Introduction
Osteoporosis is a disease where decreased bone
strength increases the risk of bone fractures. It is the most
common reason for bone fractures among older individuals. According
to published data, the National Osteoporosis Foundation (NOF)
estimates that as many as 54 million Americans have osteoporosis
and decreased bone mass (1–3).
Studies have shown that approximately half of the female and one
quarter of the male population will suffer from bone fractures due
to osteoporosis (4,5). More than 50% of non-Hispanic
Caucasian and Asian women aged 50 years and older have a decreased
bone mass, thus increasing their risk for osteoporosis (6,7). A
major reason for this is that the female body produces less
estrogen after menopause, and estrogen plays an important role in
helping to prevent bone loss (8,9).
Decreased bone mass poses a serious health concern for individuals
worldwide, as it is known that severe osteoporosis is associated
with other disorders and diseases. Osteoporosis may occur due to
lower than normal peak bone mass and greater than normal bone loss.
Bone loss increases after menopause due to lower levels of estrogen
(8,9,10,11).
Osteoporosis may also occur due to a number of
diseases or treatments, including alcoholism, anorexia,
hyperthyroidism, surgical removal of the ovaries and kidney disease
(12). Certain medications
increase the rate of bone loss, including some anti-seizure
medications, chemotherapy, proton pump inhibitors, selective
serotonin reuptake inhibitors and steroids (13). Insufficient exercise and smoking
are also considered risk factors. Among these factors, diabetes
mellitus (DM) is considered an important risk factor for the
development of osteoporosis and this has been widely noted by
researchers (14,15). Diabetes-related osteoporosis (DO)
is a general metabolic bone disorder which increases the tendency
for fractures due to osteopenia, microstructural changes in bone
tissue, decreased bone strength and increased friability, which is
one of the main complications of DM affecting the skeletal system.
With the increasing morbidity associated with DM, the morbidity
associated with DO is markedly increasing, and has become a main
cause of mutilation and death in patients with diabetes. DO
severely affects the quality of life of patients, and adds heavy
economic burden to families and society.
The pathogenesis and prevention of DO has become a
worldwide research hotspot (16).
In previous studies, the phosphoinositide 3-kinase (PI3K)/AKT,
c-Jun N-terminal kinase (JNK)/mitogen-activated protein kinase
(MAPK) and nuclear factor-κB (NF-κB) signaling pathways have been
proven to be possible targets with which to inhibit the development
and progression of DO (17,18). The underlying molecular mechanisms
of of action these pathways in DO remain unknown.
Bergapten (BP;
C12H8O4), a natural
anti-inflammatory and anti-tumor agent isolated from bergamot
essential oil, other citrus essential oils and grapefruit juice,
has been used to prevent lipopolysaccharide (LPS)-mediated
osteoclast formation, bone resorption and osteoclast survival in
vitro (19). Bergapten has
also been shown to significantly inhibit the production of
pro-inflammatory cytokines (20).
However, the effects of Bergapten on diabetes-related osteoporosis
are not yet understood.
Hence, in the present study, in an aim to elucidate
the mechanisms of action of these pathways in DO, we examined the
protective effects of BP on DO and investigated the role of the
PI3K/AKT, JNK/MAPK and NF-κB signaling pathways in the protective
effects of BP in osteoprotegerin (OPG) knockout mice.
Materials and methods
Animals
For the purposes of this study, 6-8-week-old
wild-type male C57/B6 mice (n=102) and OPG knockout [OPG(−/−) or
OPG-KO; n=56] male C57/B6 mice (weighing 25–30 g) were purchased
from the Zhejiang University Experimental Animal Center (Zhejiang,
China) and kept in a temperature and humidity-controlled
environment (25±2°C, 50±10% humidity) with a standard 12-h
light/12-h dark cycle with food and water provided in their cages.
All experiments involving the use of animals were carried out in
accordance with the Institutional Animal Care and Use Committee of
the Department of Orthopaedics, Zaozhuang Municipal Hospital. BP
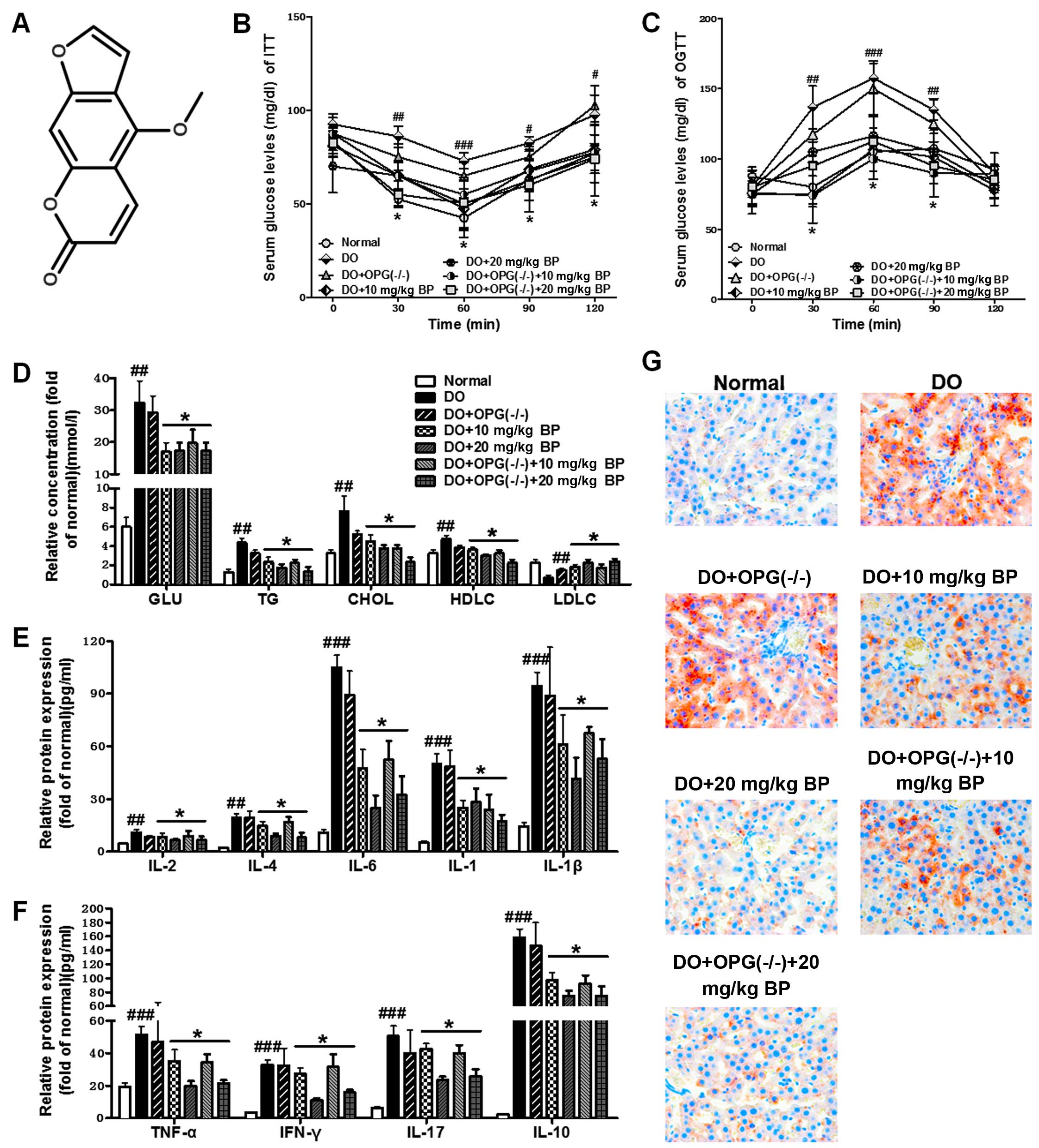
(chemical structure shown in Fig.
1A) (CAS:68144-21-8, purity ≥98%) was purchased from Nanjing
Biological Technology Co., Ltd. (Nanjing, China) and prepared in
phosphate-buffered saline (PBS). The rodent high-fat diet was
purchased from Research Diets Inc. (D12492, 60 kcal% fat; New
Brunswick, NJ. USA). The mice were randomly divided into 7 groups
as follows (6–10 mice per group): i) normal (wild-type); ii) DO;
iii) DO + OPG(−/−); iv) DO + 10 mg/kg/day BP; v) DO + 20 mg/kg/day
BP; vi) DO + OPG(−/−) + 10 mg/kg/day BP; and vii) DO + OPG(−/−) +
20 mg/kg/day BP. The mice were orally administered BP for 20 weeks.
