Introduction
The annual mortality rate due to traumatic injury in
the US is approximately 192,000 individuals annually, while
unintentional injuries are ranked among the top 10 causes of
mortality in adults 65 years of age and older, of both genders
(http://www.nationaltraumainstitute.org/home/trauma_statistics.html).
The leading complication associated with trauma is the onset of
several pathological pathways which leads to multiple-organ failure
several days post-injury (1).
Since aging is characterized by increased oxidative
stress, a heightened inflammatory response, accelerated cellular
senescence and progressive organ dysfunction, it has been proposed
that the homeostatic imbalance with aging significantly alters
cellular responses to injury (2).
In particular, previous research on aging has shown alterations in
the levels or activity of factors involved in key regulatory
processes in the maintenance of mitochondrial structural integrity,
biogenesis and function, thus leading to the hypothesis that the
cellular energetic imbalance may be important for the restoration
of organ function following severe injury in aged individuals
(3). That is, with severe
insults, the mitochondrion, which plays a pivotal role in healthy
aging (2) and, consequently, in
the physiological response to trauma and shock (4), becomes stressed and eventually,
dysfunctional. The most serious consequences of mitochondrial
dysfunction are realized when ATP production falters. The
inhibition of mitochondrial respiration leads to compromised ATP
production and increased levels of reactive oxygen species (ROS)
due to electron leakage. Mitochondrial oxidative stress damages
cellular and mitochondrial protein machinery and genomic DNA,
thereby compromising the repair and recovery capacities of the
body. Furthermore, Jacob et al, using proton nuclear
magnetic resonance (1H-NMR) spectroscopy, demonstrated
that a high intramyocellular lipid (IMCL) content is exhibited
early in the pathogenesis of insulin resistance, highlighting the
importance of the IMCL content as a biomarker of insulin resistance
in patients with type 2 diabetes and their offspring (5). Indeed, the IMCL content in the
soleus muscle has been found to be increased in insulin-resistant
elderly patients, thus providing support for the hypothesis that an
age-associated decline in mitochondrial function contributes to
insulin resistance (6).
High-resolution magic angle spinning (HRMAS)
1H-NMR spectroscopy is a novel non-invasive technique
that yields substantially improved spectral line-widths compared to
conventional NMR, and therefore facilitates the acquisition of
high-resolution spectra from intact cells (7,8)
and unprocessed tissue (9–12).
Although HRMAS 1H-NMR allows the associations between
metabolites, such as IMCLs, and cellular processes to be
investigated more closely than previously possible, it is generally
performed ex vivo (13).
Szczepaniak et al recently demonstrated that the IMCL
content can be quantified accurately in a clinical setting with
in vivo 1H-NMR spectroscopy (14). In the present study, for the first
time and to the best of our knowledge, we applied the emergent
in vivo HRMAS proton magnetic resonance spectroscopy
(1H-MRS) methodology to Drosophila in order to
examine the effects of trauma-induced injury. We anticipated that
in vivo HRMAS 1H-NMR would be a useful tool in
Drosophila since in vitro NMR can be used to
demonstrate the metabolic effects of hypoxia (15) and temperature stress (16) in flies. Drosophila
melanogaster (D. melanogaster), a small, short-lived and
genetically amenable model organism with mutants for several genes
of interest, already available, provides a useful model tool for
assessing the biomarkers of trauma pathophysiology and for
providing critical information on the development of novel
therapies for trauma in a systemic and systematic manner. In
addition, a range of antioxidant defenses has evolved in
Drosophila, protecting the flies against impending oxidative
damage, including antioxidant enzymes (e.g., superoxide dismutase,
catalase and glutathione reductase) that suppress ROS activity
before ROS can damage the vital cellular components (16,17). Although there is a lack of strong
links between antioxidant enzymes and normal aging, antioxidant
enzymes play an important role in extending the longevity of
Drosophila under stressful conditions (17). Therefore, assessing the redox
status of the mitochondria in injured flies is a rational means of
evaluating the antioxidant defense capabilities of the flies.
The present study was designed to examine the
hypothesis that trauma leads to reduced insulin signaling, a
phylogenetically conserved pathway for the regulation of glucose
and lipid metabolism (18,19),
and mitochondrial dysfunction. Insulin signaling was assessed by
cDNA microarrays, insulin resistance was evaluated by the in
vivo assessment of the IMCL content, and mitochondrial
dysfunction was evaluated by estimating in vivo ROS
production. Finally, we examined whether the Szeto-Schiller (SS)-31
peptide, which is known to interact specifically with cardiolipin,
and prevents the conversion of cytochrome c into a
peroxidase while promoting oxidative phosphorylation (20), can reverse these effects. This
hypothesis was examined in aged flies with trauma-induced injury
which were injected with saline or SS-31. In vivo HRMAS
1H-NMR and electron paramagnetic resonance (EPR)
spectroscopy were applied to eliminate the in vitro
artifacts.
Materials and methods
Flies
Aged male D. melanogaster flies (age range,
30–33 days) weighing 0.7–1.0 mg were used in all the experiments.
Oregon-R flies (wild type) were obtained from the Bloomington
Drosophila Stock Center, Department of Biology, Indiana University,
IN, USA. For the microarray genome analysis, wild-type (wt) flies
were used, and the gene expression in flies injured with a thoracic
non-lethal, needle puncture, as previously described (21,22), was compared to that in uninjured
control flies. In vivo HRMAS 1H-NMR spectroscopy
was conducted on 3 groups of flies (n=6 per group). The 3 groups of
flies were as follows: a) uninjured wt flies; b) wt flies injured
24 h prior to examination; and c) injured wt flies injected with
SS-31 12 h post-injury. SS-31 (3 mg/kg) was injected into the
thorax, using a Nanojet II injector (Drummond Scientific, Broomall,
PA, USA) at 0, 3, 6, 24 and 48 h after needle puncture injury, as
described in the study by Apidianakis and Rahme (22). The flies in groups 'a' and 'b'
received injections of saline only following the same schedule, and
a group of uninjured control flies was injected with SS-31 in order
to determine the effects of the agent alone in the absence of
trauma. To demonstrate that the HRMAS 1H-NMR spectra
obtained from whole flies are similar to those of muscle-enriched
fly thoraces, we also analyzed dissected thoraces from 6 flies. Fly
heads, abdomens and legs were removed from the flies and the
thoraces were preserved on ice for 2-6 h prior to their analysis.
In our other experiments, only whole flies were used.
EPR spectroscopy to investigate the redox status was
also conducted on the same groups of flies as used in the HRMAS
1H-NMR spectroscopy experiments. For the in vivo
HRMAS 1H-NMR and EPR spectroscopy experiments, we also
assessed chico1/2 flies (kindly donated by Dr
Robert Perrimon, Department of Genetics, Harvard Medical School,
Boston, MA, USA), bearing 2 mutated alleles of the chico
gene, a Drosophila homolog of vertebrate insulin receptor
substrate 1-4 (IRS1-4), and their genetic control,
chico1/+ flies, were also used, as previously
described (23). Naïve
chico1/2 mutant flies were used as the controls
for the injured flies.
Microarray hybridization and genomic data
analysis
Biotinylated cRNA was generated with 10 µg of
total cellular RNA from the wt flies according to the protocol
outlined by Affymetrix, Inc. (Santa Clara, CA, USA). cRNA was
hybridized onto an Affymetrix GeneChip®
Drosophila Genome oligonucleotide array (Affymetrix, Inc.),
labeled with strep-tavidin-phycoerythrin, washed and scanned
according to the manufacturer's instructions.
Data files of the scanned images of the arrays
hybridized with probes from the RNA extracted from
Drosophila muscle isolated at the specified time points from
the flies with trauma-induced thoracic injury and the control flies
(n=3/group per time point) were converted to cell intensity files
(.CEL files) in the Microarray Suite 5.0 program (MAS; Affymetrix).
The data were scaled to a target intensity of 500, and all possible
pairwise array comparisons of the replicates (injured vs. uninjured
control flies) were performed for each time point using a MAS 5.0
change call algorithm. Probe sets that had a signal value
difference >100, and for which one of the two samples being
compared was not assessed as 'absent', were scored as
differentially modulated when the following two conditions were
met: i) the number of changes registered in the same direction were
at least 3, 4 and 6, when the number of comparisons were 4, 6 and
9, respectively; and ii) the other comparisons were unaltered. This
scoring method compensates partially for biological stochasticity
and technical variation. Based on the ratios of 100 genes
determined to be invariant in the majority of conditions tested
(Affymetrix), an additional constraint of a minimum ratio of 1.65
was applied in order to control for known false-positives at a
level of 5%.
To identify functionally related sets of genes and
processes significantly associated with trauma (at p≤0.05), we used
the GeneSpring tool to characterize the genes found to be
differentially expressed in the flies with trauma-induced thoracic
injury compared to the control flies and compared to the Gene
Ontology Consortium database. The microarray data are available
online at MIAMExpress (accession no. E-MEXP-1287). The expression
levels of all presented genes were verified by reverse
transcription-quantitative polymerase chain reaction experiments
(data not shown).
