Introduction
Liver ischemia/reperfusion (I/R) injury is an
important cause of liver damage occurring during liver surgery or
transplantation, significantly influences the prognosis of liver
function (1). A variety of immune
cells, including monocytes, Kupffer cells, CD4+
lymphocytes, neutrophils, hepatocytes, and cytokines are involved
in liver I/R injury (2–4). Hepatic macrophages have an important
role in maintaining homeostasis of the liver and in the
pathogenesis of liver injury. I/R can cause the activation of
Kupffer cells to release transforming growth factor-α, and other
pro-inflammatory cytokines, including tumor necrosis factor-α
(TNF-α) and interleukin-6 (IL-6). An increase in the inflammatory
state results in the activation of the immune system as a major
intermediary during the process of I/R injury (5), which can drive injury resolution or
lead to chronic inflammation (6,7).
Intrahepatic monocyte infiltration of the damaged
liver tissue represents an important constitution of the innate
immune response (8). When the
liver is under pathological stress, such as steatohepatitis and
viral hepatitis, Kupffer cells can differentiate from infiltrated
bone marrow-derived mononuclear cells (9). In addition, it has been reported
that Kupffer cells can be constantly replenished by blood
monocytes, even in steady state conditions (10). Two major monocyte subsets in mice
have been identified that closely resemble human monocytes and vary
in migratory and differentiation properties (11). In humans, classical
CD14+CD16+ monocytes express C-C chemokine
receptor 2 (CCR2), CD64, and selectin L, whereas non-classical
CD14+CD16− monocytes lack CCR2 (12). Their counterparts in mice are
CCR2+Ly6C high (Ly6Chi) and
CCR2−Ly6C low (Ly6Clo) monocytes,
respectively (12,13). Ly6Chi monocytes are
considered to be precursors to macrophages and dendritic cells
during inflammatory conditions, whereas Ly6Clo monocytes
represent steady state precursor cells for tissue macrophages
(14-16). The differential recruitment of
these monocyte subsets appears to be crucially controlled by
chemokines released from injured tissue (17-19). As such, enhanced hepatic
expression of the ligands for CCR2, including monocyte
chemoattractant protein-1 (MCP-1)/C-C motif chemokine ligand 2
(CCL2), CCR6 and macrophage inflammatory protein-3α/CCL20, has been
reported in patients with liver cirrhosis (17,20).
CCR2 is the specific receptor for MCP-1/CCL2, but is
also the receptor for CCL7, CCL8, CCL11, CCL12 and CCL13. Recently,
CCL2 has been reported to have a critical role in the pathogenesis
of various liver diseases (21-23). CCL2 interacts with its receptor,
CCR2, and induces mononuclear cells in the blood to cross the
endothelial barrier towards sites of inflammation during an
inflammatory response (24).
Upregulated expression of CCL2 has been reported in various liver
diseases (21,25). Although the CCR2/CCL2 interaction
has been reported to be associated with various liver diseases
(26), the underlying mechanism
of this association has not been clearly elucidated. Specifically,
the role of CCL2/CCR2 in the immune response of the liver during
I/R injury remains unknown. The current study aimed to identify the
infiltrating monocyte subsets in liver I/R injury and determine how
the CCL2/CCR2 axis contributes to the hepatic monocyte recruitment
during I/R injury.
Materials and methods
Animals
Male C57BL/6 mice [wild-type (WT); weight, 22–25 g]
were purchased from the Center for Animal Experiment of Wuhan
University (Wuhan, China). Male CCL2−/− or
CCR2−/− C57BL/6 mice were bred at the sterile Animal
Management Centre of Tongji Medical College (Wuhan, China) and used
between the ages of 8–12 weeks. The original experiments used 10
mice/group [including hematoxylin and eosin (H&E) staining,
serum levels of alanine aminotransferase (ALT) and aspartate
aminotransferase (AST), mRNA/protein levels of TNF-α and IL-6 in
liver tissue; recruitment of monocytes into I/R injured liver
lobes; effect of macrophage depletion as well as CCR2 or CCL2
knockout on liver I/R injury]. For the additional experiments
performed (quantification of intrahepatic CD4+ T cells
and CD8+ T cells; immunofluorescence staining of hepatic
infiltrated monocyte subsets; effect of CCR2 inhibition on hepatic
I/R injury), 4-6 animals were used per group. All animal studies
were performed in accordance with the National Institutes of Health
Guidelines for Care and Use of Laboratory Animals and were approved
by the Scientific Affairs Committee on Animal Research and Ethics
of Tongji Hospital, Tongji Medical College, Huazhong University of
Science and Technology (Wuhan, China).
Liver I/R model
A non-lethal model of segmental (70%) hepatic warm
ischemia was performed as described previously (27). Briefly, mice were anesthetized
with sodium pentobarbital [60 mg/kg; intraperitoneally (i.p.)],
which is a traditional anesthetic used in animal research of liver,
lung or cardiac I/R injury (28–30). A midline laparotomy was performed
and ligaments surrounding each lobe were dissected carefully. The
portal vein, the hepatic artery, and the bile duct supplying the
median and the left lateral lobes of the liver were clamped with an
artery clip. The caudal and right lobes retained portal and
arterial inflow and venous outflow, preventing intestinal venous
congestion. Following surgery, the abdominal cavity was properly
closed and then the mice were placed in an incubator maintained at
32°C. Liver lobes were inspected for ischemia by visualizing the
pale blanching of the ischemic lobes intermittently. After 60 min,
the clamps were removed and liver reperfusion was initiated. For
sham controls, the animals underwent anesthesia, laparotomy, and
exposure of the portal triad without hepatic I/R. Mice were
sacrificed after 6 h of reperfusion. Blood and tissues of liver
lobes from sham and I/R injury mice were collected and stored at
−80°C for future analysis.
