Introduction
Cervical tumor is the seventh most common cancer
overall and the third among women (1). Annually, more than 500,000 new cases
of invasive cervical carcinoma are diagnosed globally, representing
~10% of all cancers in females. Furthermore, 80% of the cases occur
in developing countries and the survival rate is still poor
(2). Although significant advances
have been made in cervical cancer diagnosis, late stage diagnosis
and poor survival are major challenges. As infection by human
papillomavirus (HPV) types 16 and 18 has been shown to be a central
causal agent for cervical cancer (3,4),
consensus panels of the International Agency for Research on Cancer
(IARC) and WHO concluded that there is enough justification to
evaluate HPV testing as an adjunct to Papanikolaou (Pap) smear
screening for cervical cancer (5,6).
However, due to the lack of knowledge regarding the development and
progression of cervical cancer, it still has one of the highest
mortality rates among the common malignancies in women. It is
urgent to provide insight into the mechanism of cervical cancer,
with the expectation of finding more effective and alternative
molecular targets for treatment of this disease.
Comparative proteomic analysis provides a
comprehensive evaluation of protein expression, and has been widely
applied in cancer research (7).
For example, Annexin A2 (ANXA2) was demonstrated as a factor linked
to cell transformation and oncogenesis in cervical cancer via
two-dimensional electrophoresis (2-DE) and MALDI-TOF mass
spectrometric analysis (8). Based
on 2-DE analysis between human cervical squamous cell carcinoma
(SCC) and normal cervical tissues, overexpression of cytokeratin-19
was linked to prosoplasia and uncontrolled proliferation of
cervical epithelial tissue (9). In
another investigation, differences in the abundance of
zinca-2-glycoprotein (ZAG) in serum between endometrial
adenocarcinoma patients and cervical tumor patients were examined
by 2-DE with silver staining. Moreover, ZAG was identified as an
indicator linked to the development of cachexia (10). However, considerable inter-gel
variation, low sensitivity and excessive time/labor costs are
common disadvantages with the standard 2-DE approach (7,11).
Isotope-based quantitative proteomics have been
employed in analyzing post-translational modifications or
discovering biomarkers. Compared to other isotope-based
technologies, such as isotope-coded affinity tags (ICAT), 18O and
stable isotope labeling with amino acids in cell culture (SILAC)
(12–14), the isobaric tags for relative and
absolute quantitation (iTRAQ) method is deemed to be outstanding,
since it can analyze multiple samples in one experiment for
simultaneous analysis. In the present study, iTRAQ labeling coupled
with high resolution mass spectrometry was carried out to detect
the difference in proteins between cervical cancer and non-cancer
samples. We expected to find host proteins
Materials and methods
Tissues and cell line
The present study was approved by the Institutional
Review Board for Human Subject Review. Written informed consent was
obtained from all the patients in accordance with the revised
Helsinki Declaration. A total of 8 human cervical cancer samples
and 8 paired non-cancer samples were collected at the Second
Affiliated Hospital of Chongqing Medical University. The HeLa cell
line was purchased from the Cancer Research Department of China
Medical Science Institute.
iTRAQ labeling and peptide
fractionation
Total proteins (100 μg) extracted from each sample
were denatured, cysteine blocked, and digested as described in the
standard protocol of the iTRAQ kit (AB Sciex, Framingham, MA, USA).
Pooled non-tumor samples were labeled with iTRAQ tags 113 and 115,
and tags 114 and 116 were used to label pooled cervical tumor
samples. The labeled peptides were pooled in 1:1:1:1 ratio and
lyophilized. The mixture was separated with an LC-30 high
performance liquid chromatography system (Shimadzu, Kyoto, Japan)
by gradient elution. The detailed procedures are shown in Table I. Ten fractions were collected and
lyophilized for further analysis.
 | Table IExperimental procedures of high
performance liquid chromatography (HPLC) for peptide fractionation
and NanoLC-MS/MS analysis. |
Table I
Experimental procedures of high
performance liquid chromatography (HPLC) for peptide fractionation
and NanoLC-MS/MS analysis.
| RP-HPLC for peptide
fractionation | NanoLC-MS/MS
analysis |
|---|
|
|
|---|
| Gradient
elution | Mobile phase A (20
mM ammonium formate, pH 10.0) | Mobile phase B (80%
acetonitrile with 20 mM ammonium formate, pH 10.0) | Gradient
elution | Mobile phase A (2%
acetonitrile with 0.1% formic acid) | Mobile phase B (98%
acetonitrile with 0.1% formic acid) |
|---|
| 0 min | 95% | 5% | 0 min | 95% | 5% |
| 5 min | 95% | 5% | 5 min | 95% | 5% |
| 25 min | 77% | 23% | 85 min | 75% | 25% |
| 40 min | 55% | 45% | 105 min | 50% | 50% |
| 41 min | 10% | 90% | 106 min | 20% | 80% |
| 45 min | 10% | 90% | 110 min | 20% | 80% |
| 46 min | 95% | 5% | 111 min | 95% | 5% |
| 60 min | 95% | 5% | 120 min | 95% | 5% |
| Detector: UV 220
nm | Detector:
Triple-TOF 5600 MS/MS |
NanoLC-2D-TripleTOF 5600 analysis
A Triple TOF 5600 system coupled to an Eksigent
NanoLC-2D system (AB Sciex) was used for protein identification and
quantization. Each fraction was separated in a 2-h gradient elution
by the NanoLC-2D system (Table I).
The mass spectrometer was set in the positive ion mode at a mass
range of 350–1,500 m/z, with a 0.25-sec accumulation time, followed
by information-dependent acquisition (IDA). The top 30 precursor
ions within each cycle were automatically selected for
fragmentation, with each MS/MS spectrum accumulated for 0.1 sec
(100–1,500 m/z).
ProteinPilot v.4.5 software (AB Sciex) was used for
data search against the UniProt database. The standard searching
parameters and false discovery rate analysis were set. A threshold
of confidence >99% and a local false discovery rate (FDR) of
<1% were used for both protein identification and quantitative
analysis. More than 2 unique peptides were required for protein
identification. P-values <0.01 were required for relative
quantification. The PeakView 1.1 software was used to extract ion
chromatograms.