In vivo HRMAS 1H-NMR
spectroscopy
All HRMAS 1H-NMR spectroscopy experiments
were performed on a wide-bore Bruker Bio-Spin Avance NMR
spectrometer (600.13 MHz) using a 4-mm triple resonance
(1H, 13C and 2H) HRMAS probe (both
from Bruker Corp., Billerica, MA, USA). Prior to being placed in
the spectrometer, each fly was anesthetized by being set on ice for
<1 min. The flies were kept at 4°C while in the spectrometer.
Special care was taken to avoid inflicting further injury on the
already injured flies as they were moved in and out of the rotor.
All flies survived the HRMAS 1H-NMR spectroscopy
experiment, which lasted ~45 min for each fly.
The flies were placed individually into a zirconium
oxide (ZrO2) rotor tube (Bruker BioSpin Corp., Bruker
BioSpin AG, Billerica, MA, USA; 4 mm in diameter, 50 µl),
and 8 µl of external standard 3-(trimethylsilyl)
propionic-2,2,3,3-d4 acid (TSP), (molecular weight, 172, δ=0.00
ppm, 50 mM in D2O) that functioned as a reference for
both resonance chemical shift and quantification, was then pipetted
into each tube. Each fly was placed in the rotor using the insert,
and the insert was then closed with a screw and covered with
parafilm to prevent contact between the fly and the
TSP/D2O. The samples were secured and tightened in the
rotors with a top cap (Bruker Corp.). HRMAS 1H-NMR
spectroscopy was performed at 4°C with a spinning frequency of 2
kHz.
One dimensional (1D) water-suppressed spin-echo
Carr-Purcell-Meiboom-Gill (CPMG) pulse sequence
[90°−(τ−180°−τ)n-acquisition] (24) was performed on single flies. CPMG
is a methodological improvement which is of particular relevance
when developing HRMAS for intact tissue samples ex vivo in
1D acquisition, since it suppresses broad signals that destroy the
linear baseline in typical free induction decay (FID) spectra. As a
result, CPMG proton NMR spectra are free from the broad 'rolling'
component that contributes to the baseline of simple FID spectra.
The CPMG sequence has also been applied to 2 dimensional (2D)
sequences for the same reason. Additional parameters for the CPMG
sequence included an inter-pulse delay of τ = 2π/ωr =
250 µsec, two 180° cycles in total, 256 transients, a
spectral width of 7.2 kHz, 32,768 (32 k) data points, and a 3-sec
repetition time. We selected a spin-echo delay of 30 msec as we
observed that it enabled us to avoid line broadening without the
loss of signals from triglycerides (TG). A longer spin-echo delay
improved all lipid signals, but was not favorable to other
metabolite signals.
1H HRMAS NMR spectroscopy data
processing
NMR spectra were analyzed with MestReC software
(Mestrelab Research, www.mestrec.com). A 0.5-Hz line-broadening apodization
function was applied to CPMG HRMAS 1H FIDs prior to
Fourier transformation. The spectra were referenced relative to TSP
at δ=0.0 ppm (external standard), manually phased, and a Whittaker
baseline estimator was applied to subtract the broad components of
the baseline.
Quantification of metabolites from 1D
CPMG spectra
For metabolite quantification, we used the highly
accurate 'external standard' technique. Metabolite concentrations
were calculated using MestReC software. An automated fitting
routine based on the Levenberg-Marquardt algorithm (25,26) was applied following manual peak
selection; peak positions, intensities, line widths and
Lorentzian/Gaussian ratios were adjusted until the residual
spectrum was minimized. The metabolite concentration (mol/kg) was
calculated using the following equation, as previously described
(27):
massTSP/PMTSP*
Met(area)/TSP(area)*
NTSP/NMet* 1/wt (sample), where
massTSP was constant (0.069 mg), PMTSP was
the molecular weight of TSP (172.23 g/mol), Met signified
metabolites, NTSP denoted the TSP proton number (9 1H),
NMet denoted the metabolite proton number, and wt
signified the sample weight in mg (27).
EPR spectroscopy for post-trauma redox
assessment
X-band EPR with nitroxide was implemented as a
complementary approach to NMR since NMR cannot measure the redox
status (28,29). For redox assessment, each fly was
anesthetized by cooling on ice (4°C) for 1 min and 9.2 nl of 15
mg/kg 2,2,6,6-tetramethylpiperidine 1-oxyl (TEMPO; Sigma-Aldrich,
St. Louis, MO, USA) was then injected into the abdomen via an
automated microsyringe, as previously described (22). Due to the very small size of
Drosophila and in order to minimize repetitive injury, TEMPO
was injected into each fly only once. Therefore, we used different
flies for redox measurements at each time point. The anesthetized
flies were quickly transferred to the EPR sample tube (1/tube) and
restrained gently between cotton plugs to minimize any motion
during the measurements. The EPR sample tube was then placed in the
cavity of the Bruker X-band 9.2-GHz EPR spectrometer to follow the
decay of the nitroxide (TEMPO) for 10-15 min. This method allowed
us to assess the redox status of the flies directly, using an
X-band EPR spectrometer as previously described (28–30).
Typical spectrometer parameters were as follows:
incident microwave power, 0.5 mW; magnetic field center, 348.7 mT;
modulation frequency, 100 kHz; modulation amplitude, 40 µT;
scan width, 1.5 mT and a scan time of 10 sec/scan. The low-field
component of the EPR spectra (i.e., the first EPR line) was
averaged over 1 min, and changes in signal intensity were followed
over time. The rate at which the nitroxide was reduced over time
was taken as an index of the redox status of the flies. In a
previous study, we used a similar procedure to examine the redox
status of muscle in mice following burn injury (30). After the redox measurement, the
flies were gently removed from the tubes and transferred to
Eppendorf tubes to follow their survival.
Statistical analysis
Statistical comparisons were carried out using
analyses of variance (ANOVA) with the Bonferroni correction to
account for multiple of comparisons. A p-value <0.05 (corrected)
was considered to indicate a statistically significant difference;
p-values are reported to two significant digits. Calculations were
performed using SPSS version 12 software (SPSS Inc., Chicago, IL,
USA).
Results
Gene expression
The analysis of Affymetrix microarrays identified
245, 187 and 191 genes as differentially expressed in whole flies
at 1, 6 and 12 h post-trauma. Our comparison of these sets of
differentially expressed genes to the Gene Ontology Consortium
database pointed to electron transport being the predominant
function affected by trauma; the number of electron
transport-related genes whose expression was abnormal at each time
point is shown in Fig. 1.
As shown in Fig.
2, trauma affected the expression of several genes related to
mitochondrial function across multiple time points. As expected,
the expression of genes encoding for mitochondrial uncoupling
protein UCP4 (ucp) was highly upregulated. Additionally, we
observed an upregulation in the expression of genes encoding for
apoptosis-inducing factor (aif) and Drosophila
Forkhead box O (dfoxo). We also observed the altered
expression of genes encoding for Drosophila insulin receptor
(inr), Drosophila AKT (dakt), and
Drosophila phosphatase and tensin homolog (dpten),
which indicates trauma-induced insulin signaling dysregulation;
this insulin signaling dysregulation may be linked to the
downregulation of Drosophila Ets-like gene (delg)
that was also observed. These findings prompted us to examine the
link between mitochondrial dysfunction and insulin signaling in an
in vivo model using in vivo NMR and EPR.