Serum transaminase
Cardiac puncture blood was collected following
hepatic I/R or sham into tubes (without anticoagulant), allowed to
clot and serum was separated by centrifugation at 200 x g at 4°C
for 10 min. For CCR2 inhibitor pretreatment, mice were injected
with RS-504393 (25 mg/kg; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) or dimethyl sulfoxide (DMSO) 1 h prior to I/R injury.
Liver injury was estimated by measuring the increased activities of
serum ALT and AST, which were measured using a Synchron CX7
analyzer (Beckman Coulter, Inc., Brea, CA, USA) in the Clinical
Biochemical Laboratory of Wuhan Union Hospital (Wuhan, China).
Histopathology
At predetermined time points (6 h after
reperfusion), mice were sacrificed humanely by CO2
suffocation. The liver lobes were collected from I/R mice and sham
mice and fixed in 4% formalin at room temperature (25°C) for 4 h.
Fixed tissues were embedded in paraffin, cut into 5 µm thick
sections and were placed onto glass slides. Slides were then
stained with hematoxylin and eosin (H&E) using a conventional
protocol. The necrotic area was assessed by 2 independent
pathologists, and more than 10 random fields of view were
quantified using the Image-Pro Plus v 6.0 software (Media
Cybernetics, Silver Spring, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the lobes of the
ischemic and sham liver using TRIzol. RT was performed using the RT
kit (Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 65°C for
5 min. qPCR was performed using the SYBR-Green PCR mix (Thermo
Fisher Scientific, Inc.) in a LightCycler 480 system (Roche
Molecular Diagnostics, Pleasanton, CA, USA). The PCR program was
95°C, 2 min, ×1; 95°C, 10 sec, 60°C, 1 min, ×40. β-actin was used
as an internal reference. Many previous I/R studies (31,32) also used this gene for data
normalization, indicating I/R has no effect on the expression of
β-actin. Primers were synthesized by Sangon Biotech Co., Ltd.
(Shanghai, China) and the sequences were as follows: TNF-α, 5′-ACT
GAA CTT CGG GGT GATCG-3′ (forward) and 5′-GGC TAC AGG CTT GTC
ACTCG-3′ (reverse); IL-6, 5′-AGT TGC CTT CTT GGG ACTGA-3′ (forward)
and 5′-TCC ACG ATT TCC CAG AGAAC-3′ (reverse); β-actin, 5′-GTG ACG
TTG ACA TCC GTA AAGA-3′ (forward) and 5′-GCC GGA CTC ATC GTA
CTCC-3′ (reverse). Data were analyzed using the 2−ΔΔCq
method (33).
Serum/liver cytokines
The serum expression levels of TNF-α and IL-6 was
measured using commercial ELISA kits [TNF-α (MTA00B); IL-6
(M6000B); R&D Systems, Inc., Minneapolis, MN, USA] following
standardized techniques (34).
All samples and standards were performed in duplicate. ELISA was
also performed on tissues of liver lobes from sham and I/R
mice.
Isolation of hepatic non-parenchymal
cells (NPCs)
Hepatic NPCs were obtained from the liver using a
collagenase digestion method, as described previously (35). Briefly, the animals were
anesthetized and the liver was perfused in situ by way of
the portal vein with PBS containing 0.05% collagenase. The liver
was removed, placed in PBS and incubated at 37°C for 15 min. The
incubated liver was then torn using cell scrapers. The cell
suspension were shaken on a shaking table at 37°C for 20 min and
then filtered through a 70-µm nylon mesh. NPCs were isolated
by gradient centrifugation at 400 x g for 16 min with acceleration
and braking at 0 using OptiPrep™ (Axis-Shield Diagnostics Ltd.,
Dundee, UK) according to manufacturer's instructions. NPCs were
then washed by high-speed centrifugation (1,500 rpm for 5 min).
Depletion of Kupffer cells
Kupffer cells were depleted by using clodronate in a
liposomal formulation (36,37). Briefly, a dose of 0.2 ml/20 g
animal body weight of liposome-encapsulated clodronate (FormuMax
Scientific, Inc., Sunnyvale, CA, USA) was i.p. injected 48 h before
I/R. Macrophage depletion was successfully achieved (~90%), as
examined by flow cytometry (anti-F4/80-PerCP-Cy5.5, clone BM8 (cat.
no. 123128); from BioLegend, Inc., San Diego, CA, USA) in the liver
48 h after a single injection.
Flow cytometric analysis
Red blood cells in the isolated NPCs were lysed
using Hybri-Max red blood cell lysis buffer (Sigma-Aldrich; Merck
KGaA). Cell numbers were determined by a sequential gating scheme.