Quantitative real time-PCR analysis
(qRT-PCR)
Total RNA was extracted using the RNeasy Mini kit
(Qiagen, Hilden, Germany), and 100 ng of total RNA was reverse
transcribed into cDNA using the PrimeScript RT reagent kit (Takara,
Dalian, China). qRT-PCR assays were performed on an ABI 7900HT
system with TaqMan kits. Primers for G6PD (Hs00166169-m1), STAT1
(Hs01014007_m1), HSPB1 (Hs03044127_g1), DCN (Hs00466796_CE),
ALDH3A1 (Hs00964880_m1), EPX (Hs00417510_CE), PRG3 (Hs00196082_m1),
OGN (Hs00247901_m1), CRNN (Hs00211833_m1), AGR2 (Hs00356521_m1),
ORM2 (Hs00301996_CE) and GAPDH (Hs02758991_g1) were used. Each
sample was run in triplicate, and all reactions were performed at
least twice. The 2−ΔΔCT method was used for data
analysis.
Western blot analysis
Total protein (20 μg) extracted from each tissue
sample was separated by SDS-PAGE, and transferred to
polyvinylpyrrolidone membrane (Amersham Biosciences, Uppsala,
Sweden). Primary antibodies (1:500–1:1,000) against G6PD, STAT1,
HSPB1, ALDH3A1 and FSCN1 (Abcam, Cambridge, MA, USA) and
HRP-conjugated secondary antibodies (1:5,000; Amersham Biosciences)
were used to incubate the samples. Bands were detected with an ECL
detection system (Amersham Biosciences). Each sample was analyzed
at least twice. Detailed procedure of experiments was described in
a previous study (15).
Immunohistochemistry (IHC) and tissue
microarray
The tissue microarrays (CR802) were purchased from
US Biomax, Inc. (Rockville, MD, USA) to detect G6PD, STAT1, HSPB1
and ALDH3A1 in cores from 40 cervical tumor tissues and 40
non-tumor tissues, antibodies against G6PD (1:90), STAT1 (1:200),
HSPB1 (1:250) and ALDH3A1 (1:200) were added and incubated
overnight at 4°C. Detection was performed with the
Envision/horseradish peroxidase system (DakoCytomation, Glostrup,
Denmark). Semi-quantification of protein expression was defined by
scoring criteria. The positive cells (%) and staining intensity
(scale 0–3) were checked, which were then multiplied to yield a
score ranging from 0 to 300. To maintain consistency, the same
qualified pathologist gave interpretations for all IHC data.
Wound healing and invasion assay
G6PD-specific (50 nM) siRNAHSS103891, HSS103892 and
HSS103893 (Invitrogen, Carlsbad, CA, USA) or negative control siRNA
(12935-400; Invitrogen) were transfected into HeLa cells
(105 cell/well). After 48 h, wound healing and invasion
experiments were performed on 6-well plates seeded with HeLa cells.
After the cells reached confluency, a 200 μl pipette tip was used
to incise the cell monolayer. The debris was rinsed away and
removed. The extent of gap closure was monitored and photographed
under a microscope up to 24 h. The invasion assays were performed
using a Cell Invasion Assay kit (Cell Biolabs, San Diego, CA, USA),
following the manufacturer’s instructions. After 24 h, the number
of cells that invaded and attached to the bottom chamber was
measured by CyQuant GR fluorescent dye (560 nm).
Statistical analyses
Statistical analyses were performed by SPSS software
v13.0 using the Student’s t-test, Mann-Whitney U-test,
χ2 test or Spearman’s rank correlation analysis. A
P-value <0.05 was considered statistically significant. All
tests of significance were two-tailed.
Results
Functional profiles of differentially
expressed proteins in cervical tumors
In total, 3,647 proteins were identified with 1%
global FDR from fit in cervical cancer following a workflow shown
in Fig. 1. For subsequent relative
quantification analysis, a cut-off of 1.3-fold change, up or down,
was applied to all iTRAQ ratios to minimize false positives when
identifying proteins as overerexpressed or downregulated. This
process is widely adopted in other proteomics investigations
(16–19). Accordingly, 294 proteins were
identified as differentially expressed in pooled cervical tumor
tissues comparing to non-tumor tissues, including 130 upregulated
and 164 down-regulated proteins. The top 30 upregulated proteins
and top 30 downregulated proteins were listed in Table II.
| Table IIPartial list of the differentially
expressed proteins in cervical cancer. |
Table II
Partial list of the differentially
expressed proteins in cervical cancer.
