1. Introduction
The Hippo signal pathway was first discovered by
mosaic genetic screens on Drosophila for mutants exhibiting
tissue overgrowth in the 90s (1,2).
This signal pathway plays pivotal roles in organ growth control,
stem cell function, regeneration and tumor suppression (3,4).
Wart (Wts) was identified as the first component of the
Drosophila Hippo pathway, which encodes a protein kinase of
the nuclear Dbf-2-related (NDR) family and plays an important role
in controlling cell growth and shape as well as proliferation
(5). Subsequent studies uncovered
several other Hippo pathway components, including scaffold protein
Salvador (Sav), Hippo (Hpo) and Mats in the following years
(6). These four proteins formed
the center part of the Drosophila Hippo pathway, in which
Hpo, in association with Sav, phosphorylate and activate Wts and
the adopter protein Mats. Therefore, the Hippo pathway is also
known as the Salvador-Warts-Hippo pathway (7). Wts/Mats is a protein kinase complex,
thus, it functions through its substrates. By protein interaction
screens using Wts as bait, Huang et al identified Yorkie
(Yki) as the major substrate of the Wts/Mats complex in the
Drosophila (8).
Inactivation of Yki through phosphorylation or
mutation caused tissue atrophy, while over-expression of Yki caused
massive tissue overgrowth, phenocopying loss of Hpo, Sav or Wts
function. Functionally, unphosphorylated Yki can translocate into
the nucleus and drive growth-promoting related gene expression,
such as cell cycle regulator cyclin E, cell-death inhibitor Diap1,
and bantam microRNA (9,10). Thus the localization and
phosphorylation of Yki are often taken as a measure of the activity
of the Hippo pathway (11). The
Hippo pathway is deregulated with a high frequency in many human
cancers, which confirmed the Hippo signaling pathway is closely
related to the development of cancers (12). Therefore, the discovery of Hippo
pathway has generated excitement in both basic and clinical
research because targeting the Hippo pathway provide us with new
insight into development of anticancer drugs (13).
Herein, we provide an overview of the Hippo pathway,
with an emphasis on the mammalian system, and summarize the current
understanding of the biological functions of YAP and TAZ, and
discuss the opportunities and challenges for therapeutic
intervention.
2. The Hippo signaling pathway in
mammals
The Hippo signaling pathway is an evolutionarily
conserved pathway; hence, most components of the Drosophila
Hippo pathway have mammalian homologues (Table I). For instance, Mst1/2, Sav1,
Lats1/2, Mob1 in mammalian are homologues of Hpo, Sav, Wts and
Mats, respectively; both YAP and TAZ are homologues of Yki. Similar
to the Drosophila Hippo pathway, the center part of the
mammalian Hippo pathway is also a protein kinase cascade. Once the
upstream activation signals are received, the downstream Mst1/2
kinase phosphorylate and activate the Lats1/2 kinases and MOB1
cofactor with the help of SAV1. Then the activated Lats1/2 kinases
phosphorylate and inactivate YAP/TAZ transcription co-activators by
promoting their cytoplasmic localization and degradation.
Conversely, unphosphorylated YAP/TAZ will translocated and
accumulated in the nucleus and form complexes with transcription
factors, consequently inducing the expression of targeted genes,
such as connective tissue growth factor (CTGF), cysteine-rich 61
(Cyr61), and fibroblast growth factor (FGF1) (14) (Fig.
1).
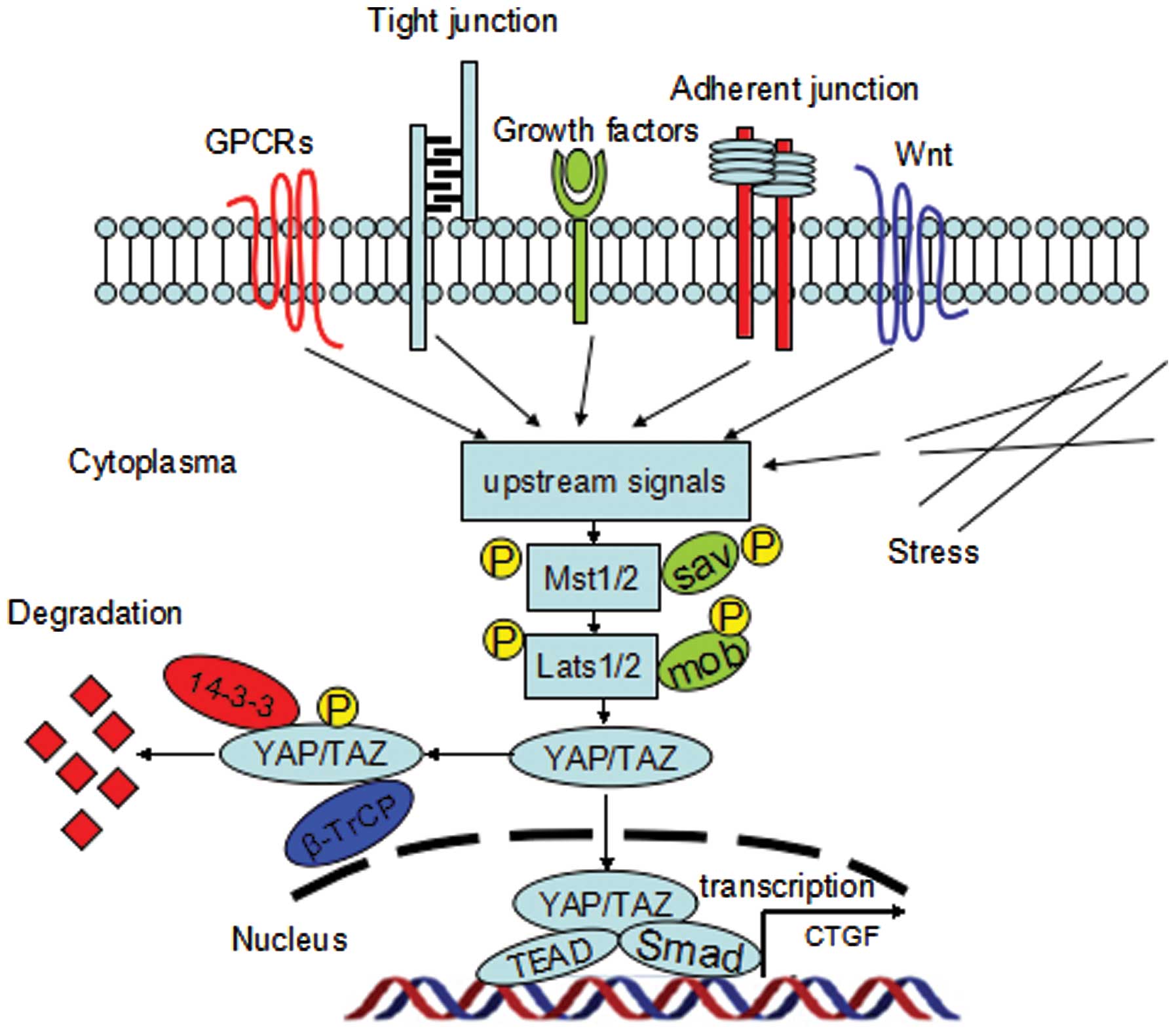 | Figure 1The mammalian Hippo signaling
pathway. Upstream signals include the GPCR signaling, Wnt
signaling, mechanical stress, tight junctions, adherent junctions,
as well as some soluble growth factors. These signals regulate the
phosphorylation of Mst1/2 and Lats1/2 kinases. Lats1/2
phosphorylates YAP/TAZ on HXRXXS motifs; and phosphorylation of
YAP/TAZ creates a 14-3-3 binding site, which promotes cytoplasmic
localization of YAP/TAZ, resulting in YAP/TAZ cytoplasmic retention
and degradation. When YAP/TAZ are dephosphorylated, they enter the
nucleus, and act as transcription coactivators by forming complexes
with various transcriptional factors such as TEAD, SMAD, to
coordinate pro-proliferating and anti-apoptotic programs. |
 | Table IHippo signaling pathway components in
mammalian and homologues in Drosophila. |
Table I
Hippo signaling pathway components in
mammalian and homologues in Drosophila.
