Introduction
Acute pancreatitis (AP) is an inflammatory disease
with high morbidity and mortality; however, the exact mechanisms
involved are not yet fully understood. Although the majority of
patients suffer mild or edematous AP with a low complication and
mortality rate, 15–20% of patients develop severe AP (SAP) with a
high mortality rate as high as 30% (1,2).
The pathogenesis of AP is not clear; however, inflammatory
cytokines, leukocytic infiltration, the activation of nuclear
factor-κB (NF-κB) and oxidative stress are important factors
(3,4). The inflammatory response is
responsible for the morbidity and mortality associated with AP
(5). During AP, particularly SAP,
pancreatic acinar cells are the primary source of various
pro-inflammatory cytokines, such as tumor necrosis factor-α
(TNF-α), interleukin (IL)-1β and IL-6, which are directly
responsible for aggravating the inflammatory response (5–9).
These cytokines are upregulated from the initial phase of AP and
directly correlate with many deleterious events locally in the
pancreas and in distant organs (10,11). Among the multitude of inflammatory
molecules, a key regulator of cytokine induction is the pleiotropic
transcription factor, NF-κB (12). NF-κB is capable of regulating a
variety of inflammatory mediators involved in AP, including TNF-α,
IL-1β and IL-6. In most silent cells, NF-κB is kept inactive in the
cytoplasm through sequestration in complexes with the inhibitor of
NF-κB (IκB) proteins, such as inhibitory κBα (IκBα) and inhibitory
κBβ (IκBβ) and it can be activated by the stimulation of nuclear
translocation (9,13). The activation of NF-κB can promote
the gene expression of TNF-α, IL-1β and IL-6, and plays a critical
role in the initiation and perpetuation of AP (14). Movever, it has been suggested that
oxidative stress plays a significant role in the pathogenesis of AP
(15,16).
The saponin, astragaloside IV (AS-IV; a
3-O-β-D-xylopyranosyl-6-O-β-D-glucopyranosylcycloastragenol), which
is purified from the known Chinese medical herb, Astragalus
membranaceus (Fisch.) Bge. is one of the major and active
components of Αstragalus membranaceus, which has been shown
to have comprehensive pharmacological function (17,18). Diverse pharmacological activities
have been found to be exerted by AS-IV such as anti-inflammatory,
anti-oxidation, anti-infarction and antinociception (19–21). Previous studies have demonstrated
the anti-inflammatory effects of AS-IV in a murine model of chronic
asthma, in rats with focal cerebral ischemia/reperfusion injury, as
well as in vitro (19,21,22). In addition, AS-IV has been shown
to exert antioxidant effects through the reduction of free
radicals, the inhibition of lipid peroxidation and the elevation of
antioxidant enzymes (23).
To the best of our knowledge, to date, the
protective effects of AS-IV on AP have not yet been investigated.
Since the activation of NF-κB and oxidative stress are the major
factors accounting for the pathogenesis of AP, we hypothesized that
AS-IV may contribute to the prevention of AP progression. The aim
of the present study was to investigate the protective effects of
AS-IV in a rat model of AP. Our results revealed that AS-IV
prevented the aggravation of AP by inhibiting the activition of
NF-κB and counteracting oxidative stress, which suggests that AS-IV
may be effective for the clinical therapy/prevention of AP.
Materials and methods
Ethics statement
All the animal-related procedures were approved by
the Animal Care and Use Committee of the Shanghai Tenth People’s
Hospital, Tongji University, Shanghai, China (permit no.
2011-RES1). This study was also approved by the Science and
Technology Commission of Shanghai Municipality (ID: SYXK
2007-0006).
Animals and materials
Male Sprague-Dawley rats weighing 250±30 g were
purchased from the Shanghai SLAC Laboratory Animal Co., Ltd.
(Shanghai, China). The animals were maintained under 12 h
light-dark cycles at 22°C, provided with water ad libitum,
fed standard laboratory chow and allowed to acclimatize for 1 week.
The environment was maintained at a relative humidity of 30–70%.
Purified AS-IV (>98%) was purchased from Shanghai Tauto Biotech
Co., Ltd. (Shanghai, China). Sodium taurocholate (NaTc), L-arginine
(L-Arg), dimethyl sulfoxide (DMSO) and hematoxylin and eosin
(H&E) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Antibodies against NF-κB p65, manganese superoxide dismutase
(SOD1), cuprum/zinc superoxide dismutase (SOD2), β-actin and
lamin-A were purchased from Abcam (Hong Kong, China).
Peroxidase-conjugated secondary antibody was purchased from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Antibodies against
IκBα and IκBβ were purchased from Cell Signaling Technology (CST;
Shanghai, China).
Experimental design
Before the experiment was initiated, the rats were
fasted overnight with continued access to water. AS-IV, NaTc and
L-Arg were dissolved in the vehicle (2% DMSO). NaTc-induced
experimental AP was induced in the rats by injecting 4% NaTc (0.1
ml/100 g) in the retrograde direction of the biliopancreatic duct.
The L-Arg-induced experimental AP was induced in the rats by 2
intraperitoneal injections of 20% L-Arg (3 mg/kg), with an interval
of 1 h between the injections. In order to determine the optimal
dose of AS-IV for the prevention of AP, we performed a preliminary
experiment. A total of 36 rats were randomly divided into 9 groups
(n=4 in each group) as follows: group 1, normal control; group 2,
NaTc + vehicle-treated; groups 3–5, NaTc + AS-IV-treated (12.5, 25
and 50 mg/kg, respectively); group 6, L-Arg + vehicle-treated; and
groups 7–9, L-Arg + AS-IV-treated (12.5, 25 and 50 mg/kg,
respectively). The rats in the normal control group were injected
with the vehicle intraperitoneally instead of 4% NaTc or 20% L-Arg.
AS-IV, L-Arg and the vehicle (DMSO) were administered 2 h prior to
the induction of AP. All the rats were sacrificed by taking blood
from the heart 24 h after the induction of AP, a time point at
which pancreatic damage had already been induced. The effects of
AS-IV on AP were assessed by determining the serum amylase level
and pancreatic staining with H&E, to obtain an optimal dose.
Accordingly, the optimal dose of AS-IV (50 mg/kg) was used for the
next series of experiments.
Subsequently, 90 rats were randomly divided into 5
groups (n=18 in each group) as follows: group 1, normal control;
group 2, NaTc + vehicle-treated; group 3, NaTc + AS-IV-treated;
group 4, L-Arg + vehicle-treated; and group 5, L-Arg +
AS-IV-treated. The induction of AP and the administration of AS-IV
or the vehicle were carried out in a similar manner as in the
preliminary experiment. The rats were sacrificed by taking blood
from the heart at 12, 24 and 48 h after the induction of AP, 6 rats
at each time point in each group. Blood samples were obtained to
determine the serum amylase, lipase and cytokine levels. A portion
of the pancreas was rapidly removed from each rat and fixed in 4%
neutral paraformaldehyde solution for morphological examination.
The remaining portion of each pancreas was stored in liquid
nitrogen for further analysis.
Isolation of pancreatic acinar cells from
rats
Pancreatic acinar cells were isolated from the rats
using a collagenase digestion procedure as previously described
(24). The isolated acinar cells
were incubated at 37°C under humidified conditions of 95% air and
5% CO2 in Dulbecco’s modified Eagle’s medium/Ham’s F12
Medium (DMEM/F12) containing 10% fetal bovine serum (FBS) and 1%
penicillin-streptomycin (all from Gibco-BRL, Grand Island, NY, USA)
with or without NaTc (3,750 nmol/l)/L-Arg (40 μmol/l) and
AS-IV at different doses (20, 40, 80 and 160 μmol/l). At 12
h following treatment with NaTc/L-Arg, the pancreatic acinar cells
were used to carry out a series of experiments, including Cell
Titer-Glo luminescent cell viability assa, cell counting kit-8
(CCK-8) assay and western blot analysis.
Histological examination
The pancreatic tissue samples were fixed in 4%
neutral paraformaldehyde solution for 24 h, dehydrated through a
graduated ethanol series, embedded in paraffin blocks and cut into
5-μm-thick sections. The pancreatic sections were dewaxed in
xylene, hydrated through an upgraded ethanol series and stained
with H&E. Morphological changes were observed under a light
microscope (DMI6000B; Leica, Wetzlar, Germany). Ten microscopic
fields were randomly selected to be observed in each paraffin
section.
Serum amylase, lipase and
pro-inflammatory cytokine assay
Blood samples of each rat were maintained at 4°C for
24 h prior to centrifugation at 3,000 × g for 15 min at 4°C, and
serum-stored at −80°C. The serum activities of amylase and lipase
were measured by enzyme dynamics chemistry using commercial kits
according to the manufacturer’s instructions in a Roche/Hitachi
modular analytics system (Roche, Mannheim, Germany). Serum TNF-α,
IL-1β and IL-6 levels were measured by enzyme-linked immunosorbent
assay (ELISA) using a commercial kit (Quantikine; R&D Systems,
Minneapoils, MN, USA).