| N | Accession | Gene sym | Protein name | Peptides (95%) | Pooled tumor
tissues: pooled non-tumor tissues (114:113)a | P-valueb | Pooled tumor
tissues: pooled non-tumor tissues (116:115) a | P-valueb |
|---|
| Top 30 proteins
downregulated in cervical cancer tissues |
| 1 |
IPI:IPI00025465.2 | OGN | cDNA FLJ59205,
highly similar to mimecan | 104 | 0.01674943 | 7.82E-12 | 0.021086279 | 5.80E-13 |
| 2 |
IPI:IPI00297056.2 | CRNN | Cornulin | 19 | 0.017864879 | 1.18E-07 | 0.02290868 | 4.43E-08 |
| 3 |
IPI:IPI00012119.1 | DCN | Isoform A of
decorin | 96 | 0.018365379 | 1.10E-09 | 0.023988331 | 4.67E-11 |
| 4 |
IPI:IPI00980755.1 | PRELP | PRELP protein
(fragment) | 73 | 0.02051162 | 7.91E-10 | 0.02831392 | 8.35E-12 |
| 5 |
IPI:IPI00297646.5 | COL1A1 | Collagen α-1(I)
chain | 473 | 0.023988331 | 1.33E-15 | 0.02831392 | 3.99E-13 |
| 6 |
IPI:IPI00940343.1 | TPM1 | cDNA FLJ16459 fis,
clone similar to BRCAN 2002473, moderately tropomyosin, fibroblast
isoform 2 | 59 | 0.02466039 | 2.96E-06 | 0.043651581 | 1.79E-07 |
| 7 |
IPI:IPI00021264.1 | CNN1 | Calponin-1 | 16 | 0.026061529 | 3.32E-06 | 0.04613176 | 4.03E-06 |
| 8 |
IPI:IPI00553177.1 | SERPINA1 | Isoform 1 of
α-1-antitrypsin | 128 | 0.026546059 | 6.37E-14 | 0.04830588 | 7.74E-13 |
| 9 |
IPI:IPI00440822.3 | ABI3BP | Isoform 1 of target
of Nesh-SH3 | 7 | 0.031045601 | 1.62E-05 | 0.02187762 | 0.000515066 |
| 10 |
IPI:IPI00007427.2 | AGR2 | AGR2 | 27 | 0.03372873 | 4.18E-08 | 0.048752852 | 8.75E-08 |
| 11 |
IPI:IPI00020091.1 | ORM2 | α-1-acid
glycoprotein 2 | 11 | 0.03404082 | 0.00138629 | 0.08550667 | 0.00133485 |
| 12 |
IPI:IPI00643384.2 | BGN | cDNA FLJ36740 fis,
clone UTERU 2013322, highly similar to biglycan | 109 | 0.034355789 | 1.80E-14 | 0.042461962 | 6.51E-14 |
| 13 |
IPI:IPI00021854.1 | APOA2 | Apolipoprotein
A-II | 46 | 0.034355789 | 1.33E-08 | 0.044874541 | 3.02E-08 |
| 14 |
IPI:IPI00292130.3 | DPT | Dermatopontin | 18 | 0.03467368 | 7.33E-05 | 0.058613818 | 2.40E-05 |
| 15 |
IPI:IPI00298828.3 | APOH | β-2-glycoprotein
1 | 24 | 0.036982819 | 8.84E-10 | 0.084722742 | 6.80E-09 |
| 16 |
IPI:IPI00216138.6 | TAGLN | Transgelin | 49 | 0.03872576 | 5.68E-12 | 0.060813501 | 2.67E-10 |
| 17 |
IPI:IPI00027827.2 | SOD3 | Extracellular
superoxide dismutase [Cu-Zn] | 15 | 0.039810721 | 7.61E-05 | 0.089536473 | 0.002630402 |
| 18 |
IPI:IPI00304962.4 | COL1A2 | Collagen α-2(I)
chain | 302 | 0.044874541 | 2.33E-10 | 0.075162292 | 5.49E-09 |
| 19 |
IPI:IPI00000860.2 | FMOD | Fibromodulin | 12 | 0.045289759 | 0.00012670 | 0.057543989 | 0.000182819 |
| 20 |
IPI:IPI00514285.2 | PTGDS | Uncharacterized
protein | 6 | 0.045289759 | 0.00596775 | 0.059156161 | 0.001384994 |
| 21 |
IPI:IPI00220327.4 | KRT1 | Keratin, type II
cytoskeletal 1 | 57 | 0.054954089 | 6.40E-09 | 0.152756602 | 3.95E-08 |
| 22 |
IPI:IPI00418431.1 | ASPN | ASPN protein | 35 | 0.057543989 | 8.29E-06 | 0.052966349 | 2.79E-07 |
| 23 |
IPI:IPI00017601.1 | CP | Ceruloplasmin | 42 | 0.064268783 | 8.88E-16 | 0.1018591 | 1.02E-14 |
| 24 |
IPI:IPI00008580.1 | SLPI |
Antileukoproteinase | 7 | 0.064268783 | 4.86E-05 | 0.100000001 | 2.92E-05 |
| 25 |
IPI:IPI00792011.1 | CAPS | Calcyphosin | 6 | 0.064863443 | 0.00500057 | 0.107646503 | 0.006311405 |
| 26 |
IPI:IPI00215894.1 | KNG1 | Isoform LMW of
kininogen-1 | 19 | 0.064863443 | 6.10E-09 | 0.084722742 | 3.88E-06 |
| 27 |
IPI:IPI00219219.3 | LGALS1 | Galectin-1 | 62 | 0.067297667 | 1.03E-10 | 0.075857759 | 2.48E-10 |
| 28 |
IPI:IPI00176903.2 | PTRF | Isoform 1 of
polymerase I and transcript release factor | 45 | 0.067297667 | 8.64E-07 | 0.077268057 | 2.99E-07 |
| 29 |
IPI:IPI00009829.4 | CPA3 | Mast cell
carboxypeptidase A | 11 | 0.067920357 | 1.34E-06 | 0.139315695 | 2.31E-05 |
| 30 |
IPI:IPI00219684.3 | FABP3 | Fatty acid-binding
protein, heart | 5 | 0.069183096 | 0.03231849 | 0.170608193 | 0.003044094 |
| Top 30 proteins
upregulated in cervical cancer tissues |
| 1 |
IPI:IPI00020021.3 | DEK | Protein DEK | 9 | 4.965922832 | 0.00015781 | 5.296635151 | 0.000133302 |
| 2 |
IPI:IPI00010090.1 | GCLM | Glutamate-cysteine
ligase regulatory subunit | 7 | 5.058247089 | 0.007747813 | 4.570881844 | 0.017547339 |
| 3 |
IPI:IPI00783625.2 | SERPINB5 | Isoform 1 of serpin
B5 | 35 | 5.105050087 | 1.36E-07 | 4.285484791 | 2.01E-07 |
| 4 |
IPI:IPI00295400.1 | WARS | Isoform 1 of
tryptophanyl-tRNA synthetase, cytoplasmic | 55 | 5.152287006 | 7.21E-08 | 5.861382008 | 6.93E-06 |
| 5 |
IPI:IPI00852685.1 | DIAPH1 | Isoform 1 of
protein diaphanous homolog 1 | 19 | 5.248075008 | 0.000127102 | 4.246195793 | 0.006647518 |
| 6 |
IPI:IPI00219078.5 | ATP2A2 | Isoform 1 of
sarcoplasmic/endoplasmic reticulum calcium ATPase 2 | 34 | 5.296635151 | 2.82E-06 | 5.058247089 | 1.17E-06 |
| 7 |
IPI:IPI00152503.1 | DTX3L | Isoform 1 of E3
