Introduction
Tumor invasion and metastasis are among the most
threatening features of the oncogenic process and are the cause of
90% of human cancer-related deaths (1). Tumor cell invasion and metastasis
are complex processes involving extracellular matrix (ECM)
degradation and cell migration through the ECM (2,3).
Matrix metalloproteinases (MMPs) are a family of zinc-dependent
endopeptidases that play a crucial role in invasion and metastasis
through the degradation of the ECM (4). MMPs are upregulated in the majority
of human cancers and uncontrolled proteolysis plays a crucial role
in several pathological conditions (5). Several studies have demonstrated
that MMP-2 and MMP-9 are critical for tumor invasion and metastasis
by degrading collagen IV, a main component of basement membranes
(6–8). A previous study indicated that
MMP-2, as opposed to MMP-9, is closely associated with invasion and
migration in MCF10A human breast epithelial cells harboring mutated
H-ras (H-ras MCF10A cells) (9).
The uncontrolled activation of the ras
signaling pathway is one of the most frequent defects in human
cancers (10). In human breast
cancer, ras mutations are infrequent, but upregulated
amounts of the ras protein have been found in 60–70% of human
breast cancers (11). A number of
studies have suggested that the expression of ras is associated
with the invasiveness of breast cancer cells and is crucial for
tumor aggressiveness, including tumor recurrence, degree of
invasion of fat tissues and infiltration into lymphatic vessels
(11–13). A previous study indicated that
H-ras, but not N-ras, induces the invasive phenotype of MCF10A
cells and suggested the important role of MMP-2 in cell invasion
(14).
The best characterized effectors of Ras are the Raf
kinases (15) and
phosphatidylinositol 3-kinases (PI3Ks) (16). PI3Ks are heterodimeric lipid
kinases that are involved in various cellular responses, including
cell growth, proliferation, cytoskeletal rearrangement and motility
(17). Activated PI3K synthesizes
the second messenger, phosphatidylinositol (3,4,5)-trisphosphate (PIP3), from
phosphatidylinositol (4,5)-bisphosphate (PIP2),
thereby affecting the activation of phosphoinositide-dependent
kinases (PDKs) and Akt signaling (18). The accumulation of PIP3
creates a docking site for Akt at the plasma membrane, which binds
to PIP3 via the pleckstrin homology domain. Akt is a
serine/threonine kinase and its activation has been implicated in
the genesis or progression of various human malignancies (17,19). Significantly upregulated PI3K
activity has been detected in highly invasive MDA-MB-231 cells
(20). One of the characteristics
of highly invasive cancers is the aberrant activation of nuclear
factor-κB (NF-κB) and PI3K activates NF-κB through distinct
mechanisms in different cell lines (20,21). It has previously been demonstrated
that the PI3K/Akt signaling pathway is involved in the invasive
phenotype of H-ras MCF10A cells (9).
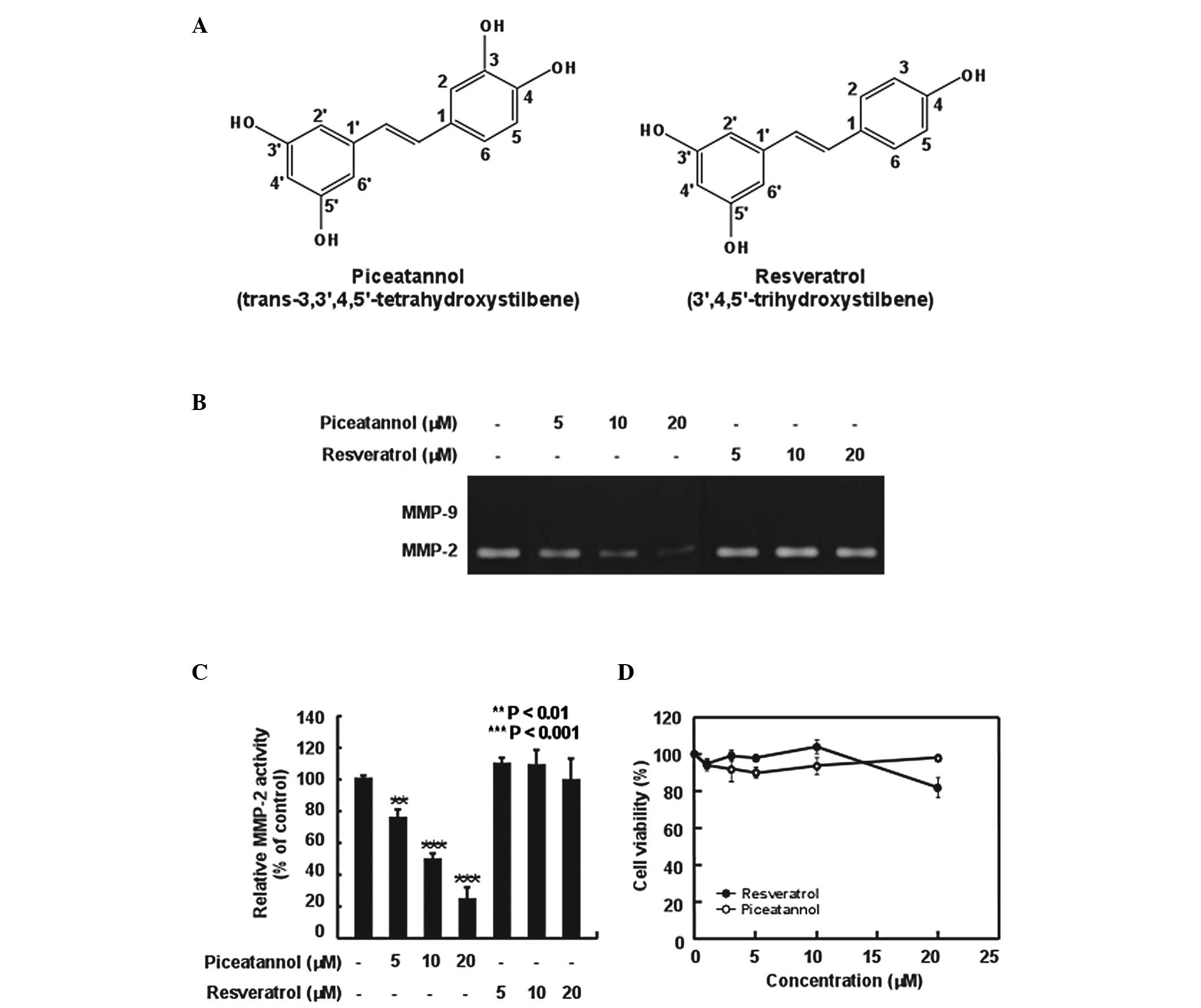
Resveratrol (3,5,4′-trihydroxystilbene; Fig. 1A), a phenolic compound of red
wine, has been reported to be a natural chemopreventive agent
against several types of cancer, such as leukemia (22), breast (23) and prostate cancer (24). Piceatannol
(3,5,3′,4′-tetrahydroxystilbene; Fig.