Meaurement of myeloperoxidase (MPO)
activity
Neutrophil sequestration in the pancreas was
quantified by measuring tissue MPO activity according to a
previously described method (25). Pancreatic tissue samples were
homogenized in 20 mM phosphate buffer (pH 7.4) and centrifuged
(12,000 × g for 10 min at 4°C). The pellet was resuspended in 50 mM
phosphate buffer (pH 6), containing 0.5% hexade
cyltrimethylammonium bromide (HETAB). The suspension was subjected
to 4 cycles of freezing and thawing and was further disrupted by
sonication for 1 min. The sample was then centrifuged (12,000 × g
for 5 min at 4°C). Aliquots of supernatant were added to the
reaction mixture containing 0.167 mg/ml of o-dianisdine
dihydrochloride and 0.0005% H2O2 solution,
which were prepared in 50 mM of phosphate buffer. The change in
absorbance at 450 nm was then measured for 5 min using a Beckman
spectrophotometer (DU640B; Beckman Coulter, Brea, CA, USA). One
unit of MPO activity was defined as that degrading 1 mmol of
peroxide/min at 25°C. The activity was expressed as unit/milligram
of tissue.
Reverse transcription-quantitative
(real-time) polymerase chain reaction (RT-qPCR)
The mRNA transcripts were analyzed by RT-qPCR of the
pancreatic tissue. Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) following the manufacturer’s
instructions and was subjected to reverse transcription using the
Prime-Script RT reagent kit (Takara, Shiga, Japan). SYBR-Green
quantitative PCR was performed using a 7900HT Fast Real-time PCR
system (Applied Biosystems, Foster City, CA, USA) according to the
instructions provided with SYBR Premix EX Taq (Takara). The
relative mRNA levels were normalized to the mRNA levels of
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and the fold
change of each mRNA was calculated using the comparative CT
(2−ΔΔCT) method. Primer sequences for these biomarkers
were as follows: rat TNF-α forward, 5′-TGATCCGAGATGTGGAACTG-3′ and
reverse, 5′-GGCCATGGAACTGATGAGAG-3′; rat IL-1β forward,
5′-CATCCAGCTTCAAATCTCAC-3′ and reverse, 5′-ACCACTTGTTGGCTTATGTT-3′;
rat IL-6 forward, 5′-TCTCTCCGCAAGAGACTTCC-3′ and reverse,
5′-TCTTGGTCCTTAGCCACTCC-3′; rat IκBα forward,
5′-CATGAAGAGAAGACACTGACCA TGGAA-3′ and reverse,
5′-TGGATAGAGGCTAAGTGTAGACACG-3′; rat IκBβ forward,
5′-GCAGGAAGATGTGGTGGA-3′ and reverse, 5′-CTGGCTCATATGGTTTCC-3′; rat
SOD1 forward, 5′-CACAAGCACAGCCTCCCTGA-3′ and reverse,
5′-GCAATCTGTAAGCGACCTTG-3′; and rat SOD2 forward,
5′-TTCGAGCAGAAGGCAAGCGG-3′ and reverse,
5′-GTACGGCCAATGATGGAATG-3′.
Western blot analysis
For western blot analysis, the rat pancreas was
rapidly ground in liquid nitrogen, as previously described
(26). The resulting powder or
isolated acinar cells were lysed in nuclear extract following the
manufacturer’s instructions for the preparation of nuclear and
cytoplasmic proteins (Pierce, Rockford, IL, USA). On the other
hand, the resulting powder or isolated acinar cells were
reconstituted in ice-cold RIPA buffer containing 1 mmol/l
phenylmethanesulfonyl fluoride (PMSF, 1 mM) and a cocktail of
protease inhibitors (1:100 dilution; Sigma-Aldrich). The samples
were centrifuged at 4°C for 15 min at 10,000 × g and supernatants
were recovered. The concentrations of nuclear, cytoplasmic and
total proteins were determined using the BCA method (Pierce). A 50
μg portion of protein or equal proportion of concentrated
supernatant was subjected to sodium dodecyl sulfate/polyacrylamide
gel electrophoresis (SDS-PAGE; Bio-Rad, Hercules, CA, USA), and
then blotted following standard methods. Non-specific binding to
the membrane was blocked by 5% (w/v) dry non-fat milk in
Tris-buffered saline/0.05% Tween-20 (TBST) at room temperature for
1 h in a covered container. The blots were incubated overnight at
4°C with rabbit polyclonal anti-NF-κB p65 antibody (1:400 dilution;
Abcam), rabbit polyclonal anti-IκBα antibody (1:200 dilution; CST),
rabbit polyclonal anti-IκB β antibody (1:200 dilution; CST), rabbit
polyclonal anti-SOD1 antibody (1:500 dilution; Abcam), rabbit
polyclonal anti-SOD2 antibody (1:2,000 dilution; Abcam), rabbit
polyclonal anti-lamin A antibody (1:500 dilution; Abcam) and mouse
monoclonal anti-β-actin (1:1,000 dilution; Abcam) diluted in 5%
BSA. Lamin A and β-actin were used as the internal reference for
nuclear proteins and cytoplasmic proteins, respectively. The
membranes were washed with TBST and incubated with a secondary
rabbit anti-mouse IhG horseradish peroxidase (HRP) antibody
(1:2,000; Santa Cruz Biotechnology, Inc.) or mouse anti-rabbit
IgG-HRP antibody (1:2,000 dilution; Santa Cruz Biotechnology, Inc.)
diluted in 5% (w/v) dry non-fat milk in TBST for 1 h at room
temperature. Finally, the membranes were washed with TBST,
developed using the ECL detection system (Santa Cruz Biotechnology,
Inc.), quickly dried and exposed to ECL film.
Immunohistochemistry
Formalin-fixed, paraffin-embedded samples were cut
at a thickness of 5 μm. Each tissue section was
deparaffinized and rehydrated with graded ethanol. For antigen
retrieval, the slides were boiled in EDTA (1 mM, pH 8.0) for 15 min
in a microwave oven. Endogenous peroxidase activity was blocked
with a 0.3% hydrogen peroxide solution for 10 min at room
temperature. After rinsing with phosphate-buffered saline (PBS),
the slides were incubated overnight at 4°C with the antibody
against NF-κB p65 (1:100 dilution), washed with 3 times with 0.02%
Tween-20 in PBS for 10 min each time and incubated with
biotinylated secondary antibody diluted at 1:500 for 40 min. The
antibody binding were detected with an Envision Detection kit,
peroxidase/DAB, rabbit/mouse (GeneTech, Ltd., Shanghai, China). The
sections were counterstained with hematoxylin. In general, NF-κB
p65 was stained in the cytoplasm, and the translocation from the
cytoplasm to the nucleus indicated the activation of NF-κB p65. The
sections were observed under a light microscope (DMI6000B; Leica)
at a magnification of x400.
Quantification of cell viability
Pancreatic acinar cell necrosis was determined by
measuring the adenosine triphosphate (ATP) levels using the Cell
Titer-Glo Luminescent Cell Viability Assay kit (Promega, Madison,
WI, USA) as previously described (27). Cell proliferation was determined
by CCK-8 assay using a CCK-8 kit (Dojindo, Kumamoto, Japan),
according to manufacturer’s instructions. The viability of the
isolated rat pancreatic acinar cells was analyzed with the above 2
methods.
Statistical analysis
All data are presented as the means ± SD.
Statistical analysis was performed using the Student’s t-test for
comparisons of 2 groups, and one way ANOVA was used to analyze the
differences among multiple groups. A value of P<0.05 was
considered to indicate a statistically significant difference. All
analyses were conducted using statistical analysis software (SPSS
17.0).