ubiquitin-protein ligase DTX3L | 7 | 5.345643997 | 0.000278658 | 6.309574127 | 0.000135678 |
| 8 |
IPI:IPI00031517.1 | MCM6 | DNA replication
licensing factor MCM6 | 11 | 5.546257019 | 0.000830936 | 5.970353127 | 0.000910989 |
| 9 |
IPI:IPI00030781.1 | STAT1 | Isoform α of signal
transducer and activator of transcription 1-α/β | 58 | 5.861382008 | 1.50E-07 | 5.152287006 | 9.05E-06 |
| 10 |
IPI:IPI00220194.6 | SLC2A1 | Solute carrier
family 2, facilitated glucose transporter member 1 | 7 | 5.861382008 | 0.01236643 | 6.918310165 | 0.008381192 |
| 11 |
IPI:IPI00450768.7 | KRT17 | Keratin, type I
cytoskeletal 17 | 247 | 6.194410801 | 1.06E-09 | 7.379042149 | 2.16E-06 |
| 12 |
IPI:IPI00019869.3 | S100A2 | Protein
S100-A2 | 18 | 6.194410801 | 6.23E-05 | 4.920394897 | 0.008037753 |
| 13 |
IPI:IPI00465343.3 | ADH1C | Alcohol
dehydrogenase 1C | 10 | 6.426877022 | 7.09E-05 | 3.767038107 | 0.004329687 |
| 14 |
IPI:IPI00292532.6 | CAMP | Cathelicidin
antimicrobial peptide precursor | 3 | 6.426877022 | 0.025978809 | 7.311390877 | 0.01136459 |
| 15 |
IPI:IPI00028564.2 | GBP1 | Interferon-induced
guanylate-binding protein 1 | 32 | 6.486343861 | 2.07E-05 | 6.668066978 | 2.52E-05 |
| 16 |
IPI:IPI00853547.1 | G6PD | Isoform 3 of
glucose-6-phosphate 1-dehydrogenase | 29 | 6.486343861 | 1.52E-06 | 6.137619972 | 4.52E-05 |
| 17 |
IPI:IPI00654709.1 | ALDH3A1 | Aldehyde
dehydrogenase | 29 | 7.04693079 | 8.37E-07 | 7.655965805 | 1.36E-08 |
| 18 |
IPI:IPI00002405.5 | OAS3 |
2′-5′-oligoadenylate synthase 3 | 11 | 7.447319984 | 0.000239752 | 7.447319984 | 0.001038955 |
| 19 |
IPI:IPI00236556.1 | MPO | Isoform H7 of
myeloperoxidase | 48 | 7.870458126 | 1.09E-08 | 6.854881763 | 1.48E-10 |
| 20 |
IPI:IPI00794567.1 | TAPBP | Uncharacterized
protein | 16 | 8.090958595 | 0.01492988 | 7.17794323 | 0.030335421 |
| 21 |
IPI:IPI00219525.10 | PGD | 6-phosphogluconate
dehydrogenase, decarboxylating | 61 | 8.165823936 | 0.000105162 | 9.20449543 | 3.39E-10 |
| 22 |
IPI:IPI00105407.2 | AKR1B10 | Aldo-ketoreductase
family 1 member B10 | 48 | 9.036495209 | 0.003378323 | 11.37627029 | 0.002583705 |
| 23 |
IPI:IPI00217468.3 | HIST1H1B | Histone H1.5 | 31 | 9.120108604 | 0.000225787 | 10.28015995 | 0.000171618 |
| 24 |
IPI:IPI00006690.1 | EPX | Eosinophil
peroxidase | 61 | 9.375619888 | 4.03E-10 | 7.311390877 | 1.20E-05 |
| 25 |
IPI:IPI00607801.2 | CES1 | Isoform 2 of liver
carboxylesterase 1 | 67 | 13.80383968 | 9.04E-05 | 14.72313023 | 8.45E-05 |
| 26 |
IPI:IPI00005778.2 | PRG3 | Proteoglycan 3 | 4 | 14.58813953 | 0.02189479 | 13.80383968 | 0.026921131 |
| 27 |
IPI:IPI00018873.1 | NAMPT |
Nicotinamidephosphoribosyltransferase | 63 | 15.13560963 | 0.001666992 | 13.55189037 | 0.002346458 |
| 28 |
IPI:IPI00908386.1 | ADH7 | alcohol
dehydrogenase class 4 mu/σ chain isoform 1 | 19 | 16.90440941 | 0.007982331 | 18.87990952 | 0.00664382 |
| 29 |
IPI:IPI00005721.1 | DEFA1 | Neutrophil defensin
1 | 11 | 17.70108986 | 6.93E-05 | 13.42765045 | 6.16E-05 |
| 30 |
IPI:IPI00005668.4 | AKR1C2 | Aldo-ketoreductase
family 1 member C2 | 44 | 22.28434944 | 0.001423413 | 20.89295959 | 0.001390291 |
To obtain the functional characteristics of proteins
associated with cervical carcinoma oncogenesis, we classified the
294 proteins using Protein Analysis through Evolutionary
Relationships Classification System (PANTHER, www.pantherdb.org). Twelve biological processes are
involved, with 55.0% of the proteins participating in metabolic
processes, followed by cellular processes (37.1%) and developmental
processes (19.4%). According to molecular function, the proteins
were divided into 10 categories, including catalytic activity
(37.8%), binding activity (32.0%) and structural molecule activity
(19.4%). The 294 proteins were grouped into 27 protein classes,
including cytoskeletal protein (14.7%), hydrolase (12.9%) and
nucleic acid binding proteins (10.4%). A total of 63 signaling
pathways were associated, with integrin signaling pathway (3.2%),
blood coagulation (2.9%) and inflammation mediated by chemokine and
cytokine signaling pathway (2.2%) at the top of the list (Fig. 2).
Validation of altered expression levels
of proteins
The iTRAQ study results were further validated by
qRT-PCR and western blot analyses. Fig. 3A shows the relative mRNA levels of
selected differentially expressed proteins in the cervical tumor
tissues, compared to those in the paired non-tumor tissues. The
mRNA levels of G6PD, HSPB1, STAT1, ALDH3A1, FSCN1, EPX and PRG3
were found to be upregulated, whereas the levels of DCN, OGN, CRNN,
AGR2 and ORM2 were down-regulated. The upregulation of protein
levels of G6PD, HSPB1, STAT1, ALDH3A1 and FSCN1 were subsequently
detected by western blot analysis (Fig. 3B). This trend matched that observed
in the iTRAQ method.