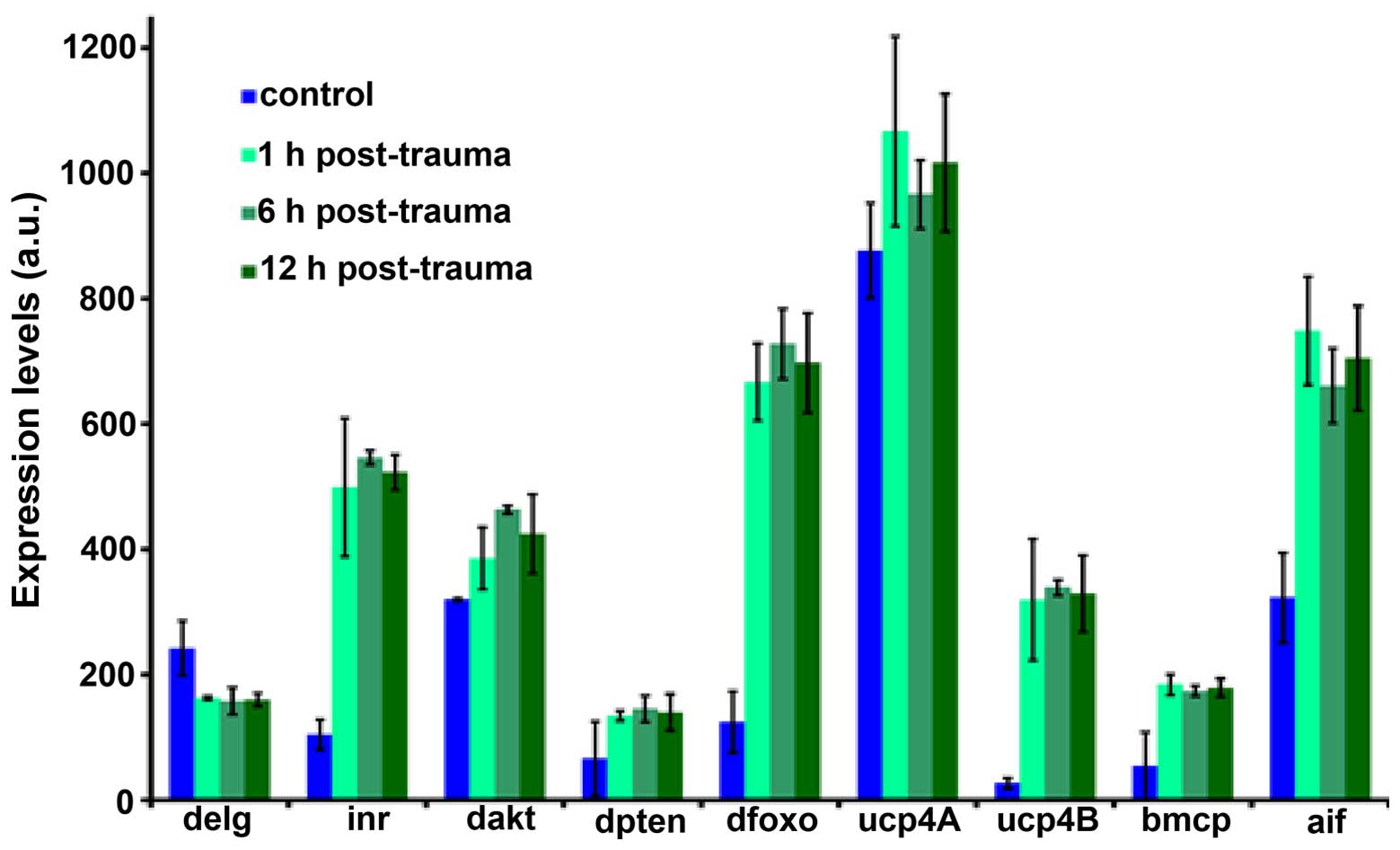 | Figure 2Changes in mitochondrial
function-related gene mRNA expression in flies at 1, 6 and 12 h
post-trauma. Expression levels are reported in arbitrary units
(a.u.). delg, Drosophila NRF-2; inr, Drosophila
insulin receptor; dakt, Drosophila AKT; dpten,
Drosophila phosphatase and tensin homolog; dfoxo,
Drosophila Forkhead box O; ucp4A, uncoupling protein4A;
ucp4b, uncoupling protein4b; bmcp, drosophila ucp5; aif,
apoptosis-inducing factor. |
In vivo 1D 1H HRMAS
Representative in vivo 1D 1H HRMAS
CPMG spectra from uninjured aged wt flies, injured aged wt flies
and chico1/2 mutants are illustrated in Fig. 3 together with a 1D 1H
HRMAS CPMG summed spectrum from the thorax of dissected wt flies
(inset), which represents primarily skeletal muscle, as the fly
thorax is highly enriched in skeletal muscle, demonstrating that
the spectra from whole flies are similar to those from the
muscle-enriched thorax. Principal lipid components [CH3
(0.89 ppm), (CH2)n (1.33 ppm), CH2C-CO (1.58
ppm), CH2C=C (2.02 ppm), CH2C=O (2.24 ppm),
CH=CH (5.33 ppm)], glycerol (4.10, 4.30 and 5.24 ppm), acetate (Ac,
1.92 ppm), β-alanine (β-Ala, 2.55 ppm), phosphocholine (PC, 3.22
ppm) and phosphoethanolamine (PE, 3.22 ppm)] were detected. Signals
at 2.02 ppm were assigned to methylene protons of the
CH2-CH=CH moiety of monounsaturated fatty acids (i.e.,
palmitoleic acid). Of note, we detected increased levels of
polyunsaturated fatty acids (PUFAs, CH2C=O at 2.24 ppm).
The unsaturated acids were identified by a signal at 5.33 ppm
produced by protons of the -CH=CH- moiety.
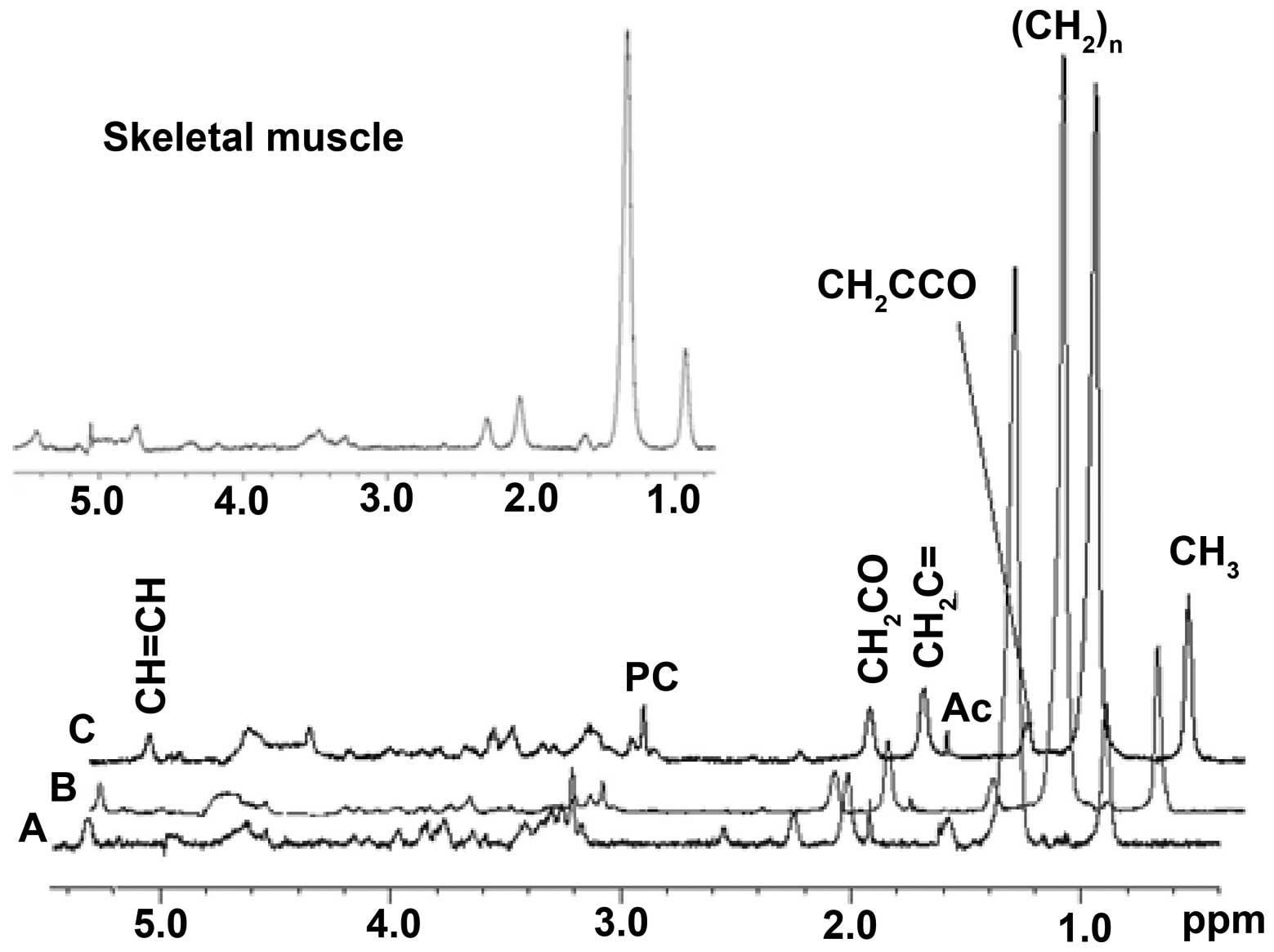 | Figure 3Representative in vivo 1D
HRMAS 1H CPMG spectra of (A) uninjured aged wild-type
(wt) flies, (B) injured old wt flies and (C) chico1/2
mutants. Lipid components: CH3 (0.89 ppm),
(CH2)n (1.33 ppm), CH2C-CO (1.58 ppm),
acetate (Ac, 1.92 ppm), CH2C=C (2.02 ppm),
CH2C=O (2.24 ppm), β-alanine (β-Ala, 2.55 ppm),
phosphocholine (PC, 3.22 ppm), and phosphoethanolamine (PE, 3.22
ppm), glycerol (4.10, 4.30 ppm 1,3-CH; 5.22 ppm 2-CH2),
CH=CH (5.33 ppm). Note that spectra from chico1/2
mutants with insulin resistance have increased insulin biomarker
(CH2)n, and that the spectra from injured wt flies have
a similar profile. The spectra in the insert are from the thorax of
dissected flies and thus represent primarily skeletal muscle; note
their similarity to spectra for whole flies. |
The metabolic HRMAS NMR profiles of the injured aged
wt flies were similar to those of the chico1/2
mutants (Fig. 3). The SS-31
injection normalized the NMR profiles of the injured aged wt flies
(Fig. 4). Quantitative
analysis revealed significant increases in (CH2)n lipids
at 1.33 ppm (an insulin resistance biomarker) and CH=CH lipids at
5.33 ppm (an apoptosis biomarker) in the injured aged wt flies
compared to the uninjured aged wt flies (Table I). The majority of the other lipid
resonances were significantly elevated in the injured flies
compared to the uninjured flies. The levels of the
(CH2)n and CH=CH lipids were normalized in the injured
flies injected with SS-31 (Table
I).