1A) is a naturally occurring polyphenol and a metabolite of the
cancer chemopreventive agent, resveratrol, via cytochrome P4501B1,
which is overexpressed in a variety of human tumors (25). It has been demonstrated that
piceatannol exerts anti-inflammatory (26) and anticancer effects (27). However, the cancer preventive
effects of piceatannol and the underlying molecular mechanisms have
not yet been fully elucidated. In this study, we investigated the
possible inhibitory effects of piceatannol on the H-ras-induced
invasiveness of MCF10A human breast epithelial cells, as well as
the underlying mechanisms.
Materials and methods
Chemicals
Piceatannol was obtained from A.G. Scientific, Inc.
(San Diego, CA, USA). Resveratrol was purchased from Sigma-Aldrich
(St. Louis, MO, USA). LY294002 and triciribine were obtained from
Calbiochem (San Diego, CA, USA). Dulbecco’s modified Eagle’s medium
(DMEM)/F12, horse serum, L-glutamine and the
penicillin/streptomycin/fungizone mixture were purchased from
Gibco-BRL (Grand Island, NY, USA). Protein A/G sepharose beads were
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). The antibodies against phosphorylated Akt (Ser473) and total
Akt were obtained from Cell Signaling Technology (Beverly, MA,
USA). The antibodies against PI3K p85 and p110 were obtained from
Upstate Biotechnology (Lake Placid, NY, USA). CNBr-Sepharose 4B and
[γ-32P]ATP were purchased from Amersham Pharmacia
Biotech (Piscataway, NJ, USA) and the protein assay kit was from
Bio-Rad Laboratories (Hercules, CA, USA).
Cell culture
H-ras MCF10A cells (kindly supplied by Dr Aree Moon,
Duksung Women’s University, Seoul, Korea) were cultured in DMEM/F12
supplemented with 5% heat-inactivated horse serum, 10 μg/ml
insulin, 100 ng/ml cholera toxin, 0.5 μg/ml hydrocortisone, 20
ng/ml recombinant epidermal growth factor, 2 mM L-glutamine and 100
ng/ml penicillin/streptomycin/fungizone mixture in a 37°C
humidified atmosphere of 5% CO2/95% air.
Gelatin zymography for MMPs
The H-ras MCF10A cells (5×105) plated on
culture dishes at 90% confluence were maintained in serum-free
medium for 24 h. The cells were cultured in serum-free medium
containing chemicals or chemical inhibitors for a further 24 h.
Conditioned medium was collected and concentrated at 10,000 × g for
1 h in a SpeedVac concentrator (Savant/E-C Instruments, Niantic,
CT, USA). The protein concentration was measured using a
dye-binding protein assay kit (Bio-Rad Laboratories) as described
in the manufacturer’s manual. Equal amounts of protein from the
conditioned medium were mixed with 2X non-reducing sample buffer,
incubated for 5 min at room temperature and then electrophoresed on
10% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis
(PAGE) gels containing 1 mg/ml gelatin. After electrophoresis, the
gels were washed with 2.5% Triton X-100 three times each for 30 min
to remove SDS, rinsed two times each for 10 min with a 50 mM
Tris-HCl buffer (pH 7.6) containing 5 mM CaCl2, 200 mM
NaCl and 0.02% Brij-35 and then incubated overnight at 37°C. The
gels were stained with 0.5% Coomassie brilliant blue R-250 solution
containing 10% acetic acid and 20% methanol for 30 min and then
destained with 7.5% acetic acid solution containing 10% methanol.
Areas of gelatinase activity appear as clear bands (zones of
gelatin degradation) against the blue-stained gelatin
background.
Cell viability assay
To estimate cell viability, H-ras MCF10A cells were
seeded in 96-well plates (n=3) and grown to near confluence in
conditioned medium. The H-ras MCF10A cells were then starved for 24
h. The cells were either treated or not treated with various
concentrations of piceatannol or resveratrol (1.25–20 μM) for 24 h.
Subsequently, 20 μl of MTT solution (0.5 mg/ml) were added to each
well containing 200 μl of conditioned medium and incubated for a
further 4 h at 37°C. The medium was then removed and 200 μl of
dimethyl sulphoxide (DMSO) were added to each well. After shaking,
cell viability was determined by reading the absorbance at 570 nm
and the results were expressed as the cell viability ratio relative
to the untreated control.
Invasion assay
An invasion assay was performed using the Transwell
system (Corning Costar Co., Cambridge, MA, USA). The lower side of
the filter was coated with 10 μl of type I collagen and the upper
side was coated with Matrigel (Upstate Biotechnology). The lower
compartment was filled with 600 μl of serum-free medium containing
0.1% bovine serum albumin (BSA). The H-ras MCF10A cells
(5×104) were resuspended with samples in 100 μl of
medium and placed in the upper part of the Transwell plate. The
cells were then incubated for 24 h in a humidified atmosphere of 5%
CO2 at 37°C. The cells on the upper surface of the
filter were completely wiped off with a cotton swab, fixed with
methanol and stained with hematoxylin for 10 min. After washing in
water, the cells were stained with eosin for 4 min. The invading
cells were quantified by counting the cells that had migrated to
the lower side of the filter under a microscope. Thirteen randomly
selected fields were counted and each sample was assayed in
duplicate.
Wound migration assay
The H-ras MCF10A cells (5×105) were
plated on culture dishes and grown to 90% confluence in 2 ml of
growth medium. The cells were cultured with mitomycin C (25 μg/ml)
for 30 min and then an injury line was made using a plastic pipette
tip (all tips used had the same width). The plates were rinsed with
phosphate-buffered saline (PBS) and then complete medium containing
the samples was added. The cells were allowed to migrate in the
medium and images were acquired using an inverted microscope
(x100/x200 magnification; Olympus Ix70, Okaya, Japan) at specific
time points. The width of the injury line was then measured in
three independent experiments and was plotted as a percentage of
the width at 0 h.
Western blot analysis
After the cells were cultured for 24 h, they were
starved in serum-free DMEM/F12 for a further 24 h. The cells were
then treated with chemicals for 30 min. Cell lysates were scraped
and treated with lysis buffer [10 mM Tris (pH 7.5), 150 mM NaCl, 5
mM EDTA, 1% Triton X-100, 1 mM DTT, 0.1 mM PMSF, 10% glycerol and a
protease inhibitor cocktail tablet] for 40 min on ice followed by
centrifugation at 14,000 rpm for 10 min. The protein concentration
of the supernatant was determined using a dye-binding protein assay
kit (Bio-Rad Laboratories) as described in the manufacturer’s
manual. Lysate protein (30 μg) was subjected to 10% SDS-PAGE and
electrophoretically transferred onto a polyvinylidene difluoride
(PVDF) membrane (Immobilon P, Millipore Corp., Bedford, MA, USA).