The cells were then incubated with fluorescent-labeled anti-mouse
antibodies in PBS containing 1% bovine serum albumin for 30 min at
4°C. Antibodies used in this experiment were as follows:
anti-CD45-Pacific Blue (clone 30-F11; cat. no. 103126),
anti-Ly6C−APC-Cy7 (clone HK1.4; cat. no. 128026),
anti-CD11b-PE-Cy7 (clone M1/70; cat. no. 101216), and
anti-CD4-PerCP-Cy5.5 (clone GK1.5; cat. no. 100434) from BioLegend;
anti-CD8-APC (clone 53-6.7; cat. no. 561093) from BD Biosciences
(San Diego, CA, USA); anti-CD3-PE-Cy7 (clone 17A2; cat. no. 100220)
from BioLegend, Inc. For intracellular cytokine staining, NPCs were
isolated following sham or I/R injury of the 3 groups of mice
(n=4–6 mice/group) and cultured with GolgiStop (cat. no. 554724; BD
Biosciences) for 6 h. The cells were then stained with CD11b, Ly6C
and anti-TNF-α-PE (IC410P) from R&D Systems. The corresponding
isotype IgGs were used when necessary as controls. Following
staining, cells were washed, fixed in 1% para-formaldehyde in 1X
PBS, and resuspended at ~5×106/ml for flow cytometry.
Cells were analyzed on a CyAn ADP analyzer (Beckman Coulter, Inc.).
FlowJo version 7.6 software (Tree Star, Inc., Ashland, OR, USA) was
used to analyze the data (38).
Data analysis
The results were expressed as the mean ± standard
error. Student's t-test was used to compare the difference between
two groups. For multigroup comparison, one-way analysis of variance
was used followed by post hoc Mann Whitney U-test. All of the data
analysis was performed using SPSS 15.0 software (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Hepatic pathology in I/R injured
liver
Compared with sham animals with normal liver
histology, mice undergoing I/R injury exhibited periportal necrosis
after 6 h, as determined by H&E staining (Fig. 1A). I/R-induced liver injury was
also indicated by elevated serum levels of ALT and AST (Fig. 1B). The levels of inflammatory
cytokines TNF-α and IL-6 were significantly higher in liver tissues
of I/R mice than in sham animals at the mRNA (Fig. 1C) and protein (Fig. 1D) levels. For the positive
control, mice were treated with CCl4 (0.3 ml/kg) and elevated TNF-α
and IL-6 in liver tissue were observed (data not shown).
Ly6 high (Ly6Chi) and low
(Ly6Clo)monocytes are increased in I/R-injured
livers
To investigate the cells that secrete the cytokines
demonstrated, the population of intrahepatic leukocytes present in
the damaged liver was characterized. The flow cytometric plots of
CD11b and Ly6C expression in isolated hepatic NPCs from one sham
mouse and one I/R mouse, representative of the 10 mice in each
group, are presented in Fig. 2.
The population of CD11b+Ly6Chi cells
(inflammatory monocytes) increased following I/R injury, from 2.1
to 6.9% of CD45+ immune cells.
CD11b+Ly6Clo cells (reparative monocytes)
also increased from 9.21 to 20.6% of CD45+ immune cells
following hepatic I/R injury (Fig.
2). Statistical analysis was then calculated based on data from
all animals of each group and there was significant difference in
CD11b+Ly6Chi/low populations between the sham
group and I/R group. These results suggest that the increased
infiltration of monocytes into liver may have an important role in
hepatic pathology in I/R-injured liver.
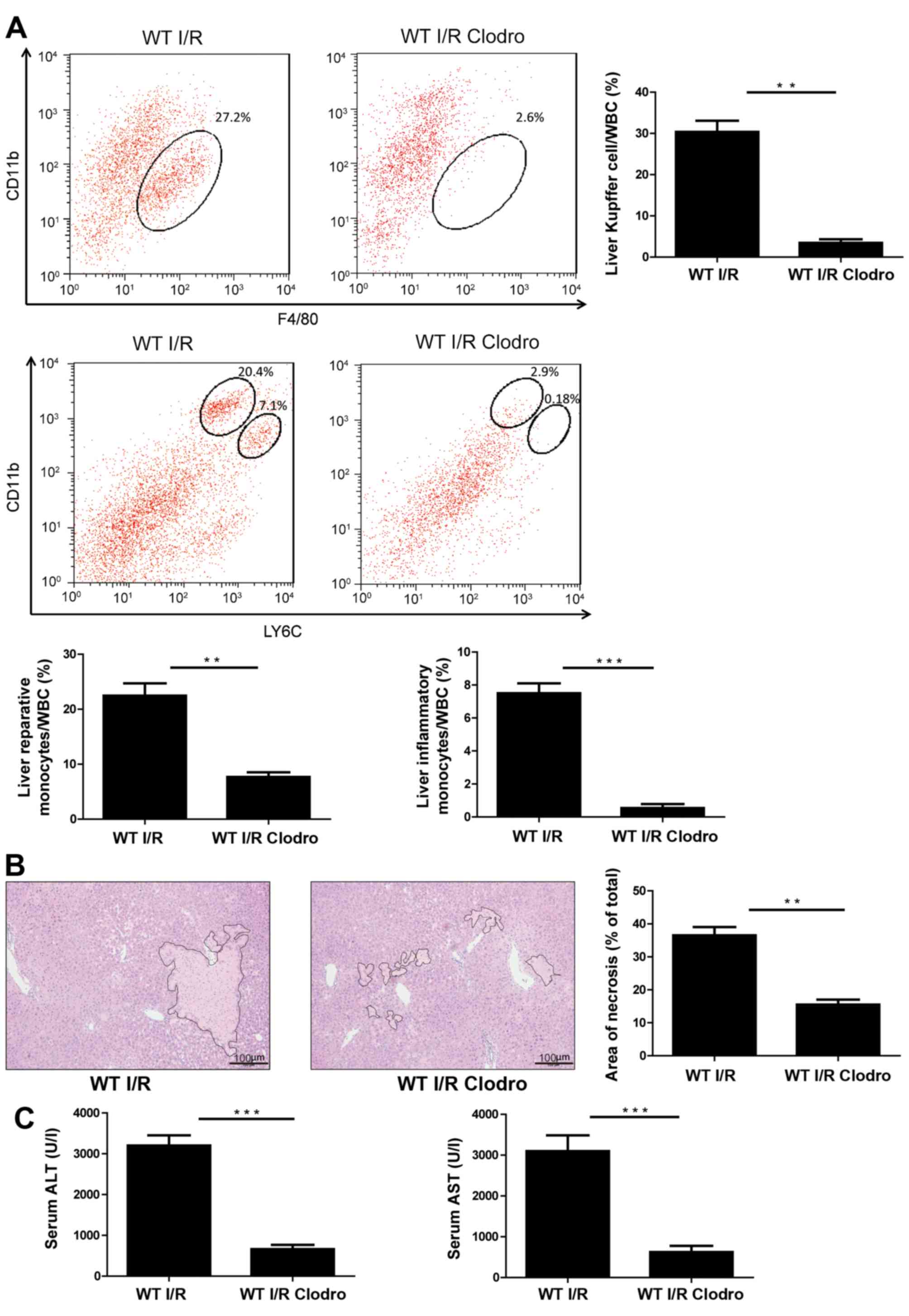
Treatment with clodronate-loaded
liposomes alleviates liver I/R injury
To determine whether monocyte-derived subsets
actively promote liver damage or represent an elemental reaction
following I/R injury, WT mice were treated with clodronate-loaded
liposomes to transiently deplete systemic macrophages 48 h before
I/R injury (39). As presented in
Fig. 3A, the population of
CD11b+Ly6Chi cells in I/R model mice
decreased from 7.1 to 0.18% following pretreatment with
clodronate-loaded liposomes. CD11b+Ly6Clo
cells also decreased from 20.4 to 2.9% in macrophage-depleted mice
(Fig. 3A). Following macrophage
depletion by clodronate-loaded liposomes, I/R-induced hepatic