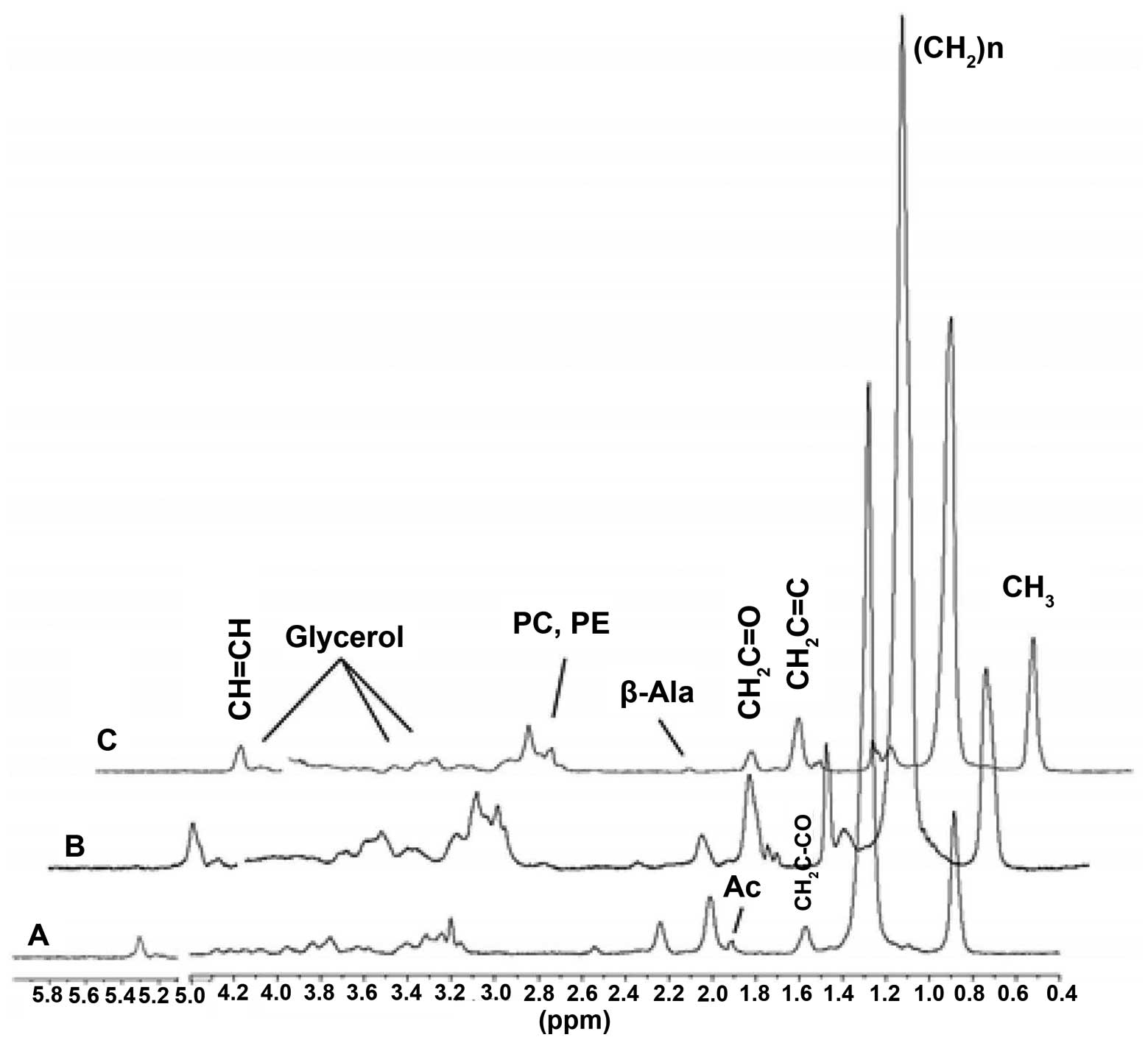 | Figure 4Representative summed in vivo
1D HRMAS 1H CPMG spectra of (A) uninjured aged wild-type
(wt) flies and (B) injured aged wt flies, (C) injured aged wt flies
treated with Szeto-Schiller (SS)-31. Lipid components:
CH3 (0.89 ppm), (CH2)n (1.33 ppm),
CH2C-CO (1.58 ppm), acetate (Ac, 1.92 ppm),
CH2C=C (2.02 ppm), CH2C=O (2.24 ppm),
β-alanine (β-Ala, 2.55 ppm), phosphocholine (PC, 3.22 ppm),
phosphoethanolamine (PE, 3.22 ppm), glycerol (4.10, 4.30 ppm
1,3-CH; 5.22 ppm 2-CH2), and CH=CH (5.33 ppm). Note that
injured flies injected with SS-31 (C) have a profile that is
similar to that of the uninjured old wt flies (A). |
 | Table IMean quantity (in µmol/g ±
standard errors) of selected lipid components in live wt flies
determined by 1H HRMAS NMR. |
Table I
Mean quantity (in µmol/g ±
standard errors) of selected lipid components in live wt flies
determined by 1H HRMAS NMR.
| Time | Lipid
components | CH3 |
(CH2)n |
CH2CCO |
CH2C= |
CH2CO | CH=CH |
|---|
| Chemical shift (δ,
ppm) | 0.89 | 1.33 | 1.58 | 2.02 | 2.24 | 5.33 |
|---|
| 6 h | Uninjured | 0.18±0.01 | 1.41±0.08 | 0.06±0.003 | 0.13±0.01 | 0.07±0.01 | 0.08±0.01 |
| Injured | 0.27±0.03 | 2.10±0.25 | 0.16±0.08 | 0.24±0.06 | 0.13±0.03 | 0.13±0.02 |
| % change | 50.0 | 48.94 | 166.67 | 84.62 | 85.71 | 62.50 |
| p-value | 0.022 | 0.024 | 0.260 | 0.085 | 0.071 | 0.015 |
| 12 h | Injured +
saline | 0.26±0.003 | 1.94±0.05 | 0.13±0.01 | 0.22±0.004 | 0.10±0.01 | 0.13±0.004 |
| Injured + SS31 | 0.17±0.002 | 1.29±0.02 | 0.06±0.002 | 0.12±0.002 | 0.06±0.001 | 0.08±0.002 |
| % change | −33.38 | −33.43 | −54.95 | −44.09 | −45.05 | −39.90 |
| p-value |
<0.001 |
<0.001 | 0.0013 | <
0.001 | <
0.001 | <
0.001 |
Analysis of the transverse relaxation time,
T2, of the metabolites and the TSP standard in the 1D
1H CPMG spectra across echo times (TE at 30, 60, 100,
300, 450 and 600 msec) revealed that the T2 decay rate
for the -CH3 groups at 0.89 ppm (1,156±72 msec) was very
close to that of TSP (1,125±103 msec). The T2 values for
(CH2)n at 1.33 ppm (516±14 msec), CH2C=C at
2.02 ppm (537±35 msec) and CH=CH at 5.33 ppm (469±27 msec) were
almost identical to each other, equating to approximately half the
T2 values for CH2CCO at 1.58 ppm (292±5.0
msec) and CH2CO at 2.24 ppm (265±16 msec). Even at
TE=600 ms, these peaks were not completely decayed, indicating that
the lipid relaxation kinetics resembled those of TSP.
EPR spectroscopy
The typical in vivo X-band EPR spectra
acquired from the flies is illustrated in Fig. 5A. Our minute-by-minute analysis of
the changes in the low field component of the TEMPO nitroxide
(Fig. 5B) indicated that the
redox status was compromised in the flies at 1 and 6 h post-injury.
The redox status (Kr) in the injured flies at 1 and 6 h post-injury
was 0.0187±0.0040/sec and 0.0187±0.0043/sec, respectively, and did
not differ significantly between these two time points. However,
these redox measures were significantly higher than those observed
in the uninjured flies in the control group (0.0131±0.0027/sec;
p=0.018 vs. 1 h post-injury and p=0.024 vs. 6 h post-injury).