| Mammalian
proteins | Drosophila
homologues | Conserved domains
and motifs | Main functions |
|---|
| Upstream
modulators |
| Fat4 | Fat | EGF-like repeats,
laminin A-G domain | Adhesion molecule
and/or signaling receptor |
| Dchs1/2 | dachsous | Cadherin repeated
domain | Cell-adhesion
protein |
| Fjx1 | Fjx1 | Golgi Ser/thr
kinase | Legs, wings and
eyes development |
| Ck1 | Dco | ser/thr kinase
domain | Protein kinase |
| ZDHHC9/14/18 | App | DHHC domain | DHHC
palmitoyltransferase |
| Patj | patj | Ribosomal protein
L27, PDZ domain | Adaptor
protein |
| FRMD6, Ex1 | ex | FERM domain | Adaptor
protein |
| PTPN14 | pez | FERM domain | Phosphates |
| NF2 | mer | FERM domain | Adaptor
protein |
| Kibra | kibra | WW domains, C2
domain | Adaptor
protein |
| Lgl1,2 | Lgl | LLGL2 domain | WD40 scaffold
protein |
| Crb1-3 | crb | EGF domains,
laminin A-G domains, transmembrane domain | Transmembrane
receptor |
| DLG1-4 | Dlg | Three PDZ domains,
SH3 domain, GUK domain | Adaptor
protein |
| SCRIB | Scrib | LRR domains, PDZ
domains | Adaptor
protein |
| Rassf1-6 | rassf | RAS association
domain, SARAH domain | Adaptor
protein |
| Ajuba | jub | LIM domains | Adaptor
protein |
| Mpp5 | sdt | Ribosomal protein
L27, PDZ domain, SH3 domain, GUK domain | Adaptor
protein |
| Amot, AmotL | - | CC domain,
PDZ-BM | Adaptor
protein |
| TAO1-3 | Tao | Kinase domain | Serine/threonine
kinase |
| E-cadherin | E-cadherin | cadherin
domain | Transmembrane
receptor |
| α-catenin | α-catenin | VH1-VH3
domains | Adaptor
protein |
| Core
components |
| Mst1/2 | Hpo | Kinase domain,
SARAH domain | Serine/threonine
kinase |
| Lats1/2 | Wts | Kinase domain | Serine/threonine
kinase |
| YAP/TAZ | Yki | WW domains, 14-3-3
BM, TEAD-BM PDZ-BM | Transcriptional
co-activator |
| Sav1 | Sav | FERM domain, WW
domains, SARAH domain | Adaptor
protein |
| Mob1/2 | Mats | MOB domain | Adaptor
protein |
| Downstream
mediators |
| TEAD1-4 | Sd | TEAD DNA-binding
domain, vestigial binding domain | Transcription
factor |
| Smads | MAD | MH1 domain | Transcription
factor |
| TSHZ1-3 | Tsh | C2H2 type
zinc-finger domain | Zn-finger
transcription factor |
| Wbp2 | Wbp2 | WW domain, GRAM
domain | Cofactor |
The upstream signals that regulate the Hippo pathway
have not been well delineated, but it consist of a large network of
proteins: including a variety of kinases, such as Src, protein
kinase A (PKA) (15); some soluble
factors, such as amphiregulin (ARGE) (14); cell adhesion and cell junction
proteins, such as Echinoid, cadherin-catenin complex (16,17);
cell polarity proteins, such as the crumbs complex, scribble
(18); other signaling pathways,
such as the G-protein-coupled receptor (GPCR) signaling, Wnt and
PI3K pathways (19–21); and the state of the actin
cytoskeleton (22,23). However, how these proteins impact
on YAP/TAZ is not entirely clear. According to the existing
experimental conclusions, in addition to regulating YAP/TAZ
phosphorylation; these upstream factors also regulate YAP/TAZ
nuclear localization through physical interaction.
3. Similarities and differences between YAP
and TAZ in structure and function
YAP was first identified as a proline-rich
phosphoprotein bound to the SH3 domain of Yes and Src protein
tyrosine kinases (24). The human
YAP gene, located at 11q13, encodes a 65 kDa protein. Eight
isoforms of YAP protein have been identified in humans to date,
which are differed by either an alteration in the activation domain
or a loss of WW domain (25,26).
YAP mRNA is not only ubiquitously expressed in a broad range of
tissues; but also expressed in the whole developmental process
(27). YAP gene is amplified in
various human cancers. TAZ, also known as WWTR1 (WW-domain
containing transcriptional regulator 1), was first identified in
2000 as a 14-3-3 binding protein. The human TAZ gene, located at
3q23-q24, encodes a 43 kDa protein. Similarly, TAZ is expressed in
various tissues, except thymus and peripheral blood leukocytes and
amplified in various human cancers (28).