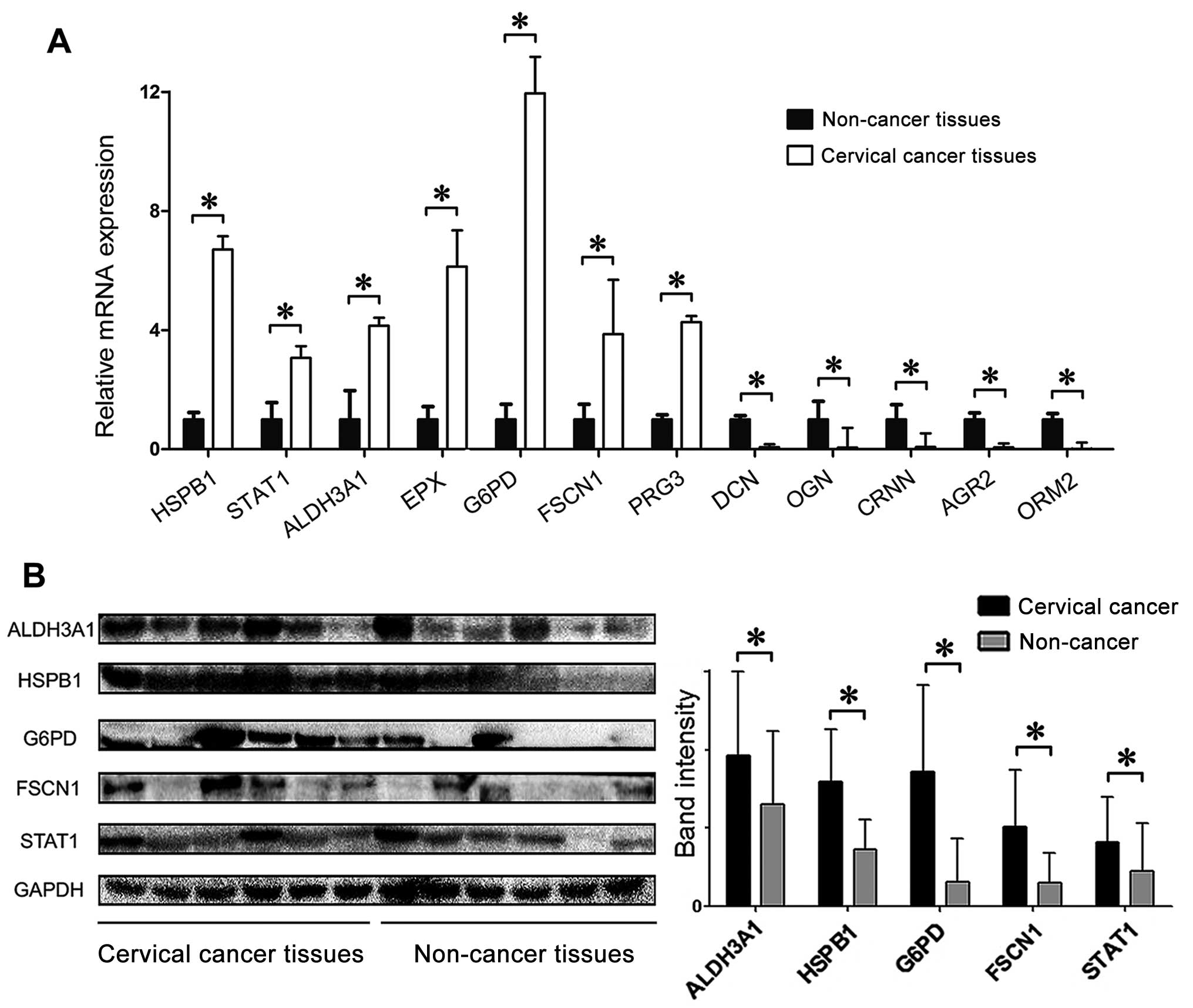 | Figure 3qRT-PCR and western blot analyses of
differentially expressed proteins in cervical cancer. The relative
mRNA expression levels of HSPB1, STAT1, ALDH3A1, EPX, G6PD, FSCN1,
PRG3, DCN, OGN, CRNN, AGR2 and ORM2 (A), and the high protein
levels of ALDH3A1, HSPB1, G6PD, FSCN1 and STAT1 (B) in cervical
cancer tissues compared with non-cancer cervical tissues. Bars
indicate SD. *P<0.05. |
Expression of G6PD, HSPB1, STAT1 and
ALDH3A1 in tissue array
The clinical relevance of G6PD, HSPB1, STAT1 and
ALDH3A1 in cervical cancer was assessed by IHC analysis (Fig. 4). A tissue microarray including 40
cervical cancer tissues and 40 matched or unmatched non-cancer
cervical tissues was analyzed. As a result, cervical cancer samples
showed significantly higher levels of G6PD, HSPB1, STAT1 and
ALDH3A1 than those in controls. Moreover, G6PD expression was
detected in 100% (40/40) of cervical cancer samples, compared to
45% (18/40) in controls. The staining intensity of G6PD in cervical
cancer cells was much stronger than that in control epithelial
cells. Similar trends were observed in the IHC analysis of HSPB1,
STAT1 and ALDH3A1.