The whole experimental period lasted 24 weeks.
Histological analysis
After the final drug administration, the mice were
anesthetized with diethyl ether and mouse blood was harvested by
eyeball extraction, 3 mouse liver tissues were collected and stored
in 4% paraformaldehyde. The liver samples obtained were subjected
to Oil Red O staining analysis and observed under a light
microscope. The Oil Red O staining was performed by Nanjing
Biotechnology, Co., Ltd. For bone tissue analysis, Masson's
staining, hematoxylin and eosin (H&E) staining, medical X-rays
and immunohistochemical analysis (IHC) were used to evaluate the
extent of osteoporosis and the involvement of related key signaling
pathways. In brief, formalin-fixed and paraffin-embedded mouse
liver specimens were sectioned at 4 mm, stained with H&E, and
used for histopathological examinations. For immunohistochemical
staining, the sections were dewaxed and dehydrated. Following
antigen retrieval in citrate buffer, we blocked the sections
overnight at 4°C. The sections were then probed with rabbit
anti-phosphorylated (p-)JNK (4668), anti-p-AKT (13038), anti-IκBα
(4812) and anti-vascular endothelial growth factor (VEGF; 9698)
antibodies (Cell Signaling Technolology, Danvers, MA, USA) and an
ultra-sensitive immunohistochemistry kit (Maixin, Fuzhou, China)
was used. X-rays were obtained using the MutiFocus X-ray imaging
system (Faxitron Bioptics LLC, Tucson, AZ, USA). Masson's staining
was performed by Biohelper Co. (Nanjing, China) according to a
standard protocol.
In order to determine the damage inflicted on the
bones of DO and DO + OPG-KO mice, we also measured the the bone
volume fraction (BV/TV), trabecular thickness (Tb.Th) and
trabecular number (Tb.N).
Biochemical analysis
The oral glucose tolerance test (OGTT) (2 g/kg
glucose) and insulin tolerance test (ITT) were conducted according
to reported methods (21). In
brief, OGTTs were performed after final weeks of treatment in mice
that had been fasted for 6 h. Glucose was orally administered (3
g/kg body weight, 660 g/l glucose solution) and blood glucose was
determined through a glucose meter using 3.5 µl of blood
collected from the tip of the tail vein before and at the
administration of the glucose load (30, 60, 90 and 120 min). Also,
the other indicators [lipid metabolism-related indicators,
including glucose (GLU), triglycerides (TG), total cholesterol
(CHOL), high-density lipoprotein cholesterol (HDLC) and low-density
lipoprotein cholesterol (LDLC); and inflammatory cytokines,
including interleukin (IL)-2, IL-4, IL-6. IL-1 and IL-1β, tumor
necrosis factor (TNF)-α, interferon (IFN)-γ, IL-17 and IL-10] shown
in Fig. 1D-1F were investigated
using respective ELISA kits obtained from Nanjing Jiancheng
Bioengineering Institute (Nanjing, China) according to the
manufacturer's specifications.
Reverse transcription-quantitative PCR
(RT-qPCR) and western blot analysis
After the final drug administration, the mice were
anesthetized with diethyl ether and mouse mice blood was harvested
by eyeball extraction, and 3 mouse liver (as mentioned
above)/visceral adipose/kidney tissues were collected and stored in
4% paraformaldehyde. The Total RNA isolation system from Omega
Bio-Tek, Inc. (Norcross, GA, USA) was used to isolate total RNA
from the visceral adipose tissue samples. Briefly, 1 µg of
total RNA was reverse transcribed using the M-MLV-RT system. The
reaction was carried at 43°C for 1 h and terminated by the
inhibition of the enzyme at 70°C for 10 min. qPCR were carried out
using SYBR-Green kits (Bio-Rad, Hercules, CA, USA) on an ABI PRISM
7900HT detection system (Applied Biosystems, Foster City, CA, USA).
Invitrogen (Carlsbad, CA, USA) produced all sequences of the
primers for qPCR. The primers used for PCR are listed in Table I.
 | Table ISequences of primers used for PCR. |
Table I
Sequences of primers used for PCR.
| Gene | Primer sequences
(5′→3′) |
|---|
| RANKL (forward) |
AGTACACCTATCATGGAG |
| RANKL (reverse) | TAGATTGTGAATACTG |
| RANK (forward) |
GACAGCCACAATGGGTAGGATGT |
| RANK (reverse) |
TCTGTGCCAGATATCTTCTGCTA |
| TAK1 (forward) |
TTGCTCCTGCTTCGTTC |
| TAK1 (reverse) |
AATAAAGTTTTATGTTGAT |
| NIK(forward) | TCGCAAAGTACGAATC |
| NIK (reverse) |
TAGTCCCACAGACAACC |
| IKKα (forward) |
GTCCAAAACGACTGTCA |
| IKKα (reverse) |
CGGACTCCATACAATCTG |
| IKKβ (forward) |
ATGAGAGTCCCTCGTGTGA |
| IKKβ (reverse) |
TTCTTGTTGCTGACTAACG |
| IκBα (forward) |
CCTTAGCCCACTACTTC |
| IκBα (reverse) |
CTCCGGCAGGAACTTGAA |
| TRAF6
(forward) |
TGCGAATGACGCAGCAA |
| TRAF6
(reverse) |
TCCGTCAAGTCGGTCA |
| GAPDH
(forward) |
AAGGCTGGGGCTCATTTG |
| GAPDH
(reverse) |
GGGCCATCCACAGTCTTC |
For western blot analysis, proteins were extracted
from the kidney tissues using the T-PER Tissue Protein Extraction
Reagent kit (Thermo Fisher Scientific, Inc., Pittsburg, PA, USA)
according to the manufacturer's instructions. Protein
concentrations were determined using the BCA protein assay kit, and
equal amounts of protein were loaded per well on a 10% sodium
dodecyl sulphatepolyacrylamide gel. Subsequently, the proteins were
transferred onto polyvinylidene difluoride membranes. The membranes
were blocked with Tris-buffered saline containing 0.05% Tween-20
(TBS-T), supplemented with 5% skim milk (Sigma, St. Louis, MO, USA)
at room temperature for 2 h on a rotary shaker, followed by washing
in TBS-T. The membranes were then incubated with specific primary
antibodies diluted in TBST at 4°C overnight. The primary polyclonal
antibodies used were as follows: rabbit anti-GAPDH (1:1000,
ab9485), receptor activator of NF-κB (RANK) ligand (RANKL; 1:1000,
ab9957), RANK (1:1000, ab200369), transforming growth factor
β-activated kinase 1 (TAK1; 1:1000, ab109526), NF-κB-inducing
kinase (NIK; 1:1000, ab203568), IKKα (1:1000, ab32041), IKKβ
(1:1000, ab32135), IκBα (ab7217), p-IκBα (1:1000, ab24783), TRAF6
(1:1000, ab33915), AKT (1:1000, ab8805), p-AKT (1:1000, ab38449),
PI3K (1:1000, ab151549), p-PI3K (1:1000, ab182651), JNK (1:1000,
ab4821), p-JNK (1:1000, ab4821) and p-mammalian target of rapamycin
(mTOR; 1:1000, ab109268) (all from Abcam, USA). diluted in TBST,
were incubated with the membranes at 4°C overnight. Subsequently,
the membranes were washed with TBS-T followed by incubation with
the peroxidase-conjugated secondary antibody [anti-rabbit antibody
(ab191866); Abcam] at room temperature for 1 h. The immunoactive
proteins were detected by using an enhanced chemiluminescence
western blotting detection kit. Western blot bands were observed
using the GE Healthcare ECL Western Blotting Analysis system (GE
Healthcare Life Sciences, Pittsburgh, PA, USA) and exposed to X-ray
film (Kodak).
Statistical analysis
Data are presented as the means ± SEM. Differences
between the treatment groups and the controls were compared using
GraphPad Prism software (version 6.0; GraphPad Software, San Diego,
CA, USA) with a one-way ANOVA and Dunn's least significant
difference tests or the Student's t-test. Differences between
groups were considered significant at a vaue of p<0.05. The bars
in the figures indicate the means ± SEM (n=10 mice per group).