After blotting, the membrane was blocked in 5% fat-free dry milk
for 1 h and then incubated with the specific primary antibody for 2
h at room temperature. Protein bands were detected by using an
enhanced chemiluminescence (ECL) detection kit (Amersham) after
hybridization with the HRP-conjugated secondary antibody.
In vitro PI3K assay
An active PI3K protein (80 ng) was incubated with
piceatannol or LY294002 at the indicated concentrations for 10 min
at 30°C. The mixtures were then incubated with 20 μl of 0.5 mg/ml
phosphatidylinositol (Avanti Polar Lipids, Inc., Alabaster, AL,
USA). After 5 min at room temperature, the mixtures were incubated
with reaction buffer [100 mM HEPES (pH 7.6), 50 mM
MgCl2, 250 μM ATP containing 10 μCi of
[γ-32P]ATP] for an additional 10 min at 30°C. The
reaction was terminated by the addition of 15 μl of 4 N HCl and 130
μl of chloroform/methanol (1:1). After vortexing, 30 μl of the
lower chloroform phase was spotted onto a 1% potassium
oxalate-coated silica gel plate, which was previously activated for
1 h at 110°C. The resulting 32P-labeled
phosphatidylinositol-3-phosphate (PIP) was separated by thin layer
chromatography (TLC) and radiolabeled spots were visualized by
autoradiography.
Immunofluorescence assay
Immunofluorescent staining was conducted on the
H-ras MCF10A cells cultured on cover slips. All staining procedures
were performed on ice or at 4°C unless otherwise stated. The cells
were grown on coverslips and cultured for 24 h. Following treatment
with the indicated doses of piceatannol for 15 min, the cells were
fixed with 4% formaldehyde for 15 min at room temperature and
permeabilized with ice-cold 100% methanol for 10 min. The cells
were stained with PIP3 primary antibody followed by
FITC-conjugated secondary antibody at dilutions recommended by the
manufacturer and cell nuclei were stained with DAPI.
Ex vivo pull-down assay
An H-ras MCF10A cellular supernatant fraction (500
μg) was incubated with piceatannol-Sepharose 4B (or Sepharose 4B as
the control) beads (100 μl, 50% slurry) in reaction buffer [50 mM
Tris-HCl (pH 7.5), 5 mM EDTA, 150 mM NaCl, 1 mM DTT, 0.01% Nonidet
P-40, 2 μg/ml bovine serum albumin, 0.02 mM PMSF and 1X protease
inhibitor mixture]. Following incubation with gentle rocking
overnight at 4°C, the beads were washed five times with buffer [50
mM Tris-HCl (pH 7.5), 5 mM EDTA, 150 mM NaCl, 1 mM DTT, 0.01%
Nonidet P-40 and 0.02 mM PMSF] and proteins bound to the beads were
analyzed by immunoblotting.
Molecular modeling
Insight II (Accelrys, Inc., San Diego, CA, USA) was
used for molecular docking and structure analysis; the crystal
coordinates of PI3K in the complex with ATP (accession code 1E8X)
are available on the Protein Data Bank (http://www.rcsb.org/pdb/).
Statistical analysis
Where appropriate, data are expressed as the means ±
SD and the Student’s t-test was used to perform statistical
analysis for single comparison. Probability values of p<0.05
were considered to indicate statistically significant
differences.
Results
Piceatannol inhibits H-ras-induced MMP-2
activity more effectively than resveratrol in MCF10A cells