Results
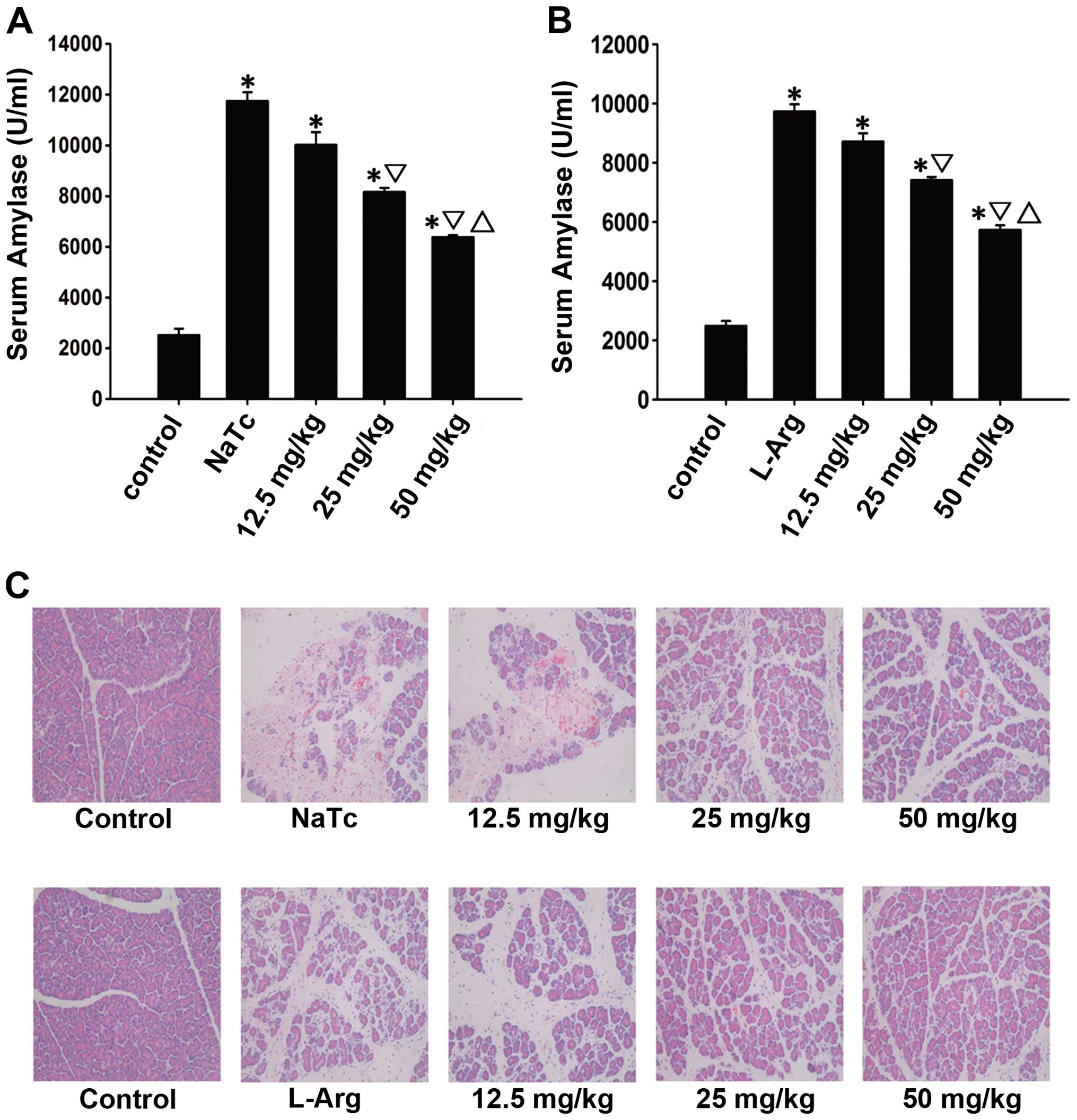
Prelilminary analysis
The optimal effective dose of AS-IV was evaluated
based on the serum amylase level and pancreatic H&E staining.
In the prelilminary experiment (Fig.
1), the highest dose (50 mg/kg) used reduced pancreatic damage
more prominently compared with the moderate dose (25 mg/kg) and the
low dose (12.5 mg/kg) used at the time point of 24 h following the
induction of AP (P<0.05). Therefore, the dose of 50 mg/kg was
selected as the optimal dose for the following experiments.
AS-IV alleviates the histopathological
alterations of the pancreas
To determine the effects of AS-IV on the development
and severity of AP, the rats were pre-treated with AS-IV (50 mg/kg)
or the vehicle (DMSO) prior to the induction of AP. In the normal
control group, the pancreatic histological characteristics were
typical of a nomal architecture (Fig.
2A, F and K). A histological examination of the rat pancreas
revealed that the rats in the NaTc + vehicle-treated group
(Fig. 2B, G and L) and L-Arg +
vehicle-treated group (Fig. 2D, I and
N) showed massive edema, hemorrhaging, inflammatory cell
infiltration and necrosis. Pre-treatment with AS-IV resulted in
lower interstitial edema, less inflammatory cell infiltration and
alleviatived acinar cell necrosis at 3 time points in the NaTc +
AS-IV-treated group (Fig. 2C, H and
M) and L-Arg + AS-IV-treated group (Fig. 2E, J and O). Therefore,
pre-treatment with AS-IV observably reduced both the NaTc- and
L-Arg-induced histological characteristics of pancreatic
injury.
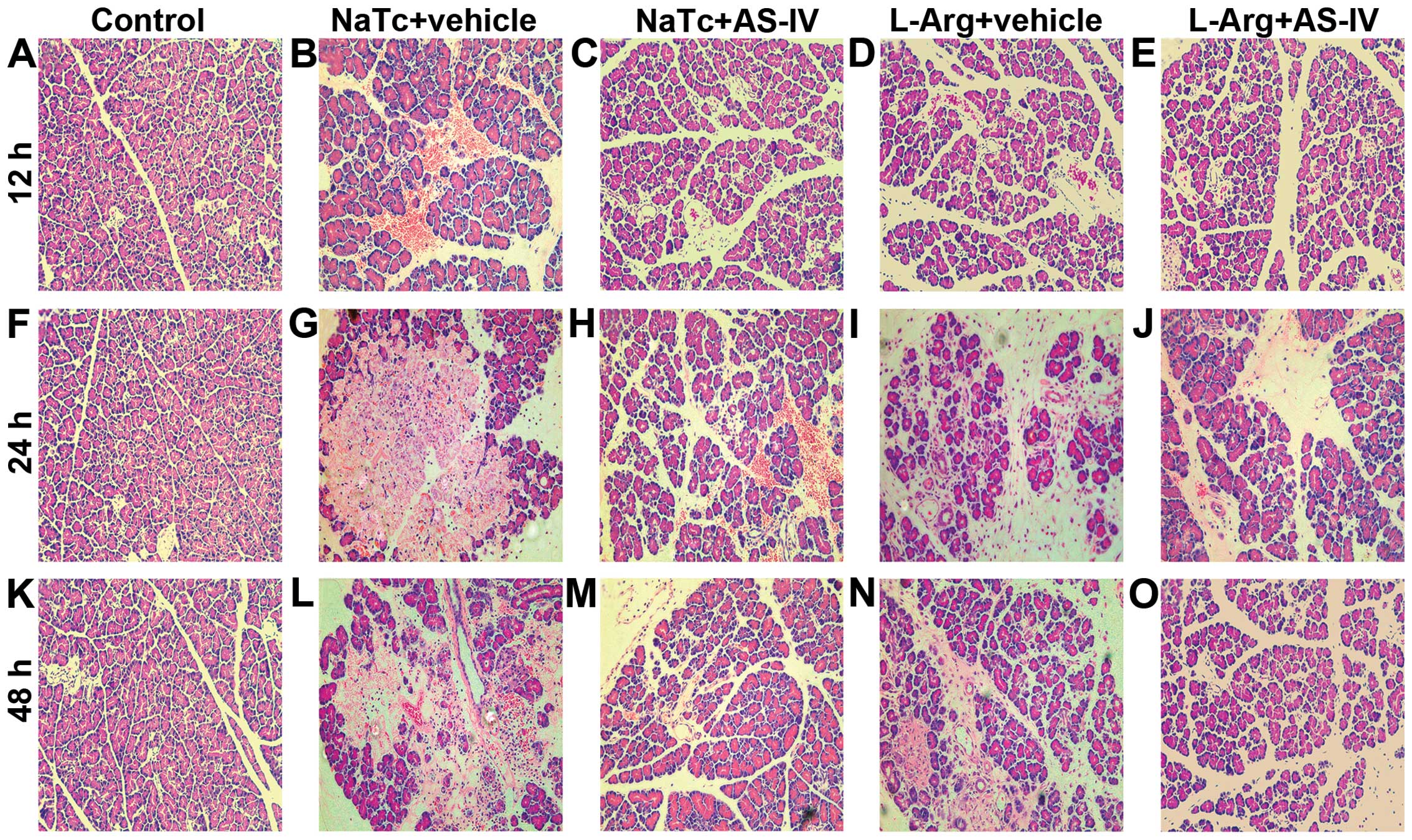 | Figure 2Pancreatic histological alterations.