Treatment with SS-31 significantly decreased the nitroxide decay
rate at 6 h post-injury to 0.0130±0.0036/sec (p=0.040) vs. injured
flies not treated with SS-31 at 6 h post-injury), a level similar
to that of the uninjured control group (p=0.933).
Discussion
In the present study, we examined the link between
insulin signaling and mitochondrial dysfunction following trauma by
combining genomic analysis with in vivo 1D 1H
HRMAS and EPR spectroscopy in Drosophila. The expression of
electron transport protein genes, in particular, was substantially
altered in the flies with trauma-induced injury. This pattern of
change is indicative of mitochondrial dysfunction following trauma
and is in agreement with the findings of a previous study of ours
on burn-associated trauma (31).
Also consistent with our prior findings in relation to burn trauma
(32), we observed the
upregulation of aif, dfoxo, as well as genes related
to mitochondrial uncoupling. Furthermore, we were intrigued to find
the altered expression of the insulin signaling-related genes,
inr, dakt, and dpten, which is also consistent
with our murine burn trauma model (33), as well as the downregulation of
delg, which could be a downstream effect of insulin
signaling dysregulation.
The results of our subsequent in vivo HRMAS
1H-NMR and EPR experiments support the hypothesis that
trauma can reduce insulin signaling through a mechanism that
involves a phylogenetically conserved pathway for the regulation of
glucose and lipid metabolism (18,19), and that SS-31 can promote the
recovery of mitochondrial function and alleviate this condition.
Our findings broaden those of prior reports using HRMAS
1H-MRS in Drosophila (34), and provide evidence for the
hypothesis that trauma in aging is linked to dysregulated insulin
signaling. This link may explain the mitochondrial dysfunction that
accompanies insulin resistance in trauma and aging in mammals.
Specifically, using in vivo HRMAS
1H-NMR (14.1 T), we detected lipids and small
metabolites in live Drosophila. At 14.1-T, we were able to
achieve a sufficiently brief acquisition time (~45 min) to attain
zero mortality. In accordance with other previously published in
vivo skeletal muscle spectra (35–37), our in vivo fly spectra
exhibited high amounts of lipids (particularly TG), but fewer
metabolites than described in certain other studies (38,39). Relative to these previous studies,
we were working with a smaller sample size and using a lower spin
rate, which may have affected spectral resolution. Given the low
body weight of the fruit fly, NMR-visible non-lipid components are
expected to contribute only a small percentage of the total signal,
with a concomitantly weaker sensitivity of detection. Indeed, even
spectra from the muscle-dense thorax of dissected flies were
similar to the spectra from whole flies (inset of Fig. 3).
The current thinking is that, following trauma, the
disruption of oxidative homeostasis, the overproduction of ROS and
the opening of the mitochondrial permeability transition pore
promote mitochondrial dysfunction, leading to the activation of
necrotic and/or apoptotic cell death pathways (4). Based on our present findings, we
propose that trauma affects mitochondrial function through the
disruption of insulin signaling, which is related to the altered
expression of delg [the Drosophila homologue of the α
subunit of nuclear factor (erythroid-derived 2)-related factor-2
(NRF-2α)] and spargel (the Drosophila homologue of
peroxisome proliferator-activated receptor-γ coactivator 1).
In light of our present findings (summarized in
Fig. 6) and other evidence in the
literature (40), suggesting that
Spargel mediates transcription in response to insulin signaling in
parallel with Dfoxo, it is important to determine whether Spargel
mediates a negative feedback loop in insulin signaling that enables
it to set a threshold for insulin signaling in the control of
metabolism. Whilst Spargel is not necessary for the maintenance of
basal mitochondrial mass under normal physiological conditions, it
becomes necessary in the absence of Delg, the fly homologue of
NRF-2a (40). The loss of
function of both Spargel and Delg leads to a greater loss of
mitochondrial mass than the loss of Delg alone (40,41), suggesting that the two factors may
act in parallel. Additionally, our previously published studies
(30–35,42–45), and the current findings, are
consistent with the possibility that Spargel promotes mitochondrial
dysfunction via uncoupling proteins; experiments testing this
possibility are currently in progress.
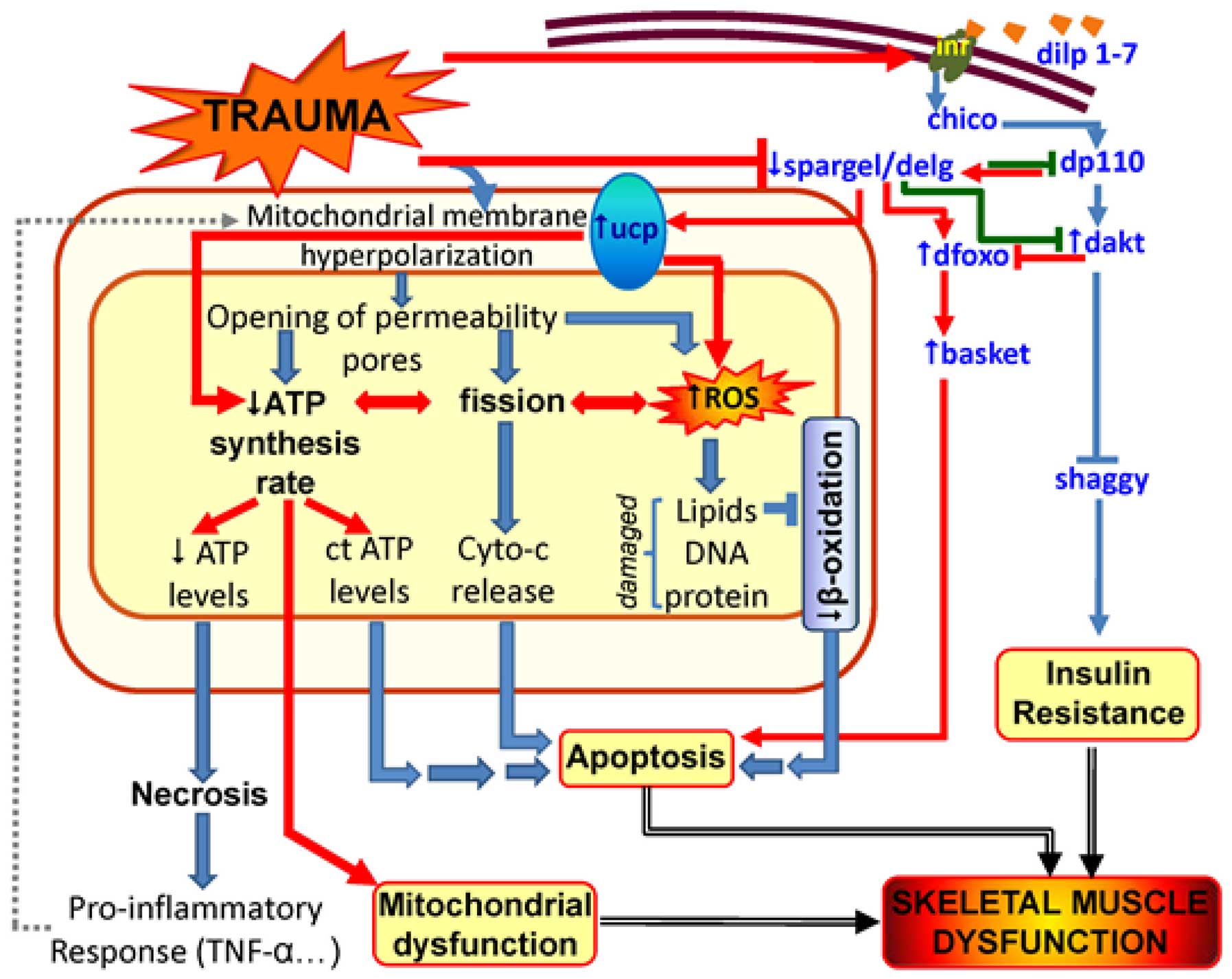 | Figure 6Proposed molecular mechanism of
trauma-induced skeletal muscle dysfunction. Red lines/arrows,
aspects of trauma-induced mitochondrial dysfunction suggested by
our data. Green lines, proposed negative feedback mechanism of
spargel in the insulin pathway. Blue arrows, aspects of
trauma-induced mitochondrial dysfunction established previously in
the literature (18–19,40–46). ct, stable; spargel, PGC-1
analogue in flies; delg, NRF-2 analogue in flies; ucp, uncoupling
proteins; dfoxo, Forkhead box O anologue in flies; basket, JNK
analogue in flies; dilp 1–7, analogues of insulin, IGF-1 and IGF-2
in flies; inr, Drosophila insulin receptor; chico, IRS 1–4
analogue in flies; dp110, PI3K analogue in flies; dakt,
Drosophila AKT analogue in flies; shaggy, GSK-3 (glycogen
synthase kinase-3) analogue in flies; ROS, reactive oxygen species;
Cyto-c, cytochrome c; ATP, adenosine triphosphate;
TNF-α, tumor necrosis factor-α. |
From a biomedical perspective, a principal finding
of our study is that mobile lipids accumulate in muscle tissue in
response to injury (Fig. 3 and
Table I). These findings support
the hypothesis that trauma leads to insulin resistance. Indeed,
insulin resistance has been suggested to develop following critical
illness and severe injury (46).