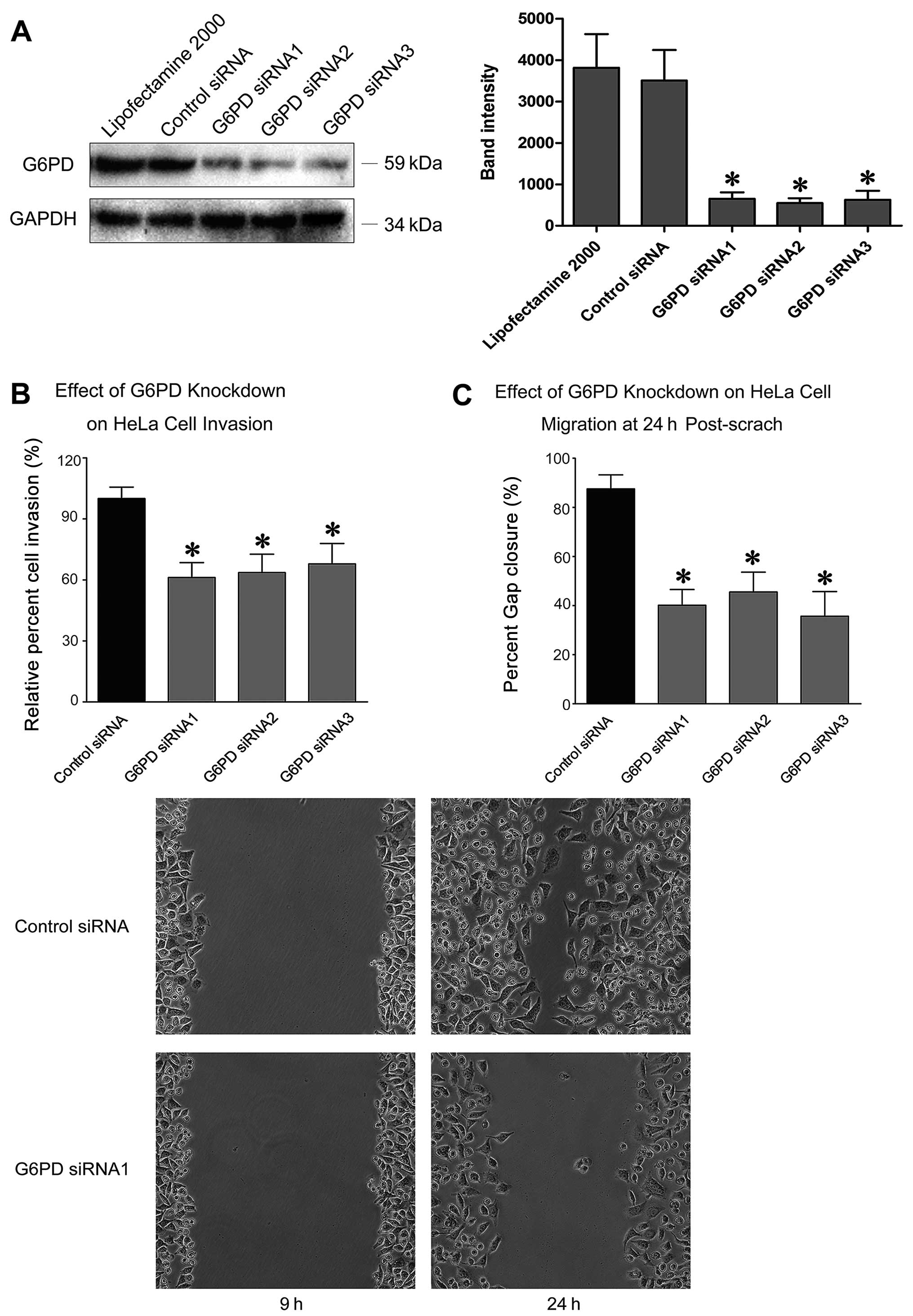
G6PD knockdown compromised HeLa cells’
invasion and migration
The dramatic increase of G6PD in cervical cancer
suggested that G6PD not only contributes to the biosynthesis of
cervical cancer cells, but also is crucial for their malignancy. To
test this hypothesis, HeLa, a human cervical cancer cell line, was
tested with an RNA interference assay. G6PD expression in HeLa
cells was initially silenced by transfection of G6PD-specific
siRNAs (Fig. 5A). G6PD-silenced
HeLa cells and control cells were then subjected to invasion and
migration assays. Invasion capacity of G6PD-silenced cells was
inhibited by 30–40% when compared to that of the control cells
(P<0.01) (Fig. 5B). The readout
of the scratch wound repair assays was reduced by 55–65% in
G6PD-silenced cells when compared to that of the control cells
(P<0.01) (Fig. 5C). Our results
supported the notion that G6PD may be an effective target in
cervical cancer treatment.
Discussion
Cervical cancer remains one of the leading causes of
death among women throughout the world. The morbidity and death
rates remain frustratingly high in spite of several emerging
biomarkers for diagnosis. Discovery of novel factors that are
related to the biology of cervical carcinoma is an urgent task for
improving current treatment strategies and prognosis of the
disease.
In the present study, we identified proteins
differentially expressed between tumor and non-tumor cervical
tissues via the iTRAQ proteomics approach. Verification studies
using qRT-PCR, immunoblot and IHC assays confirmed that the
expressions of G6PD, STAT1, ALDH3A1 and HSPB1 were, indeed,
significantly increased in tumor tissues. Functional studies
indicated that G6PD tends to act on migration and invasion of
cervical cancer cells. Our findings revealed that the iTRAQ method
for large-scale protein quantification is amenable to high
throughput processes and credible, and some novel proteins
uncovered here may serve as potential targets for cervical tumor
treatment.
G6PD, as the first and rate-limiting enzyme in the
ubiquitous pentose phosphate pathway (PPP) (20), participates in biological processes
of various cancers (21–23). Through PPP, the biosynthesis and
rapid proliferation of tumor cells are supported (24,25).
In non-tumor cells, G6PD binds with p53 protein to prevent it from
forming the active dimer (26).
This process, however, inhibits PPP by quenching functional G6PD
(27). Notably, this inhibition of
G6PD by p53 does not exist in tumor cells, because mutant p53 loses
its inhibitory effect on G6PD, and, thus, results in elevated PPP
glucose flux, which ultimately enhances glucose consumption and
promotes tumor cell biosynthesis (26). In other words, G6PD motivates
cancer cell proliferation and contributes to tumorigenesis by
increasing PPP glucose flux. Since the relationship between G6PD
and cervical cancer was not well investigated previously, we
examined the effect of G6PD proteins on HeLa cells. As a
consequence, the invasion and migration capabilities of HeLa cells
were decreased in vitro when G6PD expression was suppressed.
Our results, together with findings of other groups, indicate that
the suppression of G6PD expression may be beneficial in cervical
cancer treatment.