Results
Anti-inflammatory effects of BP on
DO-induced systemic inflammation and disorders in DO or OPG-KO
mice
In the present study, using a mouse model of DO
induced by a high-fat diet, we investigated the inflammatory
response induced by diabetes and the anti-inflammatory effects of
BP on inflammatory cytokine production. As shown in Fig. 1B and C, OGTT and ITT tests
revealed that a high-fat diet increased glucose levels in serum and
promoted the development of insulin resistance. The OPG-KO mice
exhibited higher glucose levels among most of the groups and
compared to the normal control group. We also examined the levels
of lipid metabolism-related indicators in each treatment group to
examine the effects of OPG-KO and BP treatment on metabolic
disorders caused by a high-fat diet. Our results revealed that the
levels of lipid metabolism-related indicators, including GLU, TG,
CHOL and LDLC were significantly increased in the DO and DO +
OPG-KO groups, compared with the BP treatment group and the normal
control (Fig. 1D). In addition,
the high-fat diet-induced production of inflammatory cytokines was
examined by ELISA. Our results revealed that a high-fat diet
markedly increased the inflammatory responses and cytokine
formation and release (Fig. 1E and
F). However, treatment with BP attenuated the increase in the
inflammatory response. In addition, we used H&E staining to
examine the effects of a high-fat diet on liver tissue. The shown
in Fig. 1G indicated that a
high-fat diet promoted lipid accumulation in the liver in the DO
and DO + OPG-KO group; this accumulation was inhibited by treatment
with BP (10 and 20 mg/kg). Our data indicated that DO and DO in
conjuncion with OPG-KO and with a high-fat diet cause systemic
inflammation and disorders, and that these negative effects can be
significantly suppressed by treatment with BP.
Effects of OPG knockout and BP on DO
We then examined the effects of OPG knockout and BP
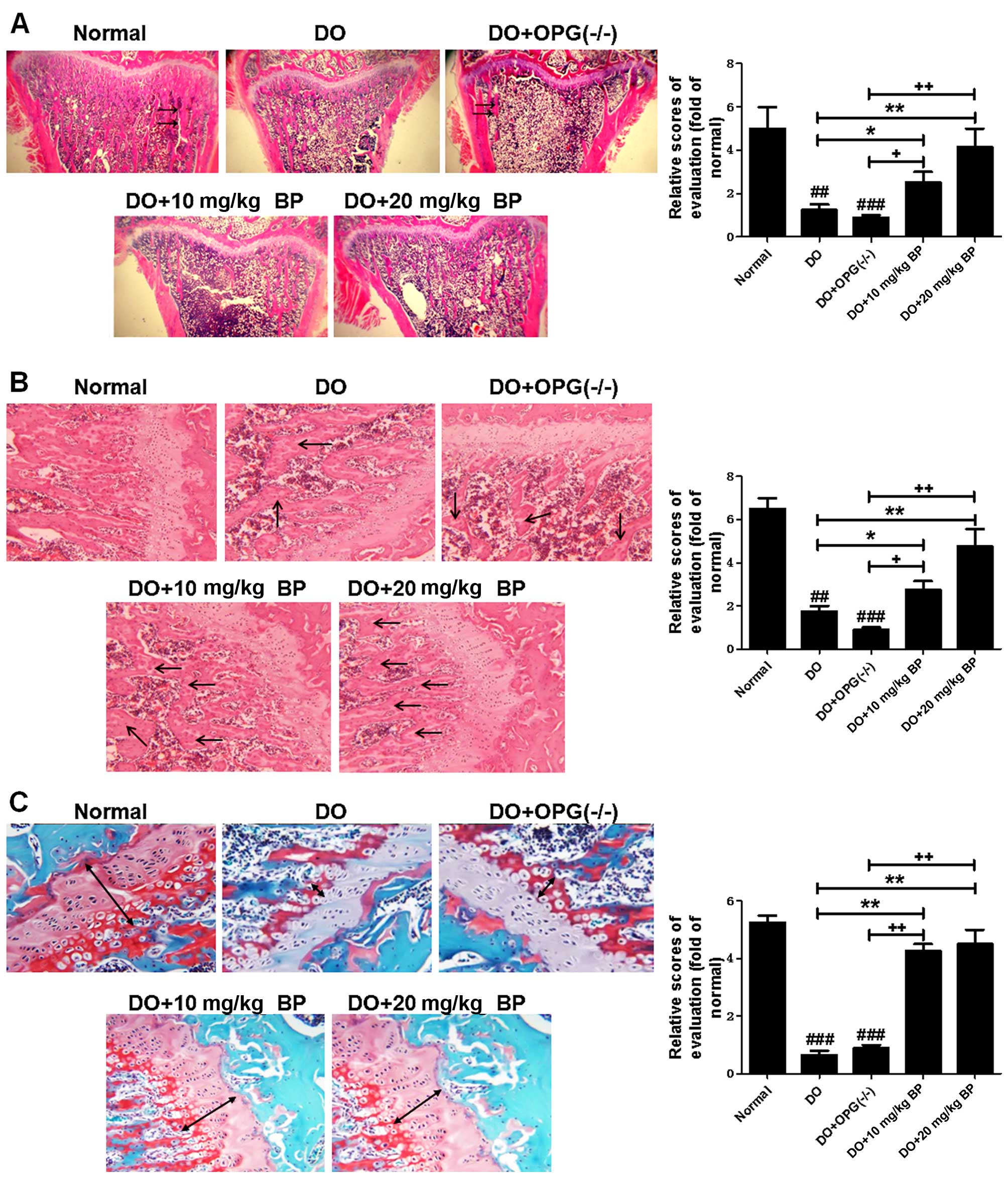
on diabetes-related osteoporosis. As shown in Fig. 2A–C, the BV/TV, Tb.Th and Tb.N were
tested to evaluate the damage inflicted on bone in DO and DO +
OPG-KO. The BV/TV, Tb.Th and Tb.N of the mice in the DO + OPG-KO
groups were significantly decreased compared with those of the
normal group. However, treatment with BP atteuated these negative
effects in a dose-independent manner. In addition, X-ray scanning
(Fig. 2D) revealed that bone
mineral density in the mice in the DO and DO + OPG-KO groups was
decreased and bone loss was more severe compared with the normal
group. In the DO + OPG-KO group, this was even more severe. These
data indicate that OPG is important for protection against
osteoporosis and that BP exerts protective effects against DO.
Furthermore, H&E and Masson's staining (Fig. 3A–C) were used to further examine
the bone injury assoicated with DO. Our results revealed that in
the DO and DO + OPG-KO groups, the trabecular number was decreased
and the trabecular bone became thin and there was a decline in
trabecular structure compared to the normal group. These effects
were more evident in the OPG-KO mice. The bone structure of mice in
the DO and DO + OPG-KO groups was more fragile and chaotic,
compared to the normal group. The bone collagen structure of mice
in the DO and DO + OPG-KO groups was significantly reduced compared
to the normal group. These effects were more evident in the OPG-KO
mice. However, treatment with BP attenuated these effects. These
data indicate that OPG gene knockout aggravates osteoporosis, and
that BP significantly suppresses osteoporosis.
