas G, Sounidaki M,
Tsogka K, Antoniadis N, Antoniadi G, Liakopoulos V and Stefanidis
I: Indoleamine 2,3-dioxygenase, by degrading L-tryptophan, enhances
carnitine palmitoyltransferase I activity and fatty acid oxidation,
and exerts fatty acid-dependent effects in human alloreactive CD4+
T-cells. Int J Mol Med. 38:1605–1613. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Choi CS, Savage DB, Abu-Elheiga L, Liu ZX,
Kim S, Kulkarni A, Distefano A, Hwang YJ, Reznick RM, Codella R, et
al: Continuous fat oxidation in acetyl-CoA carboxylase 2 knockout
mice increases total energy expenditure, reduces fat mass, and
improves insulin sensitivity. Proc Natl Acad Sci USA.
104:16480–16485. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Strable MS and Ntambi JM: Genetic control
of de novo lipogenesis: Role in diet-induced obesity. Crit Rev
Biochem Mol Biol. 45:199–214. 2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Golay A and Bobbioni E: The role of
dietary fat in obesity. Int J Obes Relat Metab Disord. 21 (Suppl
3):S2–S11. 1997.PubMed/NCBI
|
|
33
|
Heydemann A: An overview of murine high
fat diet as a model for type 2 diabetes mellitus. J Diabetes Res.
2016(2902351)2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Hariri N and Thibault L: High-fat
diet-induced obesity in animal models. Nutri Res Rev. 23:270–299.
2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ronnett GV, Kleman AM, Kim EK, Landree LE
and Tu Y: Fatty acid metabolism, the central nervous system, and
feeding. Obesity (Silver Spring). 14 (Suppl 5):S201–S207.
2006.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Brownsey RW, Boone AN, Elliott JE, Kulpa
JE and Lee WM: Regulation of acetyl-CoA carboxylase. Biochem Soc
Trans. 34:223–227. 2006.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Folmes CD and Lopaschuk GD: Role of
malonyl-CoA in heart disease and the hypothalamic control of
obesity. Cardiovasc Res. 73:278–287. 2007.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Olson DP, Pulinilkunnil T, Cline GW,
Shulman GI and Lowell BB: Gene knockout of Acc2 has little effect
on body weight, fat mass, or food intake. Proc Natl Acad Sci USA.
107:7598–7603. 2010.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Hoehn KL, Turner N, Swarbrick MM, Wilks D,
Preston E, Phua Y, Joshi H, Furler SM, Larance M, Hegarty BD, et
al: Acute or chronic upregulation of mitochondrial fatty acid
oxidation has no net effect on whole-body energy expenditure or
adiposity. Cell Metab. 11:70–76. 2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Akiyama T, Tachibana I, Shirohara H,
Watanabe N and Otsuki M: High-fat hypercaloric diet induces
obesity, glucose intolerance and hyperlipidemia in normal adult
male Wistar rat. Diabetes Res Clin Pract. 31:27–35. 1996.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Dominiczak MH: Obesity, glucose
intolerance and diabetes and their links to cardiovascular disease
Implications for laboratory medicine. Clin Chem Lab Med.
41:1266–1278. 2003.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Fernandez ML and West KL: Mechanisms by
which dietary fatty acids modulate plasma lipids. J Nutr.
135:2075–2078. 2005.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Stefan N, Kantartzis K and Haring HU:
Causes and metabolic consequences of Fatty liver. Endocr Rev.
29:939–960. 2008.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Eleftheriadis T, Pissas G, Liakopoulos V
and Stefanidis I: IDO decreases glycolysis and glutaminolysis by
activating GCN2K, while it increases fatty acid oxidation by
activating AhR, thus preserving CD4+ T-cell survival and
proliferation. Int J Mol Med. 42:557–568. 2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Eleftheriadis T, Pissas G, Antoniadi G,
Liakopoulos V and Stefanidis I: Indoleamine 2,3-dioxygenase
depletes tryptophan, activates general control non-derepressible 2
kinase and down-regulates key enzymes involved in fatty acid
synthesis in primary human CD4+ T cells. Immunology. 146:292–300.
2015.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Byersdorfer CA: The role of Fatty Acid
oxidation in the metabolic reprograming of activated t-cells. Front
Immunol. 5(641)2014.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Lochner M, Berod L and Sparwasser T: Fatty
acid metabolism in the regulation of T cell function. Trends
Immunol. 36:81–91. 2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Eleftheriadis T, Pissas G, Sounidaki M,
Antoniadi G, Rountas C, Liakopoulos V and Stefanidis L: Tryptophan
depletion under conditions that imitate insulin resistance enhances
fatty acid oxidation and induces endothelial dysfunction through
reactive oxygen species-dependent and independent pathways. Mol
Cell Biochem. 428:41–56. 2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Ghosh A, Gao L, Thakur A, Siu PM and Lai
CWK: Role of free fatty acids in endothelial dysfunction. J Biomed
Sci. 24(50)2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
de Jongh RT, Serné EH, Ijzerman RG, de
Vries G and Stehouwer CD: Free fatty acid levels modulate
microvascular function: Relevance for obesity-associated insulin
resistance, hypertension, and microangiopathy. Diabetes.
53:2873–2882. 2004.PubMed/NCBI View Article : Google Scholar
|
|
51
|
I S Sobczak A, A Blindauer C and J Stewart
A: Changes in plasma free fatty acids associated with type-2
diabetes. Nutrients. 11(E2022)2019.PubMed/NCBI View Article : Google Scholar
|