Introduction
Cross-linked fibrin and activated platelets
constitute the main components of a thrombus (1). Moreover, the activation of platelet
glycoprotein IIb/IIIa (GPIIb/IIIa), which is abundantly expressed
on the platelet surface (2), is
the final common pathway of platelet aggregation (3). Therefore, fibrin, the fibrin
degradation product (D-dimer) and GPIIb/IIIa may be used as targets
in thrombolysis. Since single-chain urokinase plasminogen activator
(scu-PA) was covalently linked to the Fab’ region of a monoclonal
antibody specific for fibrin (antibody 59D8) by Bode et al
(4), targeted thrombolytics have
become a popular research topic. Targeted thrombolytics are
synthesized by connecting thrombus-specific antibodies to
thrombolytic drugs via chemical or biological methods, thus
producing a new type of drug with high avidity and specificity for
the thrombus. This may reduce its reaction with non-target
tissues.
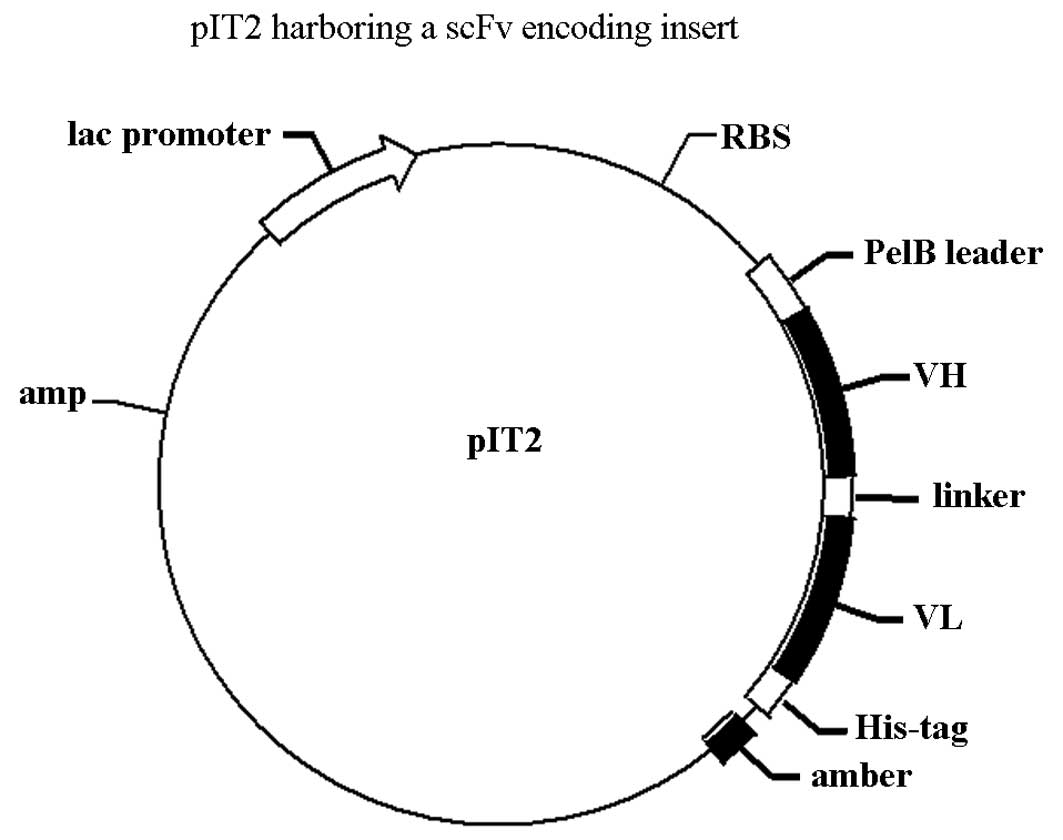
A single-chain variable fragment (scFv), which
retains the specificity of the original immunoglobulin, is a fusion
protein of the variable regions of the heavy (VH) and light (VL)
chains of immunoglobulins connected to a linker peptide (5). In previous studies, our research
group has successfully isolated specific human monoclonal
anti-D-dimer scFv antibodies (6)
and monoclonal anti-GPIIb/IIIa scFv antibodies from scFv phage
libraries (7); the two scFv
fragments were produced in Escherichia coli in soluble forms
with good retention of antigen-binding activities. Previously,
researchers devised methods for linking two scFvs to produce a
single peptide chain with two VH and two VL regions, yielding
bispecific scFvs (bs-scFvs) with a specificity for two different
antigens (8,9). Therefore, in this study, we used the
plasmids of anti-D-dimer scFv and anti-GPIIb/IIIa scFv to construct
a prokaryotic plasmid expressing GPIIb-IIIa and D-dimer bs-scFvs.