Another markedly upregulated protein in cervical
cancer found in the present study was STAT1, a member of STAT
protein family. STAT1 was noted to be critical in controlling cell
cycle progression and apoptosis. Hix et al (28) studied STAT1 effects by injecting
mouse mammary carcinoma cell TM40D with/without ectopic STAT1 into
the mammary fat pads of BALB/c mice and observed that the growth
and aggressiveness of TM40D-STAT1 cells were dramatically enhanced,
when compared to those of regular TM40D cells. Conversely, the
growth of TM40D cells was significantly delayed when STAT1 was
knocked down. Further studies demonstrated that STAT1 promoted
breast cancer by increasing interleukin-13 (IL-13), transforming
growth factor β (TGFβ) and tumor necrosis factor α (TNFα). These
factors could inhibit antitumor immune responses by recruiting and
stimulating associated cells (29). In addition, Rajkumar et al
(30) found that STAT1 rose in
early cervical intraepithelial neoplasia (CIN1/2), dropped in
CIN3/CIS, and was dramatically upregulated in invasive cervical
carcinoma, demonstrating that STAT1 may harbor double roles in HPV
infection and cervical tumorigenesis, specifically a protective
role in the early phase and an oncogenic role in the invasive
stages of the tumor. Further investigation is required to determine
the roles of STAT1 in cervical tumorigenesis.
HSPB1 protein, also named heat shock protein 27
(Hsp27), belongs to the emerging ‘survival protein’ family
(31–33). The expression of HSPB1 is involved
in cell proliferation (34) and
apoptosis (35) and is associated
with poor prognosis in various tumors (34,36).
The inhibitor of HSPB1, OGX-427, a second-generation antisense
oligonucleotide of HSPB1, downregulated the levels of HSPB1 and
induced the apoptosis of cancer cells (37). This occurs via the disruption of
the interaction of HSPB1 with cytochrome c (cytc),
procaspases-9 and -3 (35). Thus,
the attenuation of the expression of HSPB1 gene was considered as a
new therapeutic strategy. Moreover, McCollum et al (38) were able to successfully make
prostate cancer cells more sensitive to the antitumor drug
17-allylamino-demethoxygeldanamycin by employing small-interfering
RNA (siRNA) against HSPB1. Together, accumulating evidence supports
the notion that upregulation of HSPB1 stimulates cancer cell
proliferation, whereas its downregulation results in the inhibition
of tumor progression.
ALDH3A1 proteins identified in the present study has
been suggested involved in tumor progression, but not implicated in
cervical cancer. The aldehyde dehydrogenase superfamily member
ALDH3A1 belongs to the phase II group of drug-metabolizing enzymes
and plays key roles in numerous biological processes, such as cell
survival and growth (39).
Upregulation of ALDH3A1 is frequent in tumor-derived cell lines and
neoplastic tissues (40). By
removing lipid peroxidation products, the overexpression of ALDH3A1
promoted the growth of rat hepatoma cells (41). In contrast, a reduced expression of
ALDH3A1 inhibited the growth of cancer cells tested in lung tumor
cell line A549 via activating peroxisome proliferator activated
receptor γ (PPARγ), which inhibits NF-κB activity (42). These findings are consistent with
our observations made in cervical carcinoma and suggested that
ALDH3A1 may be a promising candidate target of cervical cancer
therapy.
In summary, we performed a non-targeted proteomics
study to profile differentially expressed proteins in cervical
cancer. The proteins uncovered, as well as studied, in the present
study may serve as potential targets for cervical cancer research
and treatment.
Acknowledgements
The present study was supported by grants from the
National Basic Research Program of China (2010CB945103), the
National Natural Science Foundation of China (81030013 and
81170002), the Science and Technology Project of Jiangsu Province
(BL2014053), the Ph.D. Programs Foundation of Ministry of Education
of China (20110091120032), and the Jiangsu Province Health
Department of Medicine leading talent and innovation team project
(LJ201109).
References
|
1
|
Parkin DM, Pisani P and Ferlay J:
Estimates of the worldwide incidence of eighteen major cancers in
1985. Int J Cancer. 54:594–606. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Munoz N, Bosch FX, de Sanjose S, et al:
The causal link between human papillomavirus and invasive cervical
cancer: a population-based case-control study in Colombia and
Spain. Int J Cancer. 52:743–749. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schiffman MH, Bauer HM, Hoover RN, et al:
Epidemiologic evidence showing that human papillomavirus infection
causes most cervical intraepithelial neoplasia. J Natl Cancer Inst.
85:958–964. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Franco E, Syrjanen K, de Wolf C, et al:
New developments in cervical cancer screening and prevention;
Geneva, Switzerland. June 17–19 1996; Workshop. Cancer Epidemiol
Biomarkers Prev. 5. pp. 853–856. 1996
|
|
6
|
Franco EL: Measurement errors in
epidemiological studies of human papillomavirus and cervical
cancer. IARC Sci Publ. 119:181–197. 1992.PubMed/NCBI
|
|
7
|
Sun W, Xing B, Sun Y, et al: Proteome
analysis of hepatocellular carcinoma by two-dimensional difference
gel electrophoresis: novel protein markers in hepatocellular
carcinoma tissues. Mol Cell Proteomics. 6:1798–1808. 2006.
View Article : Google Scholar
|
|
8
|
Bae SM, Lee CH, Cho YL, Nam KH, Kim YW,
Kim CK, Han BD, Lee YJ, Chun HJ and Ahn WS: Two-dimensional gel
analysis of protein expression profile in squamous cervical cancer
patients. Gynecol Oncol. 99:26–35. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bae SM, Min HJ, Ding GH, et al: Protein
expression profile using two-dimensional gel analysis in squamous
cervical cancer patients. Cancer Res Treat. 38:99–107. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abdul-Rahman PS, Lim BK and Hashim OH:
Expression of high-abundance proteins in sera of patients with
endometrial and cervical cancers: analysis using 2-DE with silver
staining and lectin detection methods. Electrophoresis.
28:1989–1996. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Görg A, Obermaier C, Boguth G, Harder A,
Scheibe B, Wildgruber R and Weiss W: The current state of
two-dimensional electrophoresis with immobilized pH gradients.
Electrophoresis. 21:1037–1053. 2007. View Article : Google Scholar
|
|
12
|
Fu C, Wu C, Liu T, Ago T, Zhai P,
Sadoshima J and Li H: Elucidation of thioredoxin target protein
networks in mouse. Mol Cell Proteomics. 8:1674–1687. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rangiah K, Tippornwong M, Sangar V, Austin
D, Tétreault MP, Rustgi AK, Blair IA and Yu K: Differential
secreted proteome approach in murine model for candidate biomarker
discovery in colon cancer. J Proteome Res. 8:5153–5164. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ross PL, Huang YN, Marchese JN, et al:
Multiplexed protein quantization in Saccharomyces cerevisiae using
amine-reactive isobaric tagging reagents. Mol Cell Proteomics.