Rats were sacrificed at 12, 24 and 48 h after the induction of AP,
6 rats at each time point in each group. Representative light
micrographs from (A, F and K) the normal control group, (B, G and
L) NaTc + vehicle-treated group, (C, H and M) NaTc + AS-IV-treated
group, (D, I and N) L-Arg + vehicle-treated group, and (E, J and O)
L-Arg + AS-IV-treated group are presented (magnification, x200). By
comparison, pre-treatment with AS-IV significantly decreased
interstitial edema, inflammatory cell infiltration and acinar cell
necrosis. AS-IV, astragaloside IV; NaTc, sodium taurocholate;
L-Arg, L-arginine; AP, acute pancreatitis. |
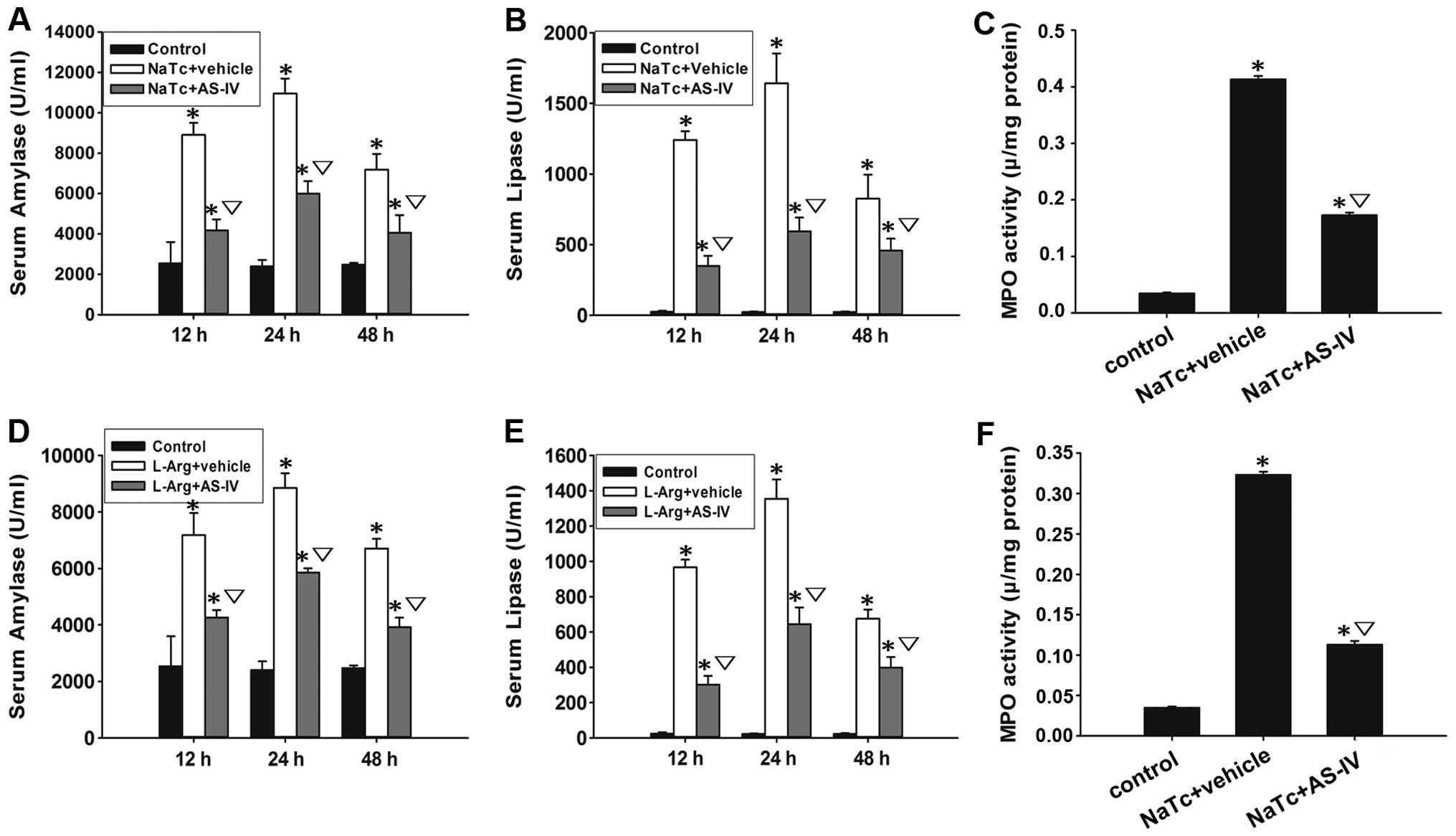
AS-IV reduces serum amylase and lipase
levels
Serum amylase and lipase are most commonly regarded
as biochemical indicators of AP (28). Thus, we assessed the development
of AP by measuring the serum amylase (Fig. 3A and D) and lipase levels
(Fig. 3B and E). Compared to the
normal control group, the serum amylase and lipase levels of the
other 4 groups were increased significantly (P<0.05). In
comparison to the NaTc/L-Arg + vehicle-treated groups,
pre-treatment with AS-IV significantly reduced the elevation of the
serum amylase and lipase levels at 3 time points (P<0.05).
AS-IV decreases the secretion of
pro-inflammatory cytokines and MPO activity
Neutrophil sequestration in the pancreas was
determined by measuring MPO activity in the pancreatic tissue at
the time point of 24 h after the induction of AP. Pre-treatment
with AS-IV reduced the NaTc- or L-Arg-induced activity of MPO in
the pancreatic tissue (Fig. 3C and
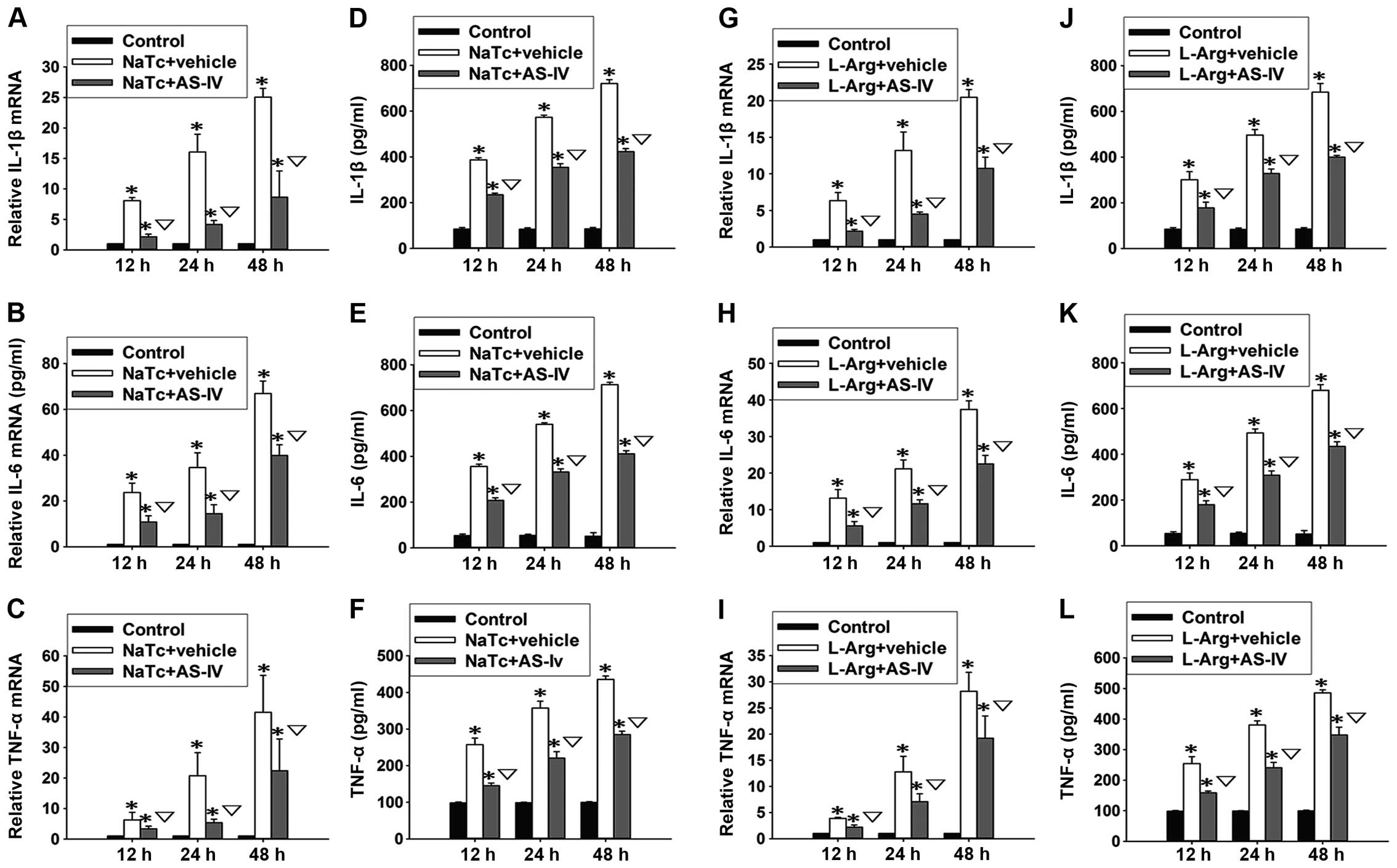
F). Based on the results of ELISA and RT-qPCR, we found that
both the mRNA expression levels (Fig.
4A-C and G-I) and the serum levels (Fig. 4D-F and J-L) of IL-1β, IL-6 and
TNF-α in the normal control group were significantly lower than
those of the other 4 groups at 3 time points (P<0.05). Compared
with the NaTc + vehicle-treated group and the L-Arg +
vehicle-treated group, pre-treatment with AS-IV significantly
reduced the elevation of the serum levels and mRNA expression
levels of IL-1β, IL-6 and TNF-α (P<0.05).