necrosis was also dramatically decreased (Fig. 3B). Serum ALT and AST levels were
also reduced in mice pretreated with clodronate-loaded liposome
compared to animals pretreated with PBS (Fig. 3C). These results suggest that the
recruitment of monocyte subsets into the injured liver may
contribute to acute hepatic damage.
CCR2/CCL2 axis is critical for the
recruitment of Ly6C+ intrahepatic monocyte-derived
cells
Ly6C+ monocytes express high levels of
CCR2, CCR1 and CCR6 (39,40). To elucidate the role of CCR2-CCL2
axis in the recruitment of monocytes to damaged liver tissue,
I/R-induced liver injury and intrahepatic recruitment of
Ly6C+ monocytes were examined in WT, CCR2−/−
and CCL2−/− mice. Compared with WT mice that exhibited
significant hepatic necrosis, CCR2−/− and
CCL2−/− mice exhibited lower levels of necrosis
following I/R injury (Fig. 4A).
Serum ALT and AST levels were also decreased compared with the WT
group (Fig. 4B). The levels of
inflammatory cytokines, TNF-α and IL-6, were also decreased in the
CCR2−/− and CCL2−/− mice (Fig. 4C and D). In addition, leukocyte
infiltration was significantly reduced following injury in CCR2 or
CCL2 deficient mice (Fig. 5A).
There was a significantly lower accumulation of intrahepatic
CD11b+Ly6C+ monocytes in CCR2−/−
and CCL2−/− mice undergoing I/R. The percentage of
CD11b+Ly6Chi inflammatory monocytes and
CD11b+Ly6Clo reparative monocytes were
decreased in CCR2−/− and CCL2−/− mice
compared with WT mice (Fig. 5A).
However, there was no difference in the number of
CD11b+Ly6Chi cells between CCR2−/−
and CCL2−/− sham group and the operation group (Fig. 5). Flow cytometric quantification
also demonstrated that CCR2 or CCL2 knockout decreased the
accumulation of CD4+ T cells and CD8+ T cells
in mice that underwent hepatic I/R injury (Fig. 5B). The decrease in
CD11b+Ly6C+ cells was also revealed by
immunofluorescence staining of infiltrated monocyte subsets in
injured liver tissue (Fig. 6A).
Intracellular cytokine staining demonstrated that
CCR2−/− or CCL2−/− mice had significantly
lower levels of TNF-α than WT mice following I/R injury, whereas
there was no induction of TNF-α in the three groups under sham
conditions (Fig. 6B).
 | Figure 4Knockout of CCR2 or CCL2 alleviates
liver I/R injury. (A) Three groups of C57BL/6 mice WT,
CCR2−/− and CCL2−/−(n=10/group] were
subjected to non-lethal segmental warm hepatic I/R injury. The
liver lobes were hematoxylin and eosin stained and areas of
necrosis were quantified. (B) Serum levels of ALT and AST in
CCR2−/−, CCL2−/− or WT I/R injured mice. (C)
mRNA levels of TNF-α and IL-6 in CCR2−/−,
CCL2−/− or WT I/R injured mice. (D) Protein levels of
TNF-α and IL-6 in the liver tissues of CCR2−/−,
CCL2−/− or WT I/R injured mice. The results are
presented as the mean + standard error of each group. *P<0.05
and **P<0.01. WT, wild-type; I/R,
ischemia/reperfusion; CCR2, C-C chemokine receptor 2; KO, knockout;
CCL2, C-C motif chemokine ligand 2; ALT, alanine aminotransferase;
AST, aspartate aminotransferase; TNF-α, tumor necrosis factor-α;
IL-6, interleukin-6. |
 | Figure 6Immunofluorescence staining of
infiltrated monocyte subsets in injured liver tissue and
quantification of TNF-α in mice of different genotypes by flow
cytometry. (A) Double immunoflueroscence staining of CD11b (red)
and Ly6C (green) in the sham-operated and I/R-induced liver tissues
of WT, CCR2 KO, and CCL2 KO mice (n=4–6 mice/group). The section
was then nuclear counter stained with DAPI. Scale bar, 50
µm. (B) non-parenchymal cells were isolated following sham
or I/R injury of the three groups of mice (n=4–6 mice/group) and
cultured with GolgiStop for 6 h. The cells were then stained with
CD11b, Ly6C and TNF-α. The MFI of TNF-α was measured in both
reparative monocytes (CD11b+ Ly6C low; upper panels) and
inflammatory monocytes (CD11b+ Ly6C high; lower panels)
by flow cytometry. Statistical analysis of TNF-α MFI is presented
as the mean + standard error of each group. **P<0.01.