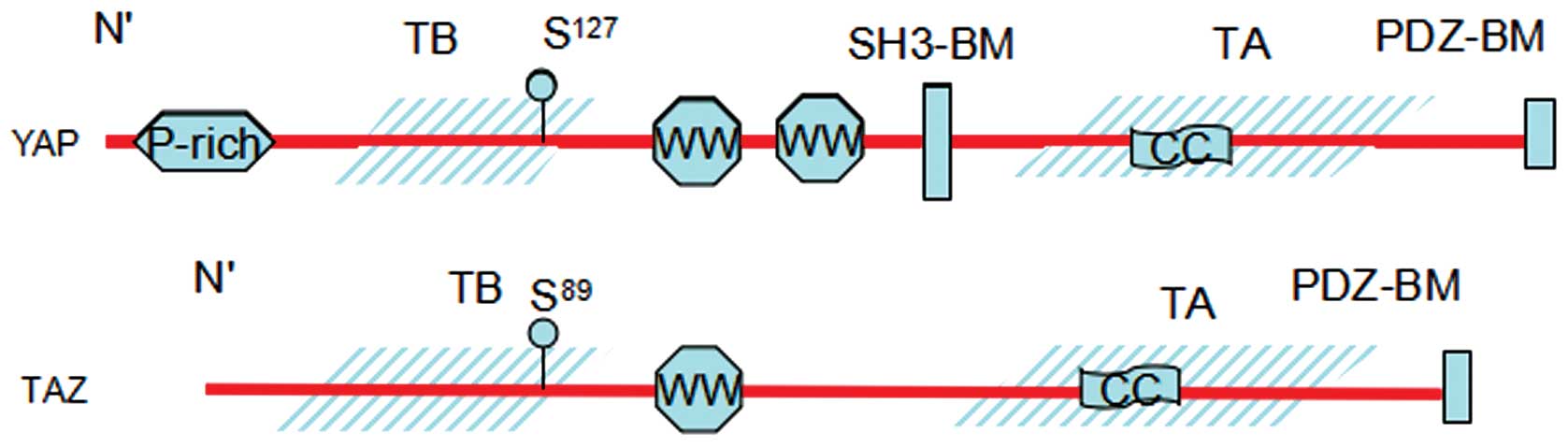
Structurally, YAP and TAZ share nearly half of the
overall amino acid sequence and have very similar topology. YAP
protein consists of 488 amino acids, has a TEAD-binding region
(TB), two WW domains, which consist of two conserved tryptophan (W)
residues separated by 20–23 amino acids, an SH3-binding motif, a
coiled-coil domain, a transcription activation domain, an
N-terminal proline-rich domain, and a C-terminal PDZ-binding motif.
TAZ protein consists of 400 amino acids, has a similar domain
organization with YAP, although it lacks the second WW domain, the
SH3-binding motif, and the proline-rich domain (Fig. 2).
Biochemically, the WW domains of the YAP and TAZ
have been shown to interact with PPXY motifs (proline/proline/any
amino acid/tyrosine) of some transcriptional factors. The TB domain
recognizes the TEAD family of transcription factors and activates
target gene expression, whereas the 14–3–3 binding motif is crucial
to the regulation of YAP and TAZ. The PDZ-binding motif is required
for binding with PDZ domain, which are found in many transmembrane
or cytoskeleton associated proteins. The PDZ-binding domains appear
to direct YAP and TAZ localization (29,30).
At the functional level, both of the YAP and TAZ are
serving as transcriptional co-activator and share some
transcription factor partners, such as TEAD and Runx. Both of them
promote cell proliferation, increases cell migration and invasion.
However, both of them have their unique target transcription
factors, such as ErbB4, p73 for YAP and PPARγ, Pax3, TBX5, TTF-1
for TAZ, which may contribute to the differential functions of YAP
to TAZ. Moreover, gene knockout mouse models demonstrated the
different physiological functions between YAP and TAZ: Knockout of
the YAP gene in mice leads to early developmental arrest,
suggesting an essential role in development. However, TAZ knockout
mice are viable, although they have defects in the kidneys and
lungs.
4. Transcriptional regulation by YAP/TAZ:
partners and targets
Although YAP and TAZ are the main downstream
effectors of the Hippo pathway in the mammals, YAP and TAZ do not
have DNA binding domains. Alternatively, once they localize to the
nucleus, they interact with transcription factor partners and
function as transcription co-activators. YAP and TAZ share some
transcription factor partners, such as TEAD and Runx proteins.
The mammalian genome contains four highly homologous
members of the TEAD protein family (TEAD1-4), which are the major
targets of the YAP and TAZ in regulation of cell contact
inhibition, epithelial-mesenchymal transition (EMT), oncogenic
transformation and apoptosis inhibition (31). The interaction of the TEAD with the
TB domains of YAP and TAZ can activate downstream gene transcripts,
such as CTGF, and Cyr61. Disrupting the binding of YAP or TAZ to
TEAD by genetic approach will abolish their ability to promote cell
proliferation and tumorigenesis (32–34).
For instance, a mutation of TEAD1 in Y421H leads to the human
genetic disease Sveinsson’s chorioretinal atrophy, which is
characterized by helicoid peripapillary chorioretinal degeneration
or atrophia areata (35). Besides
the TB domains, the WW domains of the YAP and TAZ have been shown
to interact with PPXY motifs of some transcriptional factors. Runx
transcription factors consist of 3 members (Runx1, Runx2 and
Runx3), all have a PPXY motif and are involved in carcinogenesis
and cancer metastasis. Both YAP and TAZ can bind to and potentiate
Runx activity through WW-PPXY interaction (36,37).
In addition to the above binding modes, YAP and TAZ
can regulate expression of some genes through other means. TAZ
increases the transcriptional activity of thyroid transcription
factor-1(TTF-1) on the surfactant protein C promoter by directly
interacting with the N-terminal domain of TTF-1 (38). T-box 5 gene (TBX5) belongs to the
T-box family of transcription factors that plays an essential role
in cardiac and limb development. YAP and TAZ can stimulate TBX5
transcription by interaction with multiple domains including
carboxyl-terminal structure of the TBX5 (39). Through the coiled-coil region, YAP
and TAZ are able to interact with Smad2/3/4 and consequently
activate transforming growth factor-β target gene expression and
maintain human embryonic stem cell self-renewal (40).
However, both YAP and TAZ have their own specific
binding partners, although they are very similar in their
structure. For instance, TAZ inhibits the transcriptional
activation of PPARγ, which is critical for adipocyte
differentiation, through the interaction of WW domain with PPXY
motif. This inhibition prevented mesenchymal stem cells from
differentiating into adipocytes. However, although YAP has WW
domains, it is unable to interact with PPARγ (41). TAZ, but not YAP, interacts with the
PPXY motif of the Kruppel-like factor 5 (KLF5) through the WW
domain and protects KLF5 from WWP1-mediated ubiquitination and
degradation, thereby, promotes breast cell proliferation and
tumorigenesis (42). On the other
hand, YAP also has its specific binding partners, such as p73 and
ErBb4. Strano et al demonstrated the WW domain of YAP
interacts with PPXY motif of the p73, consequently enhancing p73
transcription activity and accelerating programmed cell death
(43). Komuro et al
reported YAP activates ErbB4-dependent transcription by forming the
YAP-ErbB4 complex, which is mediated by the WW domain of YAP and
the PPXY motif of ErbB-4 (44).
5. Mechanisms of YAP/TAZ inhibition by the
Hippo pathway
Phosphorylation of YAP S127 and the
corresponding site of TAZ by Lats1/2 promote 14-3-3 protein binding
resulting in YAP/TAZ cytoplasm retention and functional inhibition,
because they were separated from their nuclear binding partners.