Elevated IMCL levels have been associated with insulin resistance,
a major metabolic dysfunction of diabetes (47,48), aging (6,48–50), burn trauma (32–36) and obesity (51–54). Previous genomic (55) and gene expression data in studies
of human diabetics (56) suggest
that elevated IMCL levels are the result of a deficiency in
mitochondrial oxidative capacity (56,57), suggesting that an elevated IMCL
content is indicative of reduced mitochondrial oxidation and
phosphorylation.
Another principal finding of the present study was
that ceramide had accumulated in aged injured flies (Table I and Fig. 3), to a greater extent than in
young injured flies, as we have previously shown (34). Ceramide accumulation decreases
insulin- stimulated glucose transporter type 4 (GLUT4)
trans-location to the plasma membrane, which results in decreased
glucose transport and, consequently, in the development of insulin
resistance. Paumen et al demonstrated that saturated fatty
acids (e.g., palmitoleic acid, signal at 2.02 ppm in our study)
induced the de novo synthesis of ceramide and programmed
cell death (58); they suggested
that the inhibition of carnitine palmitoyl transferase I activity
induced both sphingolipid synthesis and palmitate-induced cell
death. Moreover, Ruddock et al (59) suggested that long-chain saturated
fatty acids (e.g., palmitoleic acid C16:0) inhibited insulin
activity and attenuated insulin signal transduction in hepatoma
cells, and concluded that an increase in palmitoleic acid is a
harbinger of insulin resistance. If so, the signal at 2.02 ppm,
which was high in the aged flies in our study, could be a biomarker
of insulin resistance. Our NMR data further suggest that trauma
induces the activation of the sphingolipid pathway, including
ceramide, which functions as an intracellular apoptosis signal.
Indeed, our data demonstrated vinyl proton accumulation at 5.33 ppm
(Table I), including protons from
ceramide and possibly other sphingolipids, such as sphingosine and
other monounsaturated fatty acids. These enhanced signals are in
accordance with the hypothesis that the sphingolipid pathway may
contribute to trauma-mediated apoptosis (60,61).
Of note, SS-31-treated flies exhibited a
significantly reduced 5.33-ppm resonance in our study (Table I, Fig. 3), which supports our prior finding
that SS-31 promotes mitochondrial respiration and inhibits ROS
production (62). SS-31 inhibits
cardiolipin peroxidation and mitochondrial permeability transition,
thus preventing the release of cytochrome c and consequent
apoptosis (62,63), and SS-31 has been shown to promote
mitochondrial bioenergetics in skeletal muscle and prevent insulin
resistance (64–67).
SS-31 has been reported to improve mitochondrial
respiration in skeletal muscle, reduce ROS production following
immobilization, and prevent the development of insulin resistance
(64,65,68). Consistent with our prior findings
on mouse burn trauma (8), our
complementary EPR results confirm that trauma-induced changes
occurred in the redox status, and that SS-31 facilitated the
normalization of the mitochondrial redox status in injured flies
(Table I, Fig. 5). SS-31 targets and concentrates
in the inner mitochondrial membrane, where it reduces mitochondrial
oxidative stress via several mechanisms: it can scavenge electrons
directly via its dimethyltyrosine residue (69), reduce mitochondrial ROS activity
(64,68) and prevent cardiolipin peroxidation
elicited by the cardiolipin/cytochrome c complex (62). Cardiolipin peroxidation triggers
mitochondrial permeability transition, and SS-31 has been reported
to inhibit mitochondrial permeability transition and swelling and
apoptosis (69). The ability of
SS-31 to inhibit insulin resistance induced by a high fat diet
(65) and burn trauma (66) may be a result of its protective
effect on mitochondrial function.
Trauma can induce lipid peroxidation, a biomarker
of lipid damage from oxidative stress (70). Indeed, trauma-induced free
radicals can attack intra-membrane PUFAs, which our NMR data
revealed to be elevated in injured flies (see CH2CO
resonance at 0.24 ppm in Table I)
to form lipid peroxides which can disrupt membrane permeability,
integrity and function, compromise cellular components, and lead to
further injury (71,72). Our use of EPR spectroscopy to
analyze trauma-related oxidative damage and lipid peroxidation in
Drosophila, allowed us to corroborate previously described
protective effects of SS-31 against lipid peroxidation (63,73,74).
In the present study, we demonstrated that the
innovative approach of employing in vivo HRMAS NMR
complemented by EPR spectroscopy is a sensitive method for
characterizing metabolic and mitochondrial perturbations in injured
Drosophila at the molecular level. Furthermore, we
demonstrated that SS-31 can reverse injury-induced mitochondrial
dysfunction and associated insulin resistance (75), as evidenced by NMR, while also
attenuating injury-induced oxidative stress effects, as evidenced
by EPR. The relative timing and interactions of SS-31 warrant
further investigations in which the constituent events are
isolated.
As corollary benefits, our approach advances the
development of non-invasive in vivo research approaches in
Drosophila, offers biomarkers which may be used to
investigate biomedical paradigms, and thus may direct novel
mitochondrial therapeutic development. Our demonstration of the
utility of this approach in the extraordinarily well characterized
and practicable model organism D. melanogaster can pave the
way for a shift in the current research paradigm in trauma. Given
that our findings are relevant for the treatment of mitochondrial
dysfunction that occurs in as many chronic and terminal diseases as
in trauma, this study is of great relevance to public health.
Abbreviations:
|
Ac
|
acetate
|
|
Ala
|
alanine
|
|
β-Ala
|
β-alanine
|
|
Arg
|
arginine
|
|
CPMG
|
Carr-Purcell-Meiboom-Gill
|
|
EMCLs
|
extramyo-cellular lipids
|
|
FID
|
free induction decay
|
|
1H-NMR
|
proton nuclear magnetic resonance
|
|
HRMAS
|
high-resolution magic angle
spinning
|
|
EPR
|
electron paramagnetic resonance
|
|
IMCLs
|
intra myocellular lipids
|
|
NMR
|
nuclear magnetic resonance
|
|
PE
|
phosphoethanolamine
|
|
PC
|
phosphocholine
|
|
PUFA
|
polyunsaturated fatty acid
|
|
ROS
|
reactive oxygen species
|
|
SS-31
|
Szeto-Schiller-31
|
|
Tau
|
taurine
|
|
TG
|
triglycerides
|
|
wt
|
wild-type
|
Acknowledgments
The present study was supported in part by a grant
from DM103014 of the Defense Medical Research and Development
Program (DMRDP) to Laurence G. Rahme (Aria A. Tzika,
co-investigator), and Shriner's Hospital for Children research
grants to Aria A. Tzika (no. 8893) and Laurence G. Rahme (no.
8892). The authors also thank Dr Ann Power Smith (Write Science
Right, Las Vegas, NV, USA) for providing editorial assistance. The
SS peptide technology has been licensed for commercial development
to Stealth Peptides Inc. by the Cornell Research Foundation (CRF),
and both CRF and Hazel H. Szeto have financial interests. We would
also like to thank Dr Robert Perrimon for providing the
chico flies.
References
|
1
|
Sobrino J and Shafi S: Timing and causes
of death after injuries. Proc (Bayl Univ Med Cent). 26:120–123.
2013.
|
|
2
|
López-Lluch G, Irusta PM, Navas P and de
Cabo R: Mitochondrial biogenesis and healthy aging. Exp Gerontol.
43:813–819. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Poulose N and Raju R: Aging and injury:
alterations in cellular energetics and organ function. Aging Dis.
5:101–108. 2014.PubMed/NCBI
|
|
4
|
Hubbard WJ, Bland KI and Chaudry IH: The
role of the mitochondrion in trauma and shock. Shock. 22:395–402.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jacob S, Machann J, Rett K, Brechtel K,
Volk A, Renn W, Maerker E, Matthaei S, Schick F, Claussen CD and
Häring HU: Association of increased intramyocellular lipid content
with insulin resistance in lean nondiabetic offspring of type 2
diabetic subjects. Diabetes. 48:1113–1119. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Petersen KF, Befroy D, Dufour S, Dziura J,
Ariyan C, Rothman DL, DiPietro L, Cline GW and Shulman GI:
Mitochondrial dysfunction in the elderly: possible role in insulin
resistance. Science. 300:1140–1142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weybright P, Millis K, Campbell N, Cory DG
and Singer S: Gradient, high-resolution, magic angle spinning 1H
nuclear magnetic resonance spectroscopy of intact cells. Magn Reson
Med. 39:337–345. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blankenberg FG, Storrs RW, Naumovski L,
Goralski T and Spielman D: Detection of apoptotic cell death by
proton nuclear magnetic resonance spectroscopy. Blood.