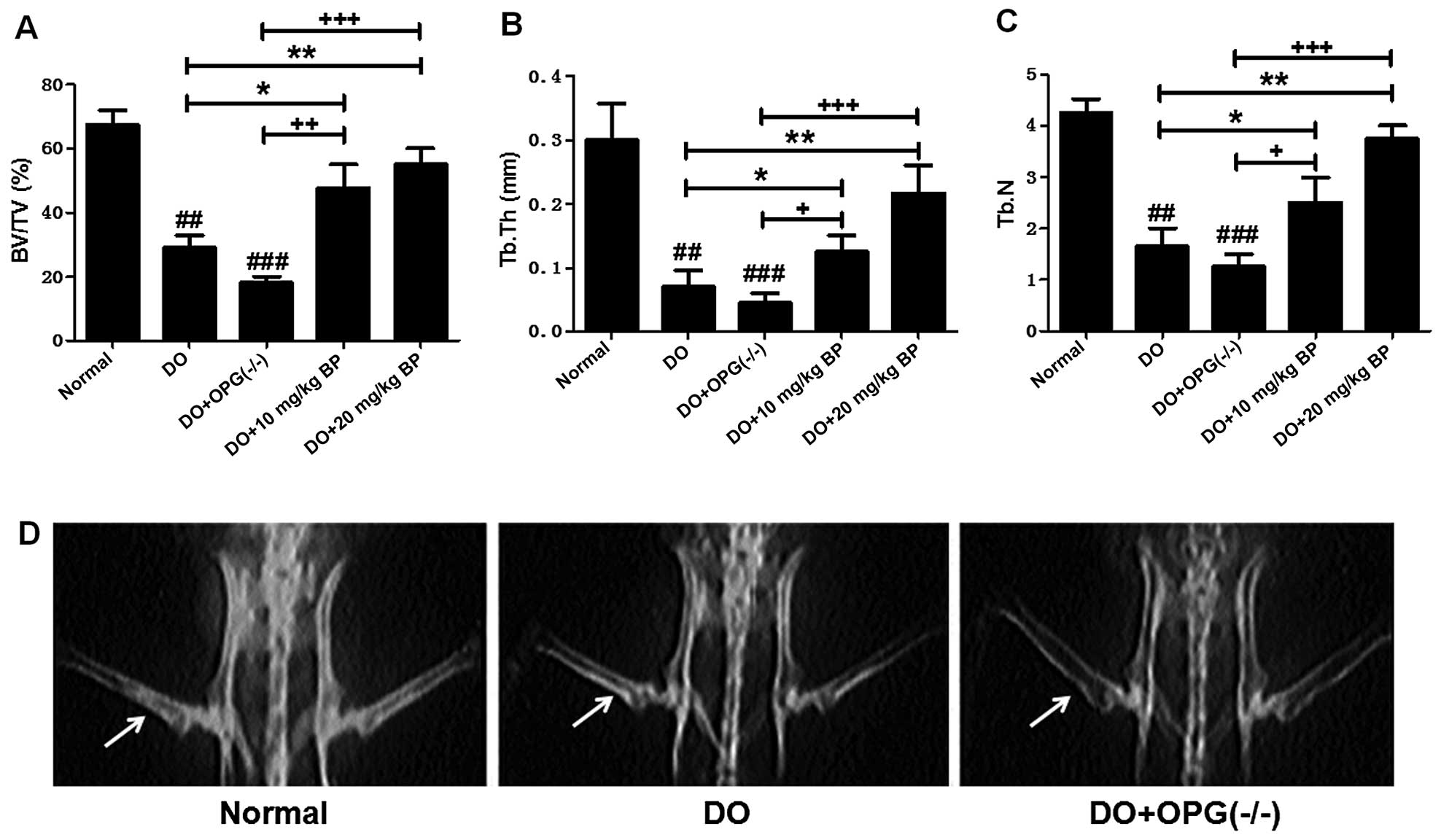 | Figure 2Effects of osteoprotegerin (OPG)
knockout on diabetes-related osteoporosis (DO). (A) Analysis of
bone volume fraction (BV/TV) in mice in the normal, DO, DO +
OPG-KO, DO + 10 mg/kg bergapten (BP) and DO + 20 mg/kg BP groups.
(B) Analysis of trabecular thickness (Tb.Th) in mice in the normal,
DO, DO + OPG-KO, DO + 10 mg/kg BP and DO + 20 mg/kg BP groups. (C)
Analysis of trabecular number (Tb.N) in mice in the normal, DO, DO
+ OPG-KO, DO + 10 mg/kg BP and DO + 20 mg/kg BP groups.
##p<0.01 and ###p<0.001 vs. normal
group; *p<0.05; +p<0.05,
++p<0.01 and +++p<0.001. (D) X-ray
scanning for determining bone mineral density. The arrows in the
images indicate the bone mineral density. |
Inhibitory effects of BP on the
activation of DO-related signaling pathways
As we mentioned above, in high-fat diet-induced DO,
the PI3K/AKT, JNK/MAPK and NF-κB signaling pathways have been
proven to be a possible target for the inhibition of the
development and progression of DO. However, the underlying
molecular mechanisms of action of these pathways in DO remain
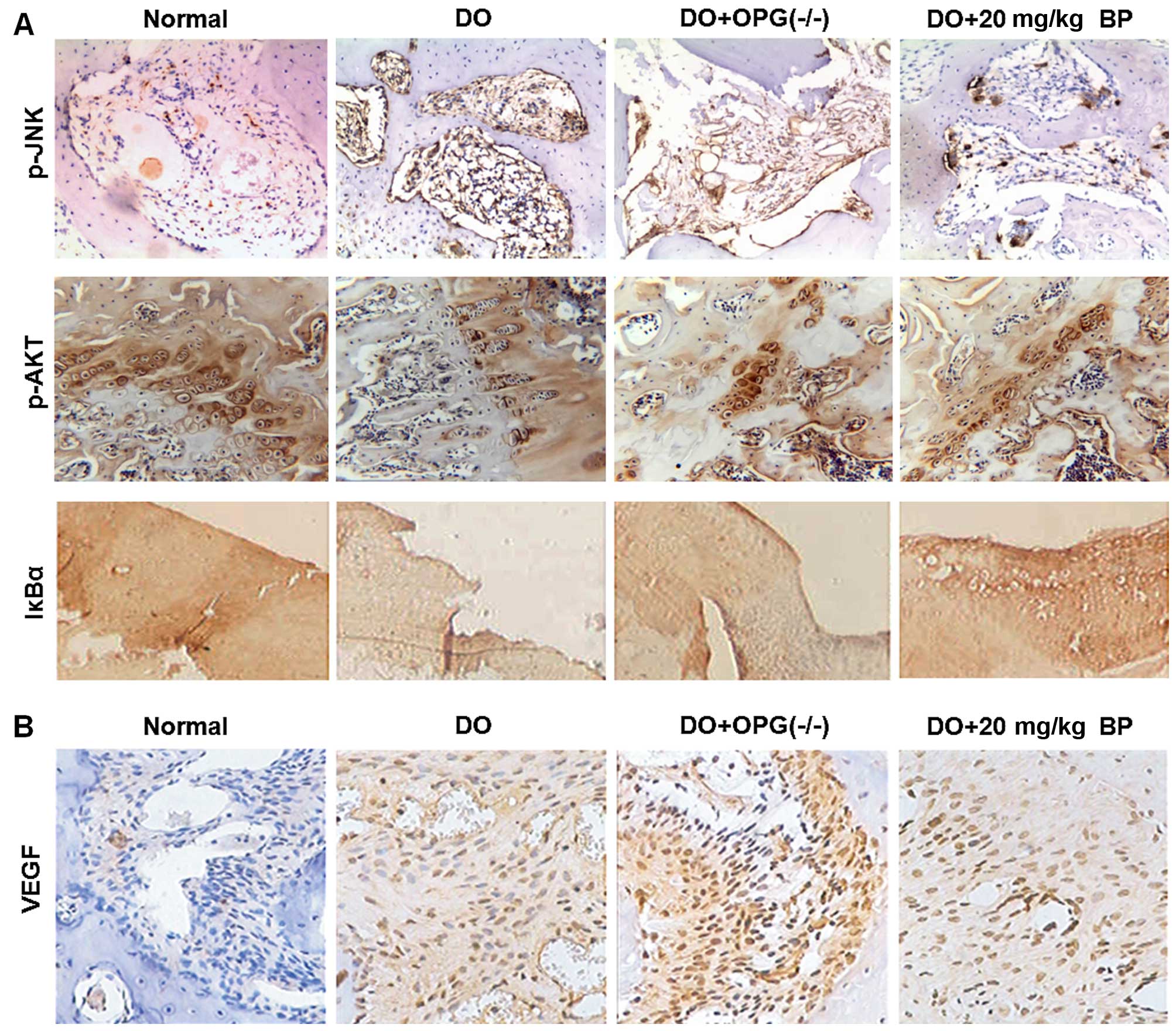
unknown. Thus in this regard, we used IHC to determine the
experssion of p-AKT, p-JNK and the key factor of NF-κB, IκBα, the
mice in the normal, DO, DO + OPG-KO and DO + 10/20 mg/kg BP groups.
As shown in Fig. 4A, the
expression of p-AKT, p-JNK and IκBα of NF-κB in the DO and DO +
OPG-KO groups was significantly higher compared with normal and BP
treatment groups. In particular, when comparing the DO group with
the DO + OPG-KO group, the DO + OPG-KO group exhibited a higher
expressoin of p-AKT, p-JNK and IκBα than the DO group, which
indicates that OPG has a protective effect against osteoporosis and
that BP has a similar effect to OPG in attenuating DO. In addition,
DO promoted the activation of the PI3K/AKT, JNK/MAPK and NF-κB
signaling pathways, enhancing osteoporosis. As shown in Fig. 4B, VEGF experssion was higher the
DO and DO + OPG-KO groups compared with the normal group,
indicating that greater bone loss occurred in the DO and OPG-KO
mice, and this was suppressed by treatment with BP.