Mutation of YAP S127 or equivalent residues in TAZ
abrogated the 14-3-3 protein interaction thus enhancing their
function as transcriptional co-activators (45).
In addition to promoting YAP/TAZ translocation,
phosphorylation by Lats1/2 also suppress their activities by
inducing proteasomal degradation. The Lats1/2 phosphorylate the
C-terminal phosphodegrons in YAP/TAZ, primed them for subsequent
phosphorylation by casein kinase 1, eventually resulting in the
recruitment of the SCFβ-TrCP ligase, thereby leading to
poly-ubiquitylation and degradation of YAP/TAZ (46). The N-terminal phosphodegron of TAZ
can be phosphorylated by GSK3 via PI3K signaling, resulting in
SCFβ-TrCP E3 ubiquitin ligase-mediated proteolysis of
TAZ (47). Disruption of the
interaction between YAP/TAZ and β-TrCP by mutation will stabilize
TAZ/YAP and promote their function.
YAP/TAZ can also be inhibited in a protein-protein
interaction manner, the PPXY motifs on Lats1/2, AMOT, and PTPN14
have been shown to directly interact with WW domains on YAP/TAZ and
thereby convey inhibitory signals by sequestering YAP/TAZ in the
cytosol or junctional compartments (48,49).
Through its PDZ domain, junctional protein zona occludens-2 (ZO-2)
can directly bind with YAP, consequently promoting YAP
translocation to the nucleus (50,51).
6. YAP/TAZ in human cancers
Numerous studies show that overexpression of YAP/TAZ
can induce EMT, inhibiting apoptosis and increase the number of
cancer stem cells in vitro (52–54).
Knockdown of YAP/TAZ expression reduced cancer cell migration,
invasion and anchorage-independent growth in soft agar, as well as
tumorigenesis in nude mice (54–56).
The protein level of TAZ is correlated with the invasiveness of
breast cancer cells (54). In
transgenic mouse models, over-expression of YAP/TAZ or attenuation
of Hippo signaling is sufficient to promote tumor formation
(45,57,58).
Indeed, the levels and nuclear localization of
YAP/TAZ are elevated in many human cancers, such as carcinomas of
the lung, thyroid, ovarian, colorectal, prostate, pancreas,
esophagus, liver, mammary gland (12,59,60),
and positively correlated with poorly differentiated tumors
(53). Higher YAP/TAZ protein
levels are associated with shorter patient overall survival
(61–63). Xie et al reported TAZ
protein was expressed in 66.8% (121/181) of non-small cell lung
cancer patients (NSCLC) cases and significantly associated with
poorer differentiation and short survival (64). Yue et al reported TAZ
protein expression was positive in 113 out of 146 (77.4%) gastric
cancer samples and especially higher expressed in signet ring cell
carcinoma (65). The studies of
Yuen et al demonstrated that the mRNA level of TAZ is a
prognostic marker in colon cancer progression (61). Wang et al reported YAP was
overexpressed in 66.3% (61/92) specimens of NSCLC and significantly
correlated with p-TNM stage and lymph node metastasis (62). Lam-Himlin et al found that
YAP expression in the cytoplasm and nucleus is significantly
increased in high-grade dysplasia and adenocarcinoma of the
esophagus as well as gastric adenocarcinoma and metastatic gastric
disease (66).
However, some researchers suggest YAP might function
as a tumor suppressor. Knockdown of YAP in breast cell lines
suppressed anoikis, increased migration and invasiveness, enhanced
tumor growth in nude mice, and decreased Taxol responsiveness
(67,68). Overexpression of YAP can induce p19
cell apoptosis by increase p73 alpha transcriptional activity
(39). YAP is also a critical
component of c-Jun-mediated induction of apoptosis (43,69).
In contrast, TAZ have not been found to have tumor suppressor
function. These research studies suggest the possibility that YAP
may have dual roles, as tumor promoter or tumor suppressor, which
may depend on the cell context.
7. YAP/TAZ as potent therapeutic targets in
cancer
Considerable number of studies show that the Hippo
pathway plays a key role in tumorigenesis. Downregulation of the
Hippo pathway activity not only promotes cancer cells growth, but
also makes cancer cells resistant to some chemotherapy drugs
(70). These founding raised the
possibility that therapeutic intervention of Hippo signaling may
improve current treatment strategies. We focus on small molecules
that are designed to target Hippo signaling itself by manipulating
core components of the Hippo pathway (Table II).
| Table IISmall-molecule modulators of the
Hippo pathway. |
Table II
Small-molecule modulators of the
Hippo pathway.
| Compounds | Targets and
effects | Refs. |
|---|
| 17-mer | YAP-like peptides,
abolish YAP-TEAD interaction | (73) |
| Verteporfin
(VP) | Binding to YAP,
thus inhibit YAP-TEAD interaction | (71) |
| 9E1 | Mst1 inhibitor,
inhibit Mst1 kinase activity, thus impair YAP/TAZ degradation | (74) |
| LPA, S1P, Thrombin,
Epinephrin, Glucagon | Targeting GPCR
signaling pathway, act through GPCRs to inhibit the Lats1/2
kinases, thereby activating YAP/TAZ | (20,82,83) |
| Butamine | G protein-coupled
β-adrenergic receptor agonist, prevent YAP nuclear
accumulation | (86) |
| Dasatinib | SRC family
inhibitor, inhibits YES1, resulted in inactivation of the
YAP1-β-catenin-TBX5 complex, and thus impaired the proliferation of
β-catenin active cells | (75,76) |
| Atrunculin A/B,
Cytochalasin D Blebbistatin | F-actin
destabilizers, inhibit YAP nuclear localization as well as YAP and
TEAD activity in various cell lines; promoting YAP/TAZ nuclear
translocation | (77–80) |
| Y27632 | RHO-ROCK signaling
inhibitor, prevent YAP from phosphorylating by Lats1/2, thus
promote YAP nuclear accumulation | (81) |
| XAV939, G007-LK,
G244-LM | Tankyrase
inhibitors, restore the activity of Axin by preventing
PAR-dependent Axin degradation, led to YAP/TAZ degradation via the
β-catenin destruction complex | (88,89) |
| Statins
Bisphosphonates | SREBP/mevalonate
pathway inhibitors, inhibit YAP/TAZ nuclear localization and
activation through metabolic control | (90) |
Targeting YAP/TAZ-TEAD interaction or
upstream kinases
In many cases, the tumorigenesis potential of YAP
and TAZ require association to TEAD proteins. Blocking the
formation of the YAP/TAZ-TEAD complex will abolish the transforming
ability of YAP and TAZ in vitro (33,34);
by screening of a small-molecule library, several porphyrin
compounds have been found out to bind to YAP, and subsequently
inhibit the interaction of YAP and TEAD. One of these porphyrin
compounds, verteporfin (VP), is already used for photodynamic
therapy of macular degeneration in the clinic; repeated
administration of VP was effective in delaying tumor progression
and suppressing liver overgrowth in a liver cancer mouse model,
without overt adverse effects in other organs (71). Vestigial-like protein 4 (VGLL4) is
a natural antagonist of YAP, because its TDU region has a structure
similar to TB domains of YAP, thus can competitively inhibit the
inter action of YAP-TEAD. These findings led to the generation of
peptide-based YAP inhibitors (72,73).