87:1951–1956. 1996.PubMed/NCBI
|
|
9
|
Cheng LL, Ma MJ, Becerra L, Ptak T, Tracey
I, Lackner A and González RG: Quantitative neuropathology by high
resolution magic angle spinning proton magnetic resonance
spectroscopy. Proc Natl Acad Sci U S A. 12:6408–6413. 1997.
View Article : Google Scholar
|
|
10
|
Cheng LL, Newell K, Mallory AE, Hyman BT
and Gonzalez RG: Quantification of neurons in Alzheimer and control
brains with ex vivo high resolution magic angle spinning proton
magnetic resonance spectroscopy and stereology. Magn Reson Imaging.
20:527–533. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Millis KK, Maas WE, Cory DG and Singer S:
Gradient, high-resolution, magic-angle spinning nuclear magnetic
resonance spectroscopy of human adipocyte tissue. Magn Reson Med.
38:399–403. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Barton SJ, Howe FA, Tomlins AM, Cudlip SA,
Nicholson JK, Bell BA and Griffiths JR: Comparison of in vivo 1H
MRS of human brain tumours with 1H HR-MAS spectroscopy of intact
biopsy samples in vitro. MAGMA. 8:121–128. 1999.PubMed/NCBI
|
|
13
|
Alves TC, Jarak I and Carvalho RA: NMR
methodologies for studying mitochondrial bioenergetics. Methods Mol
Biol. 810:281–309. 2012. View Article : Google Scholar
|
|
14
|
Szczepaniak LS, Babcock EE, Schick F,
Dobbins RL, Garg A, Burns DK, McGarry JD and Stein DT: Measurement
of intracellular triglyceride stores by H spectroscopy: validation
in vivo. Am J Physiol. 276:E977–E989. 1999.PubMed/NCBI
|
|
15
|
Feala JD, Coquin L, McCulloch AD and
Paternostro G: Flexibility in energy metabolism supports hypoxia
tolerance in Drosophila flight muscle: metabolomic and
computational systems analysis. Mol Syst Biol. 3:992007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pedersen KS, Kristensen TN, Loeschcke V,
Petersen BO, Duus JO, Nielsen NC and Malmendal A: metabolomic
signatures of inbreeding at benign and stressful temperatures in
Drosophila melanogaster. Genetics. 180:1233–1243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Le Bourg E and Le Bourg E: Oxidative
stress, aging and longevity in Drosophila melanogaster. FEBS Lett.
498:183–186. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Garofalo RS: Genetic analysis of insulin
signaling in Drosophila. Trends Endocrinol Metab. 13:156–162. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saltiel AR and Kahn CR: Insulin signalling
and the regulation of glucose and lipid metabolism. Nature.
414:799–806. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szeto HH: First-in-class
cardiolipin-protective compound as a therapeutic agent to restore
mitochondrial bioenergetics. Br J Pharmacol. 171:2029–2050. 2014.
View Article : Google Scholar :
|
|
21
|
Apidianakis Y, Mindrinos MN, Xiao W, Lau
GW, Baldini RL, Davis RW and Rahme LG: Profiling early infection
responses: Pseudomonas aeruginosa eludes host defenses by
suppressing antimicrobial peptide gene expression. Proc Natl Acad
Sci USA. 102:2573–2578. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Apidianakis Y and Rahme LG: Drosophila
melanogaster as a model host for studying Pseudomonas aeruginosa
infection. Nat Protoc. 4:1285–1294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Böhni R, Riesgo-Escovar J, Oldham S,
Brogiolo W, Stocker H, Andruss BF, Beckingham K and Hafen E:
Autonomous control of cell and organ size by CHICO, a Drosophila
homolog of vertebrate IRS1-4. Cell. 97:865–875. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meiboom S and Gill D: Modified spiin-echo
method for measuring nuclear relaxation time. Rev Sci Instrum.
29:688–691. 1958. View Article : Google Scholar
|
|
25
|
Levenberg K: Method for the solution of
certain non-linear problems in least squares. Q Appl Math.
2:164–168. 1944.
|
|
26
|
Marquardt D: An algorithm for
least-squares estimation of nonlinear parameters. SIAM J Appl Math.
11:431–441. 1963. View Article : Google Scholar
|
|
27
|
Swanson MG, Zektzer AS, Tabatabai ZL,
Simko J, Jarso S, Keshari KR, Schmitt L, Carroll PR, Shinohara K,
Vigneron DB and Kurhanewicz J: Quantitative analysis of prostate
metabolites using 1H HR-MAS spectroscopy. Magn Reson Med.
55:1257–1264. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khan N and Das DK: Application of in vivo
EPR for tissue pO2 and redox measurements. Methods Mol
Biol. 559:131–139. 2009. View Article : Google Scholar
|
|
29
|
Swartz HM, Khan N and Khramtsov VV: Use of
electron paramagnetic resonance spectroscopy to evaluate the redox
state in vivo. Antioxid Redox Signal. 9:1757–1771. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Khan N, Mupparaju SP, Mintzopoulos D,
Kesarwani M, Righi V, Rahme LG, Swartz HM and Tzika AA: Burn trauma
in skeletal muscle results in oxidative stress as assessed by in
vivo electron paramagnetic resonance. Mol Med Rep. 1:813–819.
2008.PubMed/NCBI
|
|
31
|
Padfield KE, Astrakas LG, Zhang Q, Gopalan
S, Dai G, Mindrinos MN, Tompkins RG, Rahme LG and Tzika AA: Burn
injury causes mitochondrial dysfunction in skeletal muscle. Proc
Natl Acad Sci USA. 102:5368–5373. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Q, Cao H, Astrakas LG, Mintzopoulos
D, Mindrinos MN, Schulz J III, Tompkins RG, Rahme LG and Tzika AA:
Uncoupling protein 3 expression and intramyocellular lipid
accumulation by NMR following local burn trauma. Int J Mol Med.
18:1223–1229. 2006.PubMed/NCBI
|
|
33
|
Tzika AA, Astrakas LG, Cao H, Mintzopoulos
D, Zhang Q, Padfield K, Yu H, Mindrinos MN, Rahme LG and Tompkins
RG: Murine intramyocellular lipids quantified by NMR act as
metabolic biomarkers in burn trauma. Int J Mol Med. 21:825–832.
2008.PubMed/NCBI
|
|
34
|
Righi V, Apidianakis Y, Mintzopoulos D,
Astrakas L, Rahme LG and Tzika AA: In vivo high-resolution magic
angle spinning magnetic resonance spectroscopy of Drosophila
melanogaster at 14.1 T shows trauma in aging and in innate
immune-deficiency is linked to reduced insulin signaling. Int J Mol
Med. 26:175–184. 2010.PubMed/NCBI
|
|
35
|
Astrakas LG, Goljer I, Yasuhara S,
Padfield KE, Zhang Q, Gopalan S, Mindrinos MN, Dai G, Yu YM, Martyn
JA, et al: Proton NMR spectroscopy shows lipids accumulate in
skeletal muscle in response to burn trauma-induced apoptosis. FASEB
J. 19:1431–1440. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Weis J, Johansson L, Ortiz-Nieto F and
Ahlström H: Assessment of lipids in skeletal muscle by LCModel and
AMARES. J Magn Reson Imaging. 30:1124–1129. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang L, Salibi N, Wu Y, Schweitzer ME and
Regatte RR: Relaxation times of skeletal muscle metabolites at 7T.
J Magn Reson Imaging. 29:1457–1464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Griffin JL, Williams HJ, Sang E and
Nicholson JK: Abnormal lipid profile of dystrophic cardiac tissue
as demonstrated by one- and two-dimensional magic-angle spinning
(1)H NMR spectroscopy. Magn Reson Med. 46:249–255. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen JH, Sambol EB, Decarolis P, O'Connor
R, Geha RC, Wu YV and Singer S: High-resolution MAS NMR
spectroscopy detection of the spin magnetization exchange by
cross-relaxation and chemical exchange in intact cell lines and
human tissue specimens. Magn Reson Med. 55:1246–1256. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tiefenböck SK, Baltzer C, Egli NA and Frei
C: The Drosophila PGC-1 homologue Spargel coordinates mitochondrial
activity to insulin signalling. EMBO J. 29:171–183. 2010.
View Article : Google Scholar :
|
|
41
|
Baltzer C, Tiefenböck SK, Marti M and Frei
C: Nutrition controls mitochondrial biogenesis in the Drosophila
adipose tissue through Delg and cyclin D/Cdk4. PLoS One.
4:e69352009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Padfield KE, Zhang Q, Gopalan S, Tzika AA,
Mindrinos MN, Tompkins RG and Rahme LG: Local and distant burn
injury alter immuno-inflammatory gene expression in skeletal
muscle. J Trauma. 61:280–292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tzika AA, Mintzopoulos D, Padfield K,
Wilhelmy J, Mindrinos MN, Yu H, Cao H, Zhang Q, Astrakas LG, Zhang
J, et al: Reduced rate of adenosine triphosphate synthesis by in
vivo31P nuclear magnetic resonance spectroscopy and downregulation
of PGC-1β in distal skeletal muscle following burn. Int J Mol Med.
