Introduction
Nobiletin, a major flavonoid polymethexylated
flavone (5,6,7,8,3′,4′-hexamethoxyflavone; PMF) found in citrus,
has been found in very high concentrations in immature citrus peels
(1). It can been extracted from
different citrus species, such as Citrus reticulata Blanco
(mandarin orange), Citrus unshiu Markovich, Citrus
depressa (shiikuwasa) and Citrus limon (lemon) (2–4).
Nobiletin has been reported to be an encouraging antioxidant and
anti-inflammatory agent in the treatment of asthma, colitis and
Alzheimer's disease (2,5,6).
Nobiletin has been reported to protect PC12 cells from hydrogen
peroxide (H2O2)-induced cytotoxicity
(7), and to atttenuate
ethanol-induced liver injury by augmenting the phosphorylation of
AMP-activated protein kinase (AMPK) (8). This compound possesses potent
anti-neuroinflammatory abilities by suppressing the activation of
the extracellular signal-regulated kinase (ERK), c-Jun N-terminal
kinase (JNK) and p38 mitogen-activated protein kinase (p38 MAPK)
pathways, as well as the translocation of nuclear factor-κB (NF-κB)
and the subsequent gene expression of inducible nitric oxide
synthase (iNOS), tumor necrosis factor-α (TNF-α) and interleukin
(IL)-1β (9). A previous study
demonstrated that nobiletin was the most potent inhibitor of
neuroinflammation among 8 common tangerine flavonoids (10). Another study revealed that the
administration of nobiletin protected rat brains from ischemic
damage by activating the Akt/cyclic AMP response element-binding
protein (CREB) signaling pathway (11).
Vasodilator-stimulated phosphoprotein (VASP) is a
regulator of actin reorganization in platelets. As VASP is a common
downstream target of various signaling pathways, an increasing
attention to this molecule has been paid in platelet studies
(12–14). Cyclic AMP-dependent protein kinase
(PKA; also known as protein kinase A) is considered the main
mediator of the numerous effects associated with increased cyclic
AMP levels. In platelets, PKA activation has been shown to be
involved in the phosphorylation of VASP (15). It is also noteworthy that cyclic
AMP may cross-activate the cyclic GMP-dependent protein kinase
(PKG; also known as protein kinase G) in some vascular tissues
(16). Ii has also been shown
that PKA and PKG are not the only kinases able to phosphorylate
VASP, but that protein kinase C (PKC) may also have this ability
(17). VASP phosphorylation in
response to cyclic AMP/cyclic GMP in platelets correlates with
fibrinogen receptor inhibition (18). In addition, platelets from
VASP-knockout mice exhibit enhanced thrombin-induced platelet
activation and impaired cyclic AMP-dependent inhibition (19). These studies suggest an important
role of VASP in signal transduction pathways in platelets, its
phosphorylation closely correlating with adenylate cyclase
stimulation, platelet cyclic AMP/cyclic GMP increase, and the
inhibition of platelet aggregation.
In our previous study, we found that nobiletin
(10–30 µM) inhibited collagen and arachidonic acid-induced
platelet aggregation in a concentration-dependent manner through
the inhibition of the activation of the phospholipase C (PLC)
γ2-PKC cascade and Akt/MAPK signaling pathways, and also found that
nobiletin prolonged closure time in human whole blood ex
vivo and increased the occlusion time of thrombotic platelet
plug formation in mice (20).
Moreover, at a maximum concentration of 30 µM, nobiletin had
no effects on the intracellular levels of cyclic AMP or cyclic GMP
in human platelets, which is consistent with the recent observation
by Vaiyapuri et al (21).
Of note, our preliminary experiments revealed that nobiletin (30
µM) markedly increased VASP phosphorylation in a similar
manner to prostaglandin E1 (PGE1) (Fig. 1A). Therefore, it is of
considerable interest to understand what the novel mechanisms
responsible for nobiletin-induced VASP phosphorylation, since
cyclic nucleotides do not play a role in this process.
Materials and methods
Chemicals and reagents
Nobiletin (≥97%), collagen (type I),
5,5-dimethyl-1-pyrroline N-oxide (DMPO), SQ22536 (inhibitor of
adenylate cyclase), 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one
(ODQ; inhibitor of guanylate cyclase), heparin, PGE1,
nitroglycerin (NTG), H89 (inhibitor of PKA), KT583 (PKG inhibitor),
LY294002 (Akt inhibitor), Ro318220 (PKC inhibitor), DPI (NOX
inhibitor), Bay11-7082 (NF-κB inhibitor), PD98059 (ERK2 inhibitor),
SB203580 (p38 MAPK inhibitor, SP600125 (JNK inhibitor) and
N-acetylcysteine (NAC) were all purchased from Sigma-Aldrich (St.
Louis, MO, USA). Anti-VASP (Cat. no. GTX132176) and anti-VASP
(phospho Ser157; Cat. no. GTX32362) polyclonal antibodies (pAbs)
were purchased both from GeneTex, Inc. (Irvine, CA, USA). The
anti-α-tubulin monoclonal antibody (mAb; Cat. no. MS-581-P0) was
purchased from NeoMarkers (Fremont, CA, USA). Hybond-P
polyvinylidene difluoride (PVDF) membranes, an enhanced
chemiluminescence western blotting detection reagent, the
horseradish peroxidase (HRP)-conjugated donkey anti-rabbit
immunoglobulin G (IgG; Cat. no. RPN4301), and the sheep anti-mouse
IgG (Cat. no. RPN4201) were all purchased from GE Healthcare UK
Ltd. (Buckinghamshire, UK). Nobiletin was dissolved in 0.5%
dimethyl sulfoxide (DMSO) and stored at 4°C.
Platelets
Our study was approved by the Institutional Review
Board of Taipei Medical University, Taipei, Taiwan and conformed to
the directives of the Helsinki Declaration. Human platelet
suspensions were prepared as previously described (22). Blood was collected from 20 healthy
human volunteers (who had given informed consent and had taken no
medication during the 2 weeks preceding collection) and was mixed
with an acid-citrate-dextrose solution. Following centrifugation at
120 × g for 37°C, the supernatant (platelet-rich plasma) was
supplemented with 0.5 µM PGE1 and 6.4 IU/ml of
heparin. The washed platelets were finally suspended in Tyrode's
solution containing 3.5 mg/ml of bovine serum albumin (BSA). The
final concentration of Ca2+ in Tyrode's solution was 1
mM.
Immunoblotting
The washed platelets (1.2×109 cells/ml)
were pre-incubated with 30 µM nobiletin or 0.5% DMSO for 3
min. The reaction was terminated by using 10 mM
ethylenediaminetetraacetic acid (EDTA), and the platelets were
immediately re-suspended in 200 µl of a lysis buffer.