MMP-2 and MMP-9 are critical enzymes for ECM
degradation, which is a critical step in tumor metastasis. A
previous study demonstrated that the H-ras-induced invasion of
MCF10A cells is associated more closely with the expression of
MMP-2 than of MMP-9 (9). In this
study, to investigate the protective effect of piceatannol against
cancer metastasis, we first examined the inhibitory effect of
piceatannol on MMP-2 upregulation in H-ras MCF10A cells.
Piceatannol, but not resveratrol, inhibited the upregulation of
MMP-2 activity in the H-ras MCF10A cells in a dose-dependent manner
(Fig. 1B and C). The survival of
the cells treated with piceatannol for up to 24 h was comparable to
that of the cells at 0 h, as determined by MTT assay (Fig. 1D), indicating that the inhibition
of MMP-2 upregulation was not due to piceatannol cytotoxicity.
Piceatannol suppresses H-ras-induced cell
invasion and migration more effectively than resveratrol in MCF10A
cells
Invasion and migration are important phenotypes in
cancer metastasis (28). In this
study, we examined the effects of piceatannol and resveratrol on
cell invasion and migration in H-ras MCF10A cells. The
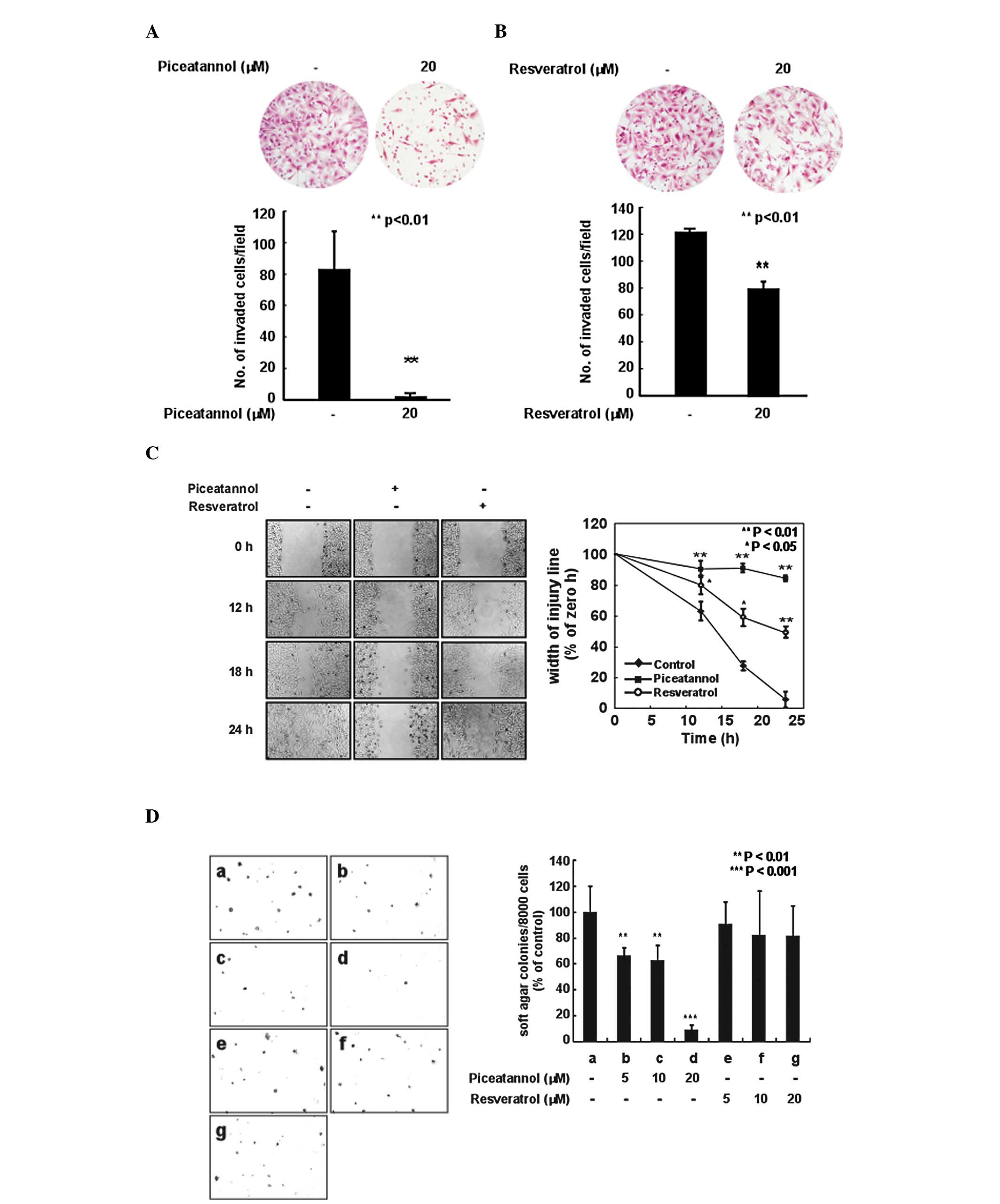
H-ras-induced invasion of MCF10A cells was markedly inhibited
following treatment with piceatannol at a concentration of 20 μM
(Fig. 2A). Resveratrol also
inhibited the invasive ability of the H-ras MCF10A cells (Fig. 2B), although piceatannol suppressed
the H-ras-induced invasive ability of the cells more effectively
than did resveratrol. Subsequently, we investigated the effects of
piceatannol and resveratrol on the H-ras-induced cell migration of
MCF10A cells. As expected, piceatannol inhibited the cell migration
ability at a concentration of 20 μM (Fig. 2C). Resveratrol also suppressed the
migration ability of the H-ras MCF10A cells; however, its effects
were not as prominent as those of piceatannol. Treatment with
piceatannol significantly reduced the H-ras-induced
anchorage-independent growth of MCF10A cells in a dose-dependent
manner (Fig. 2D). Based on the
number of colonies, piceatannol at 20 μM suppressed the
H-ras-induced anchorage-independent growth by 90%. However,
resveratrol did not significantly inhibit H-ras-induced
anchorage-independent growth.
Piceatannol attenuates the H-ras-induced
phosphorylation of Akt and PI3K activity in MCF10A cells
To elucidate the inhibitory mechanisms of
piceatannol on MMP-2 activity, we investigated the effects of
piceatannol on the phosphorylation of Akt and PI3K activity in the
H-ras MCF10A cells. Piceatannol markedly suppressed the
H-ras-induced phosphorylation of Akt in a time- and dose-dependent
manner in the MCF10A cells (Fig. 3A
and B). We also found that 20 μM of piceatannol almost
completely inhibited Akt phosphorylation in the H-ras MCF10A cells;
however, the same concentration of resveratrol did not exert an
inhibitory effect (Fig. 3B). We
also investigated whether piceatannol affects the levels of p38 and
extracellular signal-regulated kinase (ERK) phosphorylation and
found that it did not inhibit p38 and ERK phosphorylation (data not