OPG knockout promotes the activation of
the PI3K/AKT, JNK/MAPK and NF-κB signaling pathways in mice with
DO
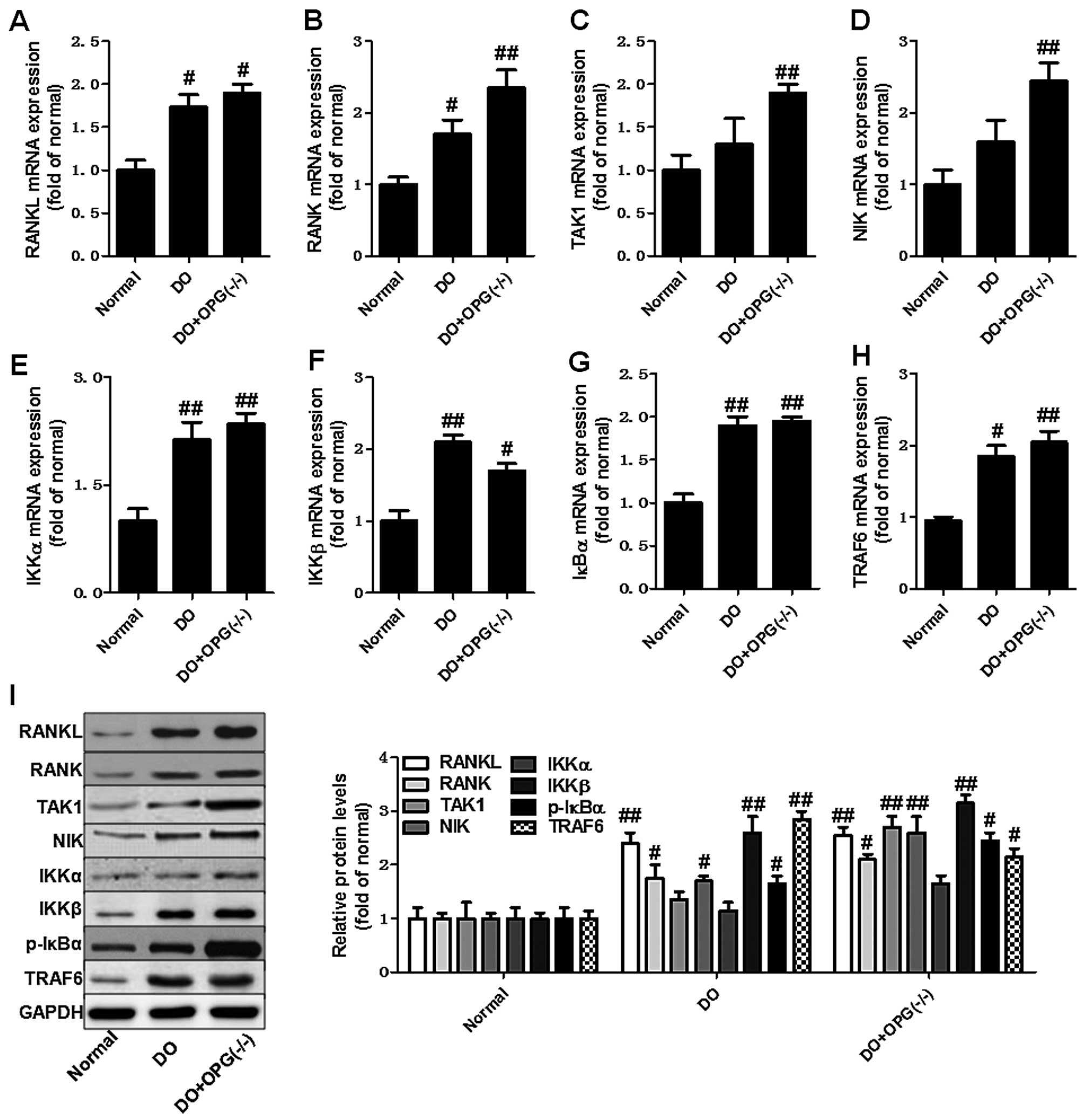
We then used qPCR and western blot analysis to
examine the expression of related indicators of the PI3K/AKT,
JNK/MAPK and NF-κB signaling pathways in OPG-KO mice. As shown in
Fig. 5A–H, the mRNA expression
levels of RANKL, RANK, TAK1, NIK, IKKα, IKKβ, IκBα and TRAF6 were
significantly increased in the OPG-KO group compared to the normal
group. In addition, the western blot analysis further indicated
that the protein expression levels of RANKL, RANK, TAK1, NIK, IKKα,
IKKβ, IκBα and TRAF6 also were further markedly upregulated in the
DO + OPG-KO group, at almost higher levels than the DO group
(Fig. 5I). This indicates that
OPG-KO significantly enhances the activation of the NF-κB signaling
pathway, thus also promoting the development of DO. Moreover, we
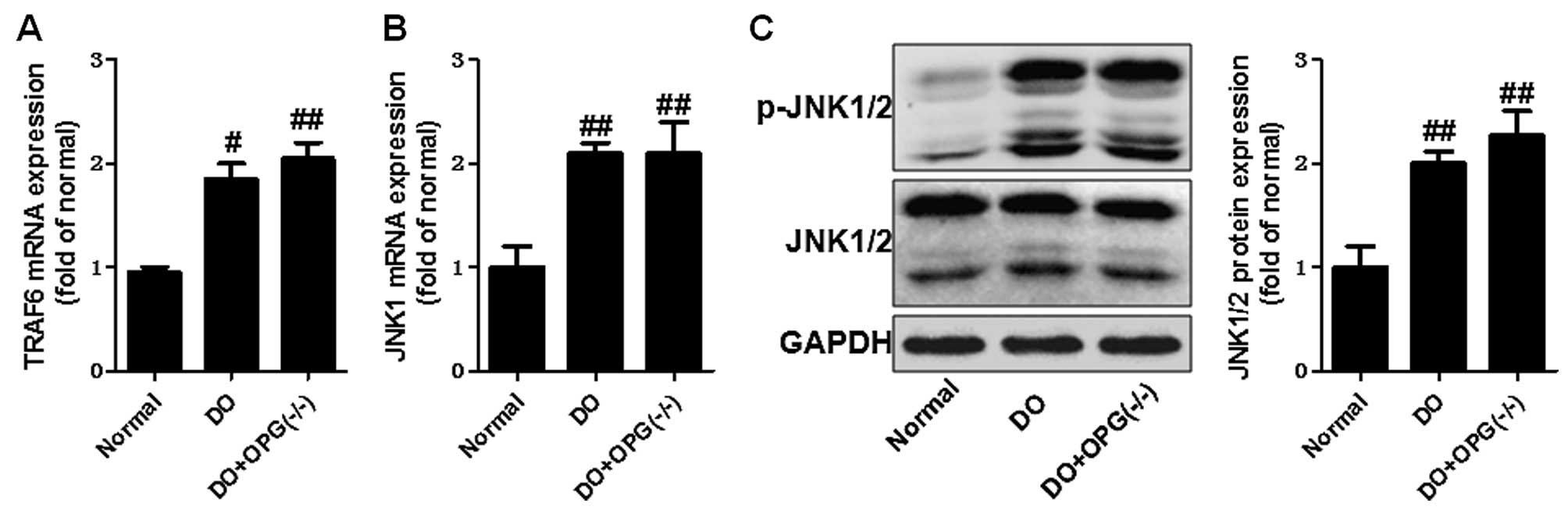
also examined the role of the JNK pathway in DO. As shown in
Fig. 6, the JNK pathway was
activated in the OPG-KO and DO groups. The levels of p-JNK1/2 were
significantly increased in the DO + OPG-KO group compared with the