Samples containing 80 µg of protein were separated on a 12%
acrylamide gel using sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), and the proteins were
electrotransferred onto the PVDF membranes by using a Bio-Rad
semidry transfer unit (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The blots were blocked with TBST (10 mM Tris-base, 100 mM
NaCl and 0.01% Tween-20) containing 5% BSA for 1 h and probed with
various primary antibodies. The membranes were incubated with the
HRP-linked anti-mouse IgG or anti-rabbit IgG (diluted 1:3,000 in
TBST) for 1 h. Immunoreactive bands were detected using an enhanced
chemiluminescence system. Ratios of the semi-quantitative results
were obtained by scanning the reactive bands and quantifying the
optical density using a video densitometer and Bio-profil Biolight
software, version V2000.01 (Vilber Lourmat, Marne-la-Vallée,
France).
Measurement of hydroxyl radicals (OH•)
using electron spin resonance (ESR) spectrometry
ESR spectrometry was performed using a Bruker EMX
ESR spectrometer (Bruker, Billerica, MA, USA) as previously
described (23). The platelet
suspensions (3.6×108 cells/ml) were treated with 1
µg/ml of collagen, 500 µM nobiletin or 0.5% DMSO for
3 min in a separate vial. The suspensions were incubated for 5 min,
and 100 µM DMPO were added before ESR was conducted. The ESR
spectrometer was operated at a power of 20 mW and 9.78 GHz, and a
scan range of 100 G and a receiver gain of 5×104 was
applied. Moreover, a Fenton reaction solution (50 µM
FeSO4 + 2 mM H2O2) was also
pre-treated with a solvent control (0.5% DMSO) or nobiletin (500
µM) for 3 min. The rate of free-radical generation is
expressed in the following equation: inhibition rate = 1 − [signal
height (nobiletin)/signal height (solvent control)], as previously
described (23).
Measurement of intracellular reactive
oxygen species (ROS) by fluorometric assay
The method, based on a fluorometric assay, was
adapted to measure the intracellular ROS. Briefly, the platelet
suspensions (3.6×108 cells/ml) were incubated (20 min,
37°C in the dark) in 24-well microtiter plates with 20 µM of
dichlorofluorescein fluorescence (DCF) stock in ethanol. The
content of each well was then transferred into a 5 ml tube for
centrifugation (335 × g, 10 min, 37°C). After removing the
supernatant, the suspensions were subsequently resuspended in
Hank's Balanced Salt Solution (HBSS) buffer and transferred to the
wells. Following treatment with various concentrations (50, 100,
and 500 µM) of nobiletin and 1 µg/ml of collagen, the
generation of the fluorescent product, DCF, was followed in an
automated plate fluorescence reader (Beckman Coulter, Miami, FL,
USA) using an excitation wavelength of 485 nm and emission
wavelength of 555 nm. The effect of nobiletin and collagen on ROS
production was compared to the resting platelets set as 100% of the
ROS-induced fluorescence.
Detection of protein carbonyls
The expression of protein carbonyl was monitored in
2,4-dinitrophenylhydrazine (DNPH)-derivatized proteins using an
OxyBlot Protein Oxidation Detection kit (Millipore, Billerica, MA,
USA). Briefly, the washed platelets (1.2×109 cells/ml)
were incubated with collagen (1 µg/ml) and nobiletin (50–500
µM) or 0.5% DMSO for 3 min. The reaction was terminated by
using EDTA, and the platelets were immediately re-suspended in 200
µl of a lysis buffer. Samples containing 30 µg of
protein were separated by electrophoresis on 12% SDS-PAGE, and
western blotting procedures were followed as described in
immunoblotting assay. A primary rabbit monoclonal antibody (1:150)
against DNP (Cat. no. S7150; Millipore) and a secondary anti-rabbit
IgG antibody (1:300) were used to detect protein carbonyl.
Statistical analysis
The experimental results are expressed as the means
± SEM and are accompanied by the number of observations (n). Values
of 'n' refer to the number of experiments, and each experiment was
conducted using different blood donors. Differences between groups
in the experiments were assessed using an analysis of variance
(ANOVA). When this analysis indicated significant differences among
group means, the groups were compared using the
Student-Newman-Keuls method. A value of P<0.05 was considered to
indicated a statistically significant difference. Statistical
analyses were performed using SAS version 9.2 (SAS Institute Inc.,
Cary, NC, USA).
Results
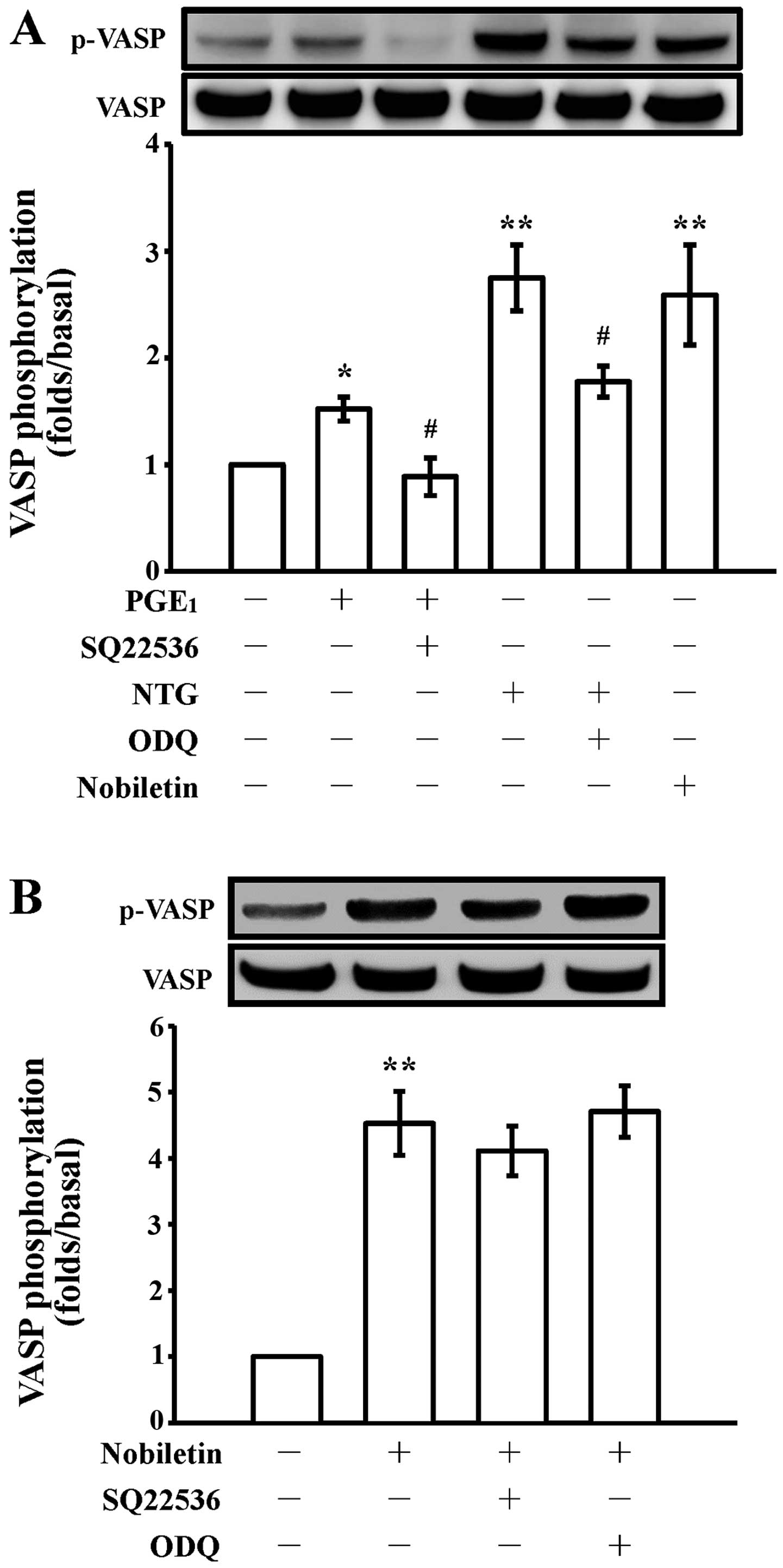
Effects of cyclic nucleotides on
nobiletin-induced VASP phosphorylation in washed human
platelets
It has been demonstrated that cyclic nucleotides can
induce VASP Ser157 phosphorylation in human platelets (24). Thus, in the present study, we
investigated whether the nobiletin-induced VASP phosphorylation is
dependent on cyclic nucleotides. The phosphorylation level of VASP
in unstimulated platelets was at lower levels; however, the
addition of 1 nM PGE1 and 10 µM NTG and 30
µM nobiletin significantly increased this phosphorylation
compared with the resting group (Fig.