shown). These results suggest that piceatannol regulates Akt
phosphorylation without affecting p38 and ERK phosphorylation.
Moreover, in order to determine the direct target of piceatannol,
we investigated the effects of piceatannol on PI3K activity, the
upstream kinase of Akt, in vitro. Piceatannol markedly
suppressed PI3K activity and its effects were similar to those of
LY294002, a commercial PI3K inhibitor (Fig. 3C). Activated PI3K generates
PIP3 and phosphorylates Akt (17). Therefore, we then examined the
inhibitory effects of piceatannol on PI3K activity in the H-ras
MCF10A cells by the immunostaining of PIP3. Piceatannol
inhibited PIP3 expression in a dose-dependent manner in
the H-ras MCF10A cells (Fig. 3D).
These results suggest that piceatannol exerts its antimetastatic
effects through the direct inhibition of PI3K activity.
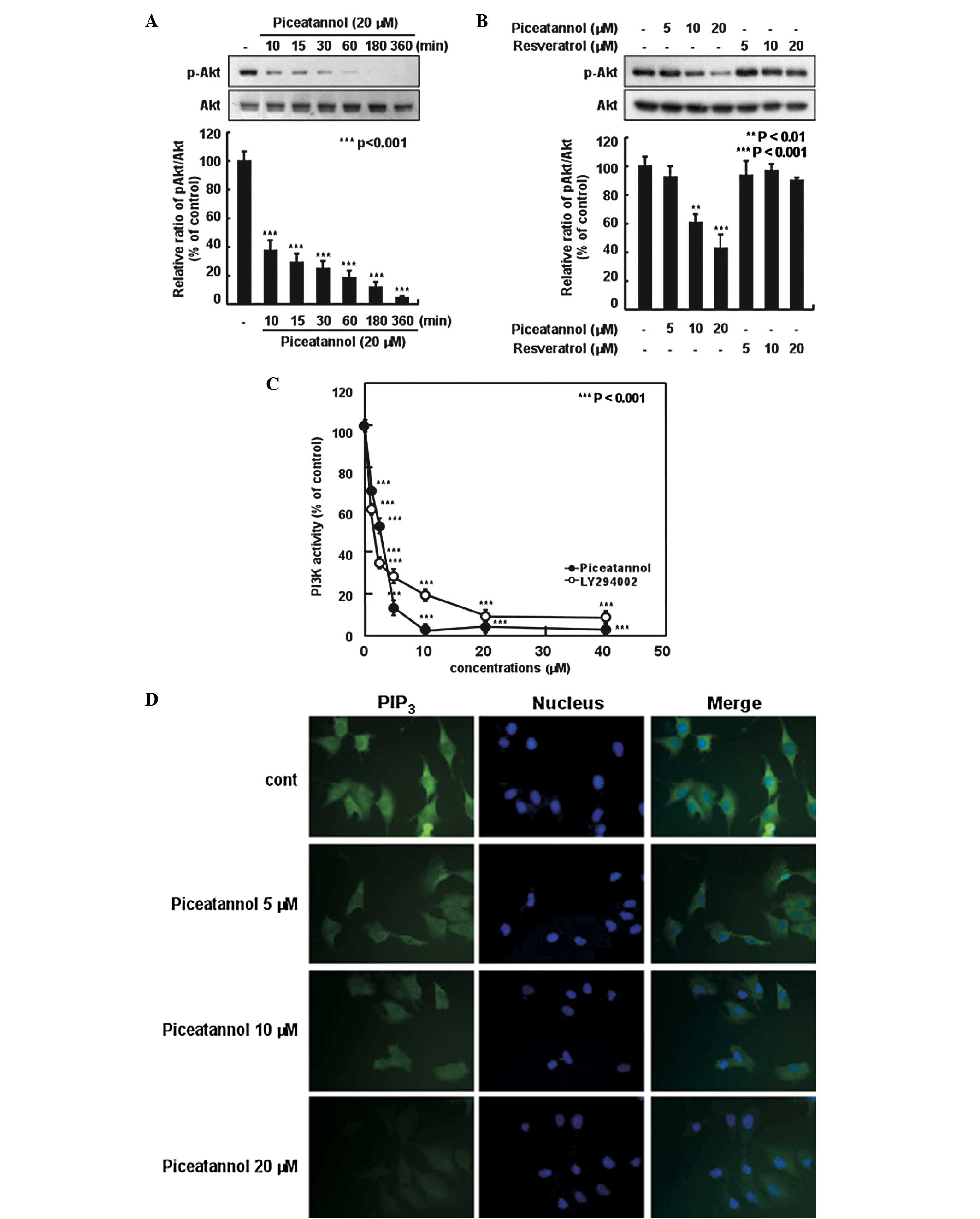 | Figure 3Effects of piceatannol and
resveratrol on the phosphorylation of Akt and phosphatidylinositol
3-kinase (PI3K) activity in H-ras MCF10A cells. (A) After 24 h of
serum starvation, the cells were treated with 20 μM piceatannol for
the indicated periods of time. (B) After 24 h of starvation, the
cells were treated with 5, 10, or 20 μM piceatannol or resveratrol
for 15 min. The levels of phosphorylated and total Akt protein were
determined by western blot analysis, as described in Materials and
methods, using specific antibodies against the corresponding
phosphorylated and total protein. Each experiment was performed in
triplicate. (C) Effects of piceatannol and LY294002 on PI3K
activity in vitro. Active PI3K (80 ng) was pre-incubated
with piceatannol and LY294002 at 1.25, 2.5, 5, 10, 20, or 40 μM for
10 min at 30°C, then incubated with phosphatidylinositol substrate
and [γ-32P]ATP for an additional 10 min at 30°C. The
resulting 32P-labeled phosphatidylinositol phosphate
(PIP) was measured as described in Materials and methods. The
relative level of PI3K activity was quantified densitometrically
using Image J software. Data are the means ± SD of three
independent experiments. (D) An immunofluorescence assay was
performed on the cells for PI3K activity using anti-PIP3
antibody. The immunostained cells (green) were observed under a
confocal microscope. |
PI3K pathway is involved in H-ras-induced
MMP-2 activity, invasion and migration in MCF10A cells
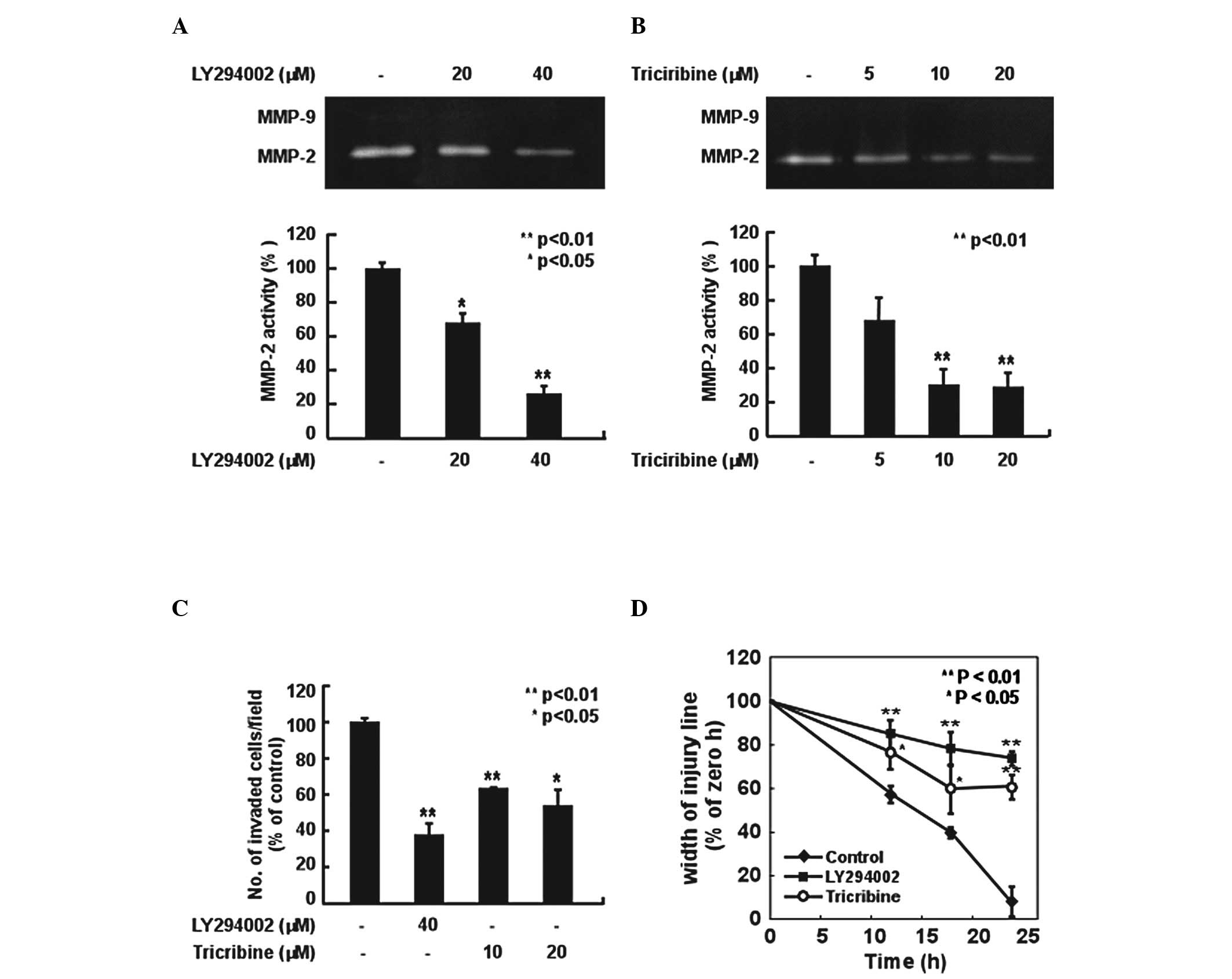
We found that piceatannol suppressed H-ras-induced
Akt phosphorylation through the direct inhibition of PI3K activity.
We then investigated the role of the PI3K/Akt pathway in MMP-2
expression, invasion and migration in H-ras MCF10A cells using a
specific inhibitor of PI3K, LY294002, and a commercial inhibitor of
Akt, triciribine. LY294002 and triciribine inhibited the
H-ras-induced MMP-2 activity in the MCF10A cells (Fig. 4A and B). Moreover, these two
inhibitors also suppressed the H-ras-induced cell invasion and
migration ability of MCF10A cells (Fig. 4C and D). These results indicate
that the PI3K/Akt pathway is involved in the induction of MMP-2
activity, as well as in the invasion and migration ability of H-ras
MCF10A cells.