The single-chain diabody binds two specific antigens simultaneously
and may remarkably improve specificity and functional avidity to a
thrombus; therefore, it lays a sound foundation for further
research on target-oriented thrombolytics.
Materials and methods
Materials
The human anti-D-dimer scFv component, designated
A1, and the human anti-GPIIb-IIIa scFv component, designated G9,
which were previously isolated from a human scFv phage display
library, were employed as fusion partners for the creation of a
bs-scFv. The two scFvs were assembled in a VH-to-VL orientation,
where the V-domains were attached by a 15 amino acid residue linker
of composition (Gly4Ser)3, which did not
interfere with antigen binding (Fig.
1). The gene sequences of A1-scFv and G9-scFv have been
determined previously (6,7,). The primers were synthesized by
Tsingke Biotechnology Co., Ltd. (Beijing, China). The primers are
shown in Table I; the primers
named linker+vlb+ and vlb− were
phosphorylated at the 5′ end. KOD Plus High Fidelity DNA polymerase
was purchased from Toyobo Co., Ltd. (Osaka, Japan). T4 DNA ligase
was purchased from New England Biolabs (Ipswich, MA, USA). The NTA
column was purchased from Merck KGaA (Darmstadt, Germany). All
other reagents were domestically produced biochemical analytical
reagents.
 | Table I.Primers for polymerase chain
reaction. |
Table I.
Primers for polymerase chain
reaction.
| Primer name | Sequence |
|---|
| vla− | CCG TTT GAT TTC CAC
CTT GGT |
|
Vector+ | GCG GCC GCA CAT CAT
CAT CAC CAT CA |
|
linker+vlb+ | GGG GGC GGG GGA TCA ATG GCC GAG
GTG CAG CTG T |
| vlb− | CCG TTT GAT TTC CAC
CTT GGT CCC TTG |
Vector construction
Taking the anti-D-dimer circular plasmid as a
template, PCR was performed using primers vla− and
Vector+ to obtain a linear plasmid, named construct I
(Fig. 2) with two blunt ends. To
introduce the Gly4Ser linker at the 5′ end, PCR
fragments with two blunt ends were generated (construct II) via two
primers: linker+vlb+ and vlb−. All
the above PCR fragments were amplified with KOD Plus High Fidelity
DNA polymerase. Following identification by agarose gel
electrophoresis, constructs I and II (Fig. 2) were retrieved and purified using
the QIA Quick Extraction kits. Then, constructs I and II were
linked together using T4 DNA ligase to generate the recombinant
circular plasmid of anti-D-dimer/anti-GPIIb-IIIa diabody,
designated pIT2-A1G9. The generation procedure of pIT2-A1G9 is
described in Fig. 2. Following the
transfection of pIT2-A1G9 into the competent cell line HB2151, the
recombinant clone was selected from the ampicillin agar plate and
characterized by PCR. Next, the clones with the correct insertion
sequences were determined by sequencing.
Bacterial expression
A single recombinant colony was selected and
inoculated overnight in 5 ml Luria-Bertani (LB) culture solution
containing 100 mg/ml ampicillin. Then, the colony was transferred
to 250 ml LB medium containing 100 mg/ml ampicillin and inoculated
to an OD600 of 1.0. Isopropylthio-β-galactoside (IPTG) was added to
a final concentration of 0.4 mM and growth was continued at 37°C
for 3.5 h. Cells were harvested by centrifugation at 1,800 × g for
30 min and frozen at −20°C. Following resuspension in 20 mM
Tris-HCl and 0.2 M NaCl (pH 8.0), cells were broken by a high
pressure homogenizer and centrifugation at 12,000 × g for 30 min.
Then, the supernatants containing the soluble product were analyzed
by sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE).
Purification of the recombinant
protein
The supernatants containing the recombinant antibody
were purified using the NTA column. The bound proteins were eluted
with a gradient of imidazole from 30 to 500 mM, then the fractions
were analyzed by SDS-PAGE. Elution occurred at a concentration of
30 mM imidazole as a distinct peak. Then, 30 mM imidazole eluate
containing the recombinant diabody was further purified with 10 mM
phosphate-buffered saline (PBS) by gel filtration chromatography
(HiPrep™ 16/60 Sephacryl S-200 high resolution column).