21:201–208. 2008.PubMed/NCBI
|
|
44
|
Righi V, Andronesi O, Mintzopoulos D and
Tzika AA: Molecular characterization and quantification using state
of the art solid-state adiabatic TOBSY NMR in burn trauma. Int J
Mol Med. 24:749–757. 2009.PubMed/NCBI
|
|
45
|
Tzika AA, Mintzopoulos D, Mindrinos M,
Zhang J, Rahme LG and Tompkins RG: Microarray analysis suggests
that burn injury results in mitochondrial dysfunction in human
skeletal muscle. Int J Mol Med. 24:387–392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Thompson LH, Kim HT, Ma Y, Kokorina NA and
Messina JL: Acute, muscle-type specific insulin resistance
following injury. Mol Med. 14:715–723. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Machann J, Thamer C, Schnoedt B, Stefan N,
Stumvoll M, Haring HU, Claussen CD, Fritsche A and Schick F: Age
and gender related effects on adipose tissue compartments of
subjects with increased risk for type 2 diabetes: a whole body
MRI/MRS study. MAGMA. 18:128–137. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Nakagawa Y, Hattori M, Harada K, Shirase
R, Bando M and Okano G: Age-related changes in intramyocellular
lipid in humans by in vivo H-MR spectroscopy. Gerontology.
53:218–223. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rouffet D, Villars C, Fissoune R,
Sappey-Marinier D, Laville M, Ibarrola D, Sothier M, Monnet MF,
Ovize M and Bonnefoy M: Intramyocellular lipid variations in active
older men: relationship with aerobic fitness. Acta Physiol (Oxf).
207:516–523. 2013. View Article : Google Scholar
|
|
50
|
Petersen KF, Morino K, Alves TC, Kibbey
RG, Dufour S, Sono S, Yoo PS, Cline GW and Shulman GI: Effect of
aging on muscle mitochondrial substrate utilization in humans. Proc
Natl Acad Sci USA. 112:11330–11334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sinha R, Dufour S, Petersen KF, LeBon V,
Enoksson S, Ma YZ, Savoye M, Rothman DL, Shulman GI and Caprio S:
Assessment of skeletal muscle triglyceride content by (1)H nuclear
magnetic resonance spectroscopy in lean and obese adolescents:
relationships to insulin sensitivity, total body fat, and central
adiposity. Diabetes. 51:1022–1027. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Schrauwen-Hinderling VB, Hesselink MK,
Schrauwen P and Kooi ME: Intramyocellular lipid content in human
skeletal muscle. Obesity (Silver Spring). 14:357–367. 2006.
View Article : Google Scholar
|
|
53
|
Consitt LA, Bell JA and Houmard JA:
Intramuscular lipid metabolism, insulin action, and obesity. IUBMB
Life. 61:47–55. 2009. View Article : Google Scholar :
|
|
54
|
Johnson AB, Argyraki M, Thow JC, Cooper
BG, Fulcher G and Taylor R: Effect of increased free fatty acid
supply on glucose metabolism and skeletal muscle glycogen synthase
activity in normal man. Clin Sci (Lond). 82:219–226. 1992.
View Article : Google Scholar
|
|
55
|
Mootha VK, Bunkenborg J, Olsen JV,
Hjerrild M, Wisniewski JR, Stahl E, Bolouri MS, Ray HN, Sihag S,
Kamal M, et al: Integrated analysis of protein composition, tissue
diversity, and gene regulation in mouse mitochondria. Cell.
115:629–640. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Petersen KF, Dufour S, Befroy D, Garcia R
and Shulman GI: Impaired mitochondrial activity in the
insulin-resistant offspring of patients with type 2 diabetes. N
Engl J Med. 350:664–671. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Patti ME, Butte AJ, Crunkhorn S, Cusi K,
Berria R, Kashyap S, Miyazaki Y, Kohane I, Costello M, Saccone R,
et al: Coordinated reduction of genes of oxidative metabolism in
humans with insulin resistance and diabetes: potential role of PGC1
and NRF1. Proc Natl Acad Sci USA. 100:8466–8471. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Paumen MB, Ishida Y, Muramatsu M, Yamamoto
M and Honjo T: Inhibition of carnitine palmitoyltransferase I
augments sphingolipid synthesis and palmitate-induced apoptosis. J
Biol Chem. 272:3324–3329. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ruddock MW, Stein A, Landaker E, Park J,
Cooksey RC, McClain D and Patti ME: Saturated fatty acids inhibit
hepatic insulin action by modulating insulin receptor expression
and post-receptor signalling. J Biochem. 144:599–607. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yasuhara S, Perez ME, Kanakubo E, Yasuhara
Y, Shin YS, Kaneki M, Fujita T and Martyn JA: Skeletal muscle
apoptosis after burns is associated with activation of proapoptotic
signals. Am J Physiol Endocrinol Metab. 279:E1114–E1121.
2000.PubMed/NCBI
|
|
61
|
Tomera JF and Martyn J: Systemic effects
of single hindlimb burn injury on skeletal muscle function and
cyclic nucleotide levels in the murine model. Burns. 14:210–219.
1988. View Article : Google Scholar
|
|
62
|
Birk AV, Liu S, Soong Y, Mills W, Singh P,
Warren JD, Seshan SV, Pardee JD and Szeto HH: The
mitochondrial-targeted compound SS-31 re-energizes ischemic
mitochondria by interacting with cardiolipin. J Am Soc Nephrol.
24:1250–1261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Szeto HH: Cell-permeable,
mitochondrial-targeted, peptide antioxidants. AAPS J. 8:E277–E283.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Min K, Smuder AJ, Kwon OS, Kavazis AN,
Szeto HH and Powers SK: Mitochondrial-targeted antioxidants protect
skeletal muscle against immobilization-induced muscle atrophy. J
Appl Physiol 1985. 111:1459–1466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Anderson EJ, Lustig ME, Boyle KE, Woodlief
TL, Kane DA, Lin CT, Price JW III, Kang L, Rabinovitch PS, Szeto
HH, et al: Mitochondrial H2O2 emission and
cellular redox state link exces fat intake to insulin resistance in
both rodents and humans. J Clin Invest. 119:573–581. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Carter EA, Bonab AA, Goverman J, Paul K,
Yerxa J, Tompkins RG and Fischman AJ: Evaluation of the antioxidant
peptide SS31 for treatment of burn-induced insulin resistance. Int
J Mol Med. 28:589–594. 2011.PubMed/NCBI
|
|
67
|
Siegel MP, Kruse SE, Percival JM, Goh J,
White CC, Hopkins HC, Kavanagh TJ, Szeto HH, Rabinovitch PS and
Marcinek DJ: Mitochondrial-targeted peptide rapidly improves
mitochondrial energetics and skeletal muscle performance in aged
mice. Aging Cell. 12:763–771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Powers SK, Hudson MB, Nelson WB, Talbert
EE, Min K, Szeto HH, Kavazis AN and Smuder AJ:
Mitochondria-targeted antioxidants protect against mechanical
ventilation-induced diaphragm weakness. Crit Care Med.
39:1749–1759. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhao K, Zhao GM, Wu D, Soong Y, Birk AV,
Schiller PW and Szeto HH: Cell-permeable peptide antioxidants
targeted to inner mitochondrial membrane inhibit mitochondrial
swelling, oxidative cell death, and reperfusion injury. J Biol
Chem. 279:34682–34690. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Smith JA, Park S, Krause JS and Banik NL:
Oxidative stress, DNA damage, and the telomeric complex as
therapeutic targets in acute neurodegeneration. Neurochem Int.
62:764–775. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ozkur MK, Bozkurt MS, Balabanli B,
Aricioglu A, Ilter N, Gürer MA and Inalöz HS: The effects of EGb
761 on lipid peroxide levels and superoxide dismutase activity in
sunburn. Photodermatol Photoimmunol Photomed. 18:117–120. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Akeo K, Amaki S, Suzuki T and Hiramitsu T:
Melanin granules prevent the cytotoxic effects of L-DOPA on retinal
pigment epithelial cells in vitro by regulation of NO and
superoxide radicals. Pigment Cell Res. 13:80–88. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Szeto HH: Mitochondria-targeted peptide
antioxidants: novel neuroprotective agents. AAPS J. 8:E521–E531.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Szeto HH: Development of
mitochondria-targeted aromatic-cationic peptides for
neurodegenerative diseases. Ann N Y Acad Sci. 1147:112–121. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lowell BB and Shulman GI: Mitochondrial
dysfunction and type 2 diabetes. Science. 307:384–387. 2005.
View Article : Google Scholar : PubMed/NCBI
|
