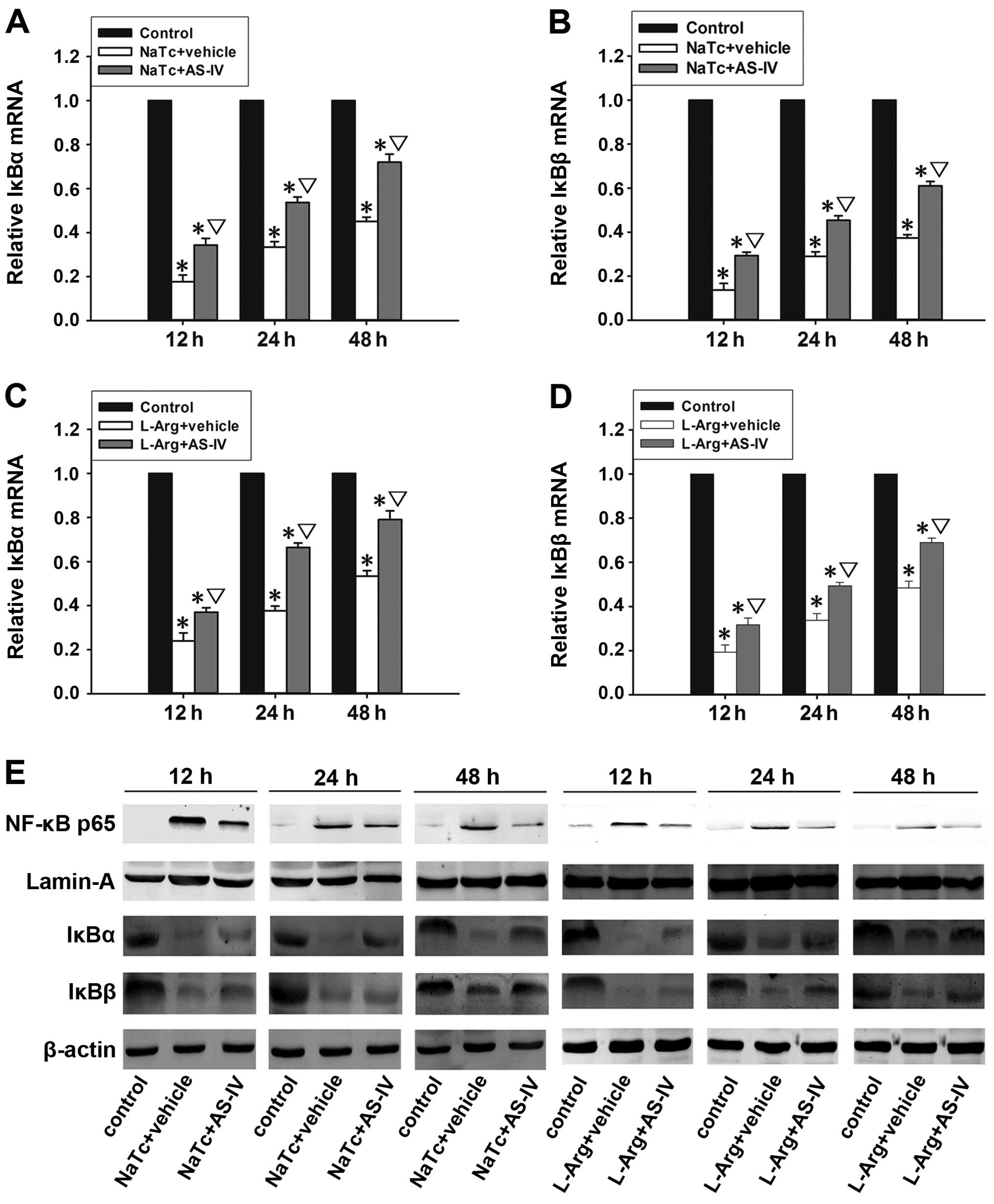
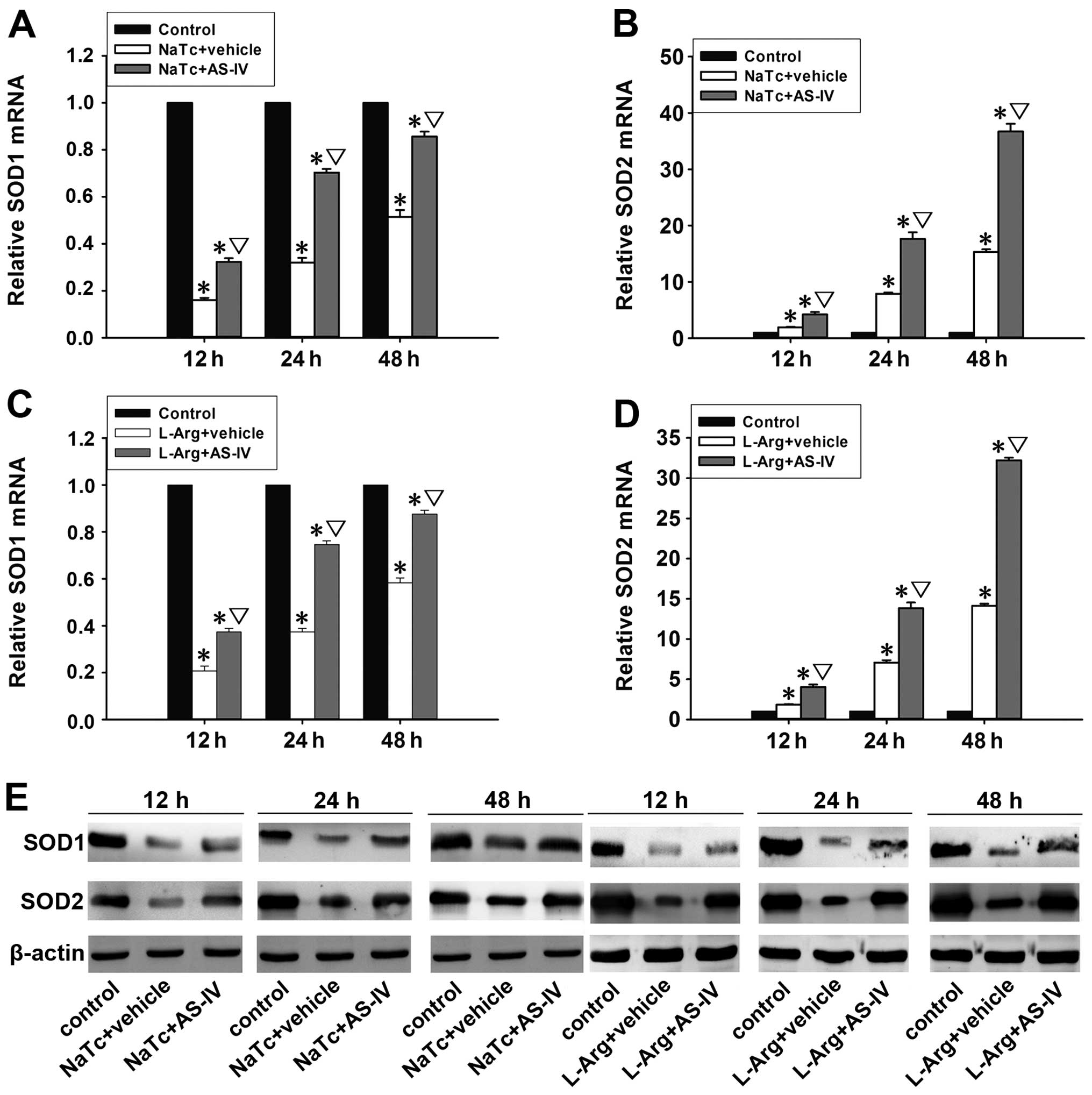
AS-IV inhibits the nuclear translocation
of NF-κB p65
The activation of NF-κB plays an important role in
the induction of pro-inflammatory cytokines (12). Generally, inactivated NF-κB is
sequestered by the inhibitor IκB in the cytoplasm, and it can be
activated by stimulation with nuclear translocation (13). The mRNA expression levels of IκBα
(Fig. 5A and C) and IκBβ
(Fig. 5B and D) in the normal
control group were significantly higher than those of the other 4
groups at 3 time points (P<0.05). In the NaTc + AS-IV-treated
group and L-Arg + AS-IV-treated group, the mRNA expression levels
of IκBα and IκBβ were significantly higher than those in the NaTc +
vehicle-treated group and L-Arg + vehicle-treated group
(P<0.05). In order to ascertain whether AS-IV inhibits the
activation of NF-κB, we examined the protein expression levels of
NF-κB p65 in the nucleus and the levels of IκBα and IκBβ in the
cytoplasm by western blot analysis at 3 time points in each group
(Fig. 5E). In the NaTc +
vehicle-treated group and L-Arg + vehicle-treated group, the
protein expression levels of IκBα and IκBβ were markedly reduced;
however, the level of nuclear NF-κB p65 was significantly increased
at the 3 time points, particularly at the time point of 12 h
following the induction of AP. Compared with NaTc/L-Arg +
vehicle-treated group at the 3 time points, the administration of
AS-IV markedly decreased the degradation of IκBα and IκBβ, as well
as the nuclear protein expression level of NF-κB p65. Subsequently,
we further measured the level of NF-κB p65 in the nucleus by
immunohistochemistry (Fig. 6). In
the normal control group (Fig. 6A, F
and K), immunoreactivity for NF-κB p65 protein was hardly
observed in the nucleus. However, the NaTc + vehicle-treated group
(Fig. 6B, G and L) and L-Arg +
vehicle-treated group (Fig. 6D, I and
N) exhibited a strong positive expression of NF-κB p65 in the
nucleus, particularly at the time point of 12 h following the
induction of AP. As expected, in the NaTc + AS-IV-treated group
(Fig. 6C, H and M) and L-Arg +
AS-IV-treated group (Fig. 6E, J and
O), the positive expression of NF-κB p65 in the nucleus was
weaker than that in the NaTc/L-Arg + vehicle-treated groups at the
3 time points. The results from western blot analysis and
immunohistochemistry confirmed that AS-IV inhibited the nuclear
translocation of NF-κB p65.