All eluted fractions were collected and measured at 280 nm for
protein.
Binding specificity of the recombinant
antibody
After the ELISA plates were coated with 25
μg/ml bovine serum albumin (BSA; negative control), 25
μg/ml GPIIb/IIIa or 25 μg/ml D-dimer, respectively,
50 μl purified recombinant antibody (20 μg/ml in PBS)
was added and detected with a 1:5,000 dilution of
peroxidase-conjugated anti-6X histidine. The absorbance values were
measured following the addition of the substrate.
Results
Successful construction of bispecific
single-chain molecules
For the generation of bispecific D-dimer GPIIb-IIIa
single-chain constructs, the anti-D-dimer scFv fragments (named
A1-scFv) and anti-GPIIb-IIIa scFv fragments (named G9-scFv) were
fused into a tandem using a five amino acid residue glycine-serine
linker (Fig. 1), which was
considered to prevent the formation of scFv molecules from the
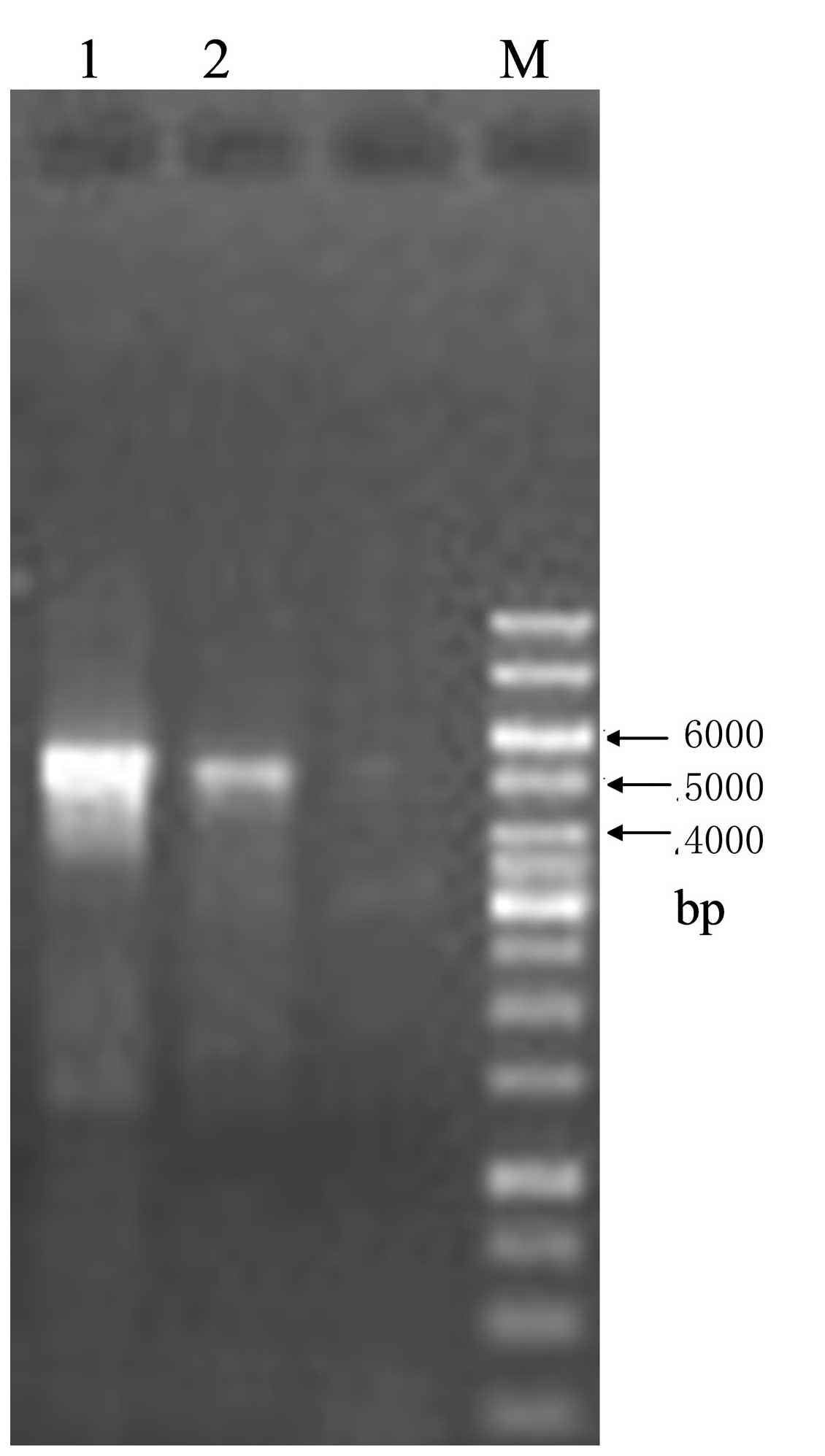
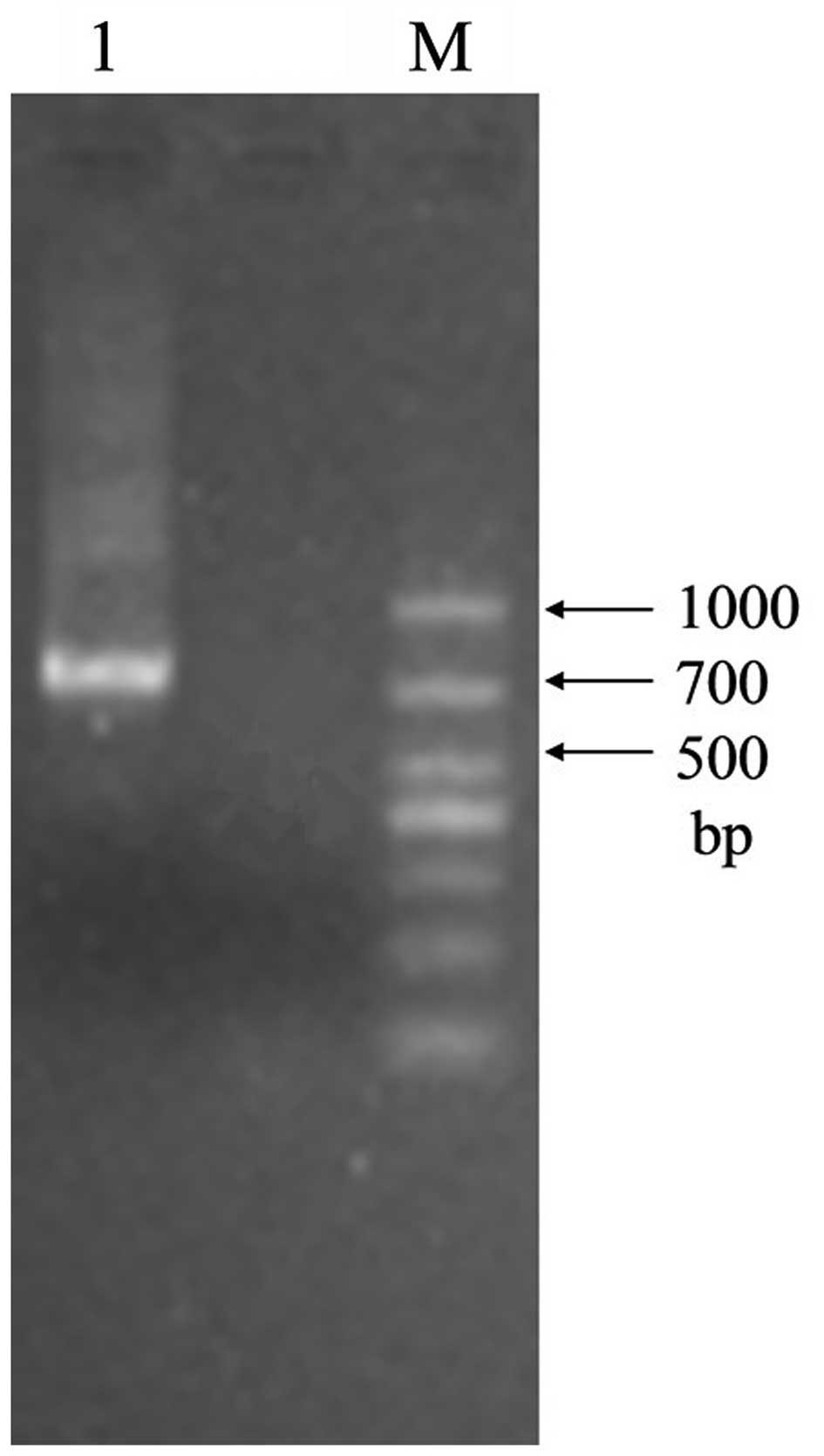
adjacent A1VL and G9VH domains. The size of the empty vector pIT2
was 4.2 kb, while the size of the scFv insert was ∼750 bp. The
expected sizes of construct I and construct II were ∼4,950 bp and
∼750 bp, respectively. Agarose gel electrophoresis confirmed that
the actual sizes of purified construct I and construct II were in
line with the expectations (Figs.