1A). Moreover, VASP phosphorylation was absolutely abolished by
pre-treatment with SQ22536 (100 µM), an inhibitor of
adenylate cyclase or ODQ (10 µM), an inhibitor of guanylate
cyclase. Moreover, pre-treatment with SQ22536 or ODQ did not
significantly reverse the nobiletin (30 µM)-induced VASP
phosphorylation (Fig. 1B). These
results indicate that nobiletin-induced VASP phosphorylation in
human platelets is not dependent on cyclic nucleotide-related
pathways.
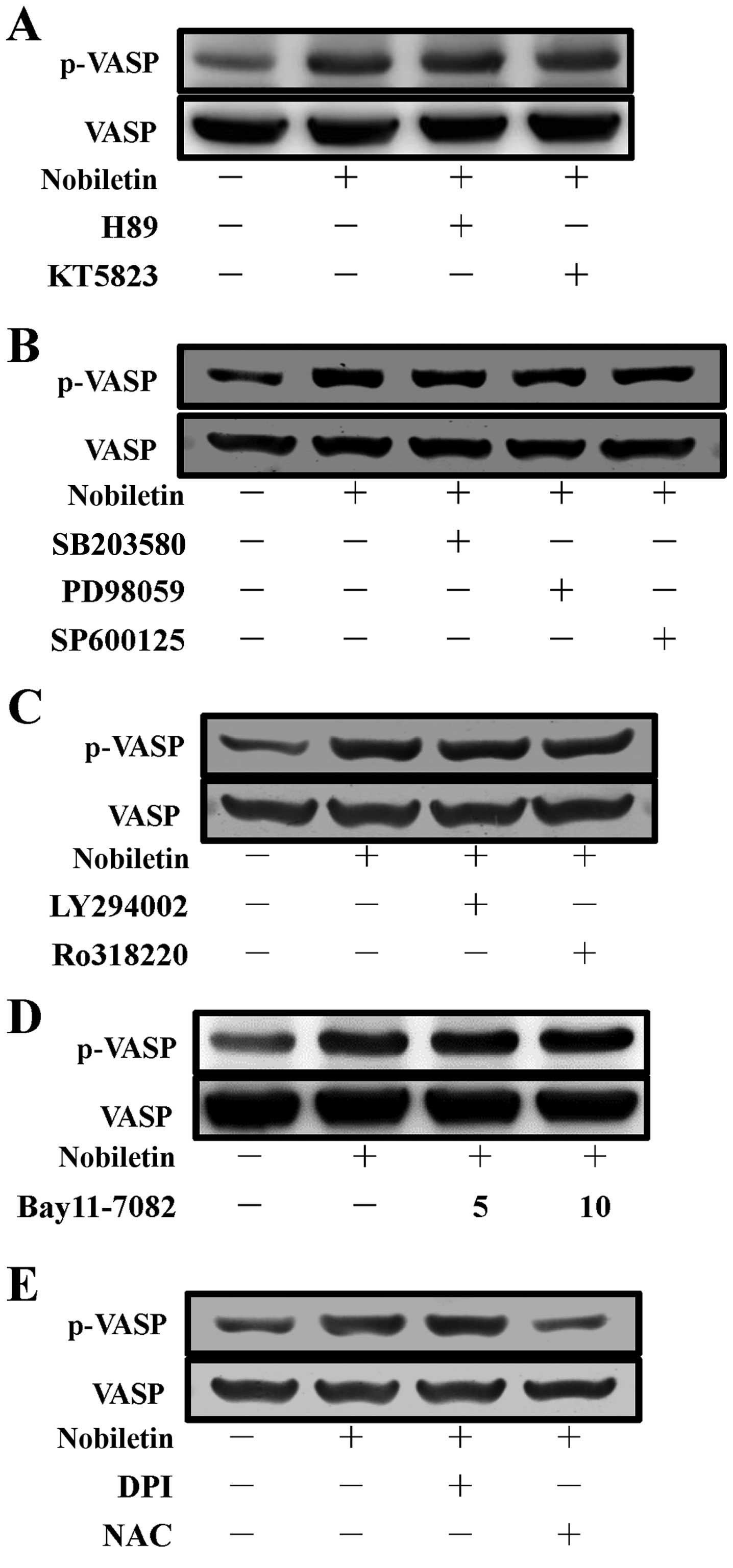
These results were further confirmed by using PKA
and PKG inhibitors, since it has previously been described that
VASP is a substrate of cyclic nucleotide (cyclic AMP/cyclic
GMP)-dependent PKA-PKG (25),
which phosphorylate it at 3 sites: Ser157, Ser239 and Thr278. These
sites are phosphorylated, with differing kinetics, both in
vitro and in intact human platelets (26). In the present study, as shown in
Fig. 2A, the inhibitors of PKA
(H89, 5 µM) and PKG (KT5823, 2 µM) were not
essentially effective in reversing the nobiletin-induced VASP
phosphorylation in platelets. These data indicate that PKA and PKG
are not involved in the regulation of nobiletin-induced VASP
phosphorylation.
MAPK, Akt and PKC have no essential
function in nobiletin-induced VASP phosphorylation
To investigate whether MAPKs, including p38 MAPK,
ERK and JNK, Akt and PKC, play a role in the stimulatory mechanisms
of nobiletin on VASP phosphorylation in human platelets, we used
the well-established inhibitors of p38 MAPK (SB203580, 10
µM), ERK (PD98059, 20 µM), JNK (SP600125, 10
µM), Akt (LY294002, 10 µM) and PKC (Ro318220, 2
µM) to examine their effects on VASP phosphorylation induced
by nobiletin. The results revealed that all the tested inhibitors
of p38 MAPK, ERK and JNK, Akt and PKC did not alter the
nobiletin-induced VASP phosphorylation (Fig. 2B and C), which indicate that the
MAPK, Akt and PKC signaling pathways do not play a role in
nobiletin-induced of VASP phosphorylation.