normal group, which demonstrates that the JNK pathway is indirectly
or directly involved in the development and progression of DO. In
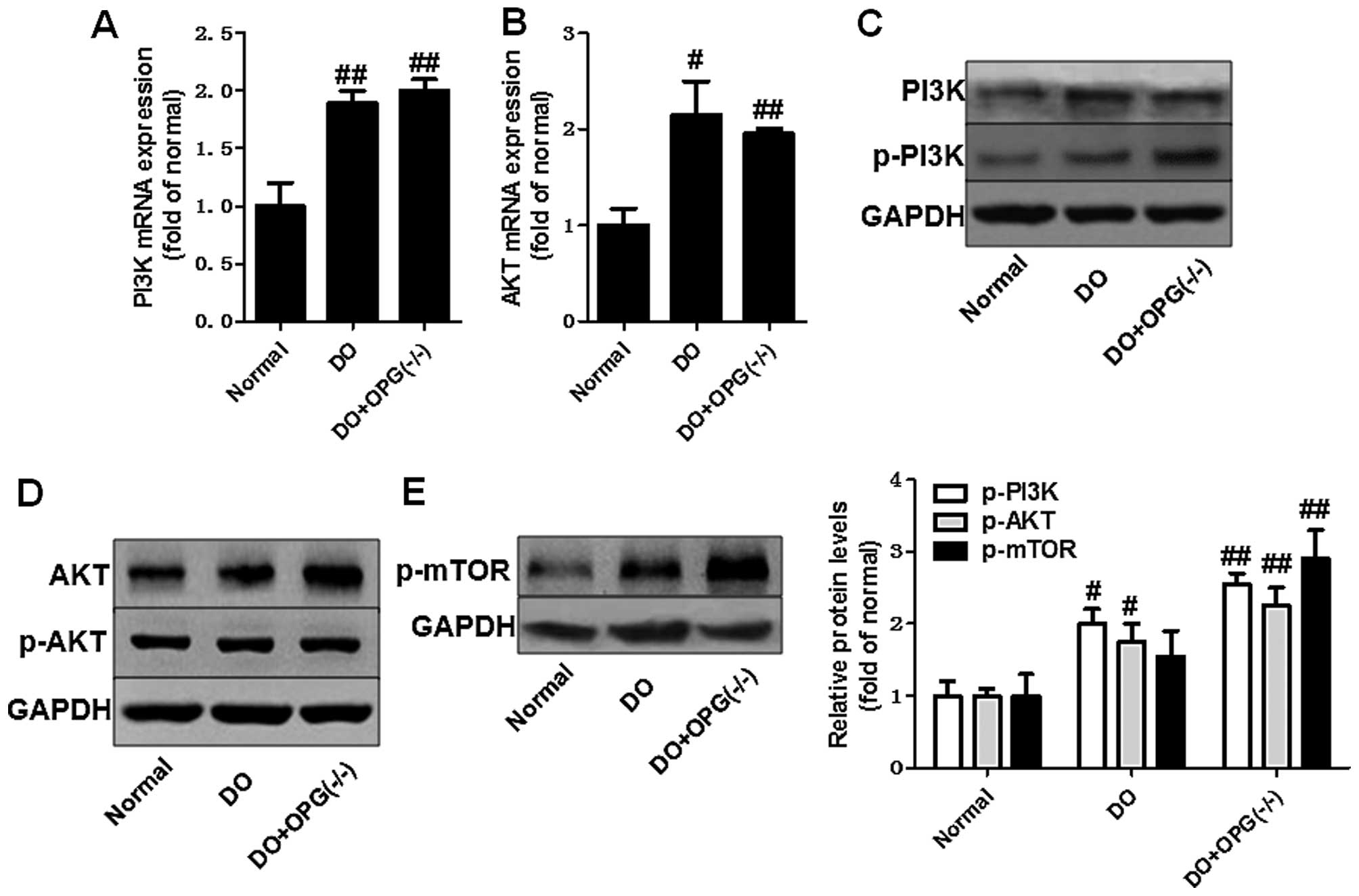
addition, we further investigated the role of the AKT signaling
pathway in the development of osteoporosis. The data shown in
Fig. 7 indicated that the AKT
pathway was activated in the OPG-KO mice. The levels of p-AKT and
p-mTOR were significantly increased compared to the normal group.
These data indicate that the PI3K/AKT, JNK/MAPK and NF-κB signaling
pathways are indirectly or directly involved in the development of
DO.
BP suppresses the activation of the
PI3K/AKT, JNK/MAPK and NF-κB signaling pathways in mice with
DO
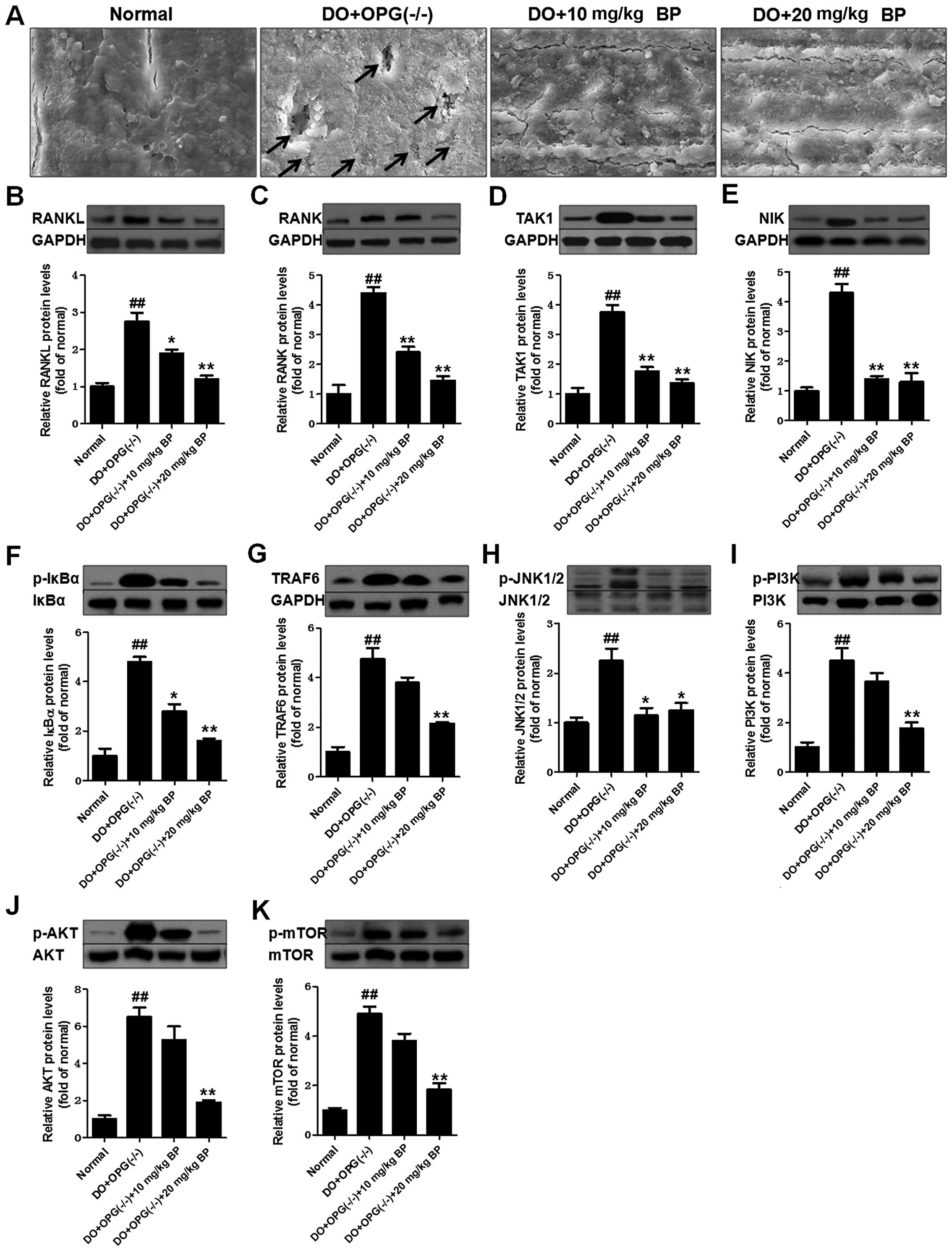
We examined the suppressive effects of BP on the
activation of the PI3K/AKT, JNK/MAPK and NF-κB signaling pathways
in our mouse model of DO. As shown in Fig. 8A, scanning electron microscopy
revealed that treatment with BP (10 and 20 mg/kg) attenuated the
development of DO, as evidenced by less surface bone damage in the
BP-treated groups compared with the DO + OPG-KO group. Osteoclast
activation was enhanced in the DO + OPG-KO group and was
significantly inhibited by BP (Fig.