In addition, small-molecule inhibitors of the upstream kinases,
such as Mst1 inhibitor 9E1, would upregulate YAP and TAZ function
(74). Src family kinase
inhibitor, Dasatinib, inhibits kinase YES1, resulting in
inactivation of the YAP1-β-catenin-TBX5 complex, and thus impaired
the proliferation of β-catenin active cells (75,76).
Targeting F-actin
F-actin can inhibit Hippo pathway activity, thereby
promoting YAP/TAZ nuclear translocation, which leads to
upregulation of proliferation and survival genes (77–79).
Therefore, it is reasonable to speculate that drugs targeting
F-actin may be effective in control of tumor growth. The F-actin
destabilizers, such as cytochalasin D, latrunculin A/B, together
with RHO kinase inhibitor Y27632 and the non-muscle myosin II
inhibitor blebbistatin all cause nuclear export of YAP and TAZ
(80,81). However, it is worth noting that the
actin cytoskeleton is important for many basic cellular functions.
Therefore, choosing a proper dosage of F-actin inhibitors would
improve the pharmacotherapy efficacy and reduce the toxic side
effects.
Targeting GPCR signaling pathway
GPCR represent one of the largest gene families in
the human genome and regulate a wide array of physiological
functions. Yu et al reported GPCR signaling can either
activate or inhibit the activities of YAP/TAZ depending on the
coupled G protein (20).
Serum-borne lysophosphatidic acid (LPA), sphingosine 1-phosphophate
(S1P), thrombin and protease-activated receptor agonist peptides
act through G12/13 coupled receptors to inhibit the
Lats1/2 kinases, thereby activating YAP/TAZ (82); while adrenaline, glucagon and
dihydrexidine activate Gαs coupled receptors, result in enhanced
YAP phosphorylation and inactivation (83). Therefore, development of antibodies
or small-molecule agonists mimicking LPA or S1P stimulation may be
an effective therapeutic option. Many of these drugs have been
developed and, such as the S1P-blocking antibody Sphingomab,
Phosphatase-resistant LPA analogs were evaluated in clinic trails
(84,85). By cell-based method of screening,
another G protein-coupled β-adrenergic receptor agonist,
dobutamine, was identified and shown to be effective in preventing
YAP nuclear accumulation and YAP-mediated transcriptional
activation. Of note, the effect of dobutamine is independent of the
Hippo pathway, which means it is possible to develop lower side
effect Hippo-independent YAP inhibitors (20,86).
Targeting Wnt/β-catenin signaling
pathway
It was reported that YAP/TAZ serve as downstream
elements of the Wnt/β-catenin signaling pathway with independent of
the Hippo signal pathway. Once Wnt signaling was turned on,
β-catenin detached from the destruction complex, formed by APC,
Axin and GSK3, resulting in inhibition of YAP/TAZ degradation then
leading to YAP/TAZ activation (19,87).
Therefore, targeting the Wnt pathway by restoring the integrity of
the destruction complex would effectively inhibit YAP/TAZ. Some
tankyrase inhibitors, such as XAV939, G007-LK and G244-LM can
restore the integrity of the destruction complex by increasing the
Axin activity, leading to YAP/TAZ degradation via the β-catenin
destruction complex (88,89).
Targeting metabolic cues
It was recently reported that YAP/TAZ activity is
controlled metabolically via the SREBP/mevalonate pathway.
Functionally, mevalonate is a precursor for geranylgeranyl
diphosphate (GGPP) that, in turn, promotes Rho GTPase membrane
localization and activity, thus leading to YAP/TAZ nuclear
localization and activation. Inhibition of this pathway by means of
statins, bisphosphonates or geranylgeranyl transferase inhibitors
attenuates YAP/TAZ biological activities (90). The significance of this finding is
that it opened up new ideas for designing anti-YAP/TAZ drugs.
8. Perspectives
The main reasons why the Hippo signaling pathway
gained so much attention since it was delineated are the crucial
roles in organ size control, cell differentiation, as well as
tumorigenesis. The Hippo pathway activity is associated with cancer
cell’s proliferative potential, multi-drug resistance, EMT and
metastasis. In the past dozen years, our understanding of the Hippo
signaling pathway, in both Drosophila and mammalian, has
largely increased; the kinase cascade and the main downstream
effectors of this pathway were basically defined. However, there
are still many questions to be answered: i) although a wide range
of upstream signals have been identified, the details mechanism by
which these upstream regulators integrated to impact on the core
components is not yet fully understood. ii) Some components also
interplay with other pathways; the in-depth mechanisms need to be
uncovered. iii) YAP and TAZ have similar structure and both serve
as downstream effectors in the Hippo pathway, but they also have
distinct functions. Is there some internal relationship between YAP
and TAZ? Are there any other transcription factors and target genes
mediating YAP/TAZ function? iv) Why does the Hippo pathway vary in
terms of cell context? Are there any predictive biomarkers that
enable us to select appropriate patients for anti-YAP/TAZ drugs
treatment? Answers to these questions would advance our
understanding of the Hippo pathway, as well as provide us valuable
information for new drug development.
There are numerous potential drug targets identified
to modulate the Hippo pathway and some new drugs are under clinical
trials. However, a batch of new problems have appeared: most of the
targets lack of specificity, such as upstream regulators GPCR,
which have broad physiological functions in normal cells. General
interference of these targets may cause serious side effects.
Directly targeting the core components, such as YAP and TAZ, would
improve specificity, but YAP and TAZ also play important roles in
organ size control and normal tissue regeneration. It means that
long-term whole-body inhibition of either of these proteins may
results in serious side effects. It would be ideal to develop
effective means to manipulate the Hippo pathway and YAP/TAZ
activity in both a tissue specific and transient manner.
Undoubtedly, the Hippo pathway and YAP/TAZ may represent valuable
targets for cancer therapy, although further research is necessary
to definite the upstream regulators, downstream targets, as well as
the regulation mechanisms of this pathway.
References
|
1
|
Harvey K and Tapon N: The
Salvador-Warts-Hippo pathway - an emerging tumour-suppressor
network. Nat Rev Cancer. 7:182–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Watson KL, Justice RW and Bryant PJ:
Drosophila in cancer research: The first fifty tumor suppressor
genes. J Cell Sci (Suppl). 18:19–33. 1994. View Article : Google Scholar
|
|
3
|
Halder G and Johnson RL: Hippo signaling:
Growth control and beyond. Development. 138:9–22. 2011. View Article : Google Scholar :
|
|
4
|
Mo JS, Park HW and Guan KL: The Hippo
signaling pathway in stem cell biology and cancer. EMBO Rep.