NF-κB is not responsible for
nobiletin-induced VASP phosphorylation in human platelets
NF-κB has been reported to function independently of
gene regulation in platelets, and follwoing platelet activation, it
is phosphorylated and degraded (27). NF-κB induction can be abrogated
using its inhibitor, Bay11-7082
[(E)3-[(4-methylphenyl)-sulfonyl]-2-propenenitrile], which is an
irreversible inhibitor of IκBα phosphorylation, resulting in the
downregulation of the induced NF-κB activation. Given the
significance of NF-κB and platelets in atherosclerosis and
inflammation, we investigated whether NF-κB plays a role in
nobiletin-induced VASP phosphorylation by using its inhibitor,
Bay11-7082. The data indicated that Bay11-7082 had no visible
effect on nobiletin-induced VASP phosphorylation (Fig. 2D), and consequently suggest that
NF-κB is not involved in the process of nobiletin-induced VASP
phosphorylation.
Involvement of NADPH oxidase (NOX) and
ROS in nobiletin-induced VASP phosphorylation
Since it has been previously demonstrated that NOX
and ROS regulate intracellular transduction pathways and many of
the structural properties of NOX have been found in platelets
(28), in the present study, the
effects of NOX inhibitor, diphenyleneiodonium chloride (DPI), and
the cell-permeable antioxidant, NAC, on nobiletin-induced VASP
phosphorylation in platelets were examined. We found that DPI (10
µM) did not affect nobiletin-induced VASP phosphorylation,
whereas, NAC (10 mM) significantly antagonized this phosphorylation
(Fig. 2E). These findings
indicate that the stimulatory effect of nobiletin on VASP
phosphorylation is also independent of NADPH oxidase, but that it
is dependent on NAC-mediated inhibitory mechanisms.
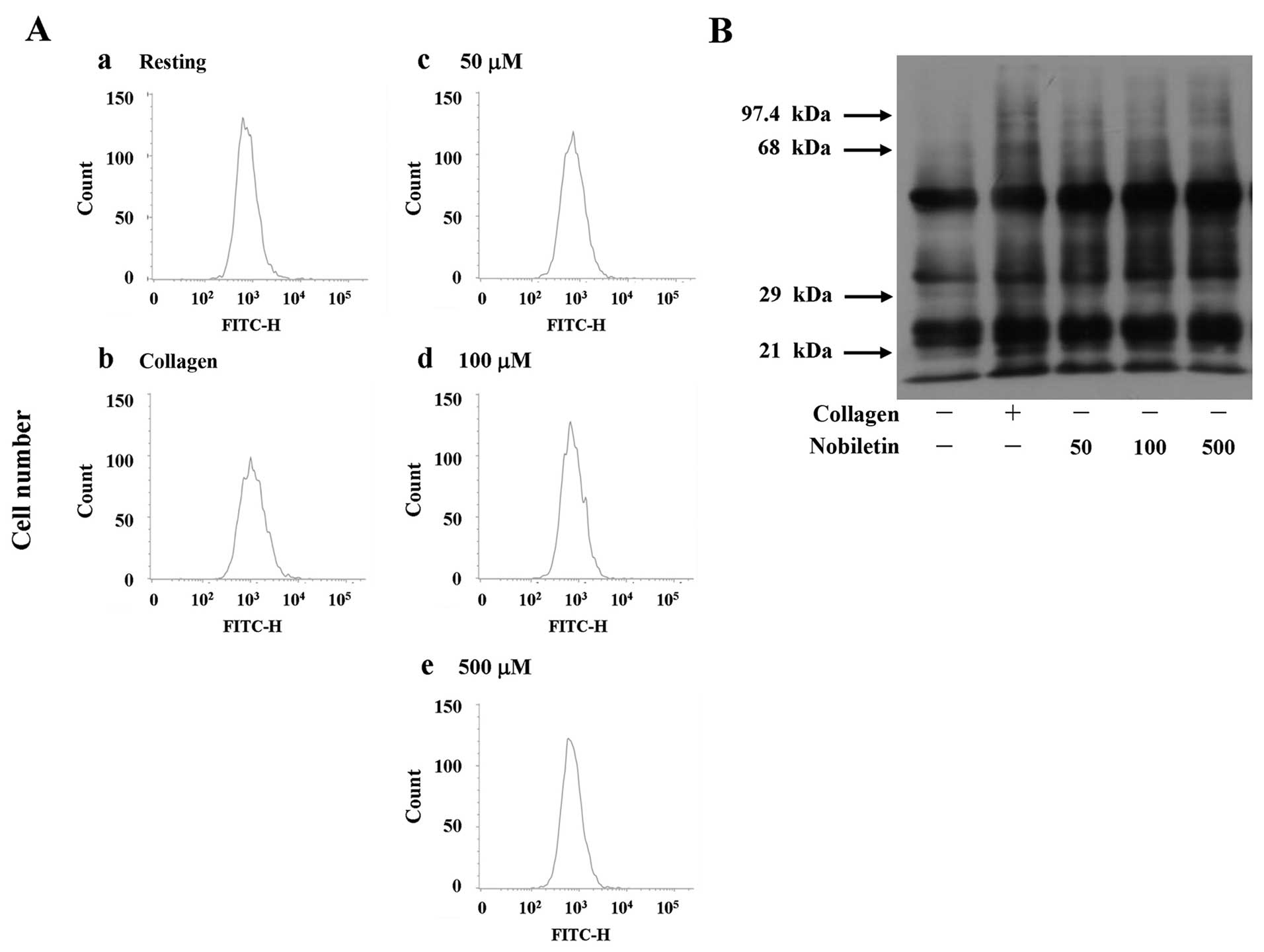
Effect of nobiletin on •OH and ROS
formation
The antioxidant properties of plants or their active
principles, mainly their radical scavenging activities, have been
intensively investigated over the years (29–31). In this study as shown in Fig. 2E, NAC, a cell-permeable
antioxidant, significantly reversed nobiletin-induced VASP
phosphorylation. Thus, we examined whether the process of
nobiletin-induced VASP phosphorylation is dependent on persuading
•OH radicals or ROS formation. ESR spectrometry and the
fluorescence-based method using DCF as a fluorescent probe revealed
that nobiletin, even at a higher concentration of 500 µM,
did not significantly stimulate •OH radical signals, as that
stimulated by either collagen (1 µg/ml; positive control)
(Fig. 3A) in washed human
platelets or by Fenton reaction (Fig.
3B). Furthermore, our results also demonstrated that nobiletin
(50 and 100 µM) did not reduce the fluorescence intensity of
DCF even at the highest concentration of 500 µM (Fig. 4A), which clearly indicated that
nobiletin does not have a stimulatory effect on •OH radicals as
well as on intracellular ROS.