Piceatannol directly binds to PI3K in
H-ras MCF10A cells
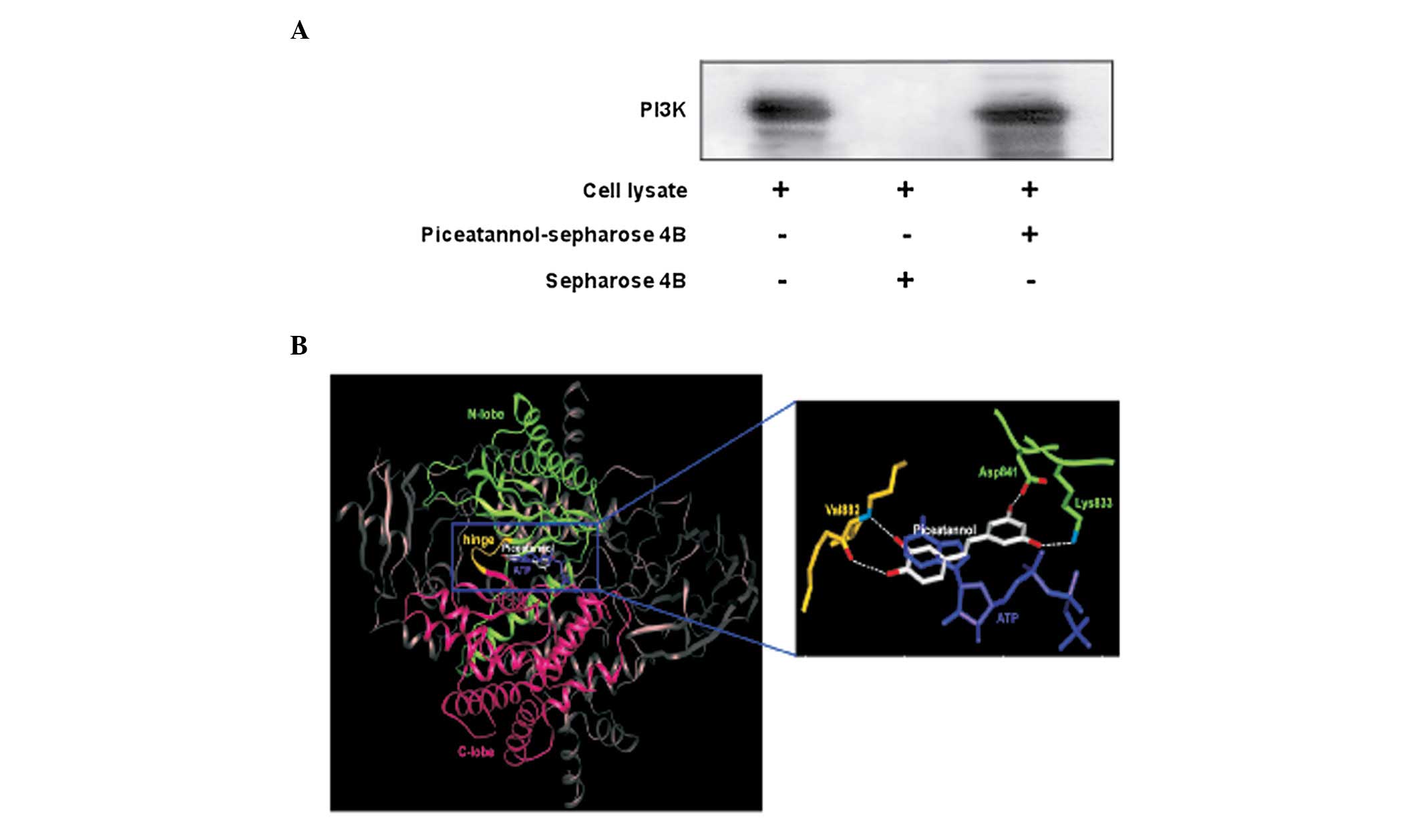
To further investigate the mechanism behind the
inhibitory effect of piceatannol on PI3K activity, we performed
ex vivo pull-down assays. We detected PI3K in the
piceatannol-Sepharose beads (Fig.
5A; lane 3), but not in the control-Sepharose beads (Fig. 5A; lane 2). The input lane
(Fig. 5A; lane 1) indicates that
only cell lysates were loaded as a marker to ensure that the band
detected the PI3K protein itself. These data indicate that
piceatannol inhibits PI3K activity and downstream signaling by
directly binding to PI3K.
Discussion
Several studies have demonstrated that red wine has
anticarcinogenic effects (29,30). Resveratrol is a well-known active
phenolic compound of red wine and has been reported to be a natural
chemopreventive agent against several types of cancer (22–24). Piceatannol is a major metabolite
of resveratrol and is generated by CYP1B1, a cytochrome p450 enzyme
(25). A previous study indicated
that resveratrol is metabolized into a glucuronidated or sulphated
form within 15 min and that a moderate consumption of red wine is
insufficient for resveratrol to reach an effective concentration
(31). The chemical structure of
piceatannol is similar to that of resveratrol, with the addition of
a hydroxyl group at the 3′-position in the B-ring moiety of
resveratrol (Fig. 1A). In the
present study, we demonstrate that piceatannol exerts significantly
greater inhibitory effects against H-ras-induced MMP-2 expression,
invasion and migration than resveratrol in MCF10A cells.
Tumor metastasis, the spread of tumor cells from the
primary site to distant sites, is the main cause of morbidity and
mortality in cancer patients. The metastatic process of quite
complex, involving the dissociation of malignant cells in the
primary tumor, local invasion, angiogenesis and the intravasation
of invading cells into the vasculature or lymphatic systems
(32,33). It is well known that invasive
tumor cells secrete matrix-degrading proteases (34) and collagen, a main component of
the ECM, is an important substrate as it constitutes the structural
scaffolding upon which the other components of the matrix are
assembled (35). The degradation
of the ECM is a crucial step in cell migration and invasion and
MMPs are essential for this step (36). MMPs are a family of zinc-dependent
endopeptidases and are upregulated in the majority of human cancers
(5). In particular, MMP-2 and
MMP-9 are collagenases and are crucial for tumor invasion and
metastasis by degrading collagen IV (6–8). A
previous study demonstrated that MMP-2 is more closely associated
with cell invasion and migration than MMP-9 in H-ras MCF10A human
breast epithelial cells (9). In
this study, we demonstrate that piceatannol, but not resveratrol,
inhibits H-ras-induced MMP-2 expression, significantly suppressing
the migration and invasion of MCF10A cells. Moreover, we
demonstrate that piceatannol suppresses the anchorage-independent
growth of H-ras MCF10A cells.