3:1154–1169. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Y, Toy W, Choong LY, Hou P, Ashktorab
H, Smoot DT, Yeoh KG and Lim YP: Discovery of SLC3A2 cell membrane
protein as a potential gastric cancer biomarker: implications in
molecular imaging. J Proteome Res. 1:5736–5747. 2012.
|
|
16
|
Chen Y, Choong LY, Lin Q, et al:
Differential expression of novel tyrosine kinase substrates during
breast cancer development. Mol Cell Proteomics. 6:2072–2087. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chong PK, Lee H, Zhou J, Liu SC, Loh MC,
So JB, Lim KH, Yeoh KG and Lim YP: Reduced plasma APOA1 level is
associated with gastric tumor growth in MKN45 mouse xenograft
model. J Proteomics. 73:1632–1640. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fujii K, Kondo T, Yokoo H, Yamada T,
Iwatsuki K and Hirohashi S: Proteomic study of human hepatocellular
carcinoma using two-dimensional difference gel electrophoresis with
saturation cysteine dye. J Proteomics. 5:1411–1422. 2005.
View Article : Google Scholar
|
|
19
|
Gan CS, Chong PK, Pham TK and Wright PC:
Technical, experimental, and biological variations in isobaric tags
for relative and absolute quantitation (iTRAQ). J Proteome Res.
6:821–827. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pandolfi PP, Sonati F, Rivi R, Mason P,
Grosveld F and Luzzatto L: Targeted disruption of the housekeeping
gene encoding glucose 6-phosphate dehydrogenase (G6PD): G6PD is
dispensable for pentose synthesis but essential for defense against
oxidative stress. EMBO J. 14:5209–5215. 1995.PubMed/NCBI
|
|
21
|
Hu T, Zhang C, Tang Q, Su Y, Li B, Chen L,
Zhang Z, Cai T and Zhu Y: Variant G6PD levels promote tumor cell
proliferation or apoptosis via the STAT3/5 pathway in the human
melanoma xenograft mouse model. BMC Cancer. 13:2512013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang P, Du W and Yang X: A critical role
of glucose-6-phosphate dehydrogenase in TAp73-mediated cell
proliferation. Cell Cycle. 12:3720–3726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin HR, Wu CC, Wu YH, Hsu CW, Cheng ML and
Chiu DT: Proteome-wide dysregulation by glucose-6-phosphate
dehydrogenase (G6PD) reveals a novel protective role for G6PD in
aflatoxin B(1)-mediated cytotoxicity. J Proteome Res. 12:3434–3448.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: the metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang P, Du W, Wang X, Mancuso A, Gao X,
Wu M and Yang X: p53 regulates biosynthesis through direct
inactivation of glucose-6 -phosphate dehydrogenase. Nat Cell Biol.
13:310–316. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Berg J, Tymoczko J and Stryer L:
Biochemistry. 6th edition. WH Freeman & Co; New York, NY: pp.
577–589. 2006
|
|
28
|
Hix LM, Karavitis J, Khan MW, Shi YH,
Khazaie K and Zhang M: Tumor STAT1 transcription factor activity
enhances breast tumor growth and immune suppression mediated by
myeloid-derived suppressor cells. J Biol Chem. 288:11676–11688.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang M: Novel function of STAT1 in breast
cancer. Oncoimmunology. 2:e251252013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rajkumar T, Sabitha K, Vijayalakshmi N,
Shirley S, Bose MV, Gopal G and Selvaluxmy G: Identification and
validation of genes involved in cervical tumourigenesis. BMC
Cancer. 11:802011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Altieri DC: Survivin, versatile modulation
of cell division and apoptosis in cancer. Oncogene. 22:8581–8589.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ciocca DR and Calderwood SK: Heat shock
proteins in cancer: diagnostic, prognostic, predictive, and
treatment implications. Cell Stress Chaperones. 10:86–103. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cory S, Huang DC and Adams JM: The Bcl-2
family: roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Romanucci M, Marinelli A, Sarli G and
Della Salda L: Heat shock protein expression in canine malignant
mammary tumours. BMC Cancer. 6:1712006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bruey JM, Ducasse C, Bonniaud P, et al:
Hsp27 negatively regulates cell death by interacting with
cytochrome. Nat Cell Biol. 2:645–652. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cornford PA, Dodson AR, Parsons KF,
Desmond AD, Woolfenden A, Fordham M, Neoptolemos JP, Ke Y and
Foster CS: Heat shock protein expression independently predicts
clinical outcome in prostate cancer. Cancer Res. 60:7099–7105.
2000.
|
|
37
|
Cohen M, Dromard M and Petignat P: Heat
shock proteins in ovarian cancer: a potential target for therapy.
Gynecol Oncol. 119:164–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
McCollum AK, Teneyck CJ, Sauer BM, Toft DO
and Erlichman C: Up-regulation of heat shock protein 27 induces
resistance to 17-allylamino-demethoxygeldanamycin through a
glutathione-mediated mechanism. Cancer Res. 66:10967–10975. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vasiliou V and Nebert DW: Analysis and
update of the human aldehyde dehydrogenase (ALDH) gene family. Hum
Genomics. 2:138–143. 2005.PubMed/NCBI
|
|
40
|
Vasiliou V, Pappa A and Estey T: Role of
human aldehyde dehydrogenases in endobiotic and xenobiotic
metabolism. Drug Metab Rev. 36:279–299. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Canuto RA, Muzio G, Ferro M, Maggiora M,
Federa R, Bassi AM, Lindahl R and Dianzani MU: Inhibition of
class-3 aldehyde dehydrogenase and cell growth by restored lipid
peroxidation in hepatoma cell lines. Free Radic Biol Med.
26:333–340. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Muzio G, Trombetta A, Maggiora M,
Martinasso G, Vasiliou V, Lassen N and Canuto RA: Arachidonic acid
suppresses growth of human lung tumor A549 cells through
down-regulation of ALDH3A1 expression. Free Radic Biol Med.
40:1929–1938. 2006. View Article : Google Scholar : PubMed/NCBI
|