15:642–656. 2014.PubMed/NCBI
|
|
5
|
Justice RW, Zilian O, Woods DF, Noll M and
Bryant PJ: The Drosophila tumor suppressor gene warts encodes a
homolog of human myotonic dystrophy kinase and is required for the
control of cell shape and proliferation. Genes Dev. 9:534–546.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu S, Huang J, Dong J and Pan D: Hippo
encodes a Ste-20 family protein kinase that restricts cell
proliferation and promotes apoptosis in conjunction with salvador
and warts. Cell. 114:445–456. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Udan RS, Kango-Singh M, Nolo R, Tao C and
Halder G: Hippo promotes proliferation arrest and apoptosis in the
Salvador/Warts pathway. Nat Cell Biol. 5:914–920. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang J, Wu S, Barrera J, Matthews K and
Pan D: The Hippo signaling pathway coordinately regulates cell
proliferation and apoptosis by inactivating Yorkie, the Drosophila
Homolog of YAP. Cell. 122:421–434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thompson BJ and Cohen SM: The Hippo
pathway regulates the bantam microRNA to control cell proliferation
and apoptosis in Drosophila. Cell. 126:767–774. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koontz LM, Liu-Chittenden Y, Yin F, Zheng
Y, Yu J, Huang B, Chen Q, Wu S and Pan D: The Hippo effector Yorkie
controls normal tissue growth by antagonizing scalloped-mediated
default repression. Dev Cell. 25:388–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hong W and Guan KL: The YAP and TAZ
transcription co-activators: Key downstream effectors of the
mammalian Hippo pathway. Semin Cell Dev Biol. 23:785–793. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harvey KF, Zhang X and Thomas DM: The
Hippo pathway and human cancer. Nat Rev Cancer. 13:246–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stanger BZ: Quit your YAPing: A new target
for cancer therapy. Genes Dev. 26:1263–1267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Chen Z, Wang Y, et al: WWTR1
promotes cell proliferation and inhibits apoptosis through cyclin A
and CTGF regulation in non-small cell lung cancer. Tumour Biol.
35:463–468. 2014. View Article : Google Scholar
|
|
15
|
Yu FX, Zhang Y, Park HW, Jewell JL, Chen
Q, Deng Y, Pan D, Taylor SS, Lai ZC and Guan KL: Protein kinase A
activates the Hippo pathway to modulate cell proliferation and
differentiation. Genes Dev. 27:1223–1232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim NG, Koh E, Chen X and Gumbiner BM:
E-cadherin mediates contact inhibition of proliferation through
Hippo signaling-pathway components. Proc Natl Acad Sci USA.
108:11930–11935. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yue T, Tian A and Jiang J: The cell
adhesion molecule echinoid functions as a tumor suppressor and
upstream regulator of the Hippo signaling pathway. Dev Cell.
22:255–267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Verghese S, Waghmare I, Kwon H, Hanes K
and Kango-Singh M: Scribble acts in the Drosophila fat-hippo
pathway to regulate warts activity. PLoS One. 7:e471732012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Konsavage WM Jr and Yochum GS:
Intersection of Hippo/YAP and Wnt/β-catenin signaling pathways.
Acta Biochim Biophys Sin (Shanghai). 45:71–79. 2013. View Article : Google Scholar
|
|
20
|
Yu FX, Zhao B, Panupinthu N, et al:
Regulation of the Hippo-YAP pathway by G-protein-coupled receptor
signaling. Cell. 150:780–791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tumaneng K, Schlegelmilch K, Russell RC,
Yimlamai D, Basnet H, Mahadevan N, Fitamant J, Bardeesy N, Camargo
FD and Guan KL: YAP mediates crosstalk between the Hippo and
PI(3)K-TOR pathways by suppressing PTEN via miR-29. Nat Cell Biol.
14:1322–1329. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dai X, She P, Chi F, et al:
Phosphorylation of angiomotin by Lats1/2 kinases inhibits F-actin
binding, cell migration, and angiogenesis. J Biol Chem.
288:34041–34051. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong W: Angiomotin’g YAP into the nucleus
for cell proliferation and cancer development. Sci Signal.
6:pe272013.
|
|
24
|
Sudol M: Yes-associated protein (YAP65) is
a proline-rich phosphoprotein that binds to the SH3 domain of the
Yes proto-oncogene product. Oncogene. 9:2145–2152. 1994.PubMed/NCBI
|
|
25
|
Sudol M, Bork P, Einbond A, Kastury K,
Druck T, Negrini M, Huebner K and Lehman D: Characterization of the
mammalian YAP (Yes-associated protein) gene and its role in
defining a novel protein module, the WW domain. J Biol Chem.
270:14733–14741. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gaffney CJ, Oka T, Mazack V, et al:
Identification, basic characterization and evolutionary analysis of
differentially spliced mRNA isoforms of human YAP1 gene. Gene.
509:215–222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Morin-Kensicki EM, Boone BN, Howell M,
Stonebraker JR, Teed J, Alb JG, Magnuson TR, O’Neal W and Milgram
SL: Defects in yolk sac vasculogenesis, chorioallantoic fusion, and
embryonic axis elongation in mice with targeted disruption of
Yap65. Mol Cell Biol. 26:77–87. 2006. View Article : Google Scholar :
|
|
28
|
Kanai F, Marignani PA, Sarbassova D, et
al: TAZ: A novel transcriptional co-activator regulated by
interactions with 14-3-3 and PDZ domain proteins. EMBO J.
19:6778–6791. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oka T and Sudol M: Nuclear localization
and pro-apoptotic signaling of YAP2 require intact PDZ-binding
motif. Genes Cells. 14:607–615. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Remue E, Meerschaert K, Oka T, Boucherie
C, Vandekerckhove J, Sudol M and Gettemans J: TAZ interacts with
zonula occludens-1 and -2 proteins in a PDZ-1 dependent manner.