8A). Western blot analysis demonstrated that the levels of
related indicators of the PI3K/AKT, JNK/MAPK and NF-κB signaling
pathways were significantly suppressed by BP, compared to the DO +
OPG-KO group. The inhibitory effects of BP on the levels of these
indicators were observed in a dose-independent manner. These data
suggest that BP is capable of inhibiting the activation of the
PI3K/AKT, JNK/MAPK and NF-κB signaling pathways.
Discussion
Diabetes is regarded as a global threat to human
health. Diabetes is a metabolic disorder which causes a dysfunction
in the production of insulin, a hormone that is required to convert
sugar, starches and other foods into energy. According to published
data, it is estimated that >50 million individuals are already
affected by diabetes, and that 11.6 million diabetics are not aware
of the existing disease (1–4).
In our current understanding, there are 3 main types of diabetes
which have been confirmed: type 1 diabetes, type 2 diabetes and
gestational diabetes. Diabetes is the primary reason for adult
blindness, end-stage renal disease (ESRD), gangrene and amputations
(22–25). Being overweight, lack of exercise,
family history and stress increase the likelihood of developing
diabetes. Constantly high blood sugar levels lead to kidney
failure, cardiovascular problems and neuropathy (26,27). Patients with diabetes are 4-fold
more likely to suffer from coronary heart disease and stroke. In
addition, gestational diabetes is more dangerous for pregnant women
and their fetus (28). However,
more seriously, in recent years, global researchers have paid more
attention to diabetes-related disorders (29,30). At present, there is increasing
concern about the many acute and chronic complications of diabetes,
among which the association between diabetes and osteoporosis is
attracting more attention. DO is a systemic metabolic disease
affecting the bone and has an intricate and complex
pathogenesis.
DO easily leads to fracture, has a high disability
rate and is a serious threat to human health. It has been found
that diabetes affects bone metabolism through various mechanisms,
including insulin dysfunction, high blood glucose, increased
accumulation of advanced glycation end products and microangiopathy
(31,32). OPG as an important protein for the
preventation of osteoporosis and has been proven to be key to
suppressing the development of DO. In this regard, in this study,
we used a mouse model of DO a model of DO with OPG knockout mice to
investigate the related signaling pathways involved in the
development and progression of osteoporosis. In addition, BP as a
natural anticancer and anti-inflammatory agent isolated from
bergamot essential oil, other citrus essential oils and grapefruit
juice, was used to examine its protective effects against DO and
whether it has a similar effect to OPG in inhibiting
osteoporosis.
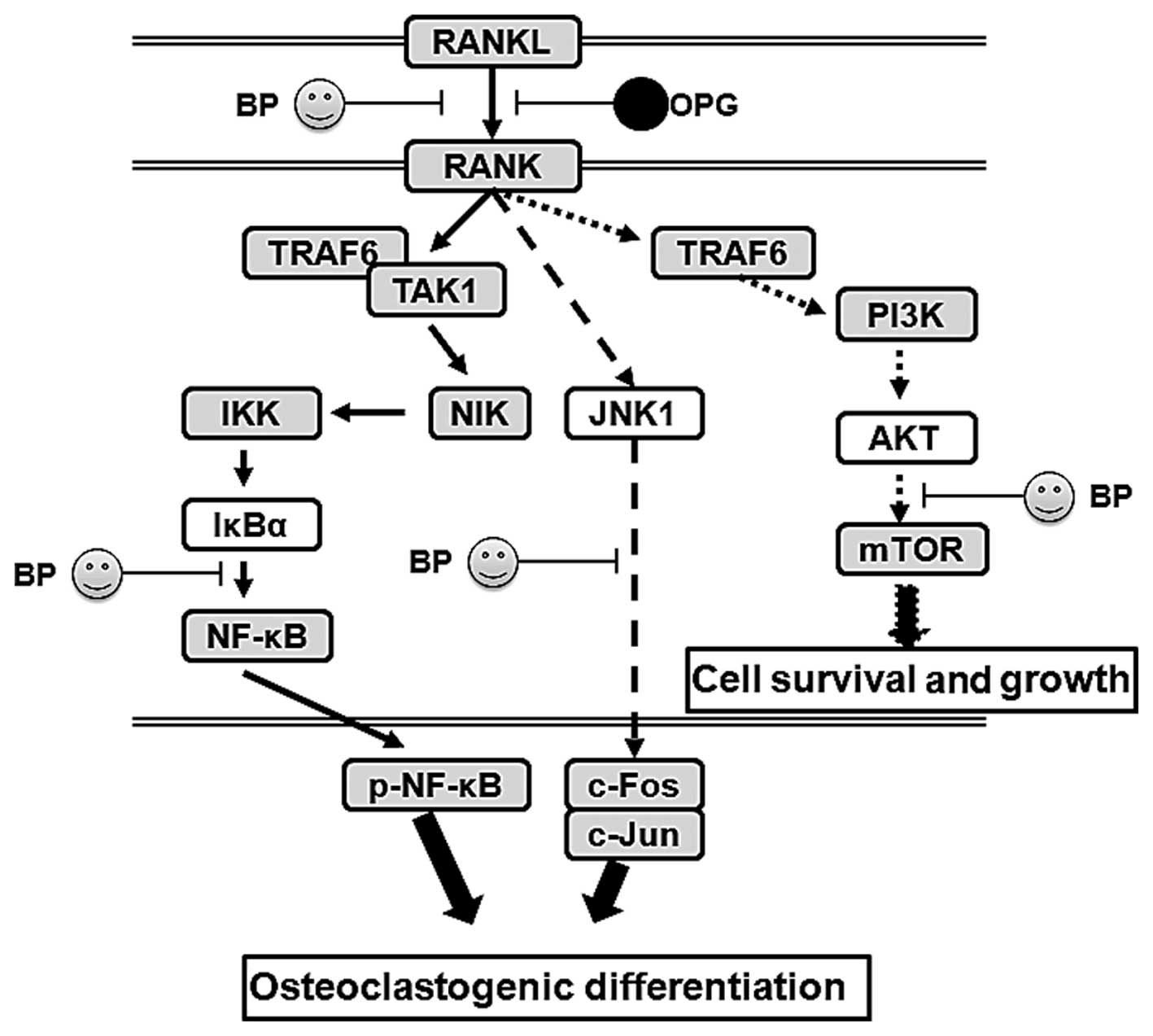
The PI3K/AKT and related pathways play an important
role in internalizing the effects of external growth factors and
membrane tyrosine kinases. As shown in Fig. 9, in DO, many downstream indicators
and pathways are activated, such as NF-κB, mTOR, Bad and FKHR. The
activation of PI3K/AKT further enhances inflammatory cyokine
expression and signaling pathway transduction, particularly
regarding NF-κB. Our data found that DO upregulate the level of
phosphorylated AKT and activates the downstream mTOR pathway, which
promotes the development of osteoporosis. In addition, OPG-KO
significantly enhanced the transduction between RANKL and RANK,
further activating the AKT pathway. On the one hand, activated AKT
further enhances the phosphorylated NF-κB level, leading to the
increased production and and release of inflammatory cytokines. On
the other hand, our results revealed that OPG-KO in DO
significantly and directly increases the level of phosphorylated
NF-κB through NIK/IKK/IκBα to promote nuclear transduction. Thus,
through these two mechanisms, phosphorylated NF-κB is indirectly or
directly activated and this causes osteoclastogenic differentiation
and loss of ossein, aggravating trabecular bone injury.
Furthermore, JNK, as the major pathway in the MAPK family, has been
proven to be involved in the development and progression of various
diseases (33,34). In this study, our data
demonstrated that the MAPK/JNK signaling pathway is involved in the
development of osteoporosis. It enhances c-Jun epxression and
promotes osteoclastogenic differentiation, in association with
increased levels of phosphorylated NF-κB, thus promoting the
development of DO. We also investigated whether the
anti-inflammatory agent, BP, exerts effects similar to those of OPG
in mice with DO and with OPG-KO. Our data indicated that BP has the
ability to indirectly or directly suppress the activation of the
NF-κB, JNK and AKT/mTOR signaling pathways. It prevents trabecular
bone injury and suppresses the loss of collagen.
In conclusion, in this study, we examined the
protective effects of BP using mouse models of DO and and DO with
OPG-KO. The DO and DO in conjunction with OPG-KO significantly
promote the activation of the PI3K/AKT, JNK/MAPK and NF-κB
signaling pathways and increases the expression of related
indicators, further promoting the release inflammatory cytokines,
which damages trabecular bone structure and promotes
osteoclastogenic differentiation.