WT, wild-type; I/R, ischemia/reperfusion; CCR2, C-C chemokine
receptor 2; KO, knockout; CCL2, C-C motif chemokine ligand 2;
TNF-α, tumor necrosis factor-α; MFI, mean fluorescence
intensity. |
CCR2 inhibitor alleviates hepatic I/R
injury
CCR2−/− or CCL2−/− mice
exhibited decreased hepatic I/R injury compared with WT mice. CCR2
has a crucial role in the CCR2/CCL2 axis and certain CCR2
inhibitors have been reported to provide renal and glycemic
benefits (41). It was tested
whether a CCR2 inhibitor could alleviate hepatic I/R injury. When
mice were pretreated with the specific CCR2 inhibitor, RS504393 (25
mg/kg), serum ALT and AST levels were significantly decreased,
compared with the control group and vehicle group (Fig. 7A). The CCR2 inhibitor group also
exhibited a decrease in the area of necrosis (Fig. 7B). In addition, inflammatory
CD11b+Ly6Chi and reparative
CD11b+Ly6Clo cells were markedly decreased
following RS504393 treatment (Fig.
7C). Similar to the magnitude of inhibition in
CCR2−/− deficient mice, CCR2 inhibition nearly entirely
suppressed CD11b+Ly6Chi cells infiltration
into the hepatic tissue after I/R injury. No difference in
CD11b+Ly6Chi and
CD11b+Ly6Clo cell populations was observed
between the untreated control group and DMSO control group. These
results demonstrate that CCR2 inhibition efficiently alleviated
hepatic I/R injury and, thus, may be an effective potential drug
for hepatic I/R injury.
Discussion
In the present study, the role of infiltrating
monocyte subsets into the liver following liver I/R injury was
investigated and demonstrated an important functional role for
inflammatory CD11b+Ly6C+ monocytes in causing
liver I/R injury. The CCR2/CCL2 axis has a critical role in
mediating monocyte infiltration into the liver following I/R
injury.
Immune cells, and the release of associated
cytokines, have been reported to be involved in the occurrence of
liver I/R injury, however the exact immunocyte populations have yet
to be determined (42).
Inflammatory cells also have a major role in liver repair and are
recruited immediately following injury (43). CD11b+Ly6C+
and CD11b+Ly6C− monocytes are the two most
well-defined subsets of monocytes and have been proposed to
infiltrate tissues during inflammation (44,45). The infiltration of
CD11b+Ly6C+ monocyte subset has been
demonstrated to initiate liver injury in infected mice and produced
pathogenic TNF (46). By
contrast, the accumulation of Ly6C−
monocytes/macrophages in the liver coincided with a drop in the
pool of Ly6C+ monocytes and provided hepatoprotective
function by secreting the anti-inflammatory cytokine IL-10
(46). The findings of the
current study provided evidence of an important role for the
recruitment of CD11b+Ly6C+ monocytes to the
site of hepatic damage following liver I/R in mice.
CD11b+Ly6Clo reparative monocytes are
predominantly recruited following acute liver I/R injury, with a
lesser extent to CD11b+Ly6Chi inflammatory
monocytes. CD11b+Ly6Chi monocytes have been
demonstrated to inhibit Th1 differentiation, but enhance
development of regulatory T cells and exhibit immunosuppressive
characteristics (14). The
subsequent release of the inflammatory cytokines, TNF-α and IL-6,
in the liver resulted in hepatocyte necrosis and elevated serum ALT
and AST levels, which suggest that hepatic injury may be in part
due to monocyte infiltration after I/R injury. Therefore, hepatic
infiltration of CD11b+Ly6Chi monocytes may be
a self-regulatory mechanism to control inflammation in I/R
injury.
The effects of macrophages in monocyte intrahepatic
recruitment most likely result from their capacity to express
numerous growth factors and cytokines (47). Macrophage depletion can be
achieved with the systemic injection of liposomes containing
clodronate (37,48). In the current study, ~90%
macrophage depletion was successfully achieved in the liver 48 h
after a single injection of liposomes containing clodronate.
Notably, the transient depletion of macrophages alleviated
I/R-induced hepatic damage, coinciding with a ecrease in
CD11b+Ly6C+ monocytes intrahepatic
infiltration. This finding strongly suggests that macrophages
contribute to the damage observed in the liver following I/R
injury.
Accumulation of Ly6Chi monocytes in the
injured liver is critically dependent on the chemokine receptor
CCR2 and its ligand, CCL2 (49).