3 and 4). The recombinant
clone in the correct A1VH-A1VL-G9VH-G9VL orientation was selected
from the ampicillin agar plate and underwent PCR and sequencing
(figure not shown).
Expression and purification of the
recombinant protein
The diabody (∼56 kDa) was successfully expressed in
a small amount with the induction of IPTG (Fig. 5, lane 2), and was identified in the
supernatant (Fig. 5, lane 3) and
precipitate (Fig. 5, lane 4) of
bacterial lysates. As the diabody mainly existed in the supernatant
of bacterial lysates, it was a soluble protein. Following affinity
chromatography using the Ni-NTA column, one main band with the
molecular weight of 56 kDa existed in the 30 and 500 mmol/l
imidazole eluates, yet the majority of the diabody was located in
the 30 mmol/l imidizole eluate (Fig.
5, lanes 6 and 7). However, with miscellaneous protein
contamination, the recombinant protein was not pure enough;
therefore, it required further purification. Following purification
by molecular sieve chromatography, the target protein presented one
single band in SDS-PAGE (Fig. 6).
As a result, we obtained the electrophoretically pure recombinant
protein. The purity was >90% and the concentration was 1.65
mg/ml.
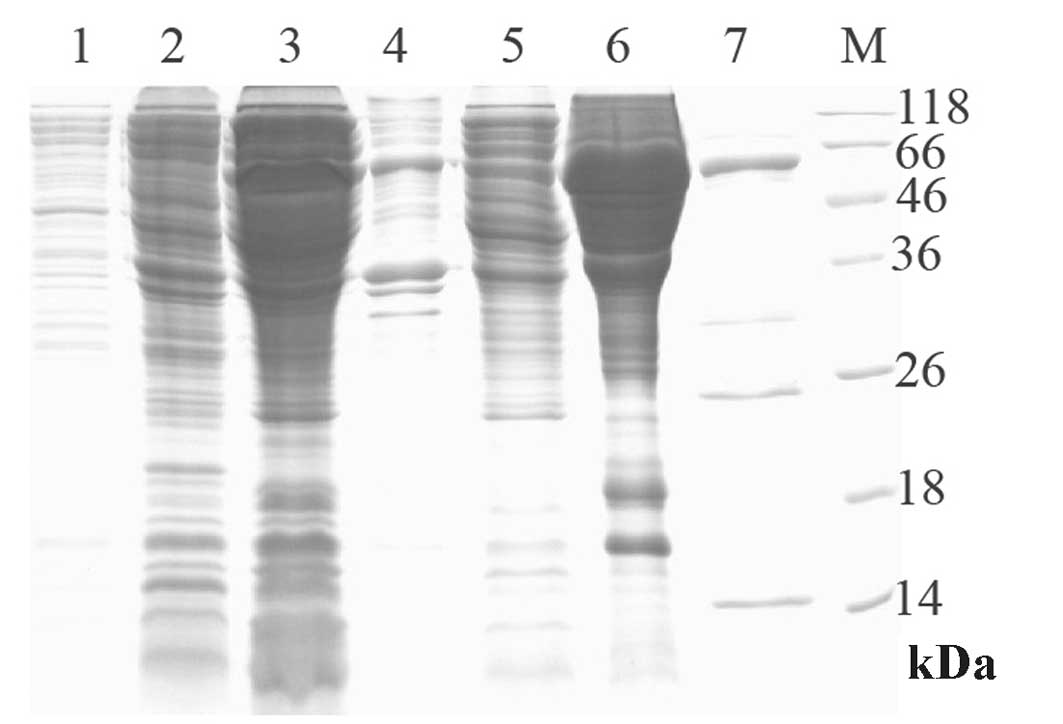 | Figure 5.Expression and purification of the
diabody (10% SDS-PAGE). Lane 1, total protein of bacterial
pIT2-A1G9 prior to induction; lane 2, total protein of bacterial
pIT2-A1G9 induced with IPTG; lane 3, periplasmic lysates of
bacterial pIT2-A1G9 induced with IPTG; lane 4, cell lysate
precipitate of bacterial pIT2-A1G9 induced with IPTG; lane 5, the
Ni-NTA column flow-through; lane 6, the eluate washed at a
concentration of 30 mM imidazole; lane 7, the eluate washed at a
concentration of 500 mM imidazole; lane M, protein marker.
SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis; IPTG, isopropylthio-β-galactoside. |
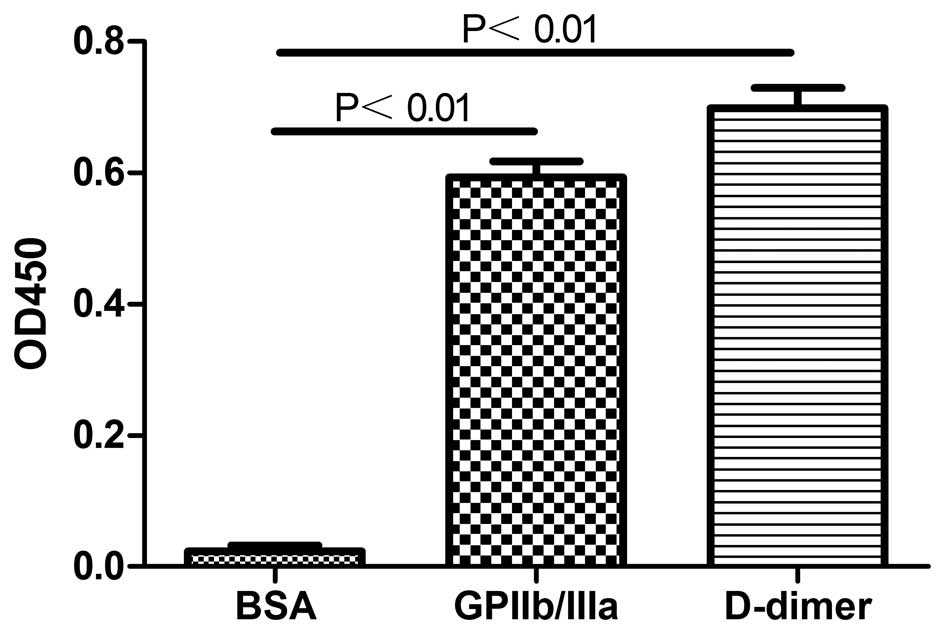
Binding specificity of the recombinant
protein
The purified recombinant protein was shown to be
specific for GPIIb/IIIa and D-dimer since there was no
cross-binding to other proteins, including BSA (Fig. 7).
Discussion
Fibrin is a clear target for antithrombotic or
fibrinolytic agents. Sufficient amounts of fibrin are present even
in platelet-rich thrombi. Certain types of fibrin-targeted
anticoagulants have been produced. Antibodies, including MA-15C5,
directed against the D-dimer fragment of cross-linked human fibrin,
have been fused to recombinant scu-PA and used successfully to
target clots (10). A study in
various in vivo models of venous thrombosis has demonstrated
that thrombolysis by 59D8-scuPA is significantly faster and more
potent compared with that by the clinically used urokinase
(11). Another fibrin-targeted
anticoagulant was successfully developed by fusing hirudin to the
generated fibrin-specific scFv of 59D8 to target a developing clot
(12). Moreover, studies
concerning platelet-targeted anticoagulants have also been reported
(13,14). In one study, an anti-GPIIb/IIIa
single-chain antibody was genetically fused with a potent, direct
factor Xa (fXa) inhibitor and tick anticoagulant peptide (TAP)
(15). However, these chimeric
proteins target only one portion of the thrombus: fibrin or
platelets. Thrombolytics that targeted fibrin and platelets
simultaneously may have enhanced potency and clot specificity. A
bispecific antifibrin-antiplatelet urokinase conjugate (BAAUC) was
created by coupling urokinase to the monovalent Fab’ from the
antifibrin monoclonal antibody 59D8 and the monovalent Fab’ from
the anti-glycoprotein GPIIb/IIIa monoclonal antibody 7E3 (16). In vitro, this bispecific
drug has the potency to lyse fibrin-rich and platelet-rich thrombi
with high efficacy and to effectively inhibit platelet aggregation.
However, for penetrating into the thrombus, a target-oriented
thrombolytic agent with a smaller molecular weight than BAAUC is
required. Moreover, the majority of monoclonal antibodies or Fab’
portions in recombinant proteins are derived from mice and may
produce human anti-mouse antibodies (17). Thus, we aimed to construct a new
and effective anticoagulant or thrombolytic agent with a small
molecular weight, low immunogenicity, strong tissue penetrating
force and a good specific binding capacity for thrombi.