FEBS Lett. 584:4175–4180. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sawada A, Kiyonari H, Ukita K, Nishioka N,
Imuta Y and Sasaki H: Redundant roles of Tead1 and Tead2 in
notochord development and the regulation of cell proliferation and
survival. Mol Cell Biol. 28:3177–3189. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lamar JM, Stern P, Liu H, Schindler JW,
Jiang ZG and Hynes RO: The Hippo pathway target, YAP, promotes
metastasis through its TEAD-interaction domain. Proc Natl Acad Sci
USA. 109:E2441–E2450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang H, Liu CY, Zha ZY, Zhao B, Yao J,
Zhao S, Xiong Y, Lei QY and Guan KL: TEAD transcription factors
mediate the function of TAZ in cell growth and
epithelial-mesenchymal transition. J Biol Chem. 284:13355–13362.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao B, Ye X, Yu J, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fossdal R, Jonasson F, Kristjansdottir GT,
Kong A, Stefansson H, Gosh S, Gulcher JR and Stefansson K: A novel
TEAD1 mutation is the causative allele in Sveinsson’s chorioretinal
atrophy (helicoid peripapillary chorioretinal degeneration). Hum
Mol Genet. 13:975–981. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yagi R, Chen LF, Shigesada K, Murakami Y
and Ito Y: A WW domain-containing yes-associated protein (YAP) is a
novel transcriptional co-activator. EMBO J. 18:2551–2562. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zaidi SK, Sullivan AJ, Medina R, Ito Y,
van Wijnen AJ, Stein JL, Lian JB and Stein GS: Tyrosine
phosphorylation controls Runx2-mediated subnuclear targeting of YAP
to repress transcription. EMBO J. 23:790–799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Park KS, Whitsett JA, Di Palma T, Hong JH,
Yaffe MB and Zannini M: TAZ interacts with TTF-1 and regulates
expression of surfactant protein-C. J Biol Chem. 279:17384–17390.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Murakami M, Nakagawa M, Olson EN and
Nakagawa O: A WW domain protein TAZ is a critical coactivator for
TBX5, a transcription factor implicated in Holt-Oram syndrome. Proc
Natl Acad Sci USA. 102:18034–18039. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Varelas X, Sakuma R, Samavarchi-Tehrani P,
Peerani R, Rao BM, Dembowy J, Yaffe MB, Zandstra PW and Wrana JL:
TAZ controls Smad nucleocytoplasmic shuttling and regulates human
embryonic stem-cell self-renewal. Nat Cell Biol. 10:837–848. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hong JH, Hwang ES, McManus MT, et al: TAZ,
a transcriptional modulator of mesenchymal stem cell
differentiation. Science. 309:1074–1078. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao D, Zhi X, Zhou Z and Chen C: TAZ
antagonizes the WWP1-mediated KLF5 degradation and promotes breast
cell proliferation and tumorigenesis. Carcinogenesis. 33:59–67.
2012. View Article : Google Scholar
|
|
43
|
Strano S, Munarriz E, Rossi M, Castagnoli
L, Shaul Y, Sacchi A, Oren M, Sudol M, Cesareni G and Blandino G:
Physical interaction with Yes-associated protein enhances p73
transcriptional activity. J Biol Chem. 276:15164–15173. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Komuro A, Nagai M, Navin NE and Sudol M:
WW domain-containing protein YAP associates with ErbB-4 and acts as
a co-transcriptional activator for the carboxyl-terminal fragment
of ErbB-4 that translocates to the nucleus. J Biol Chem.
278:33334–33341. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dong J, Feldmann G, Huang J, Wu S, Zhang
N, Comerford SA, Gayyed MF, Anders RA, Maitra A and Pan D:
Elucidation of a universal size-control mechanism in Drosophila and
mammals. Cell. 130:1120–1133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao B, Li L, Tumaneng K, Wang CY and Guan
KL: A coordinated phosphorylation by Lats and CK1 regulates YAP
stability through SCF(beta-TRCP). Genes Dev. 24:72–85. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huang W, Lv X, Liu C, Zha Z, Zhang H,
Jiang Y, Xiong Y, Lei QY and Guan KL: The N-terminal phosphodegron
targets TAZ/WWTR1 protein for SCFβ-TrCP-dependent degradation in
response to phosphatidylinositol 3-kinase inhibition. J Biol Chem.
287:26245–26253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu X, Yang N, Figel SA, Wilson KE,
Morrison CD, Gelman IH and Zhang J: PTPN14 interacts with and
negatively regulates the oncogenic function of YAP. Oncogene.
32:1266–1273. 2013. View Article : Google Scholar
|
|
49
|
Zhao B, Li L, Lu Q, Wang LH, Liu CY, Lei Q
and Guan KL: Angiomotin is a novel Hippo pathway component that
inhibits YAP oncoprotein. Genes Dev. 25:51–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Oka T, Schmitt AP and Sudol M: Opposing
roles of angiomotin-like-1 and zona occludens-2 on pro-apoptotic
function of YAP. Oncogene. 31:128–134. 2012. View Article : Google Scholar
|
|
51
|
Hamaratoglu F, Gajewski K, Sansores-Garcia
L, Morrison C, Tao C and Halder G: The Hippo tumor-suppressor
pathway regulates apical-domain size in parallel to tissue growth.
J Cell Sci. 122:2351–2359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Overholtzer M, Zhang J, Smolen GA, Muir B,
Li W, Sgroi DC, Deng CX, Brugge JS and Haber DA: Transforming
properties of YAP, a candidate oncogene on the chromosome 11q22
amplicon. Proc Natl Acad Sci USA. 103:12405–12410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cordenonsi M, Zanconato F, Azzolin L, et
al: The Hippo transducer TAZ confers cancer stem cell-related
traits on breast cancer cells. Cell. 147:759–772. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chan SW, Lim CJ, Guo K, Ng CP, Lee I,
Hunziker W, Zeng Q and Hong W: A role for TAZ in migration,
invasion, and tumorigenesis of breast cancer cells. Cancer Res.
68:2592–2598. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Piccolo S, Cordenonsi M and Dupont S:
Molecular pathways: YAP and TAZ take center stage in organ growth
and tumorigenesis. Clin Cancer Res. 19:4925–4930. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang L, Shi S, Guo Z, Zhang X, Han S, Yang
A, Wen W and Zhu Q: Overexpression of YAP and TAZ is an independent
predictor of prognosis in colorectal cancer and related to the
proliferation and metastasis of colon cancer cells. PLoS One.
8:e655392013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lu L, Li Y, Kim SM, et al: Hippo signaling
is a potent in vivo growth and tumor suppressor pathway in the
mammalian liver. Proc Natl Acad Sci USA. 107:1437–1442. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lee KP, Lee JH, Kim TS, et al: The
Hippo-Salvador pathway restrains hepatic oval cell proliferation,
liver size, and liver tumorigenesis. Proc Natl Acad Sci USA.
107:8248–8253. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Avruch J, Zhou D and Bardeesy N: YAP
oncogene over-expression supercharges colon cancer proliferation.
Cell Cycle. 11:1090–1096. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
de Cristofaro T, Di Palma T, Ferraro A,
Corrado A, Lucci V, Franco R, Fusco A and Zannini M: TAZ/WWTR1 is
over-expressed in papillary thyroid carcinoma. Eur J Cancer.