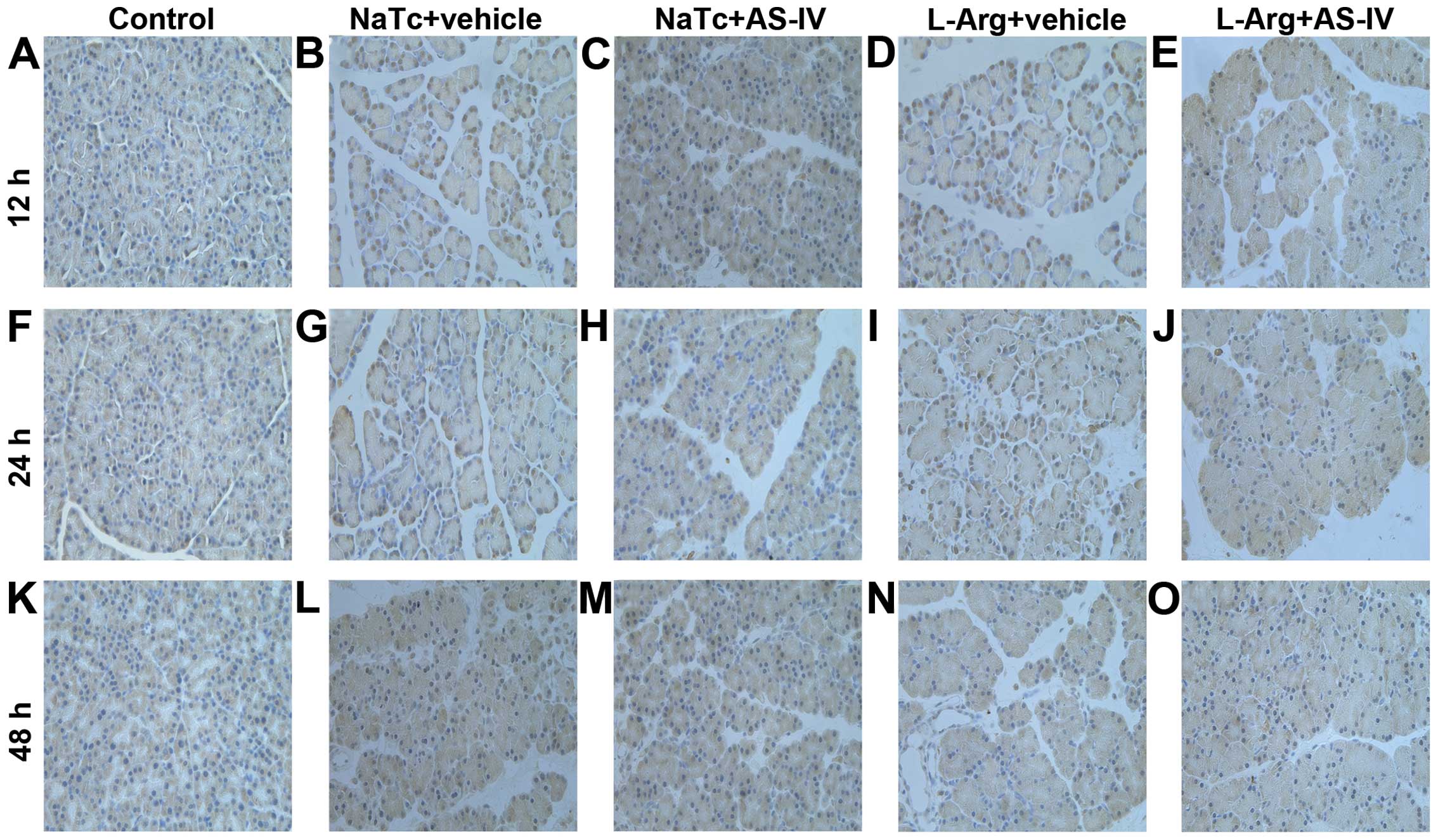 | Figure 6Immunohistochemical analysis of NF-κB
expression. (A, F and K) Normal control group, (B, G and L) NaTc +
vehicle-treated group, (C, H and M) NaTc + AS-IV-treated group, (D,
I and N) L-Arg + vehicle-treated group, and (E, J and O) L-Arg +
AS-IV-treated group (original magnification, x400). Pre-treatment
with AS-IV significantly decreased the staining intensity of NF-κB
in the nucleus, particularly at the time point of 12 h after the
induction of AP. Results are representative of 3 independent
experiments. AS-IV, astragaloside IV; NaTc, sodium taurocholate;
L-Arg, L-arginine; AP, acute pancreatitis; NF-κB, nuclear
factor-κB. |
Antioxidant effects of AS-IV on AP
Oxidative stress imposed by reactive oxygen species
(ROS) is considered one of the forerunners of AP (29). The antioxidant enzyme, SOD, plays
an improtant role in protecting cells against the generation of ROS
(30). The mRNA expression levels
of SOD1 (Fig. 7A and C) and SOD2
(Fig. 7B and D) were detected by
RT-qPCR. In the NaTc + AS-IV-treated group and L-Arg +
AS-IV-treated group, the mRNA expression levels of SOD1 and SOD2
were significantly increased in comparison with the NaTc/L-Arg +
vehicle-treated group (P<0.05). Subsequently, the protein
expression levels of SOD1 and SOD2 in the pancreatic tissue were
quantified by western blot analysis (Fig. 7E). As expected, pre-treatment with
AS-IV significantly increased the protein expression levels of SOD1
and SOD2 in the pancreatic tissue.
Effects of AS-IV on AP in vitro
The depletion of ATP can be used to measure the
degree of necrosis of pancreatic acinar cells. The proliferation of
pancreatic acinar cells can be detected by CCK-8, a sensitive
colorimetric assay. AS-IV alleviated the necrosis of pancreatic
acinar cells and improved their viability (Fig. 8A-D). Morever, using western blot
analysis, we determined the nuclear protein expression of levels of
NF-κB p65 and the cytoplasmic protein expression levels of IκBα,
IκBβ, SOD1 and SOD2 (Fig. 8E). As
expected, the results were consistent with those of the in
vivo experiments.
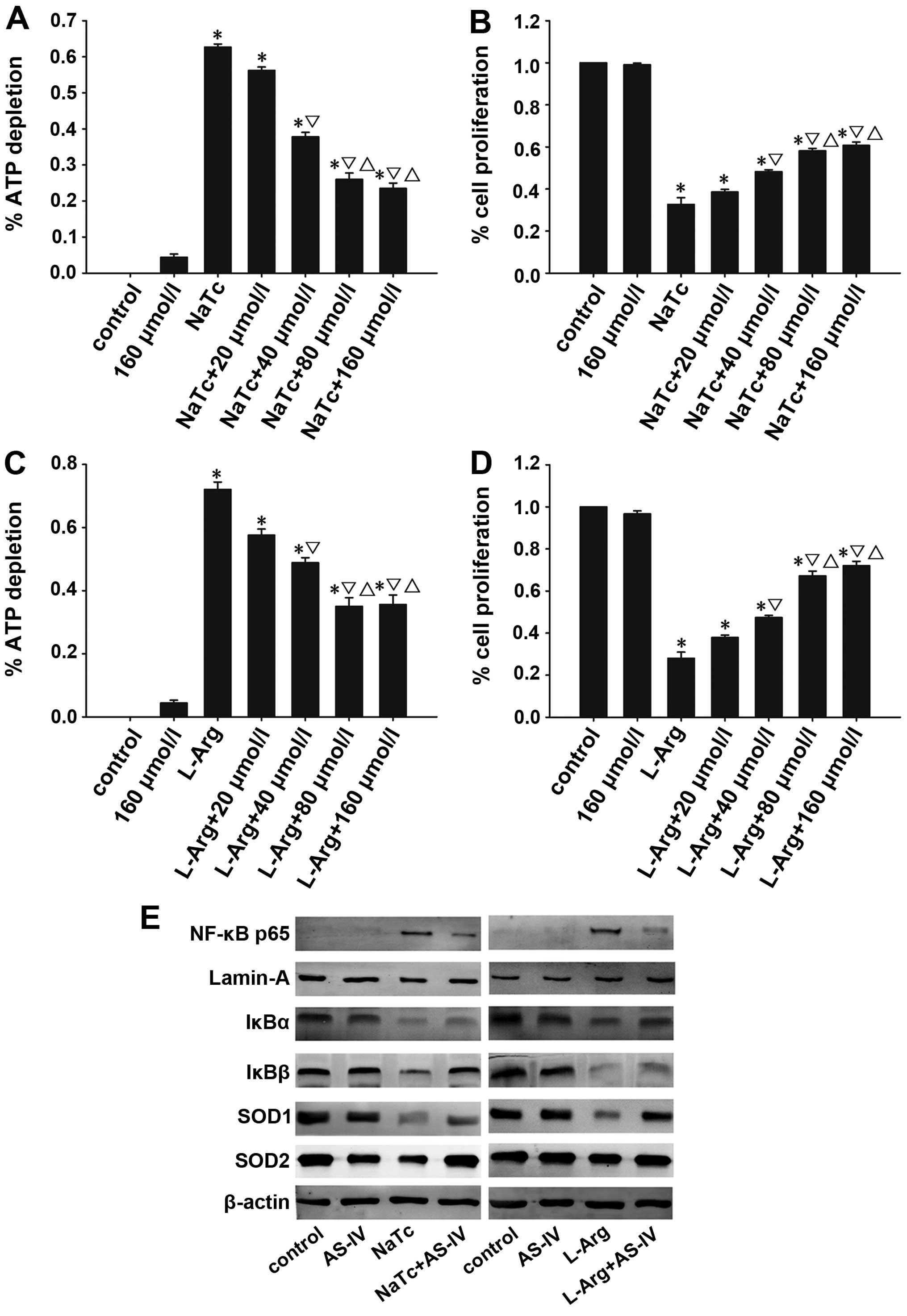 | Figure 8Effects of AS-IV on AP in
vitro. Necrosis and proliferation of pancreatic acinar cells
was detected by the depletion of ATP and CCK-8 assay, respectively.