Effects of nobiletin on protein
carbonyls
At present, much attention is focused on the protein
carbonyl content as the most widely used marker of the oxidative
modification of proteins in correlation with a variety of
pathologies (32). Since we noted
that nobiletin did not affect •OH radicals or ROS production, this
event was further confirmed by detecting carbonylated proteins in
nobiletin (50–500 µM)-treated platelets, which was compared
with collagen (positive control)-induced protein carbonylation.
When the platelets were activated by 1 µg/ml collagen,
protein carbonylation was markedly increased compared to the
resting platelets (Fig. 4B). This
augmentation is due to the oxidative stress generated by the cells
themselves when activated by the exogenous exposure of the
platelets to collagen. Furthermore, treatment with nobiletin at
various concentrations (50–500 µM) did not significantly
induce or reduce protein carbonylation, which was evidenced by
observing the unlettered intensity of the carbonylated bands, as
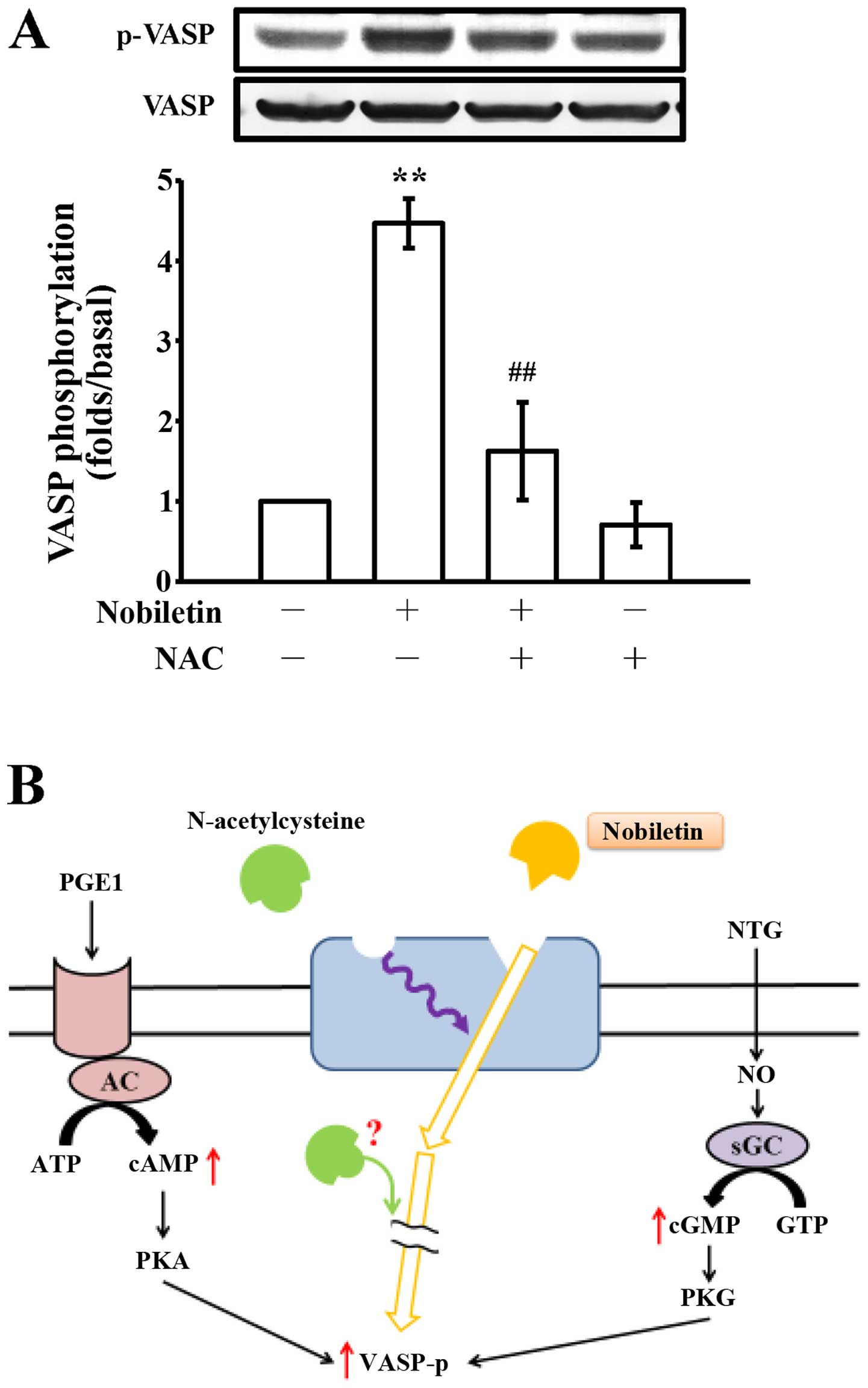
compared to the collagen-stimulated cells (Fig. 4B). Moreover, pre-treatment with
NAC (10 mM) markedly reversed the nobiletin-induced VASP
phosphorylation; however, treatment with NAC alone did not
significantly induce VASP phosphorylation (Fig. 5A).
Discussion
It has been considered that increased cyclic
AMP/cyclic GMP in platelets activates cyclic AMP-dependent PKA and
cyclic GMP-dependent PKG, and thus regulates the platelet
activation response by phosphorylating intracellular protein
substrates, such as VASP. Therefore, VASP phosphorylation may act
as a negative regulator of platelet activation. In this study, we
demonstrated that cyclic nucleotide-related pathways were not
responsible for nobiletin-induced VASP phosphorylation in washed
human platelets. Considering the fact that platelets are anuclear
cells, the projected hypothesis was thus confirmed by using various
well established cyclic nucleotide-related inhibitors, such as
adenylate cyclase inhibitor (SQ22536), guanylate cyclase inhibitor
(ODQ), PKA inhibitor (H89) and PKG inhibitor (KT5823) instead of
using their respective siRNAs. Our results indicated that
nobiletin-induced VASP phosphorylation was not affected by these
inhibitors. In addition, it was also demonstrated that this process
was independent of NF-κB, MAPKs, Akt, PLCγ2/PKC, NOX and ROS, since
the respective inhibitors of these molecules did not affect VASP
phosphorylation induced by nobiletin. Hence, it was important to
determine the actual mechanisms underlying the phosphorylation of
VASP by nobiletin.
VASP is a critical protein that plays a role in
regulating adhesive events that are involved in platelet
aggregation (15,33). Cyclic nucleotides activate cyclic
nucleotide-dependent kinases which inhibit all steps of this
cascade and simultaneously phosphorylate a number of proteins,
including VASP. Moreover, considering the results of a previous
study that suggested that VASP plays an important role in the
signaling of cyclic GMP- and cyclic AMP-dependent kinases (34), it was surprising that in this
study, nobiletin stimulated VASP phosphorylation in washed human
platelets without increasing the cyclic nucleotide levels. This
process was examined by observing whether the adenylate cyclase
inhibitor, SQ22536, and the guanylate cyclase inhibitor, ODQ,
modulate nobiletin-induced VASP phosphorylation in platelets. The
results confirmed that neither SQ22536 nor ODQ affected nobiletin
induced VASP phosphorylation as shown in Fig. 1B. A likely explanation for this
unexpected finding is that the absence of cyclic nucleotide
function in VASP phosphorylation may be substituted by other
members of proteins.