As the scFv is the smallest antibody fragment with a
complete antigen-binding site (18), we used scFv molecules specific for
D-dimer and GPIIb-IIIa to construct the diabody. This construct is
smaller than those containing whole Fab’ fragments and has improved
thrombus penetration. Furthermore, the anti-D-dimer and
anti-GPIIb-IIIa scFvs are from a fully human single-chain Fv
library; therefore, the recombinant diabody is a human
antibody.
In our study, we used a non-standard method to
construct bispecific antibodies. Since Holliger et al
(19) invented the diabody by
cross-linking the genes of the heavy-chain and light-chains of the
variable regions of two antibodies to form a hybrid scFv, the
majority of diabodies have been created by restriction enzyme
digestion and ligation (20,21).
In a change from the common method, we used blunt-end ligation to
generate the recombinant plasmid. As the gene sequences of the
anti-D-dimer and anti-GPIIb-IIIa scFvs are known, using the
circular plasmids as templates, the gene of anti-GPIIb-IIIa scFv
was conveniently amplified and inserted into the vector pIT2, which
already contained the anti-D-dimer scFv (Fig. 2). PCR and gene sequencing
demonstrated that a new plasmid of the diabody in the
A1VH-A1VL-G9VH-G9VL orientation was successfully constructed. In
this method, high fidelity PCR is the crucial step, particularly
since the amplified product (construct I) was particularly long at
∼4,950 bp (Fig. 3). However, the
superiority of this blunt-end ligation method is the reduction in
the number of processes for generating the recombinant plasmid due
to the lack of restriction enzyme digestion steps, which is
convenient for researchers.
In our recombinant plasmid, the two scFv genes share
only lac promoter and terminator without mutual influence. After
eliminating the repression function of lac by IPTG, a
double-specific (scFv)2 fragment was expressed in HB2151
cells. Linking the two scFvs tandemly with a linker is possibly the
simplest way to keep two scFv together as bispecific molecules, and
is likely to avoid undesired associations. As the molecular weight
of the A1-scFv or G9-scFv monoclonal antibody is 28 kDa (6,7), the
expected molecular weight of the double-specific (scFv)2
fragment is 56 kDa, which was verified by SDS-PAGE analysis
(Fig. 5). Diabodies represent a
class of bispecific antibody fragments similar in size to a Fab
fragment (22). Their small size
potentially gives them access to tissues such as thrombi that are
poorly accessible to intact antibodies, allows rapid clearance from
blood and non-targeted tissues and lowers the immunogenic
response.
The ELISA results demonstrated that the new D-dimer
and GPIIb/IIIa single-chain bispecific antibody was able to
simultaneously identify and bind two specific antigens (Fig. 7). Whether it is possible to apply
this diabody in vivo requires further experimental
investigation. If it presents a good specific binding capacity for
platelets and fibrin in vivo, the diabody itself may be
applied as an antiplatelet agent or be conjugated with a
thrombolytic drug for research into its target-oriented
thrombolytic function in vitro. Therefore, an ideal
target-oriented thrombolytic drug may be produced. With the
advantages of a lower molecular weight, higher antigen-binding
power and comparatively lower immunogenicity, it is likely to have
a high specificity and functional avidity for thrombi.
Acknowledgements
This study was funded by the National
Natural Science Foundation of China (No. 30872525).
References
|
1.
|
Cho J and Mosher DF: Enhancement of
thrombogenesis by plasma fibronectin cross-linked to fibrin and
assembled in platelet thrombi. Blood. 107:3555–3563. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Bennett JS: Structure and function of the
platelet integrin alpha-Ibbeta3. J Clin Invest. 115:3363–3369.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Davì G and Patrono C: Platelet activation
and atherothrombosis. N Engl J Med. 357:2482–2494. 2007.
|
|
4.
|
Bode C, Runge MS, Schönermark S, et al:
Conjugation to antifibrin Fab’ enhances fibrinolytic potency of
single-chain urokinase plasminogen activator. Circulation.
81:1974–1980. 1990.PubMed/NCBI
|
|
5.
|
Peterson E, Owens SM and Henry RL:
Monoclonal antibody form and function: manufacturing the right
antibodies for treating drug abuse. AAPS J. 8:E383–E390.