AS-IV suppressed the dose-dependent depletion of ATP in acinar
cells treated with NaTc/L-Arg (A and C), and increased the
dose-dependent proliferation of acinar cells by CCK-8 (B and D).
(E) The expression of NF-κB p65 in the nucleus, and IκBα, IκBβ,
SOD1 and SOD2 in the cytoplasm of pancreatic acinar cells was
detected by western blot analysis. Data are represented as the
means ± SD from 3 independent experiments. *P<0.05
compared with the normal control group at the same time point;
▽P<0.05 compared with the NaTc-treated group at the
same time point; △P<0.05 compared with the NaTc/L-Arg
+ 40 μmol/l AS-IV-treated group. AS-IV, astragaloside IV;
ATP, adenosine triphosphate; SOD1, manganese superoxide dismutase;
SOD2, cuprum/zinc superoxide dismutase; NaTc, sodium taurocholate;
L-Arg, L-arginine; AP, acute pancreatitis; NF-κB, nuclear
factor-κB; CCK-8, cell counting kit-8. |
Discussion
AP is a potentially fatal disease with increasing
incidence over the years. The pathogenesis of AP remains to be
elucidated despite significant advances over the past 25 years
(31). There is an urgent need to
develop effective therapeutic options for the treatment of AP. It
is generally considered that inflammatory cytokines, leukocytic
infiltration, the activation of NF-κB and oxidative stress are key
factors in the development of AP (32–34).
Inflammatory mediators play a key role in the
pathogenesis of AP and the resultant multiple organ dysfunction
syndrome, which is the primary cause of mortality under this
condition (5). Pancreatic acinar
cells are the primary source of TNF-α during the early phases of
AP. In response to TNF-α arising from acinar cells, an intricate
sequence of events involving the tissue vasculature and
inflammatory cells occur, such as the enhanement of oxidative
stress, the assembly of other cytokines and ROS, the abnormal
upregulation of adhesion molecules and the promotion of the
transmigration of leukocytes into inflamed tissue (11,35,36). All these events are involved in
the development of AP and even systemic inflammatory response
syndrome (SIRS). Previous studies have suggested that the serum
levels of IL-1β correlate with the severity of AP (37,38). A previous study also demonstrated
that IL-6 is intimately linked with pancreatic necrosis and other
organ dysfunction in experimental pancreatitis (39). The serum levels of IL-6 are
significantly higher in SAP compared with mild AP (40,41). Thus, IL-6 is an evaluating
indicator of the severity of AP. It has been suggested that the
early suppression of these pro-inflammatory cytokines relieves the
development of AP and ameliorates the severity of AP (42). The initial injury associated with
AP is closely followed by the second stage, namely, the excessive
transmigration of leukocytes into inflamed tissue, which plays a
crucial role in pancreatic injury and systemic complications. The
extent of neutrophil infiltration in the pancreas is quantified by
measuring tissue MPO activity (43). It has been confirmed that AS-IV
plays an important role in treating inflammatory diseases (18). The anti-inflammatory effects of
AS-IV may be mediated through the inhibition of the activition of
NF-κB, and activated NF-κB promotes the expression of TNF-α, IL-1β
and IL-6 (19,44). In this study, we investigated the
effects of AS-IV in 2 well-characterized models of AP induced by
NaTc/L-Arg in rats; AP in rats is similar to human AP due to the
rapid development of inflammation. Our results demonstrated that
AS-IV significantly ameliorated the pancreatic damage in
NaTc/L-Arg-induced AP as shown by histological characteristics, MPO
activity, and serum amylase and lipase levels. Morever, AS-IV
reduced the serum levels of pro-inflammatory cytokines, such as
TNF-α, IL-1β and IL-6 and inhibited the mRNA expression levels of
TNF-α, IL-1β and IL-6 in the pancreas.
The transcription factor NF-κB is a key factor in
the development of AP based on its ability to regulate the
expression of inflammatory mediators (32). It has been confirmed that the
activation of NF-κB occurs in pancreatic acinar cells in the
initial course of AP, and plays a role in the inflammatory response
during AP (4,33). NF-κB belongs to the Rel family and
exists as a heterodimer or a homodimer formed by polypeptides p50
and p65; RelA/p65 is the crucial transcription factor of the
classical pathway of NF-κB (45–47). In general, the inhibitor protein
of NF-κB (IκB) keeps NF-κB in the cytoplasm by masking its nuclear
localization sequence (48). In
response to extracellular stimuli, IκB proteins become
hyperphosphorylated, ubiquitinated and degraded in proteosomes
(48). Free NF-κB translocates to
the nucleus and binds to its specific site-κB sequences, leading to
the massive transcription of a variety of genes important for
inflammation, including TNF-α, IL-1β and IL-6 (14, 49). In light of these findings, the
NF-κB/IκB system in the pancreas presents a novel and exciting
potential target for the treatment of AP. It has been found that
AS-IV inhibits the activition of NF-κB in vitro (19). In this study, we investigated
whether AS-IV alleviates the severity of AP by inhibiting the
activation of NF-κB. The results from western blot analysis
revealed that NF-κB was activated at 12, 24 and 48 h following the
induction of AP, particularly at 12 h. However, the administration
of AS-IV significantly suppressesed the degradation of IκBα and
IκBβ, thereby decreasing the expression of NF-κB p65 in the nucleus
during AP. The results from immunohistochemistry further revealed
that AS-IV significantly inhibited the staining intensity of
nuclear NF-κB p65 in the pancreas.
Oxidative stress has a significant impact on the
pathogenesis of AP (34,50). During AP, the excessive generation
of ROS and an inefficient intrinsic antioxidative defense system
result in the accumulation of ROS and lipid membrane peroxidation
(50). As the primary defense of
the antioxidant system, SOD is an important antioxidant enzyme,
which specifically detoxifies superoxide radicals to hydrogen
peroxide. In this study, the results from RT-qPCR revealed that
AS-IV increased the mRNA expression levels of SOD1 and SOD2.
Morever, western blot analysis also revealed that AS-IV increased
the protein expression of SOD1 and SOD2. These results indicate
that AS-IV exerts antioxidant effects on the development of AP.
However, the antioxidant mechanisms of action of AS-IV require
further investigation.
In addition, we investigated the effects of AS-IV in
an in vitro model of NaTc/L-Arg-induced AP using pancreatic
acinar cells. In the in vitro model, we found that the high
dose of AS-IV reduced pancreatic acinar cell necrosis and improved
the viability of the acinar cells. Furthermore, we detected the
protein expression of NF-κB p65 in the nucleus, and the expression
of IκBα, IκBβ, SOD1 and SOD2 in the cytoplasm of pancreatic acinar
cells by western blot analysis. As expected, the results were
consistent with those of the in vivo experiments.
In conclusion, our data demonstrate that AS-IV
attenuates the severity of NaTc/L-Arg-induced experimental AP in
rats. Our results revealed that AS-IV exerted anti-inflammatory
effects by inhibiting the activation of NF-κB and suppressing the
secretion of pro-inflammatory cytokines, which are the main
mechanisms of action of AS-IV in AP. As for the antioxidant effects
of AS-IV on AP, further investigations are required. In addition,
the results of the in vitro experiments were consistent with
the results obtained in in vivo experiments. These findings
provide a basis for the further investigation of the therapeutic
role of AS-IV in AP.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81200320 and 81300350),
the Shanghai Science and Technology Commission (no. 11JC1410000),
the Fund of Shanghai Health Bureau (no. 20114315) and the Training
Plan of Excellent Academic Researcher of Shanghai Tenth People’s
Hospital (nos. 12XSGG105 and 04.01.13037).
References
|
1
|
Long J, Song N, Liu XP, Guo KJ and Guo RX:
Nuclear factor-kappaB activation on the reactive oxygen species in
acute necrotizing pancreatitic rats. World J Gastroenterol.
11:4277–4280. 2005.PubMed/NCBI
|
|
2
|
Pandol SJ, Saluja AK, Imrie CW and Banks
PA: Acute pancreatitis: bench to the bedside. Gastroenterology.
132:1127–1151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sweiry JH and Mann GE: Role of oxidative
stress in the pathogenesis of acute pancreatitis. Scand J
Gastroenterol Suppl. 219:10–15. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gukovskaya AS, Gukovsky I, Zaninovic V, et
al: Pancreatic acinar cells produce, release, and respond to tumor
necrosis factor-alpha. Role in regulating cell death and
pancreatitis. J Clin Invest. 100:1853–1862. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bhatia M, Brady M, Shokuhi S, Christmas S,
Neoptolemos JP and Slavin J: Inflammatory mediators in acute
pancreatitis. J Pathol. 190:117–125. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Norman J: The role of cytokines in the
pathogenesis of acute pancreatitis. Am J Surg. 175:76–83. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Makhija R and Kingsnorth AN: Cytokine
storm in acute pancreatitis. J Hepatobiliary Pancreat Surg.