It is well-established that PKA is an essential
negative modulator of platelet function through the inhibition of
multiple biochemical events which all converge in the inhibition of
platelet function. It is also noteworthy that cyclic AMP may
cross-activate PKG in some vascular tissues. PKG has been
identified as the intracellular mediator of the effects of cyclic
GMP-elevating agents, such as nitric oxide or natriuretic peptides
(35). Therefore, we wished to
confirm whether nobiletin-induced VASP phosphorylation is dependent
on cyclic AMP/cyclic GMP and is linked to PKA/PKG activity. In
order to examine this hypothesis, the PKA inhibitor, H89, and the
PKG inhibitor, KT5823, were added to the nobiletin pre-treated
platelets. KT5823, an in vitro inhibitor of PKG, has been
extensively used to demonstrate or rule out an involvement of PKG
in signaling processes, and the PKA inhibitor, H89, is a chemical
compound which inhibits PKA in a competitive manner (36). In this study, we found that the
elevation of nobiletin-induced VASP phosphorylation in platelets
was not modulated by either H89 or KT5823, and thus, it appears
that VASP phosphorylation stimulated by nobiletin may be
independent of cyclic AMP/PKA and cyclic GMP/PKG pathways.
A previous study suggested that PKA and PKG are not
the only kinases able to phosphorylate VASP, but that PKC may also
have this ability (17). Chitaley
et al (17) reported that,
in cultured rat aortic smooth muscle cells, VASP becomes
phosphorylated in a PKG- and PKA-independent manner. They
established that VASP is also a substrate of PKC and provide
evidence for the involvement of a classical PKC isoform in VASP
phosphorylation (17). Moreover,
in our previous study, we reported that Akt acts as an upstream
regulator of PKC (37), and this
pathway was found to be responsible for phosphorylation of VASP in
activated platelets (38). Hence,
it is interesting to speculate whether PKC and Akt play a crucial
role in regulating nobiletin-induced phosphorylation of VASP in
platelets. Our results clearly demonstrated that this
phosphorylation was independent of the PKC and Akt pathways, since
their specific inhibitors, Ro318220 (PKC) and LY294004 (Akt), did
not alter the phosphorylation status of VASP, as its level was
similar in nobiletin alone-treated platelets (Fig. 2C).
MAPKs control cell proliferation, differentiation,
mitosis, survival and apoptosis. Among these MAPKs, p38 MAPK, JNK1
and ERK2 have been reported to exist in platelets and are activated
by various stimuli (39). ERK2
and JNK1 phosphorylation stimulated by collagen has been proven to
be involved in platelet aggregation (40). In addition, p38 MAPK activation
has also been shown in collagen-induced platelet activation and
secretion, which was restored by p38 MAPK inhibitors (41). However, our findings demonstrated
that the phosphorylation of VASP-stimulated by nobiletin was not
modulated by the p38 MAPK inhibitor SB203580, ERK inhibitor,
PD98059, and also by the JNK inhibitor, SP600125, indicating that
MAPK signaling pathways do not account for the stimulatory effects
of nobiletin on VASP phosphorylation. In addition, NF-κB induction
can be abrogated using its inhibitor, Bay11-7082, which is an
irreversible inhibitor of IκBα phosphorylation, resulting in the
downregulation of the cytokine-induced NF-κB activation (42). A previous study indicated that
Bay11-7082 potently inhibited collagen- and thrombin-induced
platelet aggregation, and it induced VASP phosphorylation through
cyclic AMP elevation and PKA activation (43). Following this, it was important to
determine whether Bay11-7082 can modulate nobiletin-induced
phosphorylation of VASP, and the data showed that this inhibitor
did not account for the increased phosphorylation of VASP in
response to nobiletin, indicating that NF-κB is also not involved
in this process.
ROS derived from platelet activation may augment
platelet reactivity during in vivo thrombus formation. Our
previous study suggested that free radical species act as secondary
messengers that increase cytosolic Ca2+ during the
initial phase of platelet activation processes, and PKC is involved
in receptor-mediated free radical production in platelets (44). That study also showed
H2O2 derived from platelets is converted into
•OH, and platelet aggregation can be inhibited by •OH scavengers
(44). A previous study found
that platelet-induced ROS production was inhibited by the radical
scavenger, NAC, and the NOX inhibitor, DPI (45). We found that NAC potently reversed
the stimulatory effects of nobiletin on VASP phosphorylation, but
DPI did not alter this phosphorylation, which indicated that NADPH
is also not involved in this process.
Protein carbonyl groups serve as a biomarker of
oxidative stress, since protein oxidation typically results in
increased carbonyl contents. It has been reported that
H2O2 produces a concentration-dependent
increase in the carbonylation of platelet proteins, and it can be
quenched by antioxidant catalase, suggesting that carbonylation is
induced by the oxidative stress generated by activated platelets
(32). Several ROS, including
•OH, oxidize amino acid residues in proteins to form products with
carbonyl groups that can be measured after reaction with DNPH. In
this study, the protein carbonyl concentrations in platelets
treated with collagen, as well as with nobiletin correlated with
changes in the intensity of DNPH reactive protein bands on
immunoblots. In addition, an active role of ROS in VASP
phosphorylation stimulated by nobiletin was monitored by DCF
fluorescence as a marker for the intracellular generation of
radicals. From these experiments, it was found that
nobiletin-induced VASP phosphorylation is independent of ROS
formation. Moreover, it has been demonstrated that NAC decreases
human platelet aggregation and increases the intracellular levels
of cyclic GMP by interplaying with endogenous NO (46), which is constitutively synthesized
by platelets (47) and inhibits
aggregation by activating the soluble guanylate cyclase (48). In this study, as shown in Fig. 2E, NAC alone did not trigger VASP
phosphorylation as nobiletin did. The possible explanations for
this finding may be due to the following reasons: first, NAC may
bind to allosteric sites on the receptor distinct from that
nobiletin binds in the human platelets to block the signals of
nobiletin-triggered VASP phosphorylation; second, since NAC is a
cell-permeable antioxidant, it may interrupt the intracellular
signaling cascade which is responsible for VASP phosphorylation
through an unidentified mechanism (Fig. 5B).
In conclusion, our previous study suggested that
nobiletin represents a potential therapeutic agent for the
prevention or treatment of thromboembolic disorders. In the present
study, and to the best of our knowledge, we found for the first
time that nobiletin stimulates VASP phosphorylation via a novel
non-cyclic nucleotide-dependent mechanisms in washed human
platelets. In addition, an unexpected result was observed in that
NAC reversed the nobiletin-induced VASP phosphorylation. However,
the exact inhibitory mechanisms of action of NAC warrant further
investigation.
Acknowledgments
This study was supported by grants
(MOST103-2320-B-038-017, MOST104-2622-B-038-003 and
MOST104-2320-B-038-045-MY2) from the Ministry of Science and
Technology of Taiwan and Chi Mei Medical Center, Taipei Medical
University (104CM-TMU-04).