47:926–933. 2011. View Article : Google Scholar
|
|
61
|
Yuen HF, McCrudden CM, Huang YH, Tham JM,
Zhang X, Zeng Q, Zhang SD and Hong W: TAZ expression as a
prognostic indicator in colorectal cancer. PLoS One. 8:e542112013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang Y, Dong Q, Zhang Q, Li Z, Wang E and
Qiu X: Overexpression of yes-associated protein contributes to
progression and poor prognosis of non-small-cell lung cancer.
Cancer Sci. 101:1279–1285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Xie M, Zhang L, He CS, Hou JH, Lin SX, Hu
ZH, Xu F and Zhao HY: Prognostic significance of TAZ expression in
resected non-small cell lung cancer. J Thorac Oncol. 7:799–807.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yue G, Sun X, Gimenez-Capitan A, Shen J,
Yu L, Teixido C, Guan W, Rosell R, Liu B and Wei J: TAZ is highly
expressed in gastric signet ring cell carcinoma. Biomed Res Int.
2014:3930642014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Lam-Himlin DM, Daniels JA, Gayyed MF, Dong
J, Maitra A, Pan D, Montgomery EA and Anders RA: The hippo pathway
in human upper gastrointestinal dysplasia and carcinoma: A novel
oncogenic pathway. Int J Gastrointest Cancer. 37:103–109. 2006.
|
|
67
|
Zhao B, Li L, Wang L, Wang CY, Yu J and
Guan KL: Cell detachment activates the Hippo pathway via
cytoskeleton reorganization to induce anoikis. Genes Dev. 26:54–68.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yuan M, Tomlinson V, Lara R, et al:
Yes-associated protein (YAP) functions as a tumor suppressor in
breast. Cell Death Differ. 15:1752–1759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Danovi SA, Rossi M, Gudmundsdottir K, Yuan
M, Melino G and Basu S: Yes-associated protein (YAP) is a critical
mediator of c-Jun-dependent apoptosis. Cell Death Differ.
15:217–219. 2008. View Article : Google Scholar
|
|
70
|
Lai D, Ho KC, Hao Y and Yang X: Taxol
resistance in breast cancer cells is mediated by the hippo pathway
component TAZ and its downstream transcriptional targets Cyr61 and
CTGF. Cancer Res. 71:2728–2738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liu-Chittenden Y, Huang B, Shim JS, Chen
Q, Lee SJ, Anders RA, Liu JO and Pan D: Genetic and pharmacological
disruption of the TEAD-YAP complex suppresses the oncogenic
activity of YAP. Genes Dev. 26:1300–1305. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jiao S, Wang H, Shi Z, et al: A peptide
mimicking VGLL4 function acts as a YAP antagonist therapy against
gastric cancer. Cancer Cell. 25:166–180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhou Z, Hu T, Xu Z, et al: Targeting Hippo
pathway by specific interruption of YAP-TEAD interaction using
cyclic YAP-like peptides. FASEB J. Nov 10–2014.(Epub ahead of
print). PubMed/NCBI
|
|
74
|
Anand R, Maksimoska J, Pagano N, Wong EY,
Gimotty PA, Diamond SL, Meggers E and Marmorstein R: Toward the
development of a potent and selective organoruthenium mammalian
sterile 20 kinase inhibitor. J Med Chem. 52:1602–1611. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Rosenbluh J, Nijhawan D, Cox AG, et al:
β-Catenin-driven cancers require a YAP1 transcriptional complex for
survival and tumorigenesis. Cell. 151:1457–1473. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Frangou C, Li YW, Shen H, et al: Molecular
profiling and computational network analysis of TAZ-mediated
mammary tumorigenesis identifies actionable therapeutic targets.
Oncotarget. 5:12166–12176. 2014.PubMed/NCBI
|
|
77
|
Sansores-Garcia L, Bossuyt W, Wada K,
Yonemura S, Tao C, Sasaki H and Halder G: Modulating F-actin
organization induces organ growth by affecting the Hippo pathway.
EMBO J. 30:2325–2335. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fernández BG, Gaspar P, Brás-Pereira C,
Jezowska B, Rebelo SR and Janody F: Actin-Capping Protein and the
Hippo pathway regulate F-actin and tissue growth in Drosophila.
Development. 138:2337–2346. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wada K, Itoga K, Okano T, Yonemura S and
Sasaki H: Hippo pathway regulation by cell morphology and stress
fibers. Development. 138:3907–3914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Thomasy SM, Morgan JT, Wood JA, Murphy CJ
and Russell P: Substratum stiffness and latrunculin B modulate the
gene expression of the mechanotransducers YAP and TAZ in human
trabecular meshwork cells. Exp Eye Res. 113:66–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kono K, Tamashiro DA and Alarcon VB:
Inhibition of RHO-ROCK signaling enhances ICM and suppresses TE
characteristics through activation of Hippo signaling in the mouse
blastocyst. Dev Biol. 394:142–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Miller E, Yang J, DeRan M, Wu C, Su AI,
Bonamy GM, Liu J, Peters EC and Wu X: Identification of
serum-derived sphin-gosine-1-phosphate as a small molecule
regulator of YAP. Chem Biol. 19:955–962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Mo JS, Yu FX, Gong R, Brown JH and Guan
KL: Regulation of the Hippo-YAP pathway by protease-activated
receptors (PARs). Genes Dev. 26:2138–2143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Fleming JK, Wojciak JM, Campbell MA and
Huxford T: Biochemical and structural characterization of
lysophosphatidic Acid binding by a humanized monoclonal antibody. J
Mol Biol. 408:462–476. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ponnusamy S, Selvam SP, Mehrotra S,
Kawamori T, Snider AJ, Obeid LM, Shao Y, Sabbadini R and Ogretmen
B: Communication between host organism and cancer cells is
transduced by systemic sphingosine kinase 1/sphingosine 1-phosphate
signalling to regulate tumour metastasis. EMBO Mol Med. 4:761–775.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Bao Y, Nakagawa K, Yang Z, Ikeda M,
Withanage K, Ishigami-Yuasa M, Okuno Y, Hata S, Nishina H and Hata
Y: A cell-based assay to screen stimulators of the Hippo pathway
reveals the inhibitory effect of dobutamine on the YAP-dependent
gene transcription. J Biochem. 150:199–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Azzolin L, Zanconato F, Bresolin S,
Forcato M, Basso G, Bicciato S, Cordenonsi M and Piccolo S: Role of
TAZ as mediator of Wnt signaling. Cell. 151:1443–1456. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Huang SM, Mishina YM, Liu S, et al:
Tankyrase inhibition stabilizes axin and antagonizes Wnt
signalling. Nature. 461:614–620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lau T, Chan E, Callow M, et al: A novel
tankyrase small-molecule inhibitor suppresses APC mutation-driven
colorectal tumor growth. Cancer Res. 73:3132–3144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Sorrentino G, Ruggeri N, Specchia V, et
al: Metabolic control of YAP and TAZ by the mevalonate pathway. Nat
Cell Biol. 16:357–366. 2014. View Article : Google Scholar : PubMed/NCBI
|