References
|
1
|
Marcus R: Post-menopausal osteoporosis.
Best Pract Res Clin Obstet Gynaecol. 16:309–327. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mullin BH, Prince RL, Dick IM, Hart DJ,
Spector TD, Dudbridge F and Wilson SG: Identification of a role for
the ARHGEF3 gene in postmenopausal osteoporosis. Am J Hum Genet.
82:1262–1269. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Reppe S, Refvem H, Gautvik VT, Olstad OK,
Høvring PI, Reinholt FP, Holden M, Frigessi A, Jemtland R and
Gautvik KM: Eight genes are highly associated with BMD variation in
postmenopausal Caucasian women. Bone. 604–612. 2010. View Article : Google Scholar
|
|
4
|
Jemtland R, Holden M, Reppe S, Olstad OK,
Reinholt FP, Gautvik VT, Refvem H, Frigessi A, Houston B and
Gautvik KM: Molecular disease map of bone characterizing the
postmenopausal osteoporosis phenotype. J Bone Miner Res.
26:1793–1801. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chao TH, Yu HN, Huang CC, Liu WS, Tsai YW
and Wu WT: Association of interleukin-1 β (−511C/T) polymorphisms
with osteoporosis in postmenopausal women. Ann Saudi Med.
30:437–441. 2010.PubMed/NCBI
|
|
6
|
Finkelstein JS, Brockwell SE, Mehta V,
Greendale GA, Sowers MR, Ettinger B, Lo JC, Johnston JM, Cauley JA,
Danielson ME and Neer RM: Bone mineral density changes during the
menopause transition in a multiethnic cohort of women. J Clin
Endocrinol Metab. 93:861–868. 2008. View Article : Google Scholar
|
|
7
|
Roy DK, Berry JL, Pye SR, Adams JE,
Swarbrick CM, King Y, Silman AJ and O'Neill TW: Vitamin D status
and bone mass in UK South Asian women. Bone. 40:200–204. 2007.
View Article : Google Scholar
|
|
8
|
Bone HG, Greenspan SL, McKeever C, Bell N,
Davidson M, Downs RW, Emkey R, Meunier PJ, Miller SS, Mulloy AL, et
al: Alendronate and estrogen effects in postmenopausal women with
low bone mineral density. Alendronate/Estrogen Study Group. J Clin
Endocrinol Metab. 85:720–726. 2000.PubMed/NCBI
|
|
9
|
Slemenda C, Longcope C, Peacock M, Hui S
and Johnston CC: Sex steroids, bone mass, and bone loss. A
prospective study of pre-, peri-, and postmenopausal women. J Clin
Invest. 97:14–21. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wolff I, van Croonenborg JJ, Kemper HC,
Kostense PJ and Twisk JW: The effect of exercise training programs
on bone mass: a meta-analysis of published controlled trials in
pre- and postmenopausal women. Osteoporos Int. 9:1–12. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Clowes JA, Riggs BL and Khosla S: The role
of the immune system in the pathophysiology of osteoporosis.
Immunol Rev. 208:207–227. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cavalli L and Brandi ML: Age- and
gender-related macro- and micro-architecture changes in bone
structure and implications for treatment. Int J Clin Rheumatol.
6:359–369. 2011. View Article : Google Scholar
|
|
13
|
Sanders S and Geraci SA: Osteoporosis in
postmenopausal women: Considerations in prevention and treatment:
(Women's health series). South Med J. 106:698–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hofbauer LC, Brueck CC, Singh SK and
Dobnig H: Osteoporosis in patients with diabetes mellitus. J Bone
Miner Res. 22:1317–1328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leidig-Bruckner G and Ziegler R: Diabetes
mellitus a risk for osteoporosis? Exp Clin Endocrinol Diabetes.
109(Suppl 2): S493–S514. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McLean RR: Proinflammatory cytokines and
osteoporosis. Curr Osteoporos Rep. 7:134–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huh JE, Lee WI, Kang JW, Nam D, Choi DY,
Park DS, Lee SH and Lee JD: Formononetin attenuates
osteoclastogenesis via suppressing the RANKL-induced activation of
NF-κB, c-Fos, and nuclear factor of activated T-cells cytoplasmic 1
signaling pathway. J Nat Prod. 77:2423–2431. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Oh KW: Diabetes and Osteoporosis. Korean
Diabetes J. 33:169–177. 2009.In Korean. View Article : Google Scholar
|
|
19
|
Zheng M, Ge Y, Li H, Yan M, Zhou J and
Zhang Y: Bergapten prevents lipopolysaccharide mediated osteoclast
formation, bone resorption and osteoclast survival. Int Orthop.
38:627–634. 2014. View Article : Google Scholar :
|
|
20
|
Perez RM: Anti-inflammatory activity of
compounds isolated from plants. Scientific World Journal.
1:713–784. 2001. View Article : Google Scholar
|
|
21
|
Peng X, Nie Y, Wu J, Huang Q and Cheng Y:
Juglone prevents metabolic endotoxemia-induced hepatitis and
neuroinflammation via suppressing TLR4/NF-κB signaling pathway in
high-fat diet rats. Biochem Biophys Res Commun. 462:245–250. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Holstein P, Ellitsgaard N, Olsen BB and
Ellitsgaard V: Decreasing incidence of major amputations in people
with diabetes. Diabetologia. 43:844–847. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Romero-Aroca P: Managing diabetic macular
edema: The leading cause of diabetes blindness. World J Diabetes.
2:98–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shoji T, Emoto M, Shinohara K, Kakiya R,
Tsujimoto Y, Kishimoto H, Ishimura E, Tabata T and Nishizawa Y:
Diabetes mellitus, aortic stiffness, and cardiovascular mortality
in end-stage renal disease. J Am Soc Nephrol. 12:2117–2124.
2001.PubMed/NCBI
|
|
25
|
Schneider PA, Caps MT, Ogawa DY and Hayman
ES: Intraoperative superficial femoral artery balloon angioplasty
and popliteal to distal bypass graft: an option for combined open
and endovascular treatment of diabetic gangrene. J Vasc Surg.
33:955–692. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Balkau B, Shipley M, Jarrett RJ, Pyörälä
K, Pyörälä M, Forhan A and Eschwège E: High blood glucose
concentration is a risk factor for mortality in middle-aged
nondiabetic men. 20-year follow-up in the Whitehall Study, the
Paris Prospective Study, and the Helsinki Policemen Study. Diabetes
Care. 21:360–367. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sumner CJ, Sheth S, Griffin JW, Cornblath
DR and Polydefkis M: The spectrum of neuropathy in diabetes and
impaired glucose tolerance. Neurology. 60:108–111. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim C, Newton KM and Knopp RH: Gestational
diabetes and the incidence of type 2 diabetes: a systematic review.
Diabetes Care. 25:1862–1868. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moran C, Münch G, Forbes JM, Beare R,
Blizzard L, Venn AJ, Phan TG, Chen J and Srikanth V: Type 2
diabetes, skin autofluorescence, and brain atrophy. Diabetes.
64:279–283. 2015. View Article : Google Scholar
|
|
30
|
Zinman B, Lachin JM and Inzucchi SE:
Empagliflozin, cardiovascular outcomes, and mortality in type 2
diabetes. N Engl J Med. 374:10942016.PubMed/NCBI
|
|
31
|
Wu YY, Xiao E and Graves DT: Diabetes
mellitus related bone metabolism and periodontal disease. Int J
Oral Sci. 7:63–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li R, Xu W, Luo S, Xu H, Tong G, Zeng L,
Zhu D and Weng J: Effect of exenatide, insulin and pioglitazone on
bone metabolism in patients with newly diagnosed type 2 diabetes.
Acta Diabetol. 52:1083–1091. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu QQ, Xu M, Yuan Y, Li FF, Yang Z, Liu Y,
Zhou MQ, Bian ZY, Deng W, Gao L, et al: Cathepsin B deficiency
attenuates cardiac remodeling in response to pressure overload via
TNF-α/ASK1/JNK pathway. Am J Physiol Heart Circ Physiol.
308:H1143–H1154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weston CR and Davis RJ: The JNK signal
transduction pathway. Curr Opin Cell Biol. 19:142–149. 2007.
View Article : Google Scholar : PubMed/NCBI
|