References
|
1
|
Manthey JA and Grohmann K: Phenols in
citrus peel byproducts. Concentrations of hydroxycinnamates and
polymethoxylated flavones in citrus peel molasses. J Agric Food
Chem. 49:3268–3273. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nakajima A, Aoyama Y, Shin EJ, Nam Y, Kim
HC, Nagai T, Yokosuka A, Mimaki Y, Yokoi T, Ohizumi Y and Yamada K:
Nobiletin, a citrus flavonoid, improves cognitive impairment and
educes soluble Aβ levels in a triple transgenic mouse model of
Alzheimer's disease (3XTg-AD). Behav Brain Res. 289:69–77. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kawahata I, Yoshida M, Sun W, Nakajima A,
Lai Y, Osaka N, Matsuzaki K, Yokosuka A, Mimaki Y, Naganuma A, et
al: Potent activity of nobiletin-rich Citrus reticulata peel
extract to facilitate cAMP/PKA/ERK/CREB signaling associated with
learning and memory in cultured hippocampal neurons: identification
of the substances responsible for the pharmacological action. J
Neural Transm (Vienna). 120:1397–1409. 2013. View Article : Google Scholar
|
|
4
|
Kawaii S, Tomono Y, Katase E, Ogawa K and
Yano M: Quantitation of flavonoid constituents in citrus fruits. J
Agric Food Chem. 47:3565–3571. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiong Y, Chen D, Yu C, Lv B, Peng J, Wang
J and Lin Y: Citrus nobiletin ameliorates experimental colitis by
reducing inflammation and restoring impaired intestinal barrier
function. Mol Nutr Food Res. 59:829–842. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu YQ, Zhou CH, Tao J and Li SN:
Antagonistic effects of nobiletin, a polymethoxyflavonoid, on
eosinophilic airway inflammation of asthmatic rats and relevant
mechanisms. Life Sci. 178:2689–2696. 2006. View Article : Google Scholar
|
|
7
|
Lu YH, Su MY, Huang HY, Lin-Li and Yuan
CG: Protective effects of the citrus flavanones to PC12 cells
against cytotoxicity induced by hydrogen peroxide. Neurosci Lett.
484:6–11. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi BK, Kim TW, Lee DR, Jung WH, Lim JH,
Jung JY, Yang SH and Suh JW: A polymethoxy flavonoids-rich Citrus
aurantium extract ameliorates ethanol-induced liver injury through
modulation of AMPK and Nrf2-related signals in a binge drinking
mouse model. Phytother Res. 29:1577–1584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cui Y, Wu J, Jung SC, Park DB, Maeng YH,
Hong JY, Kim SJ, Lee SR, Kim SJ, Kim SJ, et al:
Anti-neuroinflammatory activity of nobiletin on suppression of
microglial activation. Biol Pharm Bull. 33:1814–1821. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ho SC and Kuo CT: Hesperidin, nobiletin,
and tangeretin are collectively responsible for the
anti-neuroinflammatory capacity of tangerine peel (Citri
reticulatae pericarpium). Food Chem Toxicol. 71:176–182. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Zhao H, Zhang X, Chen L, Zhao X,
Bai X and Zhang J: Nobiletin protects against cerebral ischemia via
activating the p-Akt, p-CREB, BDNF and Bcl-2 pathway and
ameliorating BBB permeability in rat. Brain Res Bull. 96:45–53.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Benz PM, Blume C, Seifert S, Wilhelm S,
Waschke J, Schuh K, Gertler F, Münzel T and Renné T: Differential
VASP phosphorylation controls remodeling of the actin cytoskeleton.
J Cell Sci. 122:3954–3965. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Major TC, Handa H, Brisbois EJ, Reynolds
MM, Annich GM, Meyerhoff ME and Bartlett RH: The mediation of
platelet quiescence by NO-releasing polymers via cGMP-induced
serine 239 phosphorylation of vasodilator-stimulated
phosphoprotein. Biomaterials. 34:8086–8096. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee DH, Cho HJ, Kim HH, Rhee MH, Ryu JH
and Park HJ: Inhibitory effects of total saponin from Korean red
ginseng via vasodilator-stimulated phosphoprotein-Ser(157)
phosphorylation on thrombin-induced platelet aggregation. J Ginseng
Res. 37:176–186. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aszódi A, Pfeifer A, Ahmad M, Glauner M,
Zhou XH, Ny L, Andersson KE, Kehrel B, Offermanns S and Fässler R:
The vasodilator-stimulated phosphoprotein (VASP) is involved in
cGMP- and cAMP-mediated inhibition of agonist-induced platelet
aggregation, but is dispensable for smooth muscle function. EMBO J.
18:37–48. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eckly-Michel A, Martin V and Lugnier C:
Involvement of cyclic nucleotide-dependent protein kinases in
cyclic AMP-mediated vasorelaxation. Br J Pharmacol. 122:158–164.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chitaley K, Chen L, Galler A, Walter U,
Daum G and Clowes AW: Vasodilator-stimulated phosphoprotein is a
substrate for protein kinase C. FEBS Lett. 556:211–215. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Horstrup K, Jablonka B, Hönig-Liedl P,
Just M, Kochsiek K and Walter U: Phosphorylation of focal adhesion
vasodilator-stimulated phosphoprotein at Ser157 in intact human
platelets correlates with fibrinogen receptor inhibition. Eur J
Biochem. 225:21–27. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hauser W, Knobeloch KP, Eigenthaler M,
Gambaryan S, Krenn V, Geiger J, Glazova M, Rohde E, Horak I, Walter
U and Zimmer M: Megakaryocyte hyperplasia and enhanced
agonist-induced platelet activation in vasodilator-stimulated
phosphoprotein knockout mice. Proc Natl Acad Sci USA. 96:8120–8125.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu WJ, Lin KC, Liu CP, Lin CY, Wu HC, Chou
DS, Geraldine P, Huang SY, Hsieh CY and Sheu JR: Prevention of
arterial thro mbosis by nobiletin: in vitro and in vivo studies. J
Nutr Biochem. 28:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vaiyapuri S, Roweth H, Ali MS, Unsworth
AJ, Stainer AR, Flora GD, Crescente M, Jones CI, Moraes LA and
Gibbins JM: Pharmacological actions of nobiletin in the modulation
of platelet function. Br J Pharmacol. 172:4133–4145. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sheu JR, Lee CR, Lin CH, Hsiao G, Ko WC,
Chen YC and Yen MH: Mechanisms involved in the antiplatelet
activity of Staphylococcus aureus lipoteichoic acid in human
platelets. Thromb Haemost. 83:777–784. 2000.PubMed/NCBI
|
|
23
|
Chou DS, Hsiao G, Shen MY, Tsai YJ, Chen
TF and Sheu JR: ESR spin trapping of a carbon-centered free radical
from agonist-stimulated human platelets. Free Radic Biol Med.
39:237–248. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Smolenski A, Bachmann C, Reinhard K,
Hönig-Liedl P, Jarchau T, Hoschuetzky H and Walter U: Analysis and
regulation of vasodilator-stimulated phosphoprotein serine 239
phosphorylation in vitro and in intact cells using a
phospho-specific monoclonal antibody. J Biol Chem. 273:20029–20035.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pula G, Schuh K, Nakayama K, Nakayama KI,
Walter U and Poole AW: PKCdelta regulates collagen-induced platelet
aggregation through inhibition of VASP-mediated filopodia
formation. Blood. 108:4035–4044. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Butt E, Abel K, Krieger M, Palm D, Hoppe
V, Hoppe J and Walter U: cAMP- and cGMP-dependent protein kinase
phosphorylation sites of the focal adhesion vasodilator-stimulated
phosphoprotein (VASP) in vitro and in intact human platelets. J
Biol Chem. 269:14509–14517. 1994.PubMed/NCBI
|
|
27
|
Liu F, Morris S, Epps J and Carroll R:
Demonstration of an activation regulated NF-kappaB/I-kappaBalpha
complex in human platelets. Thromb Res. 106:199–203. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Seno T, Inoue N, Gao D, Okuda M, Sumi Y,
Matsui K, Yamada S, Hirata KI, Kawashima S, Tawa R, et al:
Involvement of NADH/NADPH oxidase in human platelet ROS production.
Thromb Res. 103:399–409. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gülçin I, Elmastaş M and Aboul-Enein HY:
Determination of antioxidant and radical scavenging activity of
Basil (Ocimum basilicum L. Family Lamiaceae) assayed by different
methodologies. Phytother Res. 21:354–361. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kuczmannová A, Gál P, Varinská L, Treml J,
Kováč I, Novotný M, Vasilenko T, Dall'Acqua S, agy M and Mučaji P:
Agrimonia eupatoria L. and Cynara cardunculus L. Water Infusions:
Phenolic profile and comparison of antioxidant activities.
Molecules. 20:20538–20550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Timmers MA, Guerrero-Medina JL, Esposito
D, Grace MH, Paredes-López O, García-Saucedo PA and Lila MA:
Characterization of phenolic compounds and antioxidant and
anti-inflammatory activities from mamuyo (Styrax ramirezii Greenm.)
fruit. J Agric Food Chem. 63:10459–10465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alexandru N, Constantin A and Popov D:
Carbonylation of platelet proteins occurs as consequence of
oxidative stress and thrombin activation, and is stimulated by
ageing and type 2 diabetes. Clin Chem Lab Med. 46:528–536. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Massberg S, Grüner S, Konrad I, Garcia
Arguinzonis MI, Eigenthaler M, Hemler K, Kersting J, Schulz C,
Muller I, Besta F, et al: Enhanced in vivo platelet adhesion in
vasodilator-stimulated phosphoprotein (VASP)-deficient mice. Blood.
103:136–142. 2004. View Article : Google Scholar
|
|
34
|
Halbrügge M, Friedrich C, Eigenthaler M,
Schanzenbächer P and Walter U: Stoichiometric and reversible
phosphorylation of a 46-kDa protein in human platelets in response
to cGMP- and cAMP-elevating vasodilators. J Biol Chem.
265:3088–3093. 1990.PubMed/NCBI
|
|
35
|
Lucas KA, Pitari GM, Kazerounian S,
Ruiz-Stewart I, Park J, Schulz S, Chepenik KP and Waldman SA:
Guanylyl cyclases and signaling by cyclic GMP. Pharmacol Rev.
52:375–414. 2000.PubMed/NCBI
|
|
36
|
Butt E, van Bemmelen M, Fischer L, Walter
U and Jastorff B: Inhibition of cGMP-dependent protein kinase by
(Rp)-guanosine 3′,5′-monophosphorothioates. FEBS Lett. 263:47–50.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu WJ, Lee JJ, Chou DS, Jayakumar T, Fong
TH, Hsiao G and Sheu JR: A novel role of andrographolide, an
NF-kappa B inhibitor, on inhibition of platelet activation: the
pivotal mechanisms of endothelial nitric oxide synthase/cyclic GMP.
J Mol Med (Berl). 89:1261–1273. 2011. View Article : Google Scholar
|
|
38
|
Dorsam RT, Kim S, Jin J and Kunapuli SP:
Coordinated signaling through both G12/13 and G(i) pathways is
sufficient to activate GPIIb/IIIa in human platelets. J Biol Chem.
277:47588–47595. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Adam F, Kauskot A, Rosa JP and Bryckaert
M: Mitogen-activated protein kinases in hemostasis and thrombosis.
J Thromb Haemost. 6:2007–2016. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kauskot A, Adam F, Mazharian A, Ajzenberg
N, Berrou E, Bonnefoy A, Rosa JP, Hoylaerts MF and Bryckaert M:
Involvement of the mitogen-activated protein kinase c-Jun
NH2-terminal kinase 1 in thrombus formation. J Biol
Chem. 282:31990–31999. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kuliopulos A, Mohanlal R and Covic L:
Effect of selective inhibition of the p38 MAP kinase pathway on
platelet aggregation. Thromb Haemost. 92:1387–1393. 2004.PubMed/NCBI
|
|
42
|
Pierce JW, Schoenleber R, Jesmok G, Best
J, Moore SA, Collins T and Gerritsen ME: Novel inhibitors of
cytokine-induced IkappaBalpha phosphorylation and endothelial cell
adhesion molecule expression show anti-inflammatory effects in
vivo. J Biol Chem. 272:21096–21103. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee HS, Kim SD, Lee WM, Endale M,
Kamruzzaman SM, Oh WJ, Cho JY, Kim SK, Cho HJ, Park HJ, et al: A
noble function of BAY 11-7082: inhibition of platelet aggregation
mediated by an elevated cAMP-induced VASP, and decreased ERK2/JNK1
phosphorylations. Eur J Pharmacol. 627:85–91. 2010. View Article : Google Scholar
|
|
44
|
Sheu JR, Hung WC, Wu CH, Ma MC, Kan YC,
Lin CH, Lin MS, Luk HN and Yen MH: Reduction in
lipopolysaccharide-induced thrombocytopenia by triflavin in a rat
model of septicemia. Circulation. 99:3056–3062. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Berg C, Trofast C and Bengtsson T:
Platelets induce reactive oxygen species-dependent growth of human
skin fibroblasts. Eur J Cell Biol. 82:565–571. 2003. View Article : Google Scholar
|
|
46
|
Anfossi G, Russo I, Massucco P, Mattiello
L, Cavalot F and Trovati M: N-acetyl-L-cysteine exerts direct
anti-aggregating effect on human platelets. Eur J Clin Invest.
31:452–461. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Radomski MW, Palmer RMJ and Moncada S: An
L-arginine/nitric oxide pathway present in human platelets
regulates aggregation. Proc Natl Acad Sci USA. 87:5193–5197. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Moro MA, Russel RJ, Cellek S, Lizasoain I,
Su Y, Darley-Usmar VM, Radomski MW and Moncada S: cGMP mediates the
vascular and platelet actions of nitric oxide: confirmation using
an inhibitor of the soluble guanylyl cyclase. Proc Natl Acad Sci
USA. 93:1480–1485. 1996. View Article : Google Scholar : PubMed/NCBI
|