9:401–410. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zyromski N and Murr MM: Evolving concepts
in the pathophysiology of acute pancreatitis. Surgery. 133:235–237.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang XP, Wang L and Zhou YF: The
pathogenic mechanism of severe acute pancreatitis complicated with
renal injury: a review of current knowledge. Dig Dis Sci.
53:297–306. 2008. View Article : Google Scholar
|
|
10
|
Mayer J, Rau B, Gansauge F and Beger HG:
Inflammatory mediators in human acute pancreatitis: clinical and
pathophysiological implications. Gut. 47:546–552. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Malleo G, Mazzon E, Siriwardena AK and
Cuzzocrea S: Role of tumor necrosis factor-alpha in acute
pancreatitis: from biological basis to clinical evidence. Shock.
28:130–140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim H, Seo JY and Kim KH: NF-kappaB and
cytokines in pancreatic acinar cells. J Korean Med Sci. 15(Suppl):
S53–S54. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gukovsky I and Gukovskaya A: Nuclear
factor-kappaB in pancreatitis: Jack-of-all-trades, but which one is
more important? Gastroenterology. 144:26–29. 2013. View Article : Google Scholar
|
|
14
|
Algül H, Tando Y, Schneider G, Weidenbach
H, Adler G and Schmid RM: Acute experimental pancreatitis and
NF-kappaB/Rel activation. Pancreatology. 2:503–509. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schoenberg MH, Büchler M and Beger HG: The
role of oxygen radicals in experimental acute pancreatitis. Free
Radic Biol Med. 12:515–522. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schoenberg MH, Birk D and Beger HG:
Oxidative stress in acute and chronic pancreatitis. Am J Clin Nutr.
62(Suppl 6): 1306S–1314S. 1995.PubMed/NCBI
|
|
17
|
Li M, Yu L, She T, et al: Astragaloside IV
attenuates Toll-like receptor 4 expression via NF-kappaB pathway
under high glucose condition in mesenchymal stem cells. Eur J
Pharmacol. 696:203–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren S, Zhang H, Mu Y, Sun M and Liu P:
Pharmacological effects of Astragaloside IV: a literature review. J
Tradit Chin Med. 33:413–416. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang WJ, Hufnagl P, Binder BR and Wojta
J: Antiinflammatory activity of astragaloside IV is mediated by
inhibition of NF-kappaB activation and adhesion molecule
expression. Thromb Haemost. 90:904–914. 2003.PubMed/NCBI
|
|
20
|
Yang Q, Lu JT, Zhou AW, Wang B, He GW and
Chen MZ: Antinociceptive effect of astragalosides and its mechanism
of action. Acta Pharmacol Sin. 22:809–812. 2001.PubMed/NCBI
|
|
21
|
Li M, Qu YZ, Zhao ZW, et al: Astragaloside
IV protects against focal cerebral ischemia/reperfusion injury
correlating to suppression of neutrophils adhesion-related
molecules. Neurochem Int. 60:458–465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du Q, Chen Z, Zhou LF, Zhang Q, Huang M
and Yin KS: Inhibitory effects of astragaloside IV on
ovalbumin-induced chronic experimental asthma. Can J Physiol
Pharmacol. 86:449–457. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ko JK, Lam FY and Cheung AP: Amelioration
of experimental colitis by Astragalus membranaceus through
anti-oxidation and inhibition of adhesion molecule synthesis. World
J Gastroenterol. 11:5787–5794. 2005.PubMed/NCBI
|
|
24
|
Hu G, Shen J, Cheng L, et al: Reg4
protects against acinar cell necrosis in experimental pancreatitis.
Gut. 60:820–828. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shi C, Zhao X, Wang X and Andersson R:
Role of nuclear factor-kappaB, reactive oxygen species and cellular
signaling in the early phase of acute pancreatitis. Scand J
Gastroenterol. 40:103–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiao WQ, Yin GJ, Fan YT, et al: Catalpol
ameliorates sodium taurocholate-induced acute pancreatitis in rats
via inhibiting activation of nuclear factor kappa B. Int J Mol Sci.
15:11957–11972. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He S, Wang L, Miao L, et al: Receptor
interacting protein kinase-3 determines cellular necrotic response
to TNF-alpha. Cell. 137:1100–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sarr MG: 2012 revision of the Atlanta
classification of acute pancreatitis. Pol Arch Med Wewn.
123:118–124. 2013.PubMed/NCBI
|
|
29
|
Jung KH, Hong SW, Zheng HM, et al:
Melatonin ameliorates cerulein-induced pancreatitis by the
modulation of nuclear erythroid 2-related factor 2 and nuclear
factor-kappaB in rats. J Pineal Res. 48:239–250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Talalay P, Dinkova-Kostova AT and
Holtzclaw WD: Importance of phase 2 gene regulation in protection
against electrophile and reactive oxygen toxicity and
carcinogenesis. Adv Enzyme Regul. 43:121–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sah RP and Saluja A: Molecular mechanisms
of pancreatic injury. Curr Opin Gastroenterol. 27:444–451. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Ji B, Han B, Ernst SA, Simeone D
and Logsdon CD: NF-kappaB activation in pancreas induces pancreatic
and systemic inflammatory response. Gastroenterology. 122:448–457.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grady T, Liang P, Ernst SA and Logsdon CD:
Chemokine gene expression in rat pancreatic acinar cells is an
early event associated with acute pancreatitis. Gastroenterology.
113:1966–1975. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dabrowski A, Konturek SJ, Konturek JW and
Gabryelewicz A: Role of oxidative stress in the pathogenesis of
caerulein-induced acute pancreatitis. Eur J Pharmacol. 377:1–11.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Friedl HP, Till GO, Ryan US and Ward PA:
Mediator-induced activation of xanthine oxidase in endothelial
cells. FASEB J. 3:2512–2518. 1989.PubMed/NCBI
|
|
36
|
Genovese T, Mazzon E, Di Paola R, et al:
Hypericum perforatum attenuates the development of cerulein-induced
acute pancreatitis in mice. Shock. 25:161–167. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dinarello CA: Biologic basis for
interleukin-1 in disease. Blood. 87:2095–2147. 1996.PubMed/NCBI
|
|
38
|
Glasbrenner B and Adler G: Pathophysiology
of acute pancreatitis. Hepatogastroenterology. 40:517–521.
1993.PubMed/NCBI
|
|
39
|
Leindler L, Morschl E, László F, et al:
Importance of cytokines, nitric oxide, and apoptosis in the
pathological process of necrotizing pancreatitis in rats. Pancreas.
29:157–161. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kuśnierz-Cabala B, Gurda-Duda A, Dumnicka
P, et al: Analysis of selected inflammatory markers for early
prediction of severe clinical course of acute pancreatitis. Przegl
Lek. 70:392–396. 2013.(In Polish).
|
|
41
|
Inagaki T, Hoshino M, Hayakawa T, et al:
Interleukin-6 is a useful marker for early prediction of the
severity of acute pancreatitis. Pancreas. 14:1–8. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang XP, Zhang L, Chen LJ, et al:
Influence of dexamethasone on inflammatory mediators and NF-kappaB
expression in multiple organs of rats with severe acute
pancreatitis. World J Gastroenterol. 13:548–556. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bae GS, Kim MS, Jeong J, et al: Piperine
ameliorates the severity of cerulein-induced acute pancreatitis by
inhibiting the activation of mitogen activated protein kinases.
Biochem Biophys Res Commun. 410:382–388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gui D, Huang J, Guo Y, et al:
Astragaloside IV ameliorates renal injury in streptozotocin-induced
diabetic rats through inhibiting NF-kappaB-mediated inflammatory
genes expression. Cytokine. 61:970–977. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Meng Y, Ma QY, Kou XP and Xu J: Effect of
resveratrol on activation of nuclear factor kappa-B and
inflammatory factors in rat model of acute pancreatitis. World J
Gastroenterol. 11:525–528. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Treiber M, Neuhöfer P, Anetsberger E, et
al: Myeloid, but not pancreatic, RelA/p65 is required for fibrosis
in a mouse model of chronic pancreatitis. Gastroenterology.
141:1473–1485. 1485.e1471–1477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hayden MS and Ghosh S: Signaling to
NF-kappaB. Genes Dev. 18:2195–2224. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Barnes PJ: Anti-inflammatory actions of
glucocorticoids: molecular mechanisms. Clin Sci (Lond). 94:557–572.
1998.
|
|
49
|
Ethridge RT, Hashimoto K, Chung DH, Ehlers
RA, Rajaraman S and Evers BM: Selective inhibition of NF-kappaB
attenuates the severity of cerulein-induced acute pancreatitis. J
Am Coll Surg. 195:497–505. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mettu SR, Wig JD, Khullar M, Singh G and
Gupta R: Efficacy of serum nitric oxide level estimation in
assessing the severity of necrotizing pancreatitis. Pancreatology.
3:506–514. 2003. View Article : Google Scholar
|