Ras proteins are associated with the inner face of
the plasma membrane and their activity is regulated by cycling
between the inactive GDP-bound and the active GTP-bound forms
(37). The upregulation of Ras
activity induces all aspects of the malignant phenotype of cancer
cells, including cellular proliferation, transformation, invasion
and metastasis (38). Ras
regulates multiple downstream effectors that stimulate diverse
cytoplasmic signaling activities (39). PI3K is one of the
best-characterized effectors of Ras proteins (16). This kinase phosphorylates
PIP2 to form the second messenger, PIP3,
which activates Akt/PKB, a serine/threonine kinase (17,19). A previous study indicated that the
PI3K/Akt signaling pathway is involved in the H-ras-induced
invasiveness of MCF10A human breast epithelial cells (9). In this study, we found that PI3K/Akt
signaling inhibited the H-ras-induced MMP-2 expression, as well as
the invasion and migration of MCF10A cells. Moreover, piceatannol
exerted a greater inhibitory effect on Akt phosphorylation than
resveratrol.
PI3Ks are heterodimeric lipid kinases composed of
regulatory (p85) and catalytic subunits (p110) that are encoded by
different genes (40,41). Previous studies have demonstrated
that activated Ras protein directly binds with the p110 catalytic
subunit of PI3K and further stimulates PI3K activity (42,43). In our study, we found that
piceatannol markedly inhibited PI3K activity in vitro in the
H-ras MCF10A cells. Moreover, we confirmed that piceatannol
directly binds to PI3K. To investigate the molecular mechanisms
behind the inhibition of PI3K by piceatannol, we carried out a
modeling study using the crystal structure of PI3K in a complex
with ATP or inhibitors (44,45). PI3K consists of four domains: a
Ras-binding domain, a C2 domain, a helical domain and a catalytic
domain. The catalytic domain of the enzyme consists of an N-lobe
and a C-lobe with a fold similar to protein kinases and this
structural similarity is also conserved in the ATP-binding site
that is flanked by these two lobes. Consequently, ATP binds between
these lobes in a manner similar to ATP binding in protein kinases.
The N- and C-lobes are linked through a loop, which is termed the
‘hinge region’. The backbone of this loop interacts with the
adenine moiety of ATP by hydrogen bonds. Considering the
experimental result that piceatannol is an ATP-competitive
inhibitor of PI3K (46), we
docked the compound to the ATP binding site of PI3K (Fig. 5B). The B ring moiety of
piceatannol partially overlapped and was coplanar with the space
occupied by the adenine moiety of ATP, forming hydrogen bonds with
the backbone of the hinge region of PI3K as other ATP-competitive
PI3K inhibitors do (44). The
hydroxyl group at the 5′ position acts as a hydrogen bonding
acceptor in the interaction with the backbone amide group of Val882
and the hydroxyl group at the 4′ position is a hydrogen-bonding
donor in the interaction with the backbone carbonyl group of
Val882. The A ring moiety of piceatannol interacts with the N-lobe
through hydrogen bonds. The hydroxyl group at the 3′ position can
form a hydrogen bond with the side chain of Lys833, which interacts
with the α-phosphate of ATP in the structure of PI3K in complex
with ATP (45). In addition, the
hydroxyl group at the 5′ position can form hydrogen bonds with the
side chains of Asp841. The higher PI3K inhibitory activity of
piceatannol over resveratrol can also be explained by the chemical
structures of the two compounds. Based on previous competition
studies with ATP, resveratrol also acts as a competitive inhibitor
targeting the ATP-binding pocket (47). Therefore, the binding mode of
resveratrol to PI3K would be similar to that of piceatannol due to
the structural similarity between the two compounds. Piceatannol
differs from resveratrol only by the addition of a hydroxyl group
at the 5′ position in the B ring moiety. As a result of the
additional hydrogen bond between the hydroxyl group and the
backbone amide group of Val882, piceatannol is a more tight-binding
inhibitor than resveratrol. Previously, we have reported that
piceatannol inhibits PI3K activity in atherosclerotic models
(46). In this study, using
computer modeling tools, we further elucidated the structural
evidence of piceatannol functioning as a PI3K inhibitor and
clarified the differential effects of piceatannol and resveratrol
based on their chemical structures.
In conclusion, piceatannol inhibits H-ras-induced
MMP-2 expression, invasion, migration and anchorage-independent
growth, whereas resveratrol has relatively minor inhibitory effects
in MCF10A human breast epithelial cells. These inhibitory effects
were mediated mainly through the blocking of the PI3K/Akt pathway.
Piceatannol markedly suppressed PI3K activity by directly binding
to PI3K. Our computer modeling data suggest that piceatannol is a
more tight-binding inhibitor than resveratrol due to the additional
hydrogen bond between the hydroxyl group and the backbone amide
group of Val882 in the ATP binding pocket of PI3K.
Acknowledgements
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(2010-0029233 and 2010-0027204) and World Class University Program
(R31-2008-00-10056-0).
Abbreviations:
|
ECM
|
extracellular matrix
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
MMPs
|
matrix metalloproteinases
|
|
NF-κB
|
nuclear factor-κB
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
PIP3
|
phosphatidylinositol
(3,4,5)-trisphosphate
|
|
PIP2
|
phosphatidylinositol
(4,5)-bisphosphate
|
|
PDK
|
phosphoinositide-dependent kinase
|
|
H-ras MCF10A cells
|
MCF10A human breast epithelial cells
harboring mutated H-ras
|
References
|
1
|
Sporn MB: The war on cancer. Lancet.
347:1377–1381. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fidler IJ: Critical factors in the biology
of human cancer metastasis: twenty-eighth G.H.A. Clowes memorial
award lecture. Cancer Res. 50:6130–6138. 1990.PubMed/NCBI
|
|
3
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis: an imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sato H, Takino T, Okada Y, et al: A matrix
metalloproteinase expressed on the surface of invasive tumour
cells. Nature. 370:61–65. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
McCawley LJ and Matrisian LM: Matrix
metalloproteinases: multifunctional contributors to tumor
progression. Mol Med Today. 6:149–156. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kallakury BV, Karikehalli S, Haholu A,
Sheehan CE, Azumi N and Ross JS: Increased expression of matrix
metalloproteinases 2 and 9 and tissue inhibitors of
metalloproteinases 1 and 2 correlate with poor prognostic variables
in renal cell carcinoma. Clin Cancer Res. 7:3113–3119.
2001.PubMed/NCBI
|
|
7
|
Stetler-Stevenson WG: Type IV collagenases
in tumor invasion and metastasis. Cancer Metastasis Rev. 9:289–303.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tryggvason K, Hoyhtya M and Pyke C: Type
IV collagenases in invasive tumors. Breast Cancer Res Treat.
24:209–218. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shin I, Kim S, Song H, Kim HR and Moon A:
H-Ras-specific activation of Rac-MKK3/6-p38 pathway: its critical
role in invasion and migration of breast epithelial cells. J Biol
Chem. 280:14675–14683. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kiaris H and Spandidos DA: Mutations of
ras genes in human tumours. Int J Oncol. 7:413–421.
1995.
|
|
11
|
Clair T, Miller WR and Cho-Chung YS:
Prognostic significance of the expression of a ras protein with a
molecular weight of 21,000 by human breast cancer. Cancer Res.