2006.PubMed/NCBI
|
|
6.
|
Xia HL, Tan Z, Chen DJ, Qiao JG and Qiu
RF: Isolation of specific humanized anti-D-dimer scFv fragments
from scFv phage libraries. Zhonghua Wei Sheng Wu Xue He Mian Yi Xue
Za Zhi. 31:168–172. 2011.(In Chinese).
|
|
7.
|
Xia HL, Tan Z, Chen DJ, Qiao JG and Qiu
RF: Production of monoclonal anti-GP II b/IIIa scFv antibodies from
scFv phage libraries. Chin J Exp Surg. 28:529–532. 2011.
|
|
8.
|
Dincq S, Bosman F, Buyse MA, et al:
Expression and purification of monospecific and bispecific
recombinant antibody fragments derived from antibodies that block
the CD80/CD86-CD28 costimulatory pathway. Protein Expr Purif.
22:11–24. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Jongmans W, van den Oudenalder K,
Tiemessen DM, et al: Targeting of adenovirus to human renal cell
carcinoma cells. Urology. 62:559–565. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Holvoet P, Laroche Y, Stassen JM, et al:
Pharmacokinetic and thrombolytic properties of chimeric plasminogen
activators consisting of a single-chain Fv fragment of a
fibrin-specific antibody fused to single-chain urokinase. Blood.
81:696–703. 1993.
|
|
11.
|
Dewerchin M, Vandamme AM, Holvoet P, et
al: Thrombolytic and pharmacokinetic properties of a recombinant
chimeric plasminogen activator consisting of a fibrin fragment
D-dimer specific humanized monoclonal antibody and a truncated
single-chain urokinase. Thromb Haemost. 68:170–179. 1992.
|
|
12.
|
Peter K, Graeber J, Kipriyanov S, et al:
Construction and functional evaluation of a single-chain antibody
fusion protein with fibrin targeting and thrombin inhibition after
activation by factor Xa. Circulation. 101:1158–1164. 2000.
View Article : Google Scholar
|
|
13.
|
Okabayashi K, Tsujikawa M, Morita M, et
al: Secretory production of recombinant urokinase-type plasminogen
activator-annexin V chimeras in Pichia pastoris. Gene.
177:69–76. 1996. View Article : Google Scholar
|
|
14.
|
Jiang P, Changgeng R and Ru B:
Construction and expression of antibody targeted plasminogen
activator*. Enzyme Microb Technol. 27:755–760. 2000.
|
|
15.
|
Stoll P, Bassler N, Hagemeyer CE, et al:
Targeting ligand-induced binding sites on GPIIb/IIIa via
single-chain antibody allows effective anticoagulation without
bleeding time prolongation. Arterioscler Thromb Vasc Biol.
27:1206–1212. 2007. View Article : Google Scholar
|
|
16.
|
Ruef J, Nordt TK, Peter K, Runge MS,
Kübler W and Bode C: A bispecific antifibrin-antiplatelet urokinase
conjugate (BAAUC) induces enhanced clot lysis and inhibits platelet
aggregation. Thromb Haemost. 82:109–114. 1999.
|
|
17.
|
Tcheng JE, Kereiakes DJ, Lincoff AM, et
al: Abciximab read-ministration: results of the ReoPro
Readministration Registry. Circulation. 104:870–875. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Leath CA III, Douglas JT, Curiel DT and
Alvarez RD: Single-chain antibodies: A therapeutic modality for
cancer gene therapy (review). Int J Oncol. 24:765–771.
2004.PubMed/NCBI
|
|
19.
|
Holliger P, Prospero T and Winter G:
“Diabodies”: small bivalent and bispecific antibody fragments. Proc
Natl Acad Sci USA. 90:6444–6448. 1993.
|
|
20.
|
Liu F, Chen Z, Jiang W, Yang C, Xiong D
and Zhu Z: Improvement in soluble expression levels of a diabody by
exchanging expression vectors. Protein Expr Purif. 62:15–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Stamova S, Cartellieri M, Feldmann A, et
al: Unexpected recombinations in single chain bispecific
anti-CD3-anti-CD33 antibodies can be avoided by a novel linker
module. Mol Immunol. 49:474–482. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Kriangkum J, Xu B, Nagata LP, Fulton RE
and Suresh MR: Bispecific and bifunctional single chain recombinant
antibodies. Biomol Eng. 18:31–40. 2001. View Article : Google Scholar : PubMed/NCBI
|