Introduction
Thyroid cancer is the most common cancer in the
endocrine system. The incidence of thyroid cancer is increasing
every year in the United States, and the increase is not only
because of the better diagnosis technique for thyroid cancers.
There are estimated to be ~123,000 people in the world who are
diagnosed thyroid cancer cases annually, accounting for
approximately over 91.5% of the cancers of head and neck (1). Notably, papillary thyroid carcinoma
(PTC) is the most common type of all thyroid cancers, which
accounts for ~80% of the thyroid cancers.
Metabolomics is an emerging omics technology, and it
can systematically identify and quantify metabolites in a
biological sample. The technology development of nuclear magnetic
resonance (NMR) spectroscopy and mass spectrometry (MS) allows the
simultaneous analysis of various chemicals in the biological
sample, and therefore NMR and MS are the favorable platforms for
metabolomic analysis (2).
Metabolomics has been widely used in studies regarding diseases
(3), and it identifies metabolic
signatures which are related to the pathogenesis of the diseases
(3–5). There are metabolomic studies on PTC
based on gas chromatography-mass spectrometry (GC-MS) (6,7).
Compared with GC-MS, NMR is generally accepted as the gold standard
in metabolite structural elucidation, which is highly selective,
non-destructive for chemical detection. Additionally, the
metabolite coverage is different between NMR and GC-MS (8). Therefore, different metabolomic
information can be provided by NMR and GC-MS. There are limited
published metabolomic studies regarding PTC using NMR (9–11),
but these studies focused on the diagnosis of PTC, and therefore
the detailed metabolic changes especially metabolic pathway changes
related to PTC pathogenesis are still largely unknown. Given these
facts, it is necessary to adopt NMR based metabolomics technology
to study the detailed metabolic changes and disturbed metabolic
pathways in PTC.
We used 1H NMR-based metabolomic
technique to study the metabolic changes in PTC. We established
reliable statistical models which could well discriminate and
predict the tumor and normal thyroid tissue. Based on these models
and further bioinformatics analysis, we identified detailed
metabolic changes and disturbed metabolic pathways in PTC.
Patients and methods
Basic patient information and sample
collection
A total of 16 patients (4 males, 12 females; age
range, 19–59 years; tumor size, 1–4.2 cm) had undergone surgical
thyroidectomy at the First Affiliated Hospital of Nanjing Medical
University. Previous metabolomic studies using cancer tissues have
proved that similar sample number and characteristics of the
population can provide useful information on metabolic changes of
cancer (4,9–11).
Matched normal thyroid tissues and tumor tissues were obtained from
the same PTC patients. The diagnosis was confirmed by
histopathologic evaluation based on the established criteria of WHO
(12), which was conducted
independently by two pathologists. The analyzed tumor tissues which
contained over 90% cancer cells were obtained by using
microdissection, and normal tissues were not connected by
follicular adenomas (FAs) or PTC. The patients were not received