47:5290–5293. 1987.PubMed/NCBI
|
|
12
|
Clark GJ and Der CJ: Aberrant function of
the Ras signal transduction pathway in human breast cancer. Breast
Cancer Res Treat. 35:133–144. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Watson DM, Elton RA, Jack WJ, Dixon JM,
Chetty U and Miller WR: The H-ras oncogene product p21 and
prognosis in human breast cancer. Breast Cancer Res Treat.
17:161–169. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moon A, Kim MS, Kim TG, et al: H-ras, but
not N-ras, induces an invasive phenotype in human breast epithelial
cells: a role for MMP-2 in the H-ras-induced invasive phenotype.
Int J Cancer. 85:176–181. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dhillon AS, Meikle S, Yazici Z, Eulitz M
and Kolch W: Regulation of Raf-1 activation and signalling by
dephosphorylation. EMBO J. 21:64–71. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wymann MP, Zvelebil M and Laffargue M:
Phosphoinositide 3-kinase signalling - which way to target? Trends
Pharmacol Sci. 24:366–376. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang F, Lee JT, Navolanic PM, et al:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: a target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Blume-Jensen P and Hunter T: Oncogenic
kinase signalling. Nature. 411:355–365. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sliva D, Rizzo MT and English D:
Phosphatidylinositol 3-kinase and NF-kappaB regulate motility of
invasive MDA-MB-231 human breast cancer cells by the secretion of
urokinase-type plasminogen activator. J Biol Chem. S277:3150–3157.
2002. View Article : Google Scholar
|
|
21
|
Sizemore N, Leung S and Stark GR:
Activation of phosphatidylinositol 3-kinase in response to
interleukin-1 leads to phosphorylation and activation of the
NF-kappaB p65/RelA subunit. Mol Cell Biol. 19:4798–4805.
1999.PubMed/NCBI
|
|
22
|
Estrov Z, Shishodia S, Faderl S, et al:
Resveratrol blocks interleukin-1beta-induced activation of the
nuclear transcription factor NF-kappaB, inhibits proliferation,
causes S-phase arrest, and induces apoptosis of acute myeloid
leukemia cells. Blood. 02:987–995. 2003. View Article : Google Scholar
|
|
23
|
Mgbonyebi OP, Russo J and Russo IH:
Antiproliferative effect of synthetic resveratrol on human breast
epithelial cells. Int J Oncol. 12:865–869. 1998.PubMed/NCBI
|
|
24
|
Hsieh TC and Wu JM: Differential effects
on growth, cell cycle arrest, and induction of apoptosis by
resveratrol in human prostate cancer cell lines. Exp Cell Res.
249:109–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Potter GA, Patterson LH, Wanogho E, et al:
The cancer preventative agent resveratrol is converted to the
anticancer agent piceatannol by the cytochrome P450 enzyme CYP1B1.
Br J Cancer. 86:774–778. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ashikawa K, Majumdar S, Banerjee S, Bharti
AC, Shishodia S and Aggarwal BB: Piceatannol inhibits TNF-induced
NF-kappaB activation and NF-kappaB-mediated gene expression through
suppression of IkappaBalpha kinase and p65 phosphorylation. J
Immunol. 169:6490–6497. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roupe KA, Remsberg CM, Yanez JA and Davies
NM: Pharmacometrics of stilbenes: seguing towards the clinic. Curr
Clin Pharmacol. 1:81–101. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar
|
|
29
|
Briviba K, Pan L and Rechkemmer G: Red
wine polyphenols inhibit the growth of colon carcinoma cells and
modulate the activation pattern of mitogen-activated protein
kinases. J Nutr. 132:2814–2818. 2002.PubMed/NCBI
|
|
30
|
Hakimuddin F, Paliyath G and Meckling K:
Treatment of mcf-7 breast cancer cells with a red grape wine
polyphenol fraction results in disruption of calcium homeostasis
and cell cycle arrest causing selective cytotoxicity. J Agric Food
Chem. 54:7912–7923. 2006. View Article : Google Scholar
|
|
31
|
Saiko P, Szakmary A, Jaeger W and Szekeres
T: Resveratrol and its analogs: Defense against cancer, coronary
disease and neurodegenerative maladies or just a fad? Mutat Res.
658:68–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Steeg PS: Tumor metastasis: mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liotta LA, Thorgeirsson UP and Garbisa S:
Role of collagenases in tumor cell invasion. Cancer Metastasis Rev.
1:277–288. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liotta LA: Tumor invasion and
metastases-role of the extracellular matrix: Rhoads Memorial Award
lecture. Cancer Res. 46:1–7. 1986.PubMed/NCBI
|
|
36
|
Geiger TR and Peeper DS: Metastasis
mechanisms. Biochim Biophys Acta. 1796:293–308. 2009.PubMed/NCBI
|
|
37
|
Bourne HR, Sanders DA and McCormick F: The
GTPase superfamily: conserved structure and molecular mechanism.
Nature. 349:117–127. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Malumbres M and Barbacid M: To cycle or
not to cycle: a critical decision in cancer. Nat Rev Cancer.
1:222–231. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Feig LA and Buchsbaum RJ: Cell signaling:
life or death decisions of ras proteins. Curr Biol. 12:R259–R261.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Inukai K, Funaki M, Ogihara T, et al:
p85alpha gene generates three isoforms of regulatory subunit for
phosphatidylinositol 3-kinase (PI 3-Kinase), p50alpha, p55alpha,
and p85alpha, with different PI 3-kinase activity elevating
responses to insulin. J Biol Chem. 272:7873–7882. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ueki K, Algenstaedt P, Mauvais-Jarvis F
and Kahn CR: Positive and negative regulation of phosphoinositide
3-kinase-dependent signaling pathways by three different gene
products of the p85alpha regulatory subunit. Mol Cell Biol.
20:8035–8046. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rodriguez-Viciana P, Warne PH, Dhand R, et
al: Phospha-tidylinositol-3-OH kinase as a direct target of Ras.
Nature. 370:527–532. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Walker EH, Pacold ME, Perisic O, et al:
Structural determinants of phosphoinositide 3-kinase inhibition by
wortmannin, LY294002, quercetin, myricetin, and staurosporine. Mol
Cell. 6:909–919. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Walker EH, Perisic O, Ried C, Stephens L
and Williams RL: Structural insights into phosphoinositide 3-kinase
catalysis and signalling. Nature. 402:313–320. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Choi KH, Kim JE, Song NR, et al:
Phosphoinositide-3-kinase is a novel target of piceatannol for
inhibiting PDGF-BB-induced proliferation and migration in human
aortic smooth muscle cells. Cardiovasc Res. 85:836–844. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Frojdo S, Cozzone D, Vidal H and Pirola L:
Resveratrol is a class IA phosphoinositide 3-kinase inhibitor.
Biochem J. 406:511–518. 2007. View Article : Google Scholar : PubMed/NCBI
|