Increasing experimental evidence suggests that CCR2 regulates
Ly6Chi monocyte entry into inflamed tissue, albeit
indirectly, by promoting the egress of Ly6Chi monocytes
from the bone marrow into blood circulation (50). CCR2 critically controls
intrahepatic Gr1hi monocyte accumulation by mediating
their egress from bone marrow (49). The findings of the current study
demonstrated that CCR2−/− and CCL2−/− mice
had almost no excess intrahepatic infiltration of Ly6C+
monocytes, indicating a similar mechanism applies during liver I/R
injury. In addition, upon organ injury, CCR2, CCL2, CCR8 and CCL1
have been previously demonstrated to promote the accumulation of
the inflammatory Ly6C+Gr1+ monocyte subset as
precursors of tissue macrophages in the liver (8). Furthermore, novel anti-CCL2 directed
agents specifically blocked the infiltration of pro-inflammatory
monocytes into injured murine liver. In the present study, specific
CCR2 inhibitor RS504393 was used to pretreat mice prior to I/R
injury and produced a magnitude of inhibition similar to that in
CCR2−/− deficient mice. CCR2 inhibitor nearly completely
suppressed the infiltration of CD11b+Ly6Chi
cells into the hepatic tissue following I/R injury. These results
suggest that CCR2 inhibition efficiently alleviated the hepatic I/R
injury and thus, may be an effective potential drug for hepatic I/R
injury. There is a limitation of the current study. Although the
internal reference gene β-actin expressed at a high level, there
are changes in the expression upon I/R treatment. Therefore, using
an additional reference gene would strengthen the reliability and
reproducibility of our study.
In conclusion, the current study defined
intrahepatic monocyte-derived subsets in experimental murine liver
I/R injury and identified that Ly6C+ monocytes have an
important role in acute I/R hepatic damage. These findings are
likely to be relevant for the treatment of liver disease. CCR2
inhibition alleviated the inflamed hepatic environment, suggesting
a potential novel approach for interventions targeting
pro-inflammatory actions of Ly6Chi monocytes in the
liver following I/R injury. However, translation from animal model
into human clinical studies requires further investigation.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81070355 and 81570570).
We also acknowledge Dr Yin Liu (College of Life Science, Wuhan
University, Wuhan, China) for providing technical assistance with
flow cytometry.
Abbreviations:
|
I/R injury
|
ischemia/reperfusion injury
|
|
CCR2
|
C-C chemokine receptor 2
|
|
CCL2
|
C-C motif chemokine ligand 2
|
|
Ly6Chi
|
Ly6C high
|
|
Ly6Clo
|
Ly6C low
|
|
ALT
|
alanine aminotransferase
|
|
AST
|
aspartate aminotransferase
|
|
NPCs
|
non-parenchymal cells
|
|
WT
|
wild-type
|
|
TNF-α
|
tumor necrosis factor-α
|
|
IL-6
|
interleukin-6
|
References
|
1
|
Peralta C, Jiménez-Castro MB and
Gracia-Sancho J: Hepatic ischemia and reperfusion injury: Effects
on the liver sinusoidal milieu. J Hepatol. 59:1094–1106. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kupiec-Weglinski JW and Busuttil RW:
Ischemia and reperfusion injury in liver transplantation.
Transplant Proc. 37:1653–1656. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Seki E, De Minicis S, Osterreicher CH,
Kluwe J, Osawa Y, Brenner DA and Schwabe RF: TLR4 enhances TGF-beta
signaling and hepatic fibrosis. Nat Med. 13:1324–1332. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jaeschke H: Mechanisms of Liver Injury.
II. Mechanisms of neutrophil-induced liver cell injury during
hepatic ischemia-reperfusion and other acute inflammatory
conditions. Am J Physiol Gastrointest Liver Physiol.
290:G1083–G1088. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abu-Amara M, Yang SY, Tapuria N, Fuller B,
Davidson B and Seifalian A: Liver ischemia/reperfusion injury:
Processes in inflammatory networks - a review. Liver Transpl.
16:1016–1032. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ju C and Tacke F: Hepatic macrophages in
homeostasis and liver diseases: From pathogenesis to novel
therapeutic strategies. Cell Mol Immunol. 13:316–327. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang F, Yin J, Lu Z, Zhang G, Li J, Xing
T, Zhuang S and Wang N: Limb ischemic preconditioning protects
against contrast-induced nephropathy via renalase. EBioMedicine.
9:356–365. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tacke F: Functional role of intrahepatic
monocyte subsets for the progression of liver inflammation and
liver fibrosis in vivo. Fibrogenesis Tissue Repair. 5(Suppl 1):
S272012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dai LJ, Li HY, Guan LX, Ritchie G and Zhou
JX: The therapeutic potential of bone marrow-derived mesenchymal
stem cells on hepatic cirrhosis. Stem Cell Res. 2:16–25. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klein I, Cornejo JC, Polakos NK, John B,
Wuensch SA, Topham DJ, Pierce RH and Crispe IN: Kupffer cell
heterogeneity: Functional properties of bone marrow derived and
sessile hepatic macrophages. Blood. 110:4077–4085. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tacke F and Randolph GJ: Migratory fate
and differentiation of blood monocyte subsets. Immunobiology.
211:609–618. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tacke F, Alvarez D, Kaplan TJ, Jakubzick
C, Spanbroek R, Llodra J, Garin A, Liu J, Mack M, van Rooijen N, et
al: Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1
to accumulate within atherosclerotic plaques. J Clin Invest.
117:185–194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Combadière C, Potteaux S, Rodero M, Simon
T, Pezard A, Esposito B, Merval R, Proudfoot A, Tedgui A and Mallat
Z: Combined inhibition of CCL2, CX3CR1, and CCR5 abrogates Ly6C(hi)
and Ly6C(lo) monocytosis and almost abolishes atherosclerosis in
hypercholesterolemic mice. Circulation. 117:1649–1657. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma H, Wan S and Xia CQ: Immunosuppressive
CD11b+Ly6Chi monocytes in pristane-induced
lupus mouse model. J Leukoc Biol. 99:1121–1129. 2016. View Article : Google Scholar
|
|
15
|
Neal LM and Knoll LJ: Toxoplasma gondii
profilin promotes recruitment of Ly6Chi CCR2+
inflammatory monocytes that can confer resistance to bacterial
infection. PLoS Pathog. 10:e10042032014. View Article : Google Scholar
|
|
16
|
Helk E, Bernin H, Ernst T, Ittrich H,
Jacobs T, Heeren J, Tacke F, Tannich E and Lotter H: TNFα-mediated
liver destruction by Kupffer cells and Ly6Chi monocytes
during Entamoeba histolytica infection. PLoS Pathog.
9:e10030962013. View Article : Google Scholar
|
|
17
|
Marra F, DeFranco R, Grappone C, Milani S,
Pastacaldi S, Pinzani M, Romanelli RG, Laffi G and Gentilini P:
Increased expression of monocyte chemotactic protein-1 during
active hepatic fibrogenesis: Correlation with monocyte
infiltration. Am J Pathol. 152:423–430. 1998.PubMed/NCBI
|
|
18
|
Denney L, Kok WL, Cole SL, Sanderson S,
McMichael AJ and HO LP: Activation of invariant NKT cells in early
phase of experimental autoimmune encephalomyelitis results in
differentiation of Ly6Chi inflammatory monocyte to M2
macrophages and improved outcome. J Immunol. 189:551–557. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gibbons MA, MacKinnon AC, Ramachandran P,
Dhaliwal K, Duffin R, Phythian-Adams AT, van Rooijen N, Haslett C,
Howie SE, Simpson AJ, et al: Ly6Chi monocytes direct
alternatively activated profibrotic macrophage regulation of lung
fibrosis. Am J Respir Crit Care Med. 184:569–581. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shimizu Y, Murata H, Kashii Y, Hirano K,
Kunitani H, Higuchi K and Watanabe A: CC-chemokine receptor 6 and
its ligand macrophage inflammatory protein 3alpha might be involved
in the amplification of local necroinflammatory response in the
liver. Hepatology. 34:311–319. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baeck C, Wehr A, Karlmark KR, Heymann F,
Vucur M, Gassler N, Huss S, Klussmann S, Eulberg D, Luedde T, et
al: Pharmacological inhibition of the chemokine CCL2 (MCP-1)
diminishes liver macrophage infiltration and steatohepatitis in
chronic hepatic injury. Gut. 61:416–426. 2012. View Article : Google Scholar
|
|
22
|
Haukeland JW, Damås JK, Konopski Z, Løberg
EM, Haaland T, Goverud I, Torjesen PA, Birkeland K, Bjøro K and
Aukrust P: Systemic inflammation in nonalcoholic fatty liver
disease is characterized by elevated levels of CCL2. Hepatol.
44:1167–1174. 2006. View Article : Google Scholar
|
|
23
|
Klueh U, Czajkowski C, Ludzinska I, Qiao
Y, Frailey J and Kreutzer DL: Impact of CCL2 and CCR2
chemokine/receptor deficiencies on macrophage recruitment and
continuous glucose monitoring in vivo. Biosens Bioelectron.
86:262–269. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yadav A, Saini V and Arora S: MCP-1:
chemoattractant with a role beyond immunity: a review. Clin Chim
Acta. 411:1570–1579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao L, Lim SY, Gordon-Weeks AN, Tapmeier
TT, Im JH, Cao Y, Beech J, Allen D, Smart S and Muschel RJ:
Recruitment of a myeloid cell subset (CD11b/Gr1 mid) via CCL2/CCR2
promotes the development of colorectal cancer liver metastasis.
Hepatology. 57:829–839. 2013. View Article : Google Scholar
|
|
26
|
Braunersreuther V, Viviani GL, Mach F and
Montecucco F: Role of cytokines and chemokines in non-alcoholic
fatty liver disease. World J Gastroenterol. 18:727–735. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Castellaneta A, Yoshida O, Kimura S,
Yokota S, Geller DA, Murase N and Thomson AW: Plasmacytoid
dendritic cell-derived IFN-α promotes murine liver
ischemia/reperfusion injury by induction of hepatocyte IRF-1.
Hepatology. 60:267–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Le Page S, Niro M, Fauconnier J, Cellier
L, Tamareille S, Gharib A, Chevrollier A, Loufrani L, Grenier C,
Kamel R, et al: Increase in Cardiac Ischemia-Reperfusion Injuries
in Opa1+/− Mouse Model. PLoS One. 11:e01640662016.
View Article : Google Scholar
|
|
29
|
Wei Q and Dong Z: Mouse model of ischemic
acute kidney injury: Technical notes and tricks. Am J Physiol Renal
Physiol. 303:F1487–F1494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wilson GC, Freeman CM, Kuethe JW, Quillin
RC III, Nojima H, Schuster R, Blanchard J, Edwards MJ, Caldwell CC
and Lentsch AB: CXC chemokine receptor-4 signaling limits
hepatocyte proliferation after hepatic ischemia-reperfusion in
mice. Am J Physiol Gastrointest Liver Physiol. 308:G702–G709. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Andrassy M, Volz HC, Igwe JC, Funke B,
Eichberger SN, Kaya Z, Buss S, Autschbach F, Pleger ST, Lukic IK,
et al: High-mobility group box-1 in ischemia-reperfusion injury of
the heart. Circulation. 117:3216–3226. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Inoue Y, Shirasuna K, Kimura H, Usui F,
Kawashima A, Karasawa T, Tago K, Dezaki K, Nishimura S, Sagara J,
et al: NLRP3 regulates neutrophil functions and contributes to
hepatic ischemia-reperfusion injury independently of inflammasomes.
J Immunol. 192:4342–4351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
He L, Sun F, Wang Y, Zhu J, Fang J, Zhang
S, Yu Q, Gong Q, Ren B, Xiang X, et al: HMGB1 exacerbates
bronchiolitis obliterans syndrome via RAGE/NF-κB/hPSe signaling to
enhance latent TGF-β release from ECM. Am J Transl Res.
8:1971–1984. 2016.
|
|
35
|
Duret C, Gerbal-Chaloin S, Ramos J, Fabre
JM, Jacquet E, Navarro F, Blanc P, Sa-Cunha A, Maurel P and
Daujat-Chavanieu M: Isolation, characterization, and
differentiation to hepatocyte-like cells of nonparenchymal
epithelial cells from adult human liver. Stem Cells. 25:1779–1790.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Van Rooijen N and Hendrikx E: Liposomes
for specific depletion of macrophages from organs and tissues.
Methods Mol Biol. 605:189–203. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Van Rooijen N and Sanders A: Liposome
mediated depletion of macrophages: Mechanism of action, preparation
of liposomes and applications. J Immunol Methods. 174:83–93. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lv Q, Yang F, Chen K and Zhang Y:
Autophagy protects podocytes from sublytic complement induced
injury. Exp Cell Res. 341:132–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rossi L, Serafini S, Antonelli A, Pierigé
F, Carnevali A, Battistelli V, Malatesta M, Balestra E, Caliò R,
Perno CF, et al: Macrophage depletion induced by clodronate-loaded
erythrocytes. J Drug Target. 13:99–111. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Heymann F, Trautwein C and Tacke F:
Monocytes and macrophages as cellular targets in liver fibrosis.
Inflamm Allergy Drug Targets. 8:307–318. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sullivan T, Miao Z, Dairaghi DJ, Krasinski
A, Wang Y, Zhao BN, Baumgart T, Ertl LS, Pennell A, Seitz L, et al:
CCR2 antagonist CCX140-B provides renal and glycemic benefits in
diabetic transgenic human CCR2 knockin mice. Am J Physiol Renal
Physiol. 305:F1288–F1297. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang F, Zhang G, Lu Z, Geurts AM, Usa K,
Jacob HJ, Cowley AW, Wang N and Liang M: Antithrombin III/SerpinC1
insufficiency exacerbates renal ischemia/reperfusion injury. Kidney
Int. 88:796–803. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brempelis KJ and Crispe IN: Infiltrating
monocytes in liver injury and repair. Clin Transl Immunology.
5:e1132016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Höchst B, Mikulec J, Baccega T, Metzger C,
Welz M, Peusquens J, Tacke F, Knolle P, Kurts C, Diehl L, et al:
Differential induction of Ly6G and Ly6C positive myeloid derived
suppressor cells in chronic kidney and liver inflammation and
fibrosis. PLoS One. 10:e01196622015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cuervo H, Guerrero NA, Carbajosa S,
Beschin A, De Baetselier P, Gironès N and Fresno M: Myeloid-derived
suppressor cells infiltrate the heart in acute Trypanosoma cruzi
infection. J Immunol. 187:2656–2665. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Morias Y, Abels C, Laoui D, Van Overmeire
E, Guilliams M, Schouppe E, Tacke F, deVries CJ, De Baetselier P
and Beschin A: Ly6C− monocytes regulate parasite-induced
liver inflammation by inducing the differentiation of pathogenic
Ly6C+ monocytes into macrophages. PLoS Pathog.
11:e10048732015. View Article : Google Scholar
|
|
47
|
Alisi A, Carpino G, Oliveira FL, Panera N,
Nobili V and Gaudio E: The role of tissue macrophage-mediated
inflammation on NAFLD pathogenesis and its clinical implications.
Mediators Inflamm. 8162421:2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Weisser SB, van Rooijen N and Sly LM:
Depletion and reconstitution of macrophages in mice. J Vis Exp. Aug
1–2012.Epub ahead of print. View
Article : Google Scholar : PubMed/NCBI
|
|
49
|
Karlmark KR, Weiskirchen R, Zimmermann HW,
Gassler N, Ginhoux F, Weber C, Merad M, Luedde T, Trautwein C and
Tacke F: Hepatic recruitment of the inflammatory Gr1+
monocyte subset upon liver injury promotes hepatic fibrosis.
Hepatology. 50:261–274. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Engel DR, Maurer J, Tittel AP, Weisheit C,
Cavlar T, Schumak B, Limmer A, van Rooijen N, Trautwein C, Tacke F,
et al: CCR2 mediates homeostatic and inflammatory release of
Gr1(high) monocytes from the bone marrow, but is dispensable for
bladder infiltration in bacterial urinary tract infection. J
Immunol. 181:5579–5586. 2008. View Article : Google Scholar : PubMed/NCBI
|