radiation therapy or neo-adjuvant chemotherapy before operation.
All tissues were immediately frozen in liquid nitrogen, and then
stored at −80°C until NMR analysis. The study was approved by the
Ethics Committee of the First Affiliated Hospital of Nanjing
Medical University, and each participant signed an informed
consent.
1H NMR spectroscopy based
metabolomic analysis
Metabolomic analysis was conducted according to
metabolomic procedure for NMR spectroscopy of tissues (13). The metabolomic analysis was
performed with Bruker Avance III 600 NMR spectrometer. Tissue
samples were placed in a 4 mm rotor, and 5 µl aliquot of
deuterium oxide was added into the rotor. The resonance frequency
of 1H was 400 MHz, and the experimental temperature was
298 K. A Carr-Purcell-Meiboom-Gill (CPMG) filter was included in
the pulse sequence to reduce broad resonances related to background
or macromolecules. After the fourier transformation, the spectra
was manually phased and baseline corrected, and then referenced to
lactic acid CH3 resonance at δ1.33.
Multivariate pattern recognition
In order to discriminate the samples according to
tissue types and identify potential biomarkers in relation to PTC,
principal component analysis (PCA), partial least
squares-discriminant analysis (PLS-DA), and the orthogonal partial
least-squares discriminant analysis (OPLS-DA) were applied using
the normalized NMR data. We identified changed metabolites in PTC
by using OPLS-DA statistical model. SIMCA-P + software (v11.0;
Umetrics AB, Umea, Sweden) was used for data analysis. The
correlation coefficient of |r| >0.482 and the variable
importance in projection (VIP) value >1.00 were used as cut-off
value of the statistical significance based on the discrimination
significance at the level of P=0.05.
Biological significance interpretation
and informatics analysis
To identify detailed metabolic changes and disturbed
metabolic pathways, the differentiated metabolites were first
annotated with 'Kyoto Encyclopedia of Genes and Genomes' (KEGG,
http://www.genome.jp/kegg/or http://www.kegg.jp/) which is a knowledge library for
systematic analysis of metabolite functions and networks and 'Human
Metabolome Database' (HMDB, http://www.hmdb.ca/) which is a complete and
comprehensive database on metabolomics with metabolite biological
interpretation (14,15). Further, we conducted informatics
analysis by MetaboAnalyst 3.0 (http://www.metaboanalyst.ca/MetaboAnalyst/) which is
built by R software (v3.2.2) (16). MetaboAnalyst combines the results
from powerful pathway enrichment analysis with the pathway topology
analysis to discover the relevant pathways. Two modules of
MetaboAnalyst including pathway analysis and enrichment analysis
were used. The module of pathway analysis was based on the KEGG
database; the enrichment analysis was conducted based on Small
Molecule Pathway Database (SMPDB, http://smpdb.ca/)
(17).
Results
Metabolomic profiling
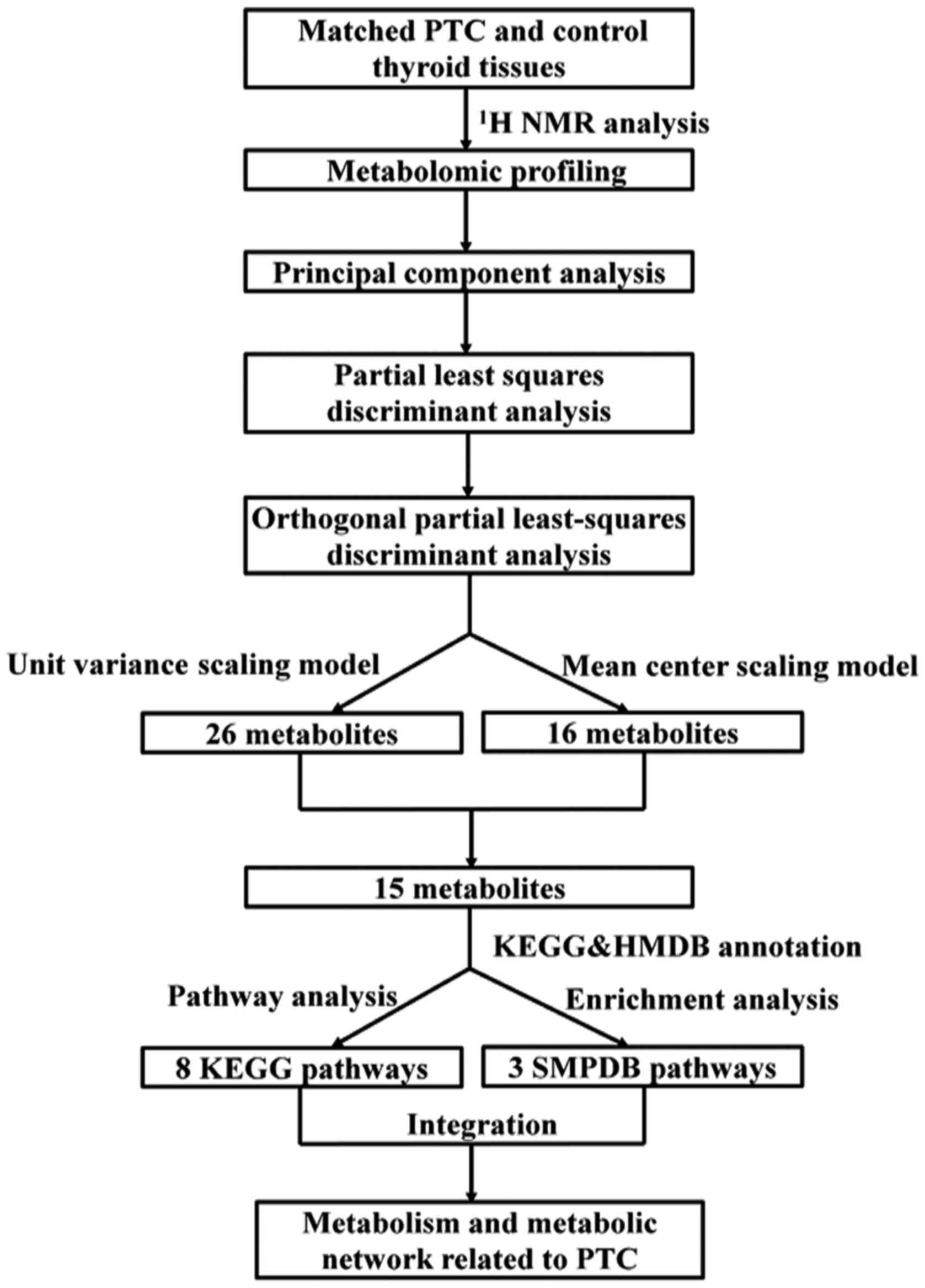
Fig. 1 shows the
workflow of this metabolomic study. The metabolomic profiles
obtained from PTC and normal thyroid tissue are shown in Fig. 2. The metabolic profiling covered
branched chain amino acids (isoleucine, leucine and valine), other
proteinogenic amino acids (alanine, phenylalanine, tyrosine,
glycine, aspartate, glutamate and lysine), and product of amino
acid (creatine) as well as amino acid derivatives (sarcosine and
taurine). Metabolites involved in the nucleotide metabolism
(hypoxanthine, uracil, uridine and allantoin), glycolysis
(lactate), tricarboxylic acid cycle (citrate and succinate),
choline metabolism (choline, ethanolamine, glycerolphosphocholine
and N,N-dimethylglycine) and lipid metabolism
[very-low-density lipoprotein (VLDL), low-density lipoprotein (LDL)
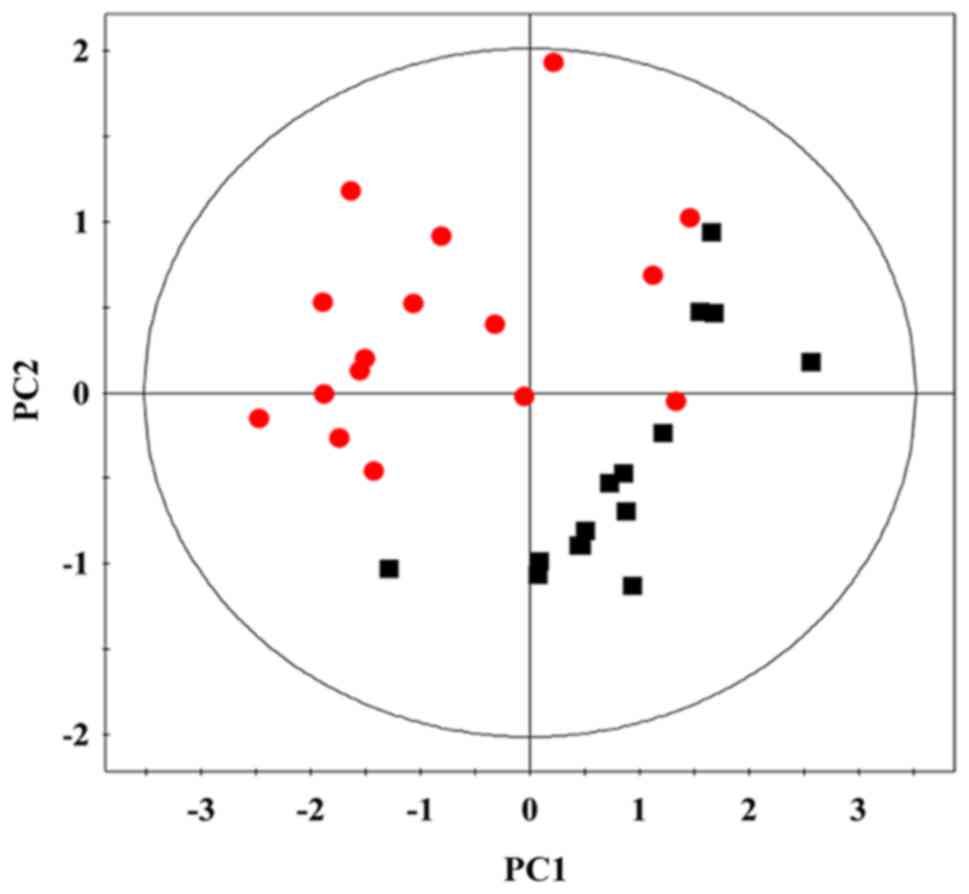
and lipids] were also profiled.PCA. PCA is the most commonly used
algorithm in metabolomics studies (18). We adopted PCA in the study to
process the NMR data based on a mean center-scaling model, which is
an unsupervised projection method used to visualize the dataset and
display the intrinsic similarity and difference in the dataset. As
shown in Fig. 3 (19), the PTC tissues were clearly
discriminated from normal tissues using PCA model, indicating
dramatic metabolic changes in PTC. The PCA model descriptors
(R2X, Q2) were 0.79/0.75, indicating the
model was reliable.
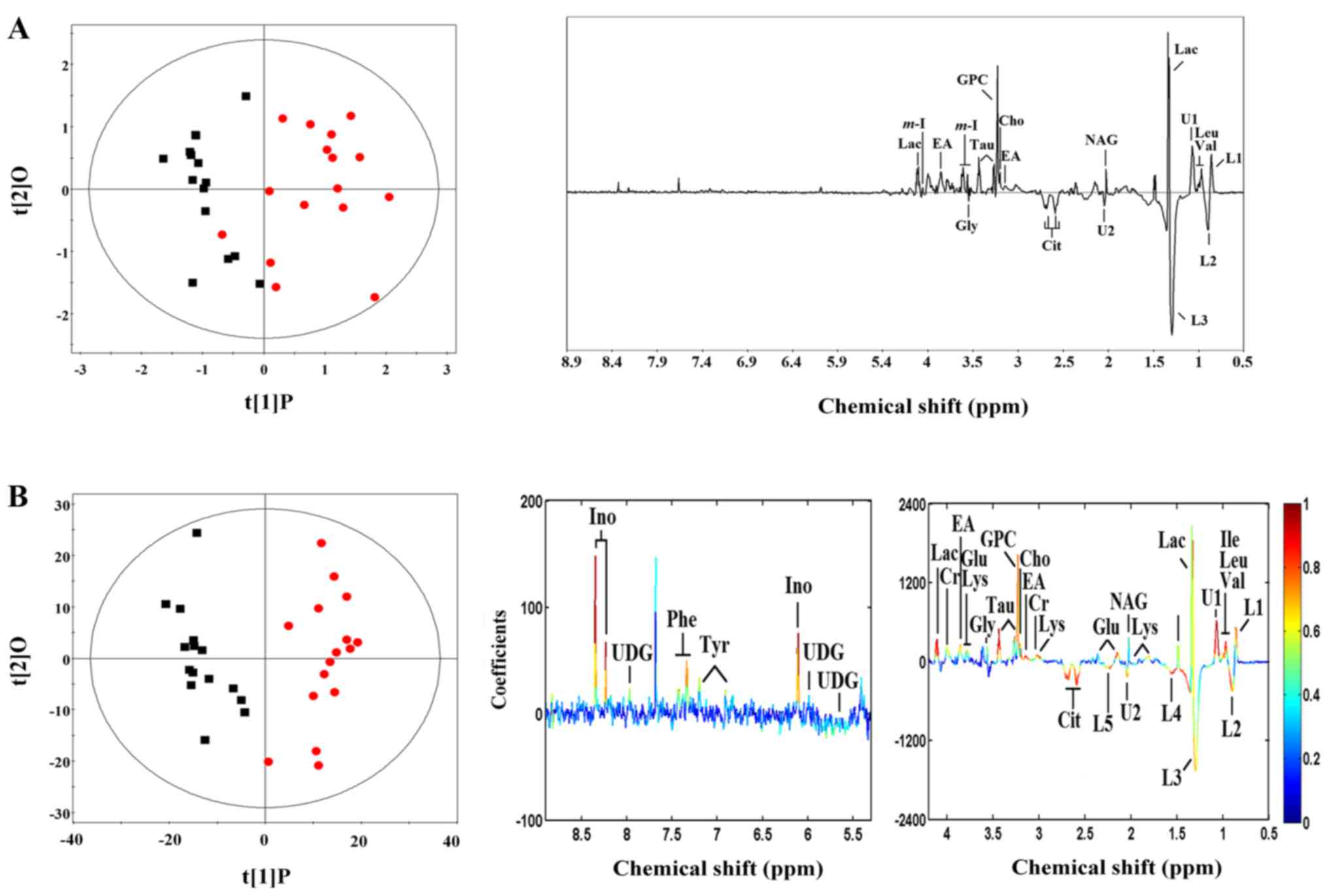
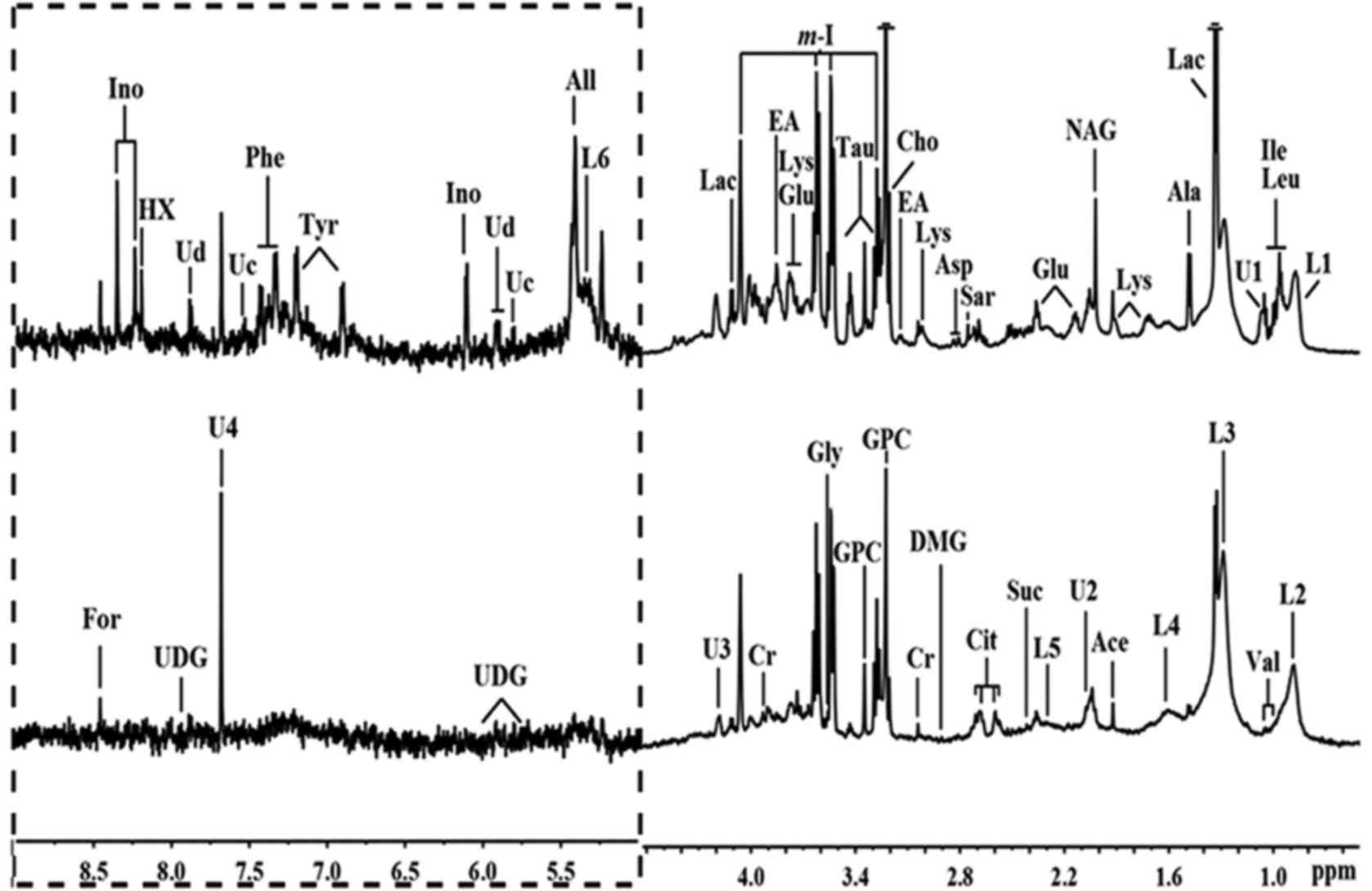 | Figure 2Representative 1H nuclear
magnetic resonance (NMR) spectra (δ0.6–4.6 and δ5.0–9.0) of thyroid
obtained from papillary thyroid carcinoma (PTC) (above) and normal
thyroid tissue (below). The region of δ5.0–9.0 (in the dashed box)
was magnified 10 times compared with corresponding region of
δ0.6–4.6 for the purpose of clarity. Ace, acetate; Ala, alanine;
All, allantoin; Asp, aspartate; Cho, choline; Cit, citrate; Cr,
creatine; EA, ethanolamine; For, formate; Glu, glutamate; Gly,
glycine; GPC, glycerolphosphocholine; HX, hypoxanthine; Ile,
isoleucine; Ino, inosine; L1: LDL, CH3–(CH2)n–;
L2: VLDL, CH3–(CH2)n–;
L3: VLDL, CH3–(CH2)n–;
L4: VLDL, –CH2–CH2–C=O;
L5: lipid, –CH2–C=O; L6: lipid, –CH=CH–; Lac, lactate;
Leu, leucine; Lys, lysine; m-I, myo-inositol; DMG,
N,N-dimethylglycine; NAG, N-acetyl
glycoprotein signals; Phe, phenylalanine; Sar, sarcosine; Suc,
succinate; Tau, taurine; Tyr, tyrosine; U, unknown; Uc, uracil; Ud,
uridine; UDG, uridine diphosphate glucose; Val, valine. |
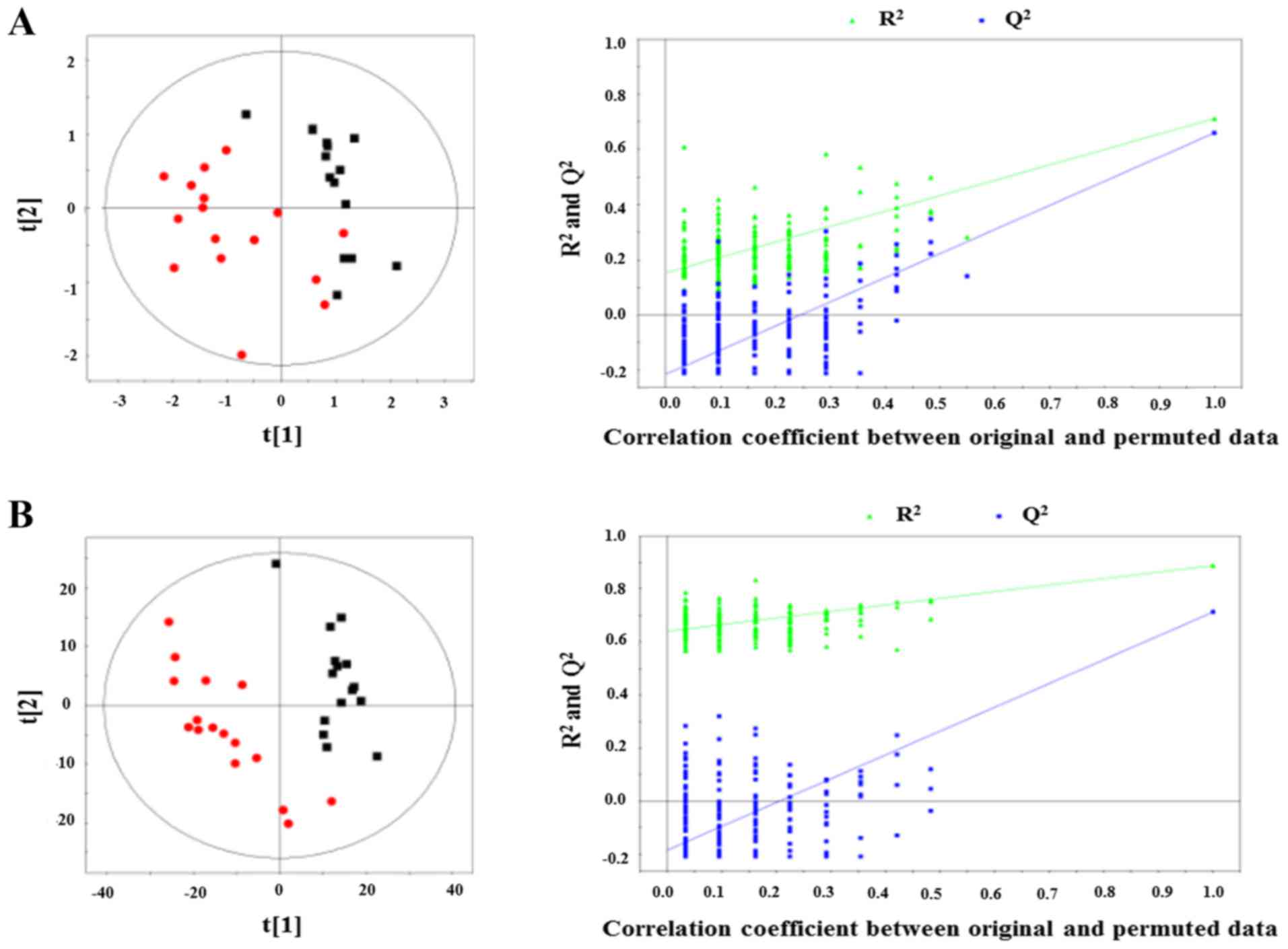
PLS-DA
PLS-DA was further conducted to evaluate the data
variance between PTC and normal tissue. As shown in Fig. 4A (20), PLS-DA score plot showed a
statistically significant discrimination between the two groups
based on the mean center-scaling model (R2X= 0.79,
R2Y= 0.71, Q2= 0.66). Similar significant
discrimination between the two groups was also observed based on
the unit variance scaling model (R2X=0.26,
R2Y= 0.82, Q2=0.71) (Fig. 4B) (20). To test the validity of the PLS-DA
models, the robustness of the models was assessed using a
200-permutations validation model, which showed that the originally
observed separation was not due to a random effect, as the
predictive discrimination values of the random models were all
lower than that of the original model.
OPLS-DA
Additional OPLS-DA was conducted to establish the
mean center-scaling model and the unit variance scaling model. The
mean center-scaling model descriptors (R2X,
Q2) were 0.79/0.66 (Fig.
5A), and the unit variance scaling mode descriptors
(R2X, Q2) were 0.26/0.77 (Fig. 5B), indicating these models were
reliable.
Differential metabolites in PTC
Based on the unit variance scaling OPLS-DA model,
there were 26 differential metabolites identified. The differential
metabolites in PTC are presented in Table I and Fig. 5B. To find the robustly changed
metabolites, we verified the changed metabolites in the mean
center-scaling model, which identified 16 differential metabolites
(Table II and Fig. 5A), among which 15 consistently
changed metabolites were found (Table III).
 | Table IOPLS-DA coefficients derived from the
NMR data of metabolites in thyroid obtained from PTC and normal
tissue in the unit variance scaling mode. |
Table I
OPLS-DA coefficients derived from the
NMR data of metabolites in thyroid obtained from PTC and normal
tissue in the unit variance scaling mode.
| Metabolites | ra |
|---|
| Alanine:
1.48(db) | 0.738 |
| Choline:
3.20(s) | 0.758 |
| Citrate: 2.57(d),
2.69(d) | −0.792 |
| Creatine: 3.04(s),
3.93(s) | 0.594 |
| Ethanolamine:
3.14(t), 3.85(t) | 0.858 |
| Glutamate: 2.15(m),
2.35(m), 3.78(t) | 0.831 |
| Glycine:
3.56(s) | 0.566 |
|
Glycerophosphocholine: 3.23(s),
3.35(s) | 0.754 |
| Isoleucine:
0.94(d), 1.01(d) | 0.597 |
| Inosine: 6.10(d),
8.23(s), 8.34(s) | 0.897 |
| L1, LDL,
CH3–(CH2)n–:
0.86(br) | 0.732 |
| L2, VLDL,
CH3–(CH2)n–:
0.90(br) | −0.762 |
| L3, VLDL,
CH3–(CH2)n–:
1.29(br) | −0.628 |
| L4, VLDL,
–CH2–CH2–C=O:
1.59(br) | −0.822 |
| L5, Lipid,
–CH2–C=O: 2.26(br) | −0.738 |
| Lactate: 1.33(d),
4.11(q) | 0.893 |
| Leucine:
0.96(t) | 0.767 |
| Lysine: 1.72(m),
1.91(m), 3.01(m), 1.76(t) | 0.750 |
| N-acetyl
glycoprotein signals: 2.03(s) | 0.520 |
| Phenylalanine:
7.33(d), 7.37(t), 7.43(dd) | 0.759 |
| Taurine: 3.28(t),
3.43(t) | 0.861 |
| Tyrosine: 6.90(d),
7.20(d) | 0.532 |
| Unknown-1:
1.07(m) | 0.854 |
| Unknown-2:
2.06(m) | −0.679 |
| Uridine diphosphate
glucose: 5.61(dd), 5.98(m), 7.96(d) | 0.569 |
| Valine: 0.99(d),
1.04(d) | 0.889 |
| Table IIVIP value derived from the NMR data
of metabolites in thyroid obtained from PTC and normal tissue in
the mean center-scaling model. |
Table II
VIP value derived from the NMR data
of metabolites in thyroid obtained from PTC and normal tissue in
the mean center-scaling model.
| Metabolites | VIPa |
|---|
| Lactate:
1.33(db), 4.11(q) | 10.81 |
| L3, VLDL,
CH3–(CH2)n–:
1.29(d) | 9.81 |
|
Glycerophosphocholine: 3.23(s),
3.35(s) | 9.24 |
| Unknown-1:
1.07(m) | 3.63 |
| L1, LDL,
CH3–(CH2)n–:
0.86(br) | 3.05 |
| Valine: 0.99(d),
1.04(d) | 2.81 |
| L2, VLDL,
CH3–(CH2)n–:
0.90(br) | 2.62 |
| Taurine: 3.28(t),
3.43(t) | 2.44 |
| N-acetyl
glycoprotein signals: 2.03(s) | 2.29 |
| Myo-inositol:
3.56(dd), 3.63(dd), 4.06(t) | 2.19 |
| Citrate: 2.57(d),
2.69(d) | 1.81 |
| Leucine:
0.96(t) | 1.63 |
| Ethanolamine:
3.14(t), 3.85(t) | 1.63 |
| Choline:
3.20(s) | 1.26 |
| Unknown-2:
2.06(m) | 1.13 |
| Glycine:
3.56(s) | 1.04 |
| Table IIIThe consistently changed metabolites
in thyroid obtained from PTC and normal tissue in the OPLS-DA
models. |
Table III
The consistently changed metabolites
in thyroid obtained from PTC and normal tissue in the OPLS-DA
models.
| Metabolites | KEGG | HMDB | ra | VIPb | Pathway |
|---|
| Choline | C00114 | HMDB0097 | 0.758 | 1.26 | Choline
metabolism |
| Ethanolamine | C00189 | HMDB0149 | 0.858 | 1.63 | Choline
metabolism |
|
Glycerophosphocholine | C00670 | HMDB0086 | 0.754 | 9.24 | Choline
metabolism |
| Lactate | C00186 | HMDB0190 | 0.893 | 10.81 | Glycolysis |
| Citrate | C00158 | HMDB0094 | −0.792 | 1.81 | Tricarboxylic acid
cycle |
| Leucine | C00123 | HMDB0687 | 0.767 | 1.63 | Branched chain
amino acid metabolism |
| Valine | C00183 | HMDB0883 | 0.889 | 2.81 | Branched chain
amino acid metabolism |
| Glycine | C00037 | HMDB0123 | 0.566 | 1.04 | Other amino acid
metabolism |
| Taurine | C00245 | HMDB0251 | 0.861 | 2.44 | Other amino acid
metabolism |
| L1, LDL | NA | NA | 0.732 | 3.05 | Lipid
metabolism |
| L2, VLDL | NA | NA | −0.762 | 2.62 | Lipid
metabolism |
| L3, VLDL | NA | NA | −0.628 | 9.81 | Lipid
metabolism |
| N-acetyl
glycoprotein signals | NA | NA | 0.520 | 2.29 | NA |
| Unknown-1:
1.07(m) | NA | NA | 0.854 | 3.63 | NA |
| Unknown-2:
2.06(m) | NA | NA | −0.679 | 1.13 | NA |
Metabolic pathway analysis and biological
significance interpretation
The 15 metabolites related to PTC with robust
results were then annotated with KEGG and HMDB (Table III). The results were submitted
to MetaboAnalyst to show the statistical analysis results of
informatics analysis. The result of pathway analysis is shown in
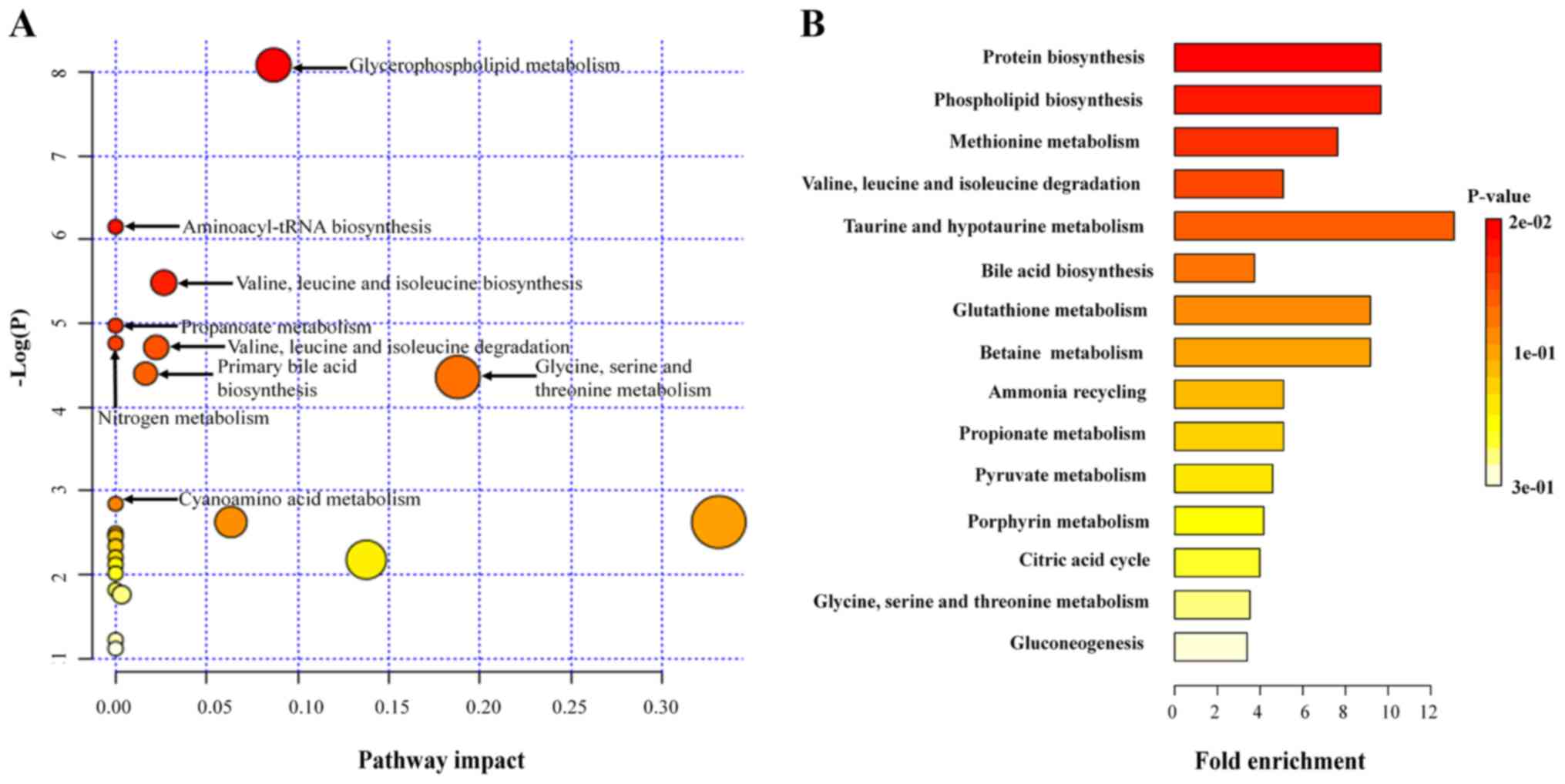
Table IV and Fig. 6A, which found 8 pathways including
glycerophospholipid metabolism, aminoacyl-tRNA biosynthesis,
valine, leucine and isoleucine biosynthesis, propanoate metabolism,
nitrogen metabolism, valine, leucine and isoleucine degradation,
primary bile acid biosynthesis, and glycine, serine and threonine
metabolism were significantly related to PTC. Furthermore, in order
to expand the understanding of metabolic pathway related to PTC,
the module of enrichment analysis of MetaboAnalyst was used, which
found 3 additional pathways including protein biosynthesis,
phospholipid biosynthesis and methionine metabolism significantly
related to PTC (Table V and
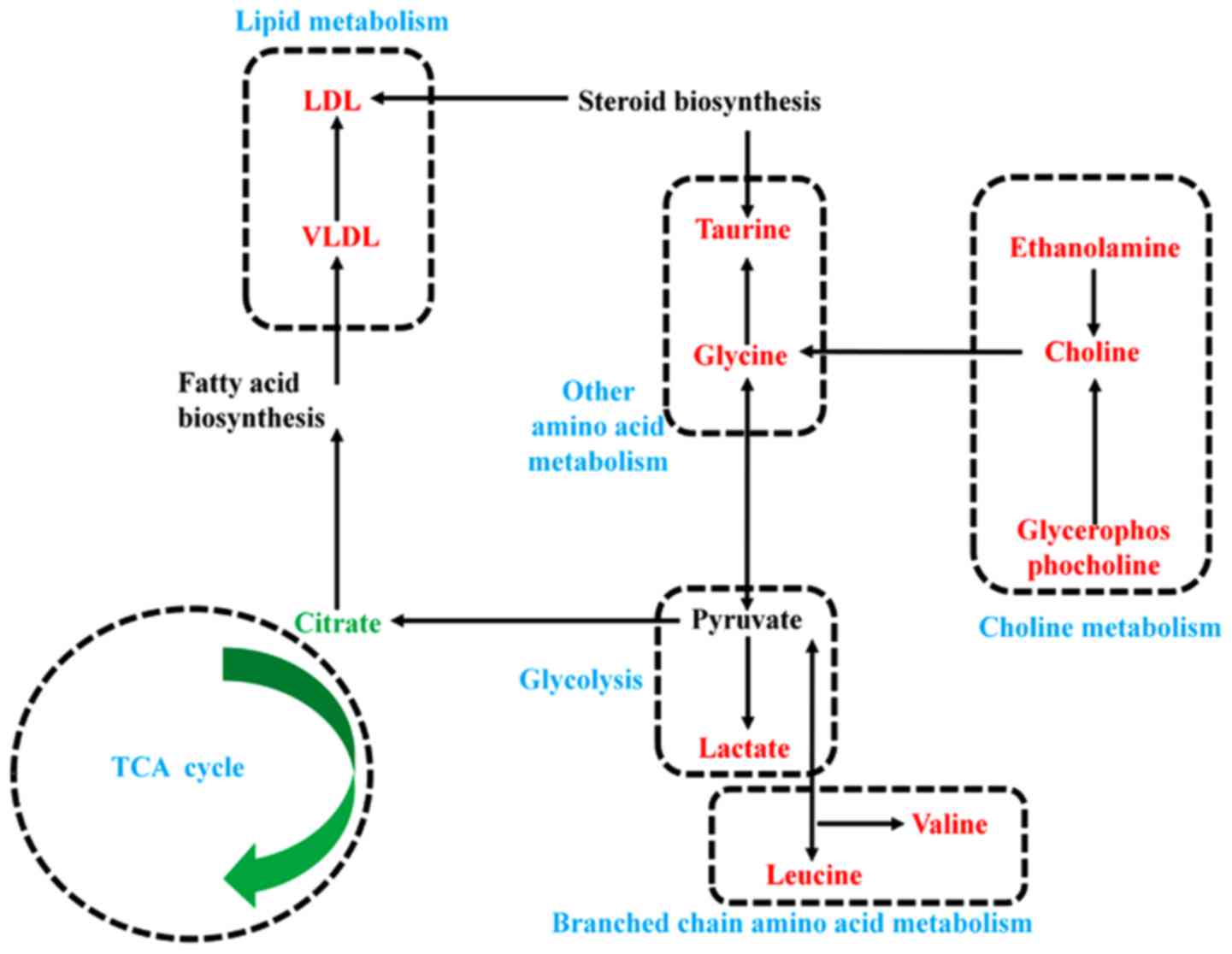
Fig. 6B). Finally, based on
biological significance and the above informatics analysis, the
metabolic network related to PTC was built (Fig. 7), which indicated the key
metabolisms related to PTC including branched chain amino acid
metabolism (leucine and valine), other amino acid metabolism
(glycine and taurine), glycolysis (lactate), tricarboxylic acid
cycle (citrate), choline metabolism (choline, ethanolamine and
glycerolphosphocholine) and lipid metabolism (VLDL and LDL).
| Table IVPathway analysis of metabolic changes
in PTC.a |
Table IV
Pathway analysis of metabolic changes
in PTC.a
| KEGG pathway | Total | Expected | Hits | Impact | P-value |
|---|
| Glycerophospholipid
metabolism | 39 | 0.146 | 3 | 0.087 | 3.09E-04 |
| Aminoacyl-tRNA
biosynthesis | 75 | 0.280 | 3 | 0.000 | 2.13E-03 |
| Valine, leucine and
isoleucine biosynthesis | 27 | 0.101 | 2 | 0.027 | 4.16E-03 |
| Propanoate
metabolism | 35 | 0.131 | 2 | 0.000 | 6.94E-03 |
| Nitrogen
metabolism | 39 | 0.146 | 2 | 0.000 | 8.57E-03 |
| Valine, leucine and
isoleucine degradation | 40 | 0.150 | 2 | 0.022 | 9.01E-03 |
| Primary bile acid
biosynthesis | 47 | 0.176 | 2 | 0.016 | 1.23E-02 |
| Glycine, serine and
threonine metabolism | 48 | 0.179 | 2 | 0.188 | 1.28E-02 |
| Cyanoamino acid
metabolism | 16 | 0.060 | 1 | 0.000 | 5.84E-02 |
| Citrate cycle (TCA
cycle) | 20 | 0.075 | 1 | 0.063 | 7.25E-02 |
| Taurine and
hypotaurine metabolism | 20 | 0.075 | 1 | 0.331 | 7.25E-02 |
| Ether lipid
metabolism | 23 | 0.086 | 1 | 0.000 | 8.29E-02 |
| Thiamine
metabolism | 24 | 0.090 | 1 | 0.000 | 8.64E-02 |
| Pantothenate and
CoA biosynthesis | 27 | 0.101 | 1 | 0.000 | 9.67E-02 |
| Glycolysis or
gluconeogenesis | 31 | 0.116 | 1 | 0.000 | 1.10E-01 |
| Pyruvate
metabolism | 32 | 0.120 | 1 | 0.138 | 1.14E-01 |
| Methane
metabolism | 34 | 0.127 | 1 | 0.000 | 1.20E-01 |
| Glutathione
metabolism | 38 | 0.142 | 1 | 0.000 | 1.34E-01 |
| Lysine
degradation | 47 | 0.176 | 1 | 0.000 | 1.63E-01 |
| Glyoxylate and
dicarboxylate metabolism | 50 | 0.187 | 1 | 0.003 | 1.72E-01 |
| Purine
metabolism | 92 | 0.344 | 1 | 0.000 | 2.96E-01 |
| Porphyrin and
chlorophyll metabolism | 104 | 0.389 | 1 | 0.000 | 3.28E-01 |
| Table VPathway enrichment of metabolic
changes in PTC.a |
Table V
Pathway enrichment of metabolic
changes in PTC.a
| Pathway from
SMPDB | Total | Expected | Hits | P-value |
|---|
| Protein
biosynthesis | 19 | 0.207 | 2 | 1.64E-02 |
| Phospholipid
biosynthesis | 19 | 0.207 | 2 | 1.64E-02 |
| Methionine
metabolism | 24 | 0.262 | 2 | 2.58E-02 |
| Valine, leucine and
isoleucine degradation | 36 | 0.393 | 2 | 5.50E-02 |
| Taurine and
hypotaurine metabolism | 7 | 0.076 | 1 | 7.42E-02 |
| Bile acid
biosynthesis | 49 | 0.535 | 2 | 9.52E-02 |
| Glutathione
metabolism | 10 | 0.109 | 1 | 1.04E-01 |
| Betaine
metabolism | 10 | 0.109 | 1 | 1.04E-01 |
| Ammonia
recycling | 18 | 0.196 | 1 | 1.81E-01 |
| Propanoate
metabolism | 18 | 0.196 | 1 | 1.81E-01 |
| Pyruvate
metabolism | 20 | 0.218 | 1 | 1.99E-01 |
| Porphyrin
metabolism | 22 | 0.240 | 1 | 2.17E-01 |
| Citric acid
cycle | 23 | 0.251 | 1 | 2.26E-01 |
| Glycine, serine and
threonine metabolism | 26 | 0.284 | 1 | 2.51E-01 |
|
Gluconeogenesis | 27 | 0.295 | 1 | 2.60E-01 |
Discussion
Based on the rapid development of analytical
techniques, metabolomics has been applied in many fields such as
biochemical and clinical study (21–24). In the study of human diseases,
metabolomics takes advantage of novel biomarker exploration and
pathophysiological interpretation at the molecular level (21–24). However, until now, there are only
NMR-based metabolomic studies focusing on the diagnosis of PTC
(9–11), and therefore the detailed
metabolic changes potentially related to PTC pathogenesis are still
largely unknown. In this study, based on the results of PCA,
PLS-DA, OPLS-DA models (Figs.
3Figure 4–5), we first identified key metabolites
related to PTC (Table III).
Then, after KEGG and HMDB annotation and following pathway and
enrichment analysis, we found significant metabolic pathways
related to PTC (Fig. 6), through
which we found the metabolic network related to PTC, indicating the
metabolic changes potentially related to PTC pathogenesis (Fig. 7).
Robust changes of metabolite levels in PTC included
increased lactate, which participates in significantly changed KEGG
pathway of propanoate metabolism directly related to glycolysis.
The significant increase of lactate in other cancers such as colon
cancer and prostate cancer has been reported in previous studies
(25,26). This metabolic response in PTC
indicated an increase of the glycolytic flux due to hypoxia and
ischemia in the tumor tissues or the consequence of the so-called
'Warburg effect', producing more waste products such as lactate
(27). Previous GC-MS based
metabolomics study did not monitor lactate but found the increased
expression of the gene LDHA encoding lactate dehydrogenase which
catalyzes the synthesis of lactate in PTC (7), which was consistent with the present
study. Citrate which was a significantly decreased metabolite in
PTC tissue participates in the tricarboxylic acid cycle which is
directly related to glycolysis. Previous study also revealed that
the decreased tricarboxylic acid cycle was accompanied by increased
glycolytic flux in gastric cardia cancer due to decreased pyruvic
acid efflux into the tricarboxylic acid cycle (28). Normal tricarboxylic acid cycle may
have an inhibitory effect on cancer progression (28). Collectively, these findings
revealed increased glycolysis and inhibited tricarboxylic acid
cycle in PTC, which may be biologically related to PTC
pathogenesis.
Notably, the robust changes of amino acids in PTC
include leucine and valine. Leucine and valine were also included
in significantly changed KEGG pathway such as amino acyl-tRNA
biosynthesis, and valine, leucine and isoleucine biosynthesis and
degradation as well as SMPDB pathway including protein
biosynthesis. Branched-chain amino acids include isoleucine,
leucine and valine. Recent metabolomics studies have consistently
revealed that branched-chain amino acids is positively related to
obesity (29) and diabetes
(23,29), which are both endocrine
abnormalities and associated with the risk of PTC (30). In the unit variance scaling mode,
isoleucine was also identified as the biomarker with increased
level in PTC. Additionally, isoleucine and leucine have been
identified as tumor promoters of bladder cancer (31). Glycine is involved in the body's
production of DNA, phospholipids and collagen, as well as in
release of energy. It participates in significantly changed KEGG
pathways such as glycine, serine and threonine metabolism. It is
reported that glycine plays an important role in rapid cancer cell
proliferation (32). Amino acid
derivative, the increase of taurine was found in PTC. Taurine was
identified as a possible fingerprint biomarker in non-muscle
invasive bladder cancer (33). It
participates in significantly changed KEGG pathways of
aminoacyl-tRNA biosynthesis, indicating the abnormal protein
biosynthesis in PTC. As suggested by Tessem et al in colon
cancer, the increase of taurine may reflect an imbalance in
osmolyte function in cancer cells (25). Therefore, the increased levels of
branched chain amino acids, glycine and taurine in PTC should
attract attention as they may be the oncogenic biomarkers of PTC
29).
Significant KEGG pathway including
glycerophospholipid metabolism and SMPDB pathways including
phospholipid biosynthesis as well as methionine metabolism both are
related to choline metabolism covering choline, ethanolamine and
glycerophosphocholine. Glycerophosphorylcholine is a choline
derivative and one of the major forms of choline storage. Indeed,
the abnormal choline metabolism has been reported in breast cancer
(34). As indicated in SMPDB,
choline metabolism is related to the body methylation status which
is associated with thyroid carcinoma pathogenesis (35). In this study, the metabolites in
lipid metabolism including VLDL and LDL also changed in PTC. This
finding is consistent with blood lipid profile alterations in
another malignant disease, acute lymphoblastic leukemia (36).
It is urgent to seek new biomarkers for thyroid
carcinoma. Metabolomic profiles from tissue have the potential to
be used in conjunction with current diagnostics to help guide the
clinical management of patients with PTC (9–11).
Our study not only established robust multivariate analysis models
that could discriminate PTC, but also identified robust metabolic
signatures of PTC based on different statistical models. The
robustly changed metabolites identified in this study may be used
as potential biomarkers for PTC, among which consistent results
were found on lactate and taurine in another metabolomics study
regarding PTC (10).
In conclusion, in this study, we found that the
metabolomic profiling could discriminate PTC in conjunction with
multivariate analysis, and identified robust metabolic signatures
of PTC. After informatics analysis, we found the PTC is
characterized with increased glycolysis and inhibited tricarboxylic
acid cycle, increased oncogenic amino acids as well as abnormal
choline and lipid metabolism, which needs further research to
deeply study the underlying mechanism and the usage of the study
findings in the intervention of PTC. The findings in this study
provide new insights into metabolic changes of PTC, and hold great
potential in the treatment of PTC.
Glossary
Abbreviations
Abbreviations:
|
PTC
|
papillary thyroid carcinoma
|
|
NMR
|
nuclear magnetic resonance
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
HMDB
|
human metabolome database
|
|
PCA
|
principal component analysis
|
|
PLS-DA
|
partial least squares discriminant
analysis
|
|
OPLS-DA
|
orthogonal partial least-squares
discriminant analysis
|
|
SMPDB
|
Small Molecule Pathway Database
|
|
VLDL
|
very-low-density lipoprotein
|
|
LDL
|
low-density lipoprotein
|
|
MS
|
mass spectrometry
|
|
GC-MS
|
gas chromatography-mass
spectrometry
|
|
VIP
|
variable importance in the
projection
|
|
FAs
|
follicular adenomas
|
|
CPMG
|
Carr-Purcell-Meiboom-Gill
|
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by the National
Natural Science Foundation (no. 81770773), the Natural Science
Foundation of Jiangsu Province (no. BK20171499); the Jiangsu
Province Key Medical Talents (co-construction) Program, the Talent
Project of '333' Project in Jiangsu Province, the Jiangsu Province
Official Hospital Scientific Research Initial Funding (no.
RPF201501), the Jiangsu Province Official Hospital Talents
Construction Fund Research Project (no. IR2015101), the National
Natural Science Foundation (no. 81402713), the Young Scholars of
Jiangsu Province (no. BK20140909), the Innovation Fund Project of
the State Key Laboratory of Reproductive Medicine (general) (no.
SKLRM-GA201802). The funding bodies had no role in the design of
the study or collection, analysis, and interpretation of data or in
writing the manuscript.
[2] Availability
of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
YL performed the metabolomic analysis and drafted
the manuscript. MC analyzed and interpreted the metabolomic data,
and made substantial contributions to manuscript revision. CL
participated in sample collection and sample preparation for
metabolomic analysis. YX provided analytical tools, and contributed
to study design and metabolic network establishment. BX was a major
contributor in sample preparation, and participated in the
instrumental analysis. YH performed metabolic pathway analysis, and
prepared figures. TC participated in multivariate pattern
recognition analysis and table preparation. MS collected the
sample, and was a major contributor in study design. WT made
substantial contributions to conception and design, and was
involved in acquisition of data and manuscript revision. All
authors read and approved the final manuscript.
[4] Ethics
approval and consent to participate
The study was approved by the Ethics Committee of
the First Affiliated Hospital of Nanjing Medical University, and
each participant signed an informed consent.
[5] Consent for
publication
The participants provided written informed consent
for the publication of any associated data and accompanying
images.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng LL, Burns MA, Taylor JL, He W,
Halpern EF, McDougal WS and Wu CL: Metabolic characterization of
human prostate cancer with tissue magnetic resonance spectroscopy.
Cancer Res. 65:3030–3034. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brindle JT, Antti H, Holmes E, Tranter G,
Nicholson JK, Bethell HW, Clarke S, Schofield PM, McKilligin E,
Mosedale DE, et al: Rapid and noninvasive diagnosis of the presence
and severity of coronary heart disease using
1H-NMR-based metabonomics. Nat Med. 8:1439–1444. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sreekumar A, Poisson LM, Rajendiran TM,
Khan AP, Cao Q, Yu J, Laxman B, Mehra R, Lonigro RJ, Li Y, et al:
Metabolomic profiles delineate potential role for sarcosine in
prostate cancer progression. Nature. 457:910–914. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
El-Bacha T, Struchiner CJ, Cordeiro MT,
Almeida FC, Marques ET Jr and Da Poian AT: 1H nuclear
magnetic resonance metabolomics of plasma unveils liver dysfunction
in dengue patients. J Virol. 90:7429–7443. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shang X, Zhong X and Tian X: Metabolomics
of papillary thyroid carcinoma tissues: potential biomarkers for
diagnosis and promising targets for therapy. Tumour Biol.
37:11163–11175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen M, Shen M, Li Y, Liu C, Zhou K, Hu W,
Xu B, Xia Y and Tang W: GC-MS-based metabolomic analysis of human
papillary thyroid carcinoma tissue. Int J Mol Med. 36:1607–1614.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan Z and Raftery D: Comparing and
combining NMR spectroscopy and mass spectrometry in metabolomics.
Anal Bioanal Chem. 387:525–527. 2007. View Article : Google Scholar
|
|
9
|
Jordan KW, Adkins CB, Cheng LL and Faquin
WC: Application of magnetic-resonance-spectroscopy-based
metabolomics to the fine-needle aspiration diagnosis of papillary
thyroid carcinoma. Acta Cytol. 55:584–589. 2011. View Article : Google Scholar
|
|
10
|
Miccoli P, Torregrossa L, Shintu L,
Magalhaes A, Chandran J, Tintaru A, Ugolini C, Minuto MN, Miccoli
M, Basolo F, et al: Metabolomics approach to thyroid nodules: a
high-resolution magic-angle spinning nuclear magnetic
resonance-based study. Surgery. 152:1118–1124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deja S, Dawiskiba T, Balcerzak W,
Orczyk-Pawiłowicz M, Głód M, Pawełka D and Młynarz P: Follicular
adenomas exhibit a unique metabolic profile. 1H NMR
studies of thyroid lesions. PLoS One. 8:e846372013. View Article : Google Scholar
|
|
12
|
DeLellis RA, Lloyd RV, Heitz PU and Eng C:
Organization Classification of Tumours Pathology and Genetics of
Tumours of Endocrine Organs. 8. 3rd edition. IARC Press; Lyon:
2004
|
|
13
|
Jordan KW, Nordenstam J, Lauwers GY,
Rothenberger DA, Alavi K, Garwood M and Cheng LL: Metabolomic
characterization of human rectal adenocarcinoma with intact tissue
magnetic resonance spectroscopy. Dis Colon Rectum. 52:520–525.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Du J, Yuan Z, Ma Z, Song J, Xie X and Chen
Y: KEGG-PATH: Kyoto Encyclopedia of Genes and Genomes-based pathway
analysis using a path analysis model. Mol Biosyst. 10:2441–2447.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wishart DS, Jewison T, Guo AC, Wilson M,
Knox C, Liu Y, Djoumbou Y, Mandal R, Aziat F, Dong E, et al: HMDB
3.0 - the Human Metabolome Database in 2013. Nucleic Acids Res.
41:D801–D807. 2013. View Article : Google Scholar
|
|
16
|
Xia J, Sinelnikov IV, Han B and Wishart
DS: Metabo-Analyst 3.0 - making metabolomics more meaningful.
Nucleic Acids Res. 43:W251–W257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Frolkis A, Knox C, Lim E, Jewison T, Law
V, Hau DD, Liu P, Gautam B, Ly S, Guo AC, et al: SMPDB: The Small
Molecule Pathway Database. Nucleic Acids Res. 38(Suppl 1):
D480–D487. 2010. View Article : Google Scholar :
|
|
18
|
Anthony ML, Rose VS, Nicholson JK and
Lindon JC: Classification of toxin-induced changes in 1H
NMR spectra of urine using an artificial neural network. J Pharm
Biomed Anal. 13:205–211. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lamour SD, Straschil U, Saric J and Delves
MJ: Changes in metabolic phenotypes of Plasmodium falciparum in
vitro cultures during gametocyte development. Malar J. 13:468–478.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bahety P, Tan YM, Hong Y, Zhang L, Chan EC
and Ee PL: Metabotyping of docosahexaenoic acid - treated
Alzheimer's disease cell model. PLoS One. 9:e901232014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Z, Klipfell E, Bennett BJ, Koeth R,
Levison BS, Dugar B, Feldstein AE, Britt EB, Fu X, Chung YM, et al:
Gut flora metabolism of phosphatidylcholine promotes cardiovascular
disease. Nature. 472:57–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Dai Y, Wen J, Zhang W, Grenz A,
Sun H, Tao L, Lu G, Alexander DC, Milburn MV, et al: Detrimental
effects of adenosine signaling in sickle cell disease. Nat Med.
17:79–86. 2011. View Article : Google Scholar
|
|
23
|
Wang TJ, Larson MG, Vasan RS, Cheng S,
Rhee EP, McCabe E, Lewis GD, Fox CS, Jacques PF, Fernandez C, et
al: Metabolite profiles and the risk of developing diabetes. Nat
Med. 17:448–453. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen M, Zhou K, Chen X, Qiao S, Hu Y, Xu
B, Xu B, Han X, Tang R, Mao Z, et al: Metabolomic analysis reveals
metabolic changes caused by bisphenol A in rats. Toxicol Sci.
138:256–267. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tessem MB, Selnaes KM, Sjursen W, Tranø G,
Giskeødegård GF, Bathen TF, Gribbestad IS and Hofsli E:
Discrimination of patients with microsatellite instability colon
cancer using 1H HR MAS MR spectroscopy and chemometric
analysis. J Proteome Res. 9:3664–3670. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tessem MB, Swanson MG, Keshari KR, Albers
MJ, Joun D, Tabatabai ZL, Simko JP, Shinohara K, Nelson SJ,
Vigneron DB, et al: Evaluation of lactate and alanine as metabolic
biomarkers of prostate cancer using 1H HR-MAS
spectroscopy of biopsy tissues. Magn Reson Med. 60:510–516. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Koppenol WH, Bounds PL and Dang CV: Otto
Warburg's contributions to current concepts of cancer metabolism.
Nat Rev Cancer. 11:325–337. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cai Z, Zhao JS, Li JJ, Peng DN, Wang XY,
Chen TL, Qiu YP, Chen PP, Li WJ, Xu LY, et al: A combined
proteomics and metabolomics profiling of gastric cardia cancer
reveals characteristic dysregulations in glucose metabolism. Mol
Cell Proteomics. 9:2617–2628. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Newgard CB, An J, Bain JR, Muehlbauer MJ,
Stevens RD, Lien LF, Haqq AM, Shah SH, Arlotto M, Slentz CA, et al:
A branched-chain amino acid-related metabolic signature that
differentiates obese and lean humans and contributes to insulin
resistance. Cell Metab. 9:311–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng SP, Chi CW, Tzen CY, Yang TL, Lee
JJ, Liu TP and Liu CL: Clinicopathologic significance of leptin and
leptin receptor expressions in papillary thyroid carcinoma.
Surgery. 147:847–853. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nishio Y, Kakizoe T, Ohtani M, Sato S,
Sugimura T and Fukushima S: L-isoleucine and L-leucine: tumor
promoters of bladder cancer in rats. Science. 231:843–845. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jain M, Nilsson R, Sharma S, Madhusudhan
N, Kitami T, Souza AL, Kafri R, Kirschner MW, Clish CB and Mootha
VK: Metabolite profiling identifies a key role for glycine in rapid
cancer cell proliferation. Science. 336:1040–1044. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Srivastava S, Roy R, Singh S, Kumar P,
Dalela D, Sankhwar SN, Goel A and Sonkar AA: Taurine - a possible
fingerprint biomarker in non-muscle invasive bladder cancer: a
pilot study by 1H NMR spectroscopy. Cancer Biomark.
6:11–20. 2010. View Article : Google Scholar
|
|
34
|
Katz-Brull R, Seger D, Rivenson-Segal D,
Rushkin E and Degani H: Metabolic markers of breast cancer:
enhanced choline metabolism and reduced choline-ether-phospholipid
synthesis. Cancer Res. 62:1966–1970. 2002.PubMed/NCBI
|
|
35
|
Venkataraman GM, Yatin M, Marcinek R and
Ain KB: Restoration of iodide uptake in dedifferentiated thyroid
carcinoma: relationship to human Na+/I−
symporter gene methylation status. J Clin Endocrinol Metab.
84:2449–2457. 1999.PubMed/NCBI
|
|
36
|
Halton JM, Nazir DJ, McQueen MJ and Barr
RD: Blood lipid profiles in children with acute lymphoblastic
leukemia. Cancer. 83:379–384. 1998. View Article : Google Scholar : PubMed